Introduction
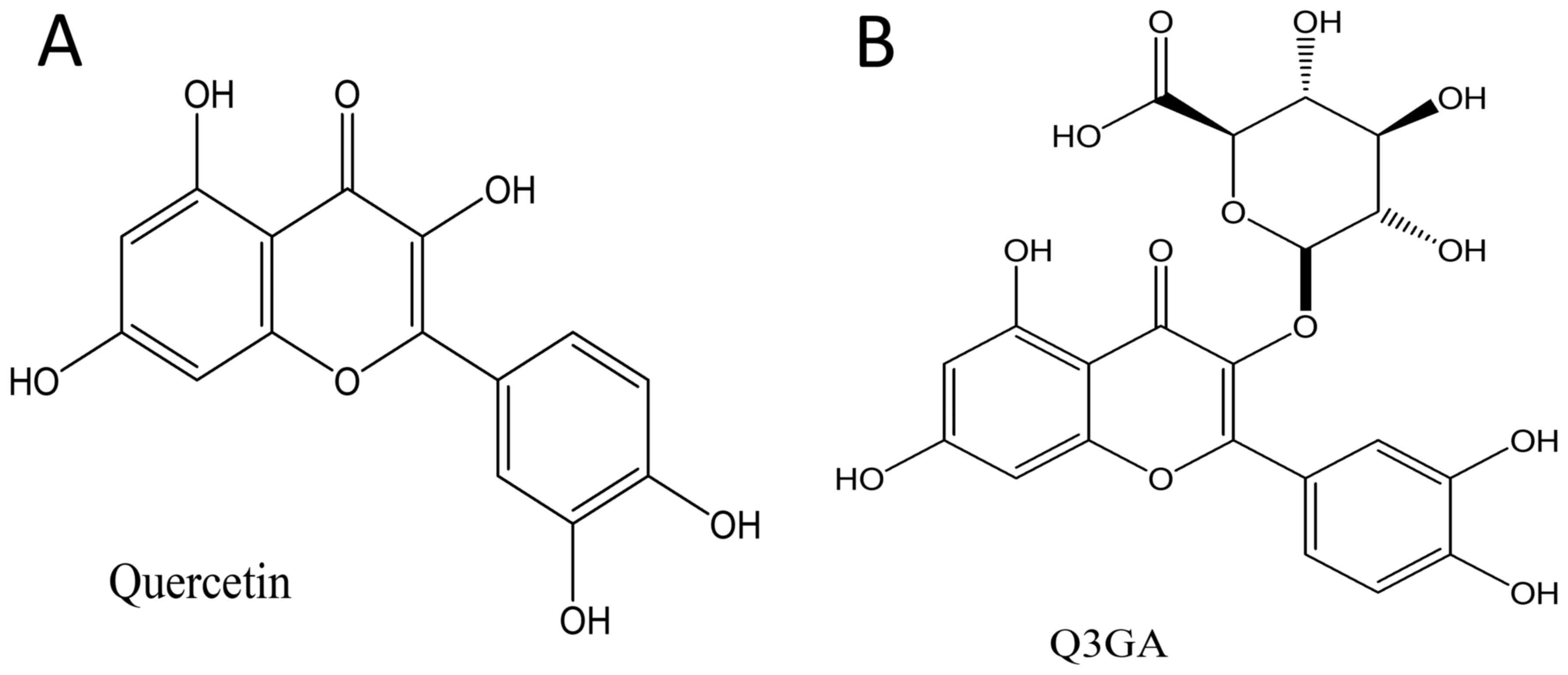
The flavonoid quercetin
(3,3',4',5,7-pentahydroxyflavone; Fig.
1A) is one of the most abundant dietary polyphenols. It is
mainly present in fruits and vegetables and ~3-40 mg quercetin is
consumed in daily diets (1,2). Quercetin is widespread in the flowers,
leaves and fruits of various plants and exhibits a multitude of
pharmacological activities, including anti-neoplastic (3-5),
anti-oxidative (6,7), anti-inflammatory (8,9),
anti-thrombotic (10,11), antiviral (12,13),
cardiovascular-protective (14,15)
and immune-regulatory (16,17) effects. Isolated quercetin is a
dietary supplement and its recommended maximum daily dosage is
1,000 mg (18). Following oral
ingestion, quercetin is extensively conjugated with glucuronic acid
and/or sulfate in the small intestine and liver.
Quercetin-3-O-β-D-glucoside (Q3GA; Fig.
1B) is one of the primary metabolites found in the blood
circulation (19). Q3GA exerts
various pharmacological properties. Quercetin inhibits the
viability of neural stem cells via the Akt signaling pathway.
However, Q3GA provides a novel therapeutic potential in
neurodegenerative diseases (20).
In addition, the anti-inflammatory activity of Q3GA was evaluated
by assessing the inhibition of LPS-induced NO release in
vitro (21).
The biological activity of quercetin is notably
affected by phase II, and not phase I, metabolism enzymes. As a
plant-derived polyphenol, quercetin contains more than one free
hydroxyl group, which makes it easy to be metabolized by different
types of UGT enzyme isoforms, including UGT1As (22,23).
UGTs are considered the indispensable enzymes of phase II
metabolism and catalyze the conjugation of several endobiotics or
xenobiotics with UDP-glucuronic acid in order to produce more
hydrophilic metabolites that are easily excreted via the kidneys or
the bile and the gut (24,25). CYP450-mediated herb-drug
interactions (HDIs) have been previously investigated by in
vitro assays using cocktails of probe substrates (26-29).
Subsequent studies involving UGT enzymes have demonstrated that
drug interactions based on the inhibition of UGTs may lead to
clinically important side effects (30,31).
Therefore, from a clinical point of view, the study of the
inhibition of herbal compounds on UGT-mediated metabolism may aid
the understanding of HDIs.
The in vitro UGT enzyme assay utilizes the
nonspecific substrate 4-MU as a substrate and has various
advantages over the use of human liver microsomes that include
several specific probe-substrates (32). In the present study, the inhibition
type and inhibitory effects of quercetin and its major metabolite
Q3GA were assessed on various UGT isoforms (UGT1A1, UGT1A3, UGT1A6
and UGT1A9) by liquid chromatography-tandem mass spectrometry
(LC-MS/MS). This method was used to detect the changes in the
concentration levels of 4-methylumbelliferyl-β-D-glucuronides
(4-MU-G). This may provide insight into the potential HDIs
regarding quercetin and Q3GA, providing the basis for further drug
research and safe drug use.
Materials and methods
Chemicals and reagents
4-MU and 4-MU-G were purchased from Shanghai Yuanye
Bio-Technology Co., Ltd. Q3GA was purchased from Chengdu Sino
Standards Bio-Tech Co., Ltd. 7-Hydroxycoumarin was obtained from
Dalian Meilun Biotechnology Co., Ltd. Quercetin and
uridine-5'-diphosphoglucuronic acid (UDPGA; trisodium salt) were
purchased from Sigma-Aldrich (Merck KGaA). Recombinant human UGT
isoforms (UGT1A1, UGT1A3, UGT1A6 and UGT1A9) were expressed in
baculovirus-infected insect cells that, where were obtained from
Corning, Inc. All other reagents were of the highest analytical
grade commercially available. The specific reagents were sourced
from companies mentioned previously (33).
Inhibition of recombinant
UGTs-catalyzed 4-MU glucuronidation by quercetin and Q3GA
The experimental protocol and incubation has been
accurately presented in previous studies (34-36).
Typical incubations were performed in 200 µl reaction mixture
containing 5 mM UDPGA, 5 mM MgCl2, 50 mM Tris-HCl buffer
(pH 7.4), 4-MU and recombinant UGTs. In addition, a series of
quercetin concentrations and Q3GA were dissolved in dimethyl
sulfoxide (DMSO) and the final concentration of DMSO in the total
mixture was <0.5% (v/v). Table I
indicates the final concentration of UGT1A isoforms and 4-Mu, and
the incubation time used for each UGT enzyme. Following
pre-incubation at 37˚C for 5 min, the UDPGA was added in the
incubation system to initiate the reaction. The samples were
incubated for the aforementioned incubation time periods and the
reaction was terminated by the addition of 200 µl ice-cold
methanol, containing 500 ng·ml-1 7-hydroxycoumarin as an
internal standard. Subsequently, the samples were centrifuged at
20,000 x g for 10 min at 4˚C, and 100 µl supernatant was obtained
and injected into the LC-MS/MS system for analysis.
 | Table ISubstrate concentration, enzyme
concentration and incubation time of each UGT enzyme. |
Table I
Substrate concentration, enzyme
concentration and incubation time of each UGT enzyme.
| UGT enzyme | Enzyme
concentration, mg/ml | Incubation time,
min | Substrate
concentration, µM |
|---|
| UGT1A1 | 0.125 | 120 | 30 |
| UGT1A3 | 0.05 | 120 | 1,200 |
| UGT1A6 | 0.025 | 30 | 110 |
| UGT1A9 | 0.05 | 30 | 30 |
Detection of 4-MU-G by LC-MS/MS
4-MU-G and 7-hydroxycoumarin (internal standard)
were analyzed on an API-4000 triple quadruple mass spectrometer
(Applied Biosystems; Thermo Fisher Scientific, Inc.) coupled with a
Waters ACQUITY Ultra Performance Liquid Chromatograph (Waters
Corporation). The separation was performed on an Inertsil ODS-SP
column (100x2.1 mm; 3 µm; GL Sciences) with a column temperature of
40˚C. The mobile phase consisted of ultrapure water, containing
0.1% formic acid (A) and acetonitrile containing 0.1% formic acid
(B). The following gradient conditions were used: 0-4.00 min, 5-80%
B; 4.00-4.10 min, 80-5% B; 4.10-7.00 min, 5% B. The flow rate used
was 0.2 ml/min, and the LC retention times of 4-MU-G and
7-hydroxycoumarin were 3.7 and 3.53 min, respectively. The turbo
ion spray interface was operated at -4,500 V and the ion source
temperature was set at 500℃ in the negative electrospray ionization
mode. The multiple reaction monitoring (MRM) mode was employed for
quantification using specific precursor/product ion transition. The
precursor/product ion transitions were monitored at m/z 351.1→175.1
and 161.0→89.0 for 4-MU-G and 7-hydroxycoumarin, respectively. The
optimized working parameters for mass detection of 4-MU-G and
7-hydroxycoumarin were as follows: i) Declustering potential, -50
and -80 V; ii) collision energy, -19 and -40 V; iii) curtain gas,
30 psi; iv) collision activated dissociation gas, 8 psi; v) Gas1,
55 psi and Gas2, 55 psi. The peak areas for all analytes were
automatically integrated using the Analyst software (version 1.5.1;
Applied Biosystems).
The specificity of this method was optimal. The
linear range was estimated to be 50-5,000 ng/ml with the lower
limit of quantification at 50 ng/ml. The RSD% of the intra-assay
and inter-assay precisions were both <10%. The extraction
recovery ranged between 100.99 and 106.34%. The internal standard
normalized matrix factors for the low-, moderate- and high-quality
control samples were 1.02, 1.07 and 0.99, respectively. The
residues were negligible, and the samples were placed in a sampler
at 4˚C for 9 h and left at room temperature for 2 h.
Determination of inhibition kinetic
parameters of quercetin and Q3GA on recombinant UGTs
The glucuronidation velocity was determined at
various 4-MU, quercetin or Q3GA concentrations. A preliminary
screening experiment was performed to assess the inhibitory effects
of quercetin and Q3GA. A total of 50 µM was selected for quercetin
and Q3GA as the experimental group concentration. The remaining
activity of UGTs=average concentration of 4-MUGUGT1A
enzyme-quercetin/average concentration of
4-MUGUGT1A enzyme-blank x100%. The remaining
activity of the blank group without quercetin and Q3GA was 100% in
the incubation system. Five or six concentration ranges of
quercetin and Q3GA were selected to assess the IC50
values for the UGT1A enzymes. The concentration ranges of quercetin
used were as follows: 0-10 µM for UGT1A1, 0-20 µM for UGT1A3 and
0-50 µM for UGT1A6 and UGT1A9. In addition, 0-50 µM Q3GA was
selected for UGT1A1, UGT1A3 and UGT1A6. The IC50 values
of quercetin and Q3GA towards UGT1A enzyme activities were
calculated by nonlinear regression analysis using the GraphPad
Prism 5 (GraphPad Software, Inc.). When the remaining activities of
these enzymes in the experimental groups were <50% of the
control group, the Ki was calculated. The Ki
resulted from fitting data into competitive inhibition,
non-competitive inhibition, or mixed inhibition models. The type of
inhibition was assessed graphically from the Lineweaver-Burk and
Dixon plots (37).
Results
Inhibitory activities of quercetin and
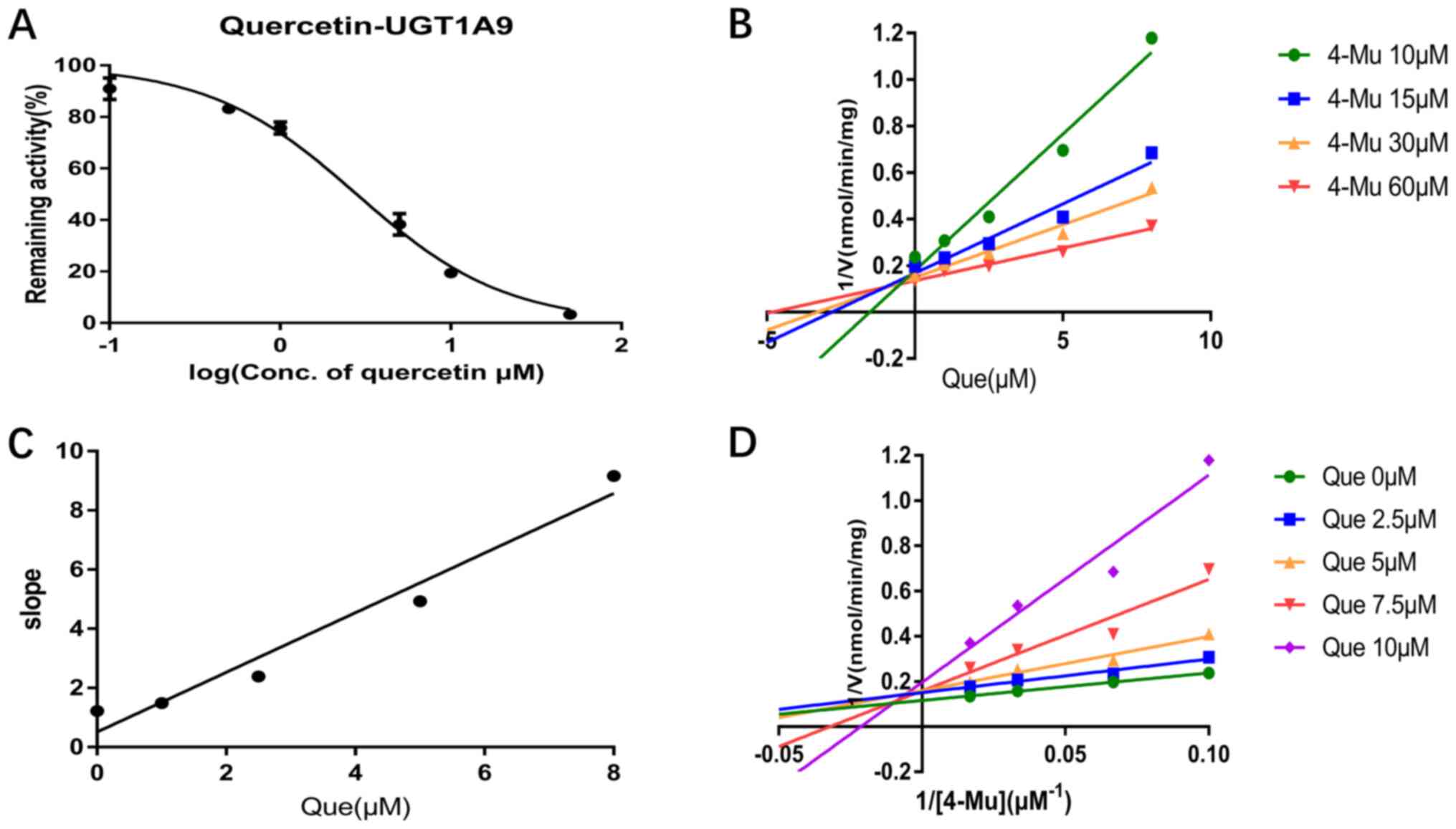
Q3GA on recombinant UGT1A isoforms
As shown in Table
II, in the presence of 50 µM quercetin, the remaining activity
of UGT1A1, UGT1A3, UGT1A6 and UGT1A9 enzymes were <8.4,
<30.8, 23.1 and 11.7%, respectively, while the remaining
activity of Q3GA on UGT1A1, UGT1A3, UGT1A6 and UGT1A9 was 36.1,
33.9, 28.4 and 74.2%, respectively. As the remaining activity of
UGT1A isoforms was <50%, except for UGTIA9, quercetin and Q3GA
exhibited inhibitory effects on UGT1A1, UGT1A3, UGT1A6 and UGT1A9
enzymes.
| Table IIPreliminary inhibition screening of
quercetin and Q3GA toward activities of recombinant UGT1A
isoforms. |
Table II
Preliminary inhibition screening of
quercetin and Q3GA toward activities of recombinant UGT1A
isoforms.
| | Remaining enzyme
activity |
|---|
| UGT1A enzyme | Quercetin, (%) | Q3GA, (%) |
|---|
| UGT1A1 | <8.4 | 36.1 |
| UGT1A3 | <30.8 | 33.9 |
| UGT1A6 | 23.1 | 28.4 |
| UGT1A9 | 11.7 | 74.2 |
In the presence of 50 µM quercetin, the final
concentrations of the metabolite 4-MUG were too low to detect in
UGT1A1 and UGT1A3 incubation systems. However, the lowest limit of
quantitation for 4-MUG was 50 ng/ml in the present study. The
remaining activity of UGT1A1 enzymes <50 ng/ml/593
ng/mlx100%=8.4%, and the remaining activity of UGT1A1 enzymes
<50 ng/ml/162.3 ng/mlx100%=30.8%. Therefore, the remaining
activities of UGT1A1 and UGT1A3 were less than 8.4 and 30.8%,
respectively.
Inhibition type and kinetics of
quercetin towards UGT1A1, UGT1A3, UGT1A6 and UGT1A9
The inhibitory parameters of quercetin on these UGT
isoforms were characterized/calculated by further kinetic
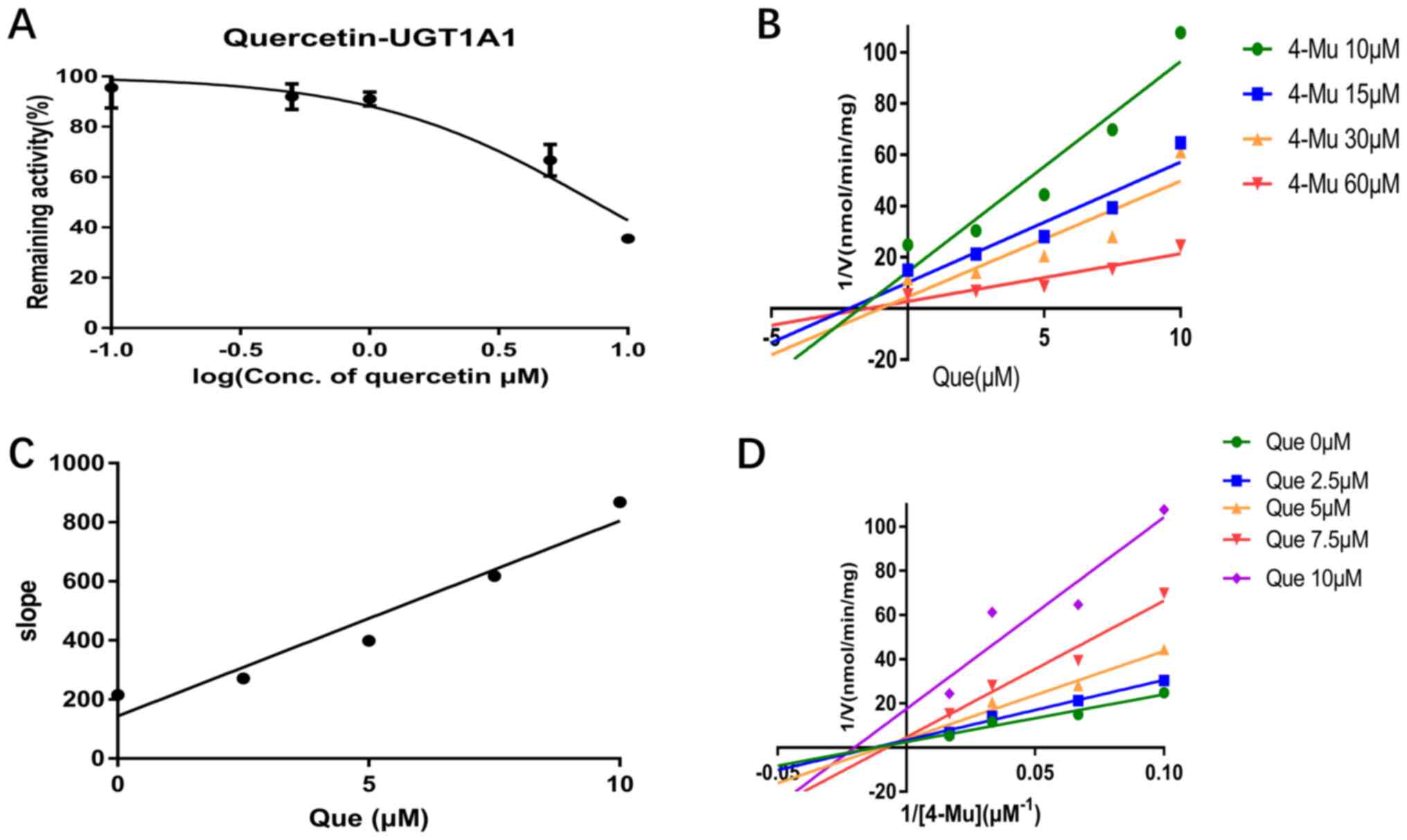
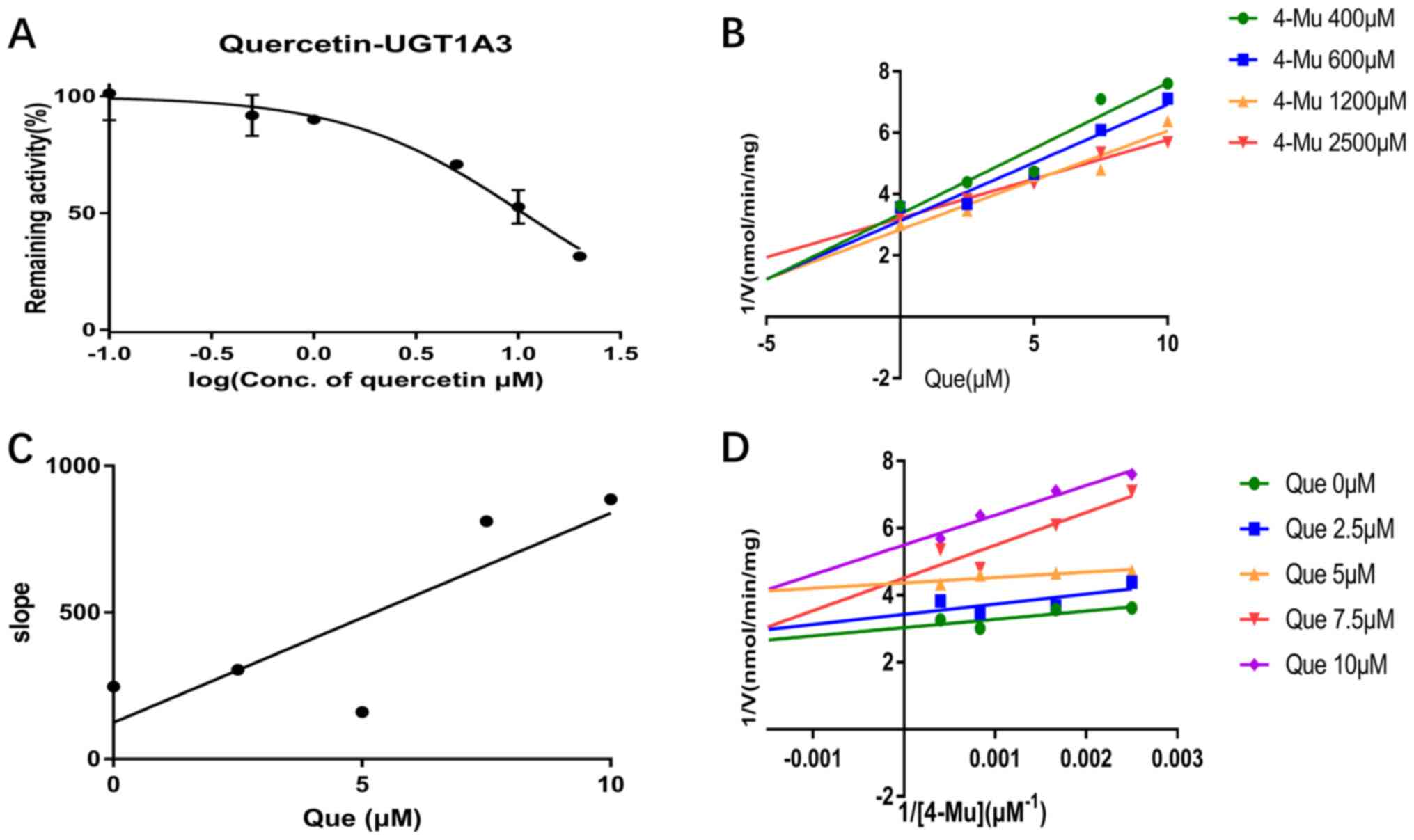
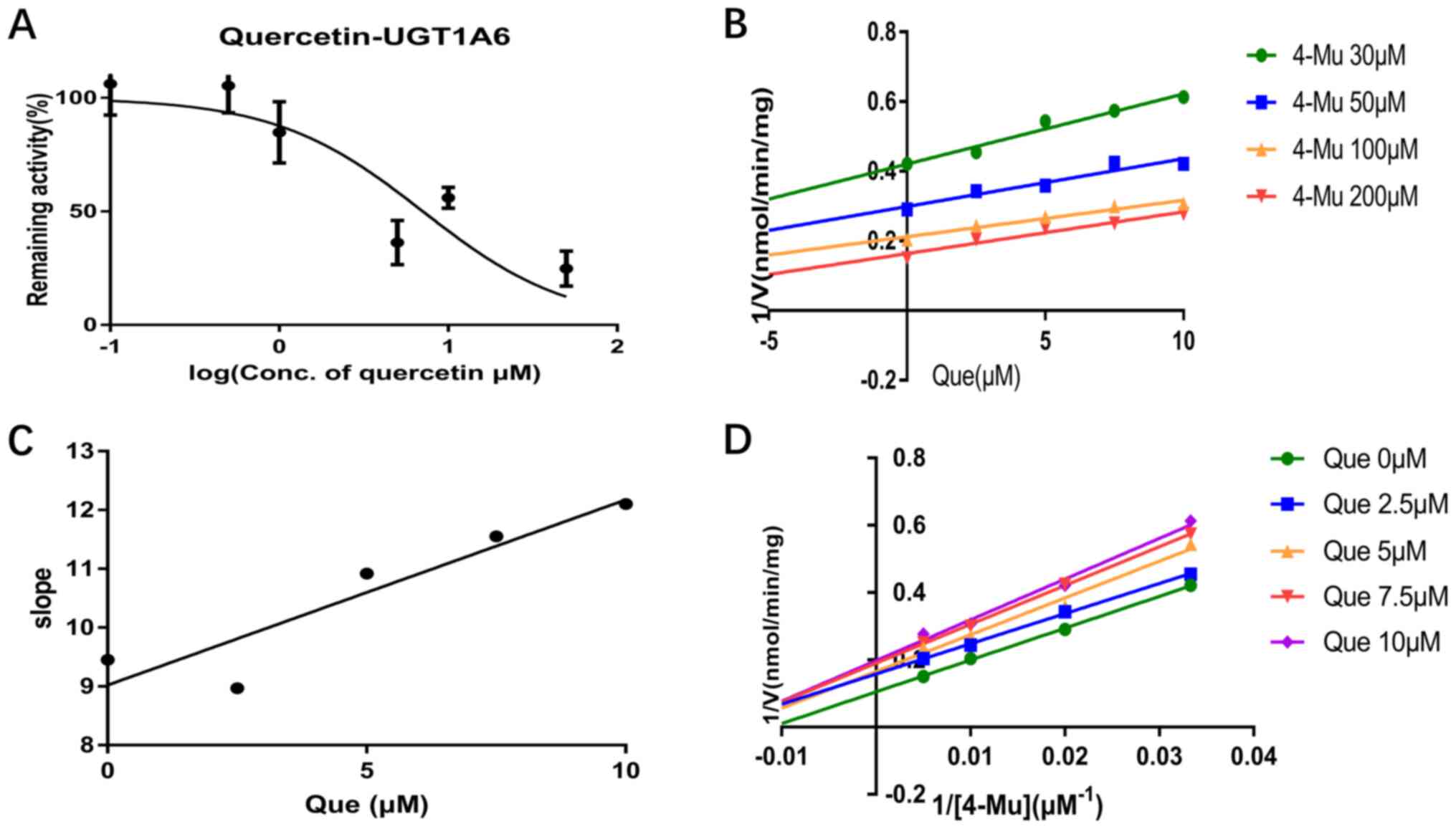
experiments. Quercetin inhibited UGT1A1, UGT1A3, UGT1A6 and UGT1A9
activity in a dose-dependent manner and the IC50 values
were 7.47, 10.58, 7.07 and 2.81 µM, respectively (Figs. 2A, 3A, 4A and
5A). Furthermore, the inhibition
types were determined from Dixon and Lineweaver-Burk plots. As
shown in Fig. 2B and D and in Fig.
4B and D, the inhibitory effect
of quercetin on UGT1A1 and UGT1A6-catalyzed 4-MU glucuronidation
was characterized as non-competitive inhibition, while the
inhibitory activity of quercetin on UGT1A3 and UGT1A6 was
characterized as competitive inhibition (Figs. 3B and D; 5B and
D). The calculated Ki
were 2.18, 1.60, 28.87 and 0.51 µM for the inhibitory effect of
quercetin on UGT1A1, UGT1A3, UGT1A6 and UGT1A9, respectively
(Figs. 2C, 3C, 4C and
5C).
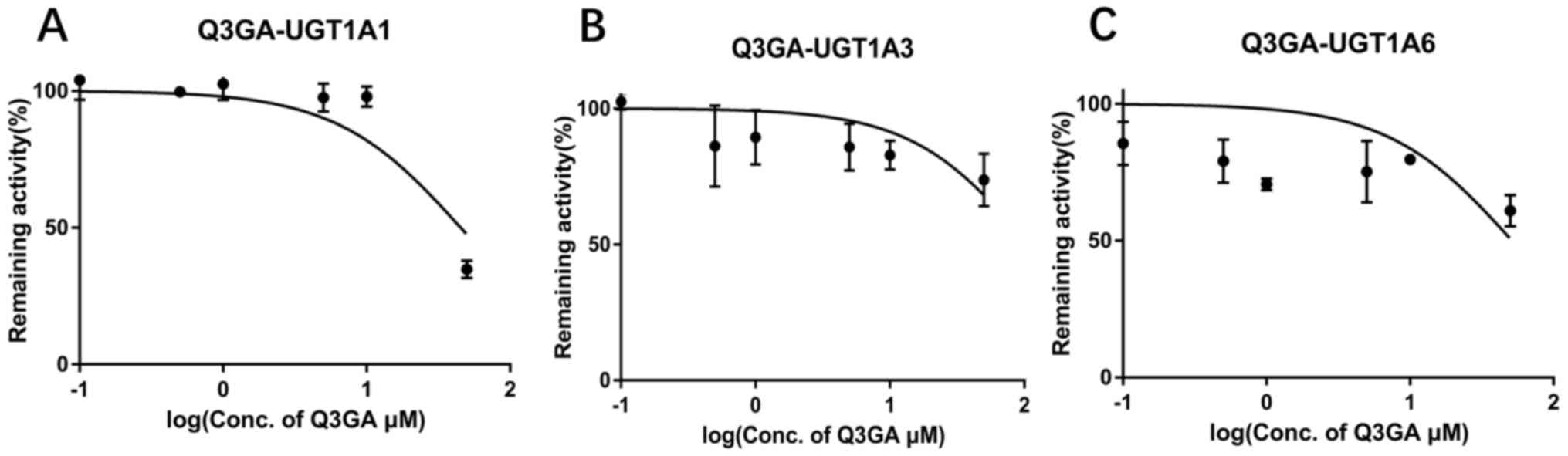
Inhibition type and kinetics of Q3GA
towards UGT1A1, UGT1A3 and UGT1A6
Preliminary screening experiments indicated that
Q3GA exhibited inhibitory effects on UGT1A1, UGT1A3 and UGT1A6. The
parameters of the kinetic experiments are presented in Fig. 6. The inhibitory activity of Q3GA on
UGT1A1, UGT1A3 and UGT1A6 enzymes was dose-dependent, with
IC50 values of 45.21, 106.5 and 51.37 µM, respectively.
Therefore, Q3GA may display weakly inhibit UGT1A1, UGT1A3 and
UGT1A6.
Discussion
In recent years, dietary supplements and alternative
medicine therapies have become increasingly accepted. A previous
study (38) has demonstrated that
the absorption and disposition of the active ingredients of dietary
supplements are significant in determining their biological
activity. In addition, accumulated evidence with regard to HDIs
between drug-metabolizing enzymes and drug transporters may be used
to predict the pharmacokinetic profile of drugs and the underlying
HDIs (39,40). These potential interactions may
cause significant risks to the patients, particularly for drugs
with narrow therapeutic indices.
It has been demonstrated that quercetin may prevent
cyclosporine A-induced nephrotoxicity and hepatotoxicity (41,42) as
it exerts various effects on the pharmacokinetics of cyclosporine
A. However, the underlying mechanisms for these interactions remain
unclear. Our previous studies have demonstrated that Q3GA increases
the Cmax, AUC0-t and AUC0-∞ of
cyclosporine A (19,43). In order to further elucidate the
underlying mechanisms of quercetin and Q3GA on the pharmacokinetics
of cyclosporine A, the inhibitory effects of quercetin and Q3GA on
the enzyme activity of recombinant UGT1A isoforms were investigated
in vitro.
The inhibitory potential of the compounds was
determined based on their Ki values and they were classified as
potent inhibitors (Ki<1 µM), moderate inhibitors (1 µM <
Ki<10 µM) and weak inhibitors (Ki>10 µM). Based on the
IC50 values of Q3GA on UGT1A1, UGT1A3 and UGT1A6 enzymes
that were much higher than 10 µM, and the fact that the remaining
activity of UGT1A9 was >50% following Q3GA incubation, these
data suggested that Q3GA displayed limited inhibitory effects on
these UGT isoforms. Therefore, the subsequent enzymatic kinetics
experiments were not performed, so the inhibition constant Ki of
Q3GA on UGT1A enzymes could not be calculated. However, quercetin
was more active in suppressing the activity of UGT isoforms
compared with Q3GA. Its inhibitory effect on UGT1A1 and UGT1A6 was
noncompetitive, with IC50 values of 7.47 and 7.07 µM,
respectively, and Ki values of 2.18 and 28.87 µM, respectively. By
contrast to these two isoforms, quercetin inhibited UGT1A3 and
UGT1A9 competitively with IC50 values of 10.58 and 2.81
µM, respectively, and Ki values of 1.60 and 0.51 µM, respectively.
These results indicated that quercetin was a moderate inhibitor of
UGT1A1 and UGT1A3, and a strong inhibitor of UGT1A9, while Q3GA
exhibited weak inhibitory effects on the activity of UGT1A1,
UGT1A3, UGT1A6 and UGT1A9 isoforms. There was a small slope in
Fig. 4C, which may be due to the
fact that quercetin only exerted a weak inhibitory effect on
UGT1A6. Furthermore, in the UGT1A3 incubation system, the
inhibitory effect of 5 µM quercetin did not have a linear
relationship with the inhibitory effect of other concentrations of
quercetin, which may be the reason for the small slope in Fig. 3C.
Quercetin and Q3GA exhibited similar inhibitory
effects on the protein and mRNA expression levels of UGT1A1 in the
small intestine and the liver. This is consistent with our
preliminary data and previously published studies (19,43).
It has also been reported that quercetin inhibits glucuronidation
of ethanol in human liver microsomes and in recombinant UGT1A1,
UGT1A3, UGT 1A4, UGT1A6 and UGT1A9 enzymes (44). However, quercetin has been reported
to induce UGT1A6 mRNA expression in human intestinal tissues in
vitro (45). Notably, quercetin
induces UGT1A expression in Caco-2 cells (46). It has also been reported that this
flavonoid increases UGT enzyme activities in hepatic, and small and
large intestinal tissues of male Wistar rats (47). A previous study has reported that
the age-associated differences in the UGT-catalyzed glucuronidation
of quercetin depend on the intestinal segment, particularly the
proximal and distal segments (48).
In brief, humans and animals express different protein and activity
levels of UGT1A isoforms.
There are limitations to the present study,
including the fact that quercetin or Q3GA were added separately to
the UGT1A incubation systems evaluate their inhibitory effects.
However, to the best of our knowledge, there are no effective
methods to distinguish the role of the quercetin/Q3GA vs. quercetin
plus Q3GA, as quercetin may be metabolized to Q3GA by UGT1A in the
incubation (49-51),
and Q3GA may be hydrolyzed to quercetin in the incubation in
activated mouse macrophages (49).
The results of the present study demonstrated that the inhibitory
effects of quercetin and Q3GA on UGT1A enzymes were significantly
different. Additionally, compared with quercetin, the inhibitory
effect of Q3GA on UGT1A enzymes was very weak. Therefore, we
hypothesized that Q3GA had a separate inhibitory effect on the
UGT1A enzymes, rather than inhibiting the UGT1A enzymes through
hydrolysis to quercetin.
A previous study (52) has reported that 3-hydroxyflavone has
a higher catalytic rate than 7-hydroxyflavone, due to differences
in the hydroxyl positions. Therefore, following conjugation of the
C-3 hydroxyl group with the glucuronic acid to yield Q3GA, a
decrease in the catalytic rate of Q3GA may be noted. In addition,
Boersma et al (53) reported
that quercetin inhibited UGT1A6 less efficiently than luteolin,
which was possibly due to the lack of the C-3 hydroxyl group. This
result suggested that flavonoids without the C-3 hydroxyl group
exhibit a significant inhibitory effect on the UGT1A6-catalyzed
reaction. This blocking effect is more pronounced for UGT1A6 when a
larger group is conjugated with the C-3 hydroxyl group. In general,
when Q3GA and quercetin are combined with UGT1As, the steric
hindrance effect of Q3GA greatly decreases the rate and extent of
this catalytic reaction.
In conclusion, the results of the present study
indicated that the inhibitory effect of quercetin on UGT1A1,
UGT1A3, UGT1A6 and UGT1A9 was more potent than that of Q3GA.
Furthermore, the results demonstrated that quercetin was a moderate
inhibitor of UGT1A1 and UGT1A3, and a strong inhibitor of UGT1A9,
and therefore HDIs may occur when quercetin is co-administered with
drugs that are mainly metabolized by UGT1A1, UGT1A3 and UGT1A9.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant nos. 81874326 and 81503161); The
Chinese Medicine Research Project of Health Commission of Hubei
Province (grant no. ZY2019Z004); and the National Key R&D
Program of China (grant no. 2017YFC0909900).
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
RZ, YL and SS designed all the experiments. YW and
TY performed the incubation experiments. Statistical analysis was
performed by XH, JZ, JNZ and CY. YL and SS confirm the authenticity
of all the raw data. YW, RZ, YL and SS wrote the manuscript. All
authors read and approved the final manuscript. RZ, YL and SS
designed all the experiments. YW and TY performed the incubation
experiments. Statistical analysis was performed by XH, JZ, JNZ and
CY. YL and SS confirm the authenticity of all the raw data. YW, RZ,
YL and SS wrote the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Babaei F, Mirzababaei M and Nassiri-Asl M:
Quercetin in food: Possible mechanisms of its effect on memory. J
Food Sci. 83:2280–2287. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Costa LG, Garrick JM, Roque PJ and
Pellacani C: Mechanisms of neuroprotection by quercetin:
Counteracting oxidative stress and more. Oxid Med Cell Longev.
2016(2986796)2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Jana N, Břetislav G, Pavel S and Pavla U:
Potential of the flavonoid quercetin to prevent and treat
cancer-current status of research. Klin Onkol. 31:184–190.
2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Primikyri A, Sayyad N, Quilici G, Vrettos
EI, Lim K, Chi SW, Musco G, Gerothanassis IP and Tzakos AG: Probing
the interaction of a quercetin bioconjugate with Bcl-2 in living
human cancer cells with in-cell NMR spectroscopy. FEBS Lett.
592:3367–3379. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Sharmila G, Athirai T, Kiruthiga B,
Senthilkumar K, Elumalai P, Arunkumar R and Arunakaran J:
Chemopreventive effect of quercetin in MNU and testosterone induced
prostate cancer of sprague-dawley rats. Nutr Cancer. 66:38–46.
2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Tang SH, Li R, Tan J, Wang Y and Jiang ZT:
One pot synthesis of water-soluble quercetin derived
multifunctional nanoparticles with photothermal and antioxidation
capabilities. Colloids Surf B Biointerfaces.
183(110429)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Zheng YZ, Deng G, Liang Q, Chen DF, Guo R
and Lai RC: Antioxidant activity of quercetin and its glucosides
from propolis: A theoretical study. Sci Rep. 7(7543)2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Newton K and Dixit VM: Signaling in innate
immunity and inflammation. Cold Spring Harb Perspect Biol.
4(a006049)2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Byun EB, Yang MS, Choi HG, Sung NY, Song
DS, Sin SJ and Byun EH: Quercetin negatively regulates TLR4
signaling induced by lipopolysaccharide through tollip expression.
Biochem Biophys Res Commun. 431:698–705. 2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lee SM, Moon J, Chung JH, Cha YJ and Shin
MJ: Effect of quercetin-rich onion peel extracts on arterial
thrombosis in rats. Food Chem Toxicol. 57:99–105. 2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Pan W, Chang MJ, Booyse FM, Grenett HE,
Bradley KM, Wolkowicz PE, Shang Q and Tabengwa EM: Quercetin
induced tissue-type plasminogen activator expression is mediated
through Sp1 and p38 mitogen-activated protein kinase in human
endothelial cells. J Thromb Haemost. 6:976–985. 2008.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Qiu X, Kroeker A, He S, Kozak R, Audet J,
Mbikay M and Chrétien M: Prophylactic efficacy of quercetin
3-β-O-d-glucoside against ebola virus infection. Antimicrob Agents
Chemother. 60:5182–5188. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gonzalez O, Fontanes V, Raychaudhuri S,
Loo R, Loo J, Arumugaswami V, Sun R, Dasgupta A and French SW: The
heat shock protein inhibitor quercetin attenuates hepatitis C virus
production. Hepatology. 50:1756–1764. 2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu H, Guo X, Chu Y and Lu S: Heart
protective effects and mechanism of quercetin preconditioning on
anti-myocardial ischemia reperfusion (IR) injuries in rats. Gene.
545:149–155. 2014.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Lin R, Liu J, Gan W and Ding C: Protective
effect of quercetin on the homocysteine-injured human umbilical
vein vascular endothelial cell line (ECV304). Basic Clin Pharmacol
Toxicol. 101:197–202. 2007.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Kobori M, Takahashi Y, Sakurai M, Akimoto
Y, Tsushida T, Oike H and Ippoushi K: Quercetin suppresses immune
cell accumulation and improves mitochondrial gene expression in
adipose tissue of diet-induced obese mice. Mol Nutr Food Res.
60:300–312. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Singh D, Tanwar H, Jayashankar B, Sharma
J, Murthy S, Chanda S, Singh SB and Ganju L: Quercetin exhibits
adjuvant activity by enhancing Th2 immune response in ovalbumin
immunized mice. Biomed Pharmacother. 90:354–360. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Andres S, Pevny S, Ziegenhagen R, Bakhiya
N, Schäfer B, Hirsch-Ernst KI and Lampen A: Safety aspects of the
use of quercetin as a dietary supplement. Mol Nutr Food Res.
62:2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Yang T, Liu Y, Huang X, Zhang R, Yang C,
Zhou J, Zhang Y, Wan J and Shi S: Quercetin-3-O-β-D-glucoside
decreases the bioavailability of cyclosporin A through regulation
of drug metabolizing enzymes, transporters and nuclear receptors in
rats. Mol Med Rep. 18:2599–2612. 2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Baral S, Pariyar R, Kim J, Lee HS and Seo
J: Quercetin-3-O-glucuronide promotes the proliferation and
migration of neural stem cells. Neurobiol Aging. 52:39–52.
2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li F, Sun XY, Li XW, Yang T and Qi LW:
Enrichment and separation of quercetin-3-O-β-d-glucuronide from
lotus leaves (nelumbo nucifera gaertn.) and evaluation of its
anti-inflammatory effect. J Chromatogr B Analyt Technol Biomed Life
Sci. 1040:186–191. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Oliveira EJ and Watson DG: In vitro
glucuronidation of kaempferol and quercetin by human UGT-1A9
microsomes. FEBS Lett. 471:1–6. 2000.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhou H, Shi R, Ma B, Ma Y, Wang C, Wu D,
Wang X and Cheng N: CYP450 1A2 and multiple UGT1A isoforms are
responsible for jatrorrhizine metabolism in human liver microsomes.
Biopharm Drug Dispos. 34:176–185. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
24
|
Stingl JC, Bartels H, Viviani R, Lehmann
ML and Brockmöller J: Relevance of UDP-glucuronosyltransferase
polymorphisms for drug dosing: A quantitative systematic review.
Pharmacol Ther. 141:92–116. 2014.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Miners JO, Mackenzie PI and Knights KM:
The prediction of drug-glucuronidation parameters in humans:
UDP-glucuronosyltransferase enzyme-selective substrate and
inhibitor probes for reaction phenotyping and in vitro-in vivo
extrapolation of drug clearance and drug-drug interaction
potential. Drug Metab Rev. 42:196–208. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Jeong HU, Lee JY, Kwon SS, Kim JH, Kim YM,
Hong SW, Yeon SH, Lee SM, Cho YY and Lee HS: Metabolism-mediated
drug interaction potential of HS-23, a new herbal drug for the
treatment of sepsis in human hepatocytes and liver microsomes. Arch
Pharm Res. 38:171–177. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Tan ML and Lim LE: The effects of
Andrographis paniculata (Burm.f.) Nees extract and diterpenoids on
the CYP450 isoforms' activities, a review of possible herb-drug
interaction risks. Drug Chem Toxicol. 38:241–253. 2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li G, Huang K, Nikolic D and van Breemen
RB: High-throughput cytochrome P450 cocktail inhibition assay for
assessing drug-drug and drug-botanical interactions. Drug Metab
Dispos. 43:1670–1678. 2015.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Weissenstein U, Kunz M, Oufir M, Wang JT,
Hamburger M, Urech K, Regueiro U and Baumgartner S: Absence of
herb-drug interactions of mistletoe with the tamoxifen metabolite
(E/Z)-endoxifen and cytochrome P450 3A4/5 and 2D6 in vitro. BMC
Complement Altern Med. 19(23)2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Tuteja S, Pyrsopoulos NT, Wolowich WR,
Khanmoradi K, Levi DM, Selvaggi G, Weisbaum G, Tzakis AG and Schiff
ER: Simvastatin-ezetimibe-induced hepatic failure necessitating
liver transplantation. Pharmacotherapy. 28:1188–1193.
2008.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Magee CN, Medani SA, Leavey SF, Conlon PJ
and Clarkson MR: Severe rhabdomyolysis as a consequence of the
interaction of fusidic acid and atorvastatin. Am J Kidney Dis.
56:e11–e15. 2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Huang T, Fang ZZ and Yang L: Strong
inhibitory effect of medroxyprogesterone acetate (MPA) on
UDP-glucuronosyltransferase (UGT) 2B7 might induce drug-drug
interactions. Pharmazie. 65:919–921. 2010.PubMed/NCBI
|
|
33
|
Lu H, Fang ZZ, Cao YF, Hu CM, Hong M, Sun
XY, Li H, Liu Y, Fu X and Sun H: Isoliquiritigenin showed strong
inhibitory effects towards multiple UDP-glucuronosyltransferase
(UGT) isoform-catalyzed 4-methylumbelliferone (4-MU)
glucuronidation. Fitoterapia. 84:208–212. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Liu C, Cao YF, Fang ZZ, Zhang YY, Hu CM,
Sun XY, Huang T, Zeng J, Fan XR and Mo H: Strong inhibition of
deoxyschizandrin and schisantherin A toward
UDP-glucuronosyltransferase (UGT) 1A3 indicating UGT
inhibition-based herb-drug interaction. Fitoterapia. 83:1415–1419.
2012.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Jiang HM, Fang ZZ, Cao YF, Hu CM, Sun XY,
Hong M, Yang L, Ge GB, Liu Y, Zhang YY, et al: New insights for the
risk of bisphenol A: Inhibition of UDP-glucuronosyltransferases
(UGTs). Chemosphere. 93:1189–1193. 2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Sun D, Zhang CZ, Ran RX, Cao YF, Du Z, Fu
ZW, Huang CT, Zhao ZY, Zhang WH and Fang ZZ: In vitro comparative
study of the inhibitory effects of mangiferin and its aglycone
norathyriol towards UDP-glucuronosyl transferase (UGT) isoforms.
Molecules. 22(1008)2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhu L, Ge G, Liu Y, He G, Liang S, Fang Z,
Dong P, Cao Y and Yang L: Potent and selective inhibition of
magnolol on catalytic activities of UGT1A7 and 1A9. Xenobiotica.
42:1001–1008. 2012.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Miclaus MO, Filip X, Filip C, Martin FA
and Grosu IG: Highly sensitive solid forms discrimination on the
whole tablet of the active ingredients in quercetin dietary
supplements by NMR crystallography approaches. J Pharm Biomed Anal.
124:274–280. 2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Shi S and Li Y: Interplay of
drug-metabolizing enzymes and transporters in drug absorption and
disposition. Curr Drug Metab. 15:915–941. 2014.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wu B: Pharmacokinetic interplay of phase
II metabolism and transport: A theoretical study. J Pharm Sci.
101:381–393. 2012.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zal F, Mostafavi-Pour Z and Vessal M:
Comparison of the effects of vitamin E and/or quercetin in
attenuating chronic cyclosporine A-induced nephrotoxicity in male
rats. Clin Exp Pharmacol Physiol. 34:720–724. 2007.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Mostafavi-Pour Z, Zal F, Monabati A and
Vessal M: Protective effects of a combination of quercetin and
vitamin E against cyclosporine A-induced oxidative stress and
hepatotoxicity in rats. Hepatol Res. 38:385–392. 2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Liu Y, Luo X, Yang C, Yang T, Zhou J and
Shi S: Impact of quercetin-induced changes in drug-metabolizing
enzyme and transporter expression on the pharmacokinetics of
cyclosporine in rats. Mol Med Rep. 14:3073–3085. 2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Stachel N and Skopp G: Formation and
inhibition of ethyl glucuronide and ethyl sulfate. Forensic Sci
Int. 265:61–64. 2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
van de Kerkhof EG, de Graaf IA, Ungell AL
and Groothuis GM: Induction of metabolism and transport in human
intestine: Validation of precision-cut slices as a tool to study
induction of drug metabolism in human intestine in vitro. Drug
Metab Dispos. 36:604–613. 2008.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Galijatovic A, Walle UK and Walle T:
Induction of UDP-glucuronosyltransferase by the flavonoids chrysin
and quercetin in Caco-2 cells. Pharm Res. 17:21–26. 2000.PubMed/NCBI View Article : Google Scholar
|
|
47
|
van der Logt EM, Roelofs HM, Nagengast FM
and Peters WH: Induction of rat hepatic and intestinal
UDP-glucuronosyltransferases by naturally occurring dietary
anticarcinogens. Carcinogenesis. 24:1651–1656. 2003.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Bolling BW, Court MH, Blumberg JB and Chen
CY: Microsomal quercetin glucuronidation in rat small intestine
depends on age and segment. Drug Metab Dispos. 39:1406–1414.
2011.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Kawai Y, Nishikawa T, Shiba Y, Saito S,
Murota K, Shibata N, Kobayashi M, Kanayama M, Uchida K and Terao J:
Macrophage as a target of quercetin glucuronides in human
atherosclerotic arteries: Implication in the anti-atherosclerotic
mechanism of dietary flavonoids. J Biol Chem. 283:9424–9434.
2008.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Ishizawa K, Yoshizumi M, Kawai Y, Terao J,
Kihira Y, Ikeda Y, Tomita S, Minakuchi K, Tsuchiya K and Tamaki T:
Pharmacology in health food: Metabolism of quercetin in vivo and
its protective effect against arteriosclerosis. J Pharmacol Sci.
115:466–470. 2011.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Chao CL, Hou YC, Chao PD, Weng CS and Ho
FM: The antioxidant effects of quercetin metabolites on the
prevention of high glucose-induced apoptosis of human umbilical
vein endothelial cells. Brit J Nutr. 101:1165–1170. 2009.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Senafi SB, Clarke DJ and Burchell B:
Investigation of the substrate specificity of a cloned expressed
human bilirubin UDP-glucuro-nosyltransferase: UDP-sugar specificity
and involvement in steroid and xenobiotic glucuronidation. Biochem
J. 303:233–240. 1994.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Boersma MG, van der Woude H, Bogaards J,
Boeren S, Vervoort J, Cnubben NH, van Iersel ML, van Bladeren PJ
and Rietjens IM: Regioselectivity of phase II metabolism of
luteolin and quercetin by UDP-glucuronosyl transferases. Chem Res
Toxicol. 15:662–670. 2002.PubMed/NCBI View Article : Google Scholar
|