Introduction
Low back pain caused by intervertebral disc
degeneration (IDD) seriously affects the quality of life of
patients with this condition (1).
Therapy for IDD includes conservative treatment (including physical
therapy and pain management) and surgical treatment, but neither
can reverse the pathological status of IDD (2). IDD is caused by a decrease in nucleus
pulposus cells (NPCs) and a subsequent decrease in proteoglycans in
the extracellular matrix (ECM) (3).
The cell density and cell regeneration rate in the nucleus pulposus
are low; therefore, repair after degeneration is difficult
(4). To alleviate and treat IDD,
researchers have focused on maintaining and increasing the number
of NPCs and enhancing ECM deposition (5). In a previous study, regenerative
therapy with stem cells has provided a novel approach for the
treatment of IDD. This approach can help repopulate intervertebral
discs by promoting the production of ECM, restoration of damaged
tissue and secretion of growth factors to regulate inflammation and
strengthen tissue regeneration, without causing additional
intervertebral disc injury (6).
The native intervertebral disc is typically composed
of two distinct anatomic regions, the annulus fibrosus and the
nucleus pulposus (7). NPCs can be
obtained from intervertebral disc tissues and have been used to
regenerate the disc tissue. Although NPCs have been used to
regenerate disc tissue (8,9), access to healthy NPCs, especially
autologous cells, is limited in clinical settings. NPC-like cells
are not obtained from the intervertebral disc tissue, but they
exhibit a nucleus pulposus-like phenotype and can potentially
provide a suitable autologous cell source for nucleus pulposus
tissue regeneration (10).
Continuous progress has been made with in vitro and in
vivo experiments of stem cell therapies for IDD, and biological
scaffolds have been widely used (11). In stem cell therapy for disc
degeneration, the seed cell selection is the first consideration
(12). Stem cells used in the
biological treatment of IDD should exhibit the following
characteristics (13): i) Be
abundant and easy to obtain; ii) be able to differentiate well into
NPCs; iii) be able to adapt to the local microenvironment of IDD
with low oxygen, low glucose and slight acidity; iv) not elicit a
strong immune response after transplantation; and v) have a low
potential for tumor growth. At present, commonly used stem cells
include mesenchymal stem cells (MSCs), induced pluripotent stem
cells (iPSCs), hematopoietic stem cells and embryonic stem cells.
Commonly used MSCs include bone marrow-derived MSCs (BMSCs),
adipose-derived MSCs (ADSCs) and umbilical cord MSCs (13).
BMSCs and ADSCs are two of the most widely studied
types of MSCs, and they are widely available and easy to culture
in vitro (14). It was
indicated that TGF-β1(15) and bone
morphogenetic protein 7(16) could
effectively induce the differentiation of BMSCs into NPC-like
cells. Elabd et al (17)
injected autologous BMSCs cultured in a hypoxic environment into
the intervertebral disc of 5 patients with intervertebral
discogenic low back pain and observed that the waist activity in 4
patients improved to different degrees. Noriega et al
(18) conducted allogeneic BMSC
transplantation in 24 patients with discogenic low back pain and
demonstrated that allogeneic BMSC transplantation was also
effective. Xu et al (19)
revealed that ADSCs could be induced to differentiate into NPC-like
cells after co-culture with NPCs. Clarke et al (20) confirmed that TGF-β1, growth
differentiation factor 5 or growth differentiation factor 6 could
induce better differentiation of ADSCs into NPC-like cells compared
with BMSCs, and could promote the expression of ECM molecules.
In tissue engineering, biological scaffolds can
maintain cell function and provide mechanical protection for cells
(10). The selection of stem cell
scaffolds for the treatment of IDD is an important issue (21,22).
Gelatinous scaffolds exhibit low viscosity during cell delivery and
can be gelled in situ after injection, thereby filling
micro- and macro-cracks and restoring the height of intervertebral
discs (23,24). Therefore, gelatinous scaffolds are
an ideal scaffold system. Feng et al (10) successfully established a biological
scaffold containing glucan-gelatin hydrogel-TGF-β1 and used the
scaffold to effectively induce mouse BMSCs to differentiate into
NPCs and promote the expression of related ECM genes.
Under appropriate conditions, BMSCs (25) and ADSCs (26) can differentiate into NPCs and reduce
the apoptosis of NPCs. However, a comparative study on the
differentiation of these two types of cells into NPC-like cells in
3D culture has not yet been reported, to the best of our knowledge.
The present study aimed to investigate the differences between
BMSCs and ADSCs in their ability to differentiate into cells with
an NPC-like phenotype in 3D culture to provide a reference for the
selection of candidate cells for the biotherapy of IDD-associated
diseases.
Materials and methods
Isolation of BMSCs and ADSCs
BMSCs were isolated and purified using the whole
bone marrow cell repeated adherent culture method (27-30).
All animal experiments were approved by the Animal Experiment and
Ethics Committee of Kunming Medical University (Kunming, China;
approval no. KM20190301). Three male Sprague-Dawley rats (6-8 weeks
old; ~200 g in weight) were purchased from the Laboratory Animal
Center of Kunming Medical University and housed in a humidity
(50-65%) pathogen-free environment at 20-25˚C with free access to
food and water under a 12/12 h light/dark cycle. Rats were
euthanized with sodium pentobarbital overdose (160 mg/kg;
intraperitoneal injection). Euthanasia was confirmed when the rats
exhibited no heartbeat and breathing for 2-3 min and no blinking
reflex. The femur and tibia of the rats were removed under aseptic
conditions, and the bone marrow cavity was exposed after the two
ends were cut off. The bone marrow cavity was washed with PBS three
times. The wash solution was collected and centrifuged at 800 x g
for 5 min at room temperature. Subsequently, the bone marrow cell
pellet was suspended in 1 ml DMEM/F12 (Thermo Fisher Scientific,
Inc.) medium containing 10% FBS (Thermo Fisher Scientific, Inc.)
and plated into a 25 cm2 cell culture flask. Cells were
then maintained at 37˚C with 5% CO2. The medium was
changed every 3 days. When the cell culture reached 90% confluence,
the cells were passaged at a ratio of 1:3.
Adipose tissues in the subcutaneous inguinal region
were separated, and the fascial tissue and lymph nodes were
eliminated with tweezers as much as possible. The adipose tissue
was washed with PBS three times. After being cut into pieces, the
tissue was centrifuged at 800 x g for 10 min at 4˚C, and the
pelleted layer of adipose tissue was retained. After being
resuspended in PBS, the tissue was centrifuged at 800 x g for 10
min at 4˚C. The pelleted layer of adipose tissue was mixed with
0.1% collagenase (Beijing Solarbio Science & Technology Co.,
Ltd.) solution (prepared in DMEM/F12 containing 10% FBS; each,
Thermo Fisher Scientific, Inc.) and digested in a 5% CO2
incubator at 37˚C for 2 h. The solution was shaken every half hour.
After the digested tissue solution was centrifuged at 600 x g for 5
min at 4˚C, the supernatant was discarded and the pellet was
resuspended in PBS.
BMSCs and ADSCs were cultured in a 5% CO2
incubator at 37˚C with DMEM/F12 (Thermo Fisher Scientific, Inc.)
containing 10% FBS, 1 IU/ml penicillin (Beijing Solarbio Science
& Technology Co., Ltd.) and 1 µg/ml streptomycin (Beijing
Solarbio Science & Technology Co., Ltd.). Cells from the third
passage were preserved for further experiments.
Identification of BMSCs and ADSCs
BMSC and ADSC cell surface markers were examined via
flow cytometry using the forward scatter/side scatter method. Cells
were trypsinized and resuspended in PBS at 1x105 cells
per 1.5 ml. BMSCs were incubated for 2 h at room temperature in the
dark with the following FITC-conjugated antibodies: Anti-CD29
(1:50; cat. no. bs-20630R-FITC), anti-CD90 (1:200; cat. no.
bs-0778R-FITC), anti-CD45 (1:100; cat. no. bs-10599R-FITC) and
anti-CD34 (1:50; cat. no. bs-0646R-FITC; all from BIOSS). ADSCs
were incubated for 2 h at room temperature with the following
FITC-conjugated antibodies: Anti-rat stem cells antigen-1 (Sca-1;
1:100; cat. no. bs-3752R-FITC; BIOSS), anti-CD44 (1:50; cat. no.
bs-0521R-FITC), anti-CD45 (1:100; cat. no. bs-10599R-FITC) and
anti-CD11b (1:200; cat. no. bs-1014R-FITC; all from BIOSS).
Subsequently, the cells were washed twice with PBS and analyzed by
a CyFlow™ Space flow cytometer (Sysmex Partec GmbH) with FloMax 2.8
software (Sysmex Partec GmbH).
3D culture and grouping
The experiments were performed with four groups: i)
BMSC control group; ii) ADSC control group; iii) BMSC 3D culture
group; and iv) ADSC 3D culture group. In the control groups, cells
were cultured with differentiating medium in 2% O2 and
5% CO2 at 37˚C. The 3D culture system was based on the
bioactive hydrogel method (3D cell culture hydrogel kit; cat. no.
FS0469; Shanghai Fushen Biotechnology Co., Ltd.) and was performed
according to the manufacturer's protocol. In the 3D culture groups,
gel solution and 1x106/ml BMSC or ADSC suspension were
gently mixed in a centrifuge tube, which was gently and quickly
inverted 1-2 times for 1 sec. After the cell culture plate was
washed with 1X PBS, the gel-cell mixture was added and gently
shaken to evenly spread the mixture in the plate (under different
circumstances the mixture could quickly adhere to the walls of the
plates). Subsequently, the cells in the 3D culture were incubated
at 37˚C for 5-10 min to form a gel. After incubation, the culture
medium was added by pipetting down towards the cell plate wall, and
the cells were cultured at 37˚C in 5% CO2 for 24 h.
After 24 h, half of the upper cell differentiation medium was
gently removed with a pipette, and the same amount of fresh medium
for NPC-like cells was added, after which the medium was regularly
changed. To induce differentiation, cells in 3D culture were
cultured with differentiating medium consisting of DMEM/F12
supplemented with 10 ng/ml TGF-β1 (cat. no. 96-100-21-2; PeproTech,
Inc.), 100 nmol/l dexamethasone (cat. no. D8040; Beijing Solarbio
Science & Technology Co., Ltd.), 50 mg/ml aseorbie acid (cat.
no. A8100; Beijing Solarbio Science & Technology Co., Ltd.),
100 mg/ml sodium pyruvate (cat. no. P8380; Beijing Solarbio Science
& Technology Co., Ltd.), 40 mg/ml proline (cat. no. P0011;
Beijing Solarbio Science & Technology Co., Ltd.) and ITS-Plus
Media Supplement (Collaborative Biomedical Products, Inc.) in 2%
O2. After 7 days of culture, the cells were subjected to
subsequent experiments.
Cell viability
The viability of BMSC and ADSC in 3D and 2D culture
conditions was evaluated with the Cell Counting Kit-8 (CCK-8) assay
(BIOSS). A total of 3x104 BMSCs or ADSCs were plated
into 96-well-plates for 3D and 2D culture. After culture at 37˚C
for 48 h, a CCK-8 assay was used to detect cell viability. For 3D
culture, cells were released from the gel via treatment with
pyrolysis liquid (cat. no. FS0469-B; Shanghai Fushen Biotechnology
Co., Ltd.) for 10 min at room temperature. The cells were washed
with PBS, and the CCK-8 solution (10 µl/well) was added to the
plates. Subsequently, the cells were incubated at 37˚C for 2 h. The
optical density (OD) at 450 nm was measured using a microplate
reader (BioTek Instruments, Inc.).
Visualization of sulfated
glycosaminoglycans (GAGs) and proteoglycans
BMSCs and ADSCs from the control and 3D culture
groups were stained with Alcian blue (cat. no. G1563; Beijing
Solarbio Science & Technology Co., Ltd.) and safranin O (cat.
no. G2540; Beijing Solarbio Science & Technology Co., Ltd.),
respectively, to visualize the formation of GAGs and proteoglycans.
For Alcian blue staining, cells and gels were washed with PBS three
times, fixed in 4% paraformaldehyde (cat. no. P1110; Beijing
Solarbio Science & Technology Co., Ltd.) for 20 min at room
temperature, then incubated with a 1% Alcian blue staining solution
for 4 h at room temperature. For safranin O staining, cells and
gels were washed with PBS three times, fixed in 4% paraformaldehyde
for 30 min at room temperature, then incubated with safranin O for
30 min at room temperature. After removing the dye solution, the
cells and gels were briefly washed with ddH2O three
times, and the staining intensity was then observed via light
microscopy (magnification, x200).
Reverse transcription-quantitative PCR
(RT-qPCR) assay
BMSCs and ADSCs were released from the gel via
treatment with pyrolysis liquid (cat. no. FS0469-B; Shanghai Fushen
Biotechnology Co., Ltd.) for 10 min at room temperature in the 3D
cell culture hydrogel kit. Total RNA was extracted from cells using
Trizol reagent (Takara Biotechnology Co., Ltd.) according to the
manufacturer's instructions. cDNA was synthesized using a
PrimeScript RT Reagent kit (Takara Biotechnology Co., Ltd.)
according to the manufacturer's protocol. mRNA levels were measured
via qPCR (ABI 7900; Applied Biosystems; Thermo Fisher Scientific,
Inc.) using a TB Green Premix ExTaq kit (Takara Biotechnology Co.,
Ltd.). Relative gene expression was analyzed with the
2-ΔΔCq method (31).
GAPDH was used as the reference gene. The primers for the genes
were as follows: GAPDH forward, 5'-AGAACATCATCCCTGCATCC-3' and
reverse, 5'-TTACTCCTTGGAGGCCATGT-3'; collagen II forward,
5'-CACTCATCTGTTGTGATGAGTTCTCC-3' and reverse,
5'-CAACACACACCAGCGCAGTTT-3'; aggrecan forward,
5'-GGGTGAGGTCTTTTATGCCA-3' and reverse, 5'-GCTTTGCAGTGAGGATCACA-3';
Sox-9 forward, 5'-TTGCTCGGAACTGTCTGGAA-3' and reverse,
5'-CCTGCTCGTCGGTCATCTT-3'; hypoxia-inducible factor 1-α (HIF-1α)
forward, 5'-ACTATGTCGCTTTCTTGG-3' and reverse,
5'-GTTTCTGCTGCCTTGTAT-3'; and glucose transporter 1 (GLUT1)
forward, 5'-GCCCTGGATGTCCTATCTGA-3' and reverse,
5'-CCCACGATGAAGTTTGAGGT-3'.
Western blot assay
BMSCs and ADSCs were released from the gel via
treatment with pyrolysis liquid as aforementioned. Cell proteins
were extracted using RIPA lysis buffer (Beyotime Institute of
Biotechnology) and quantified using a BCA Protein Assay Kit
(Beyotime Institute of Biotechnology). A total of 30 µg protein
lysate of each group was separated via 10% SDS-PAGE and transferred
to a PVDF membrane (Merck KGaA). Membranes were blocked with 5%
skimmed milk in TBS -0.05% Tween 20 (TBST) for 1.5 h at room
temperature and then incubated with primary antibodies at 4˚C
overnight. Primary antibodies included rabbit anti-Sox-9 antibody
(1:1,000; cat. no. bs-4177R; BIOSS), rabbit anti-HIF-1α antibody
(1:1,000; cat. no. bs-0737R; BIOSS), rabbit anti-GLUT1 antibody
(1:1,000; cat. no. bs-20173R; BIOSS), rabbit anti-aggrecan antibody
(1:1,000; cat. no. bs-1223R; BIOSS), mouse anti-collagen II
antibody (1:1,000; cat. no. bsm-33409M; BIOSS) and rabbit
anti-β-actin antibody (1:2,000; cat. no. AC038; ABclonal Biotech
Co., Ltd.). The membranes were then washed with TBST three times
and incubated with HRP-conjugated goat anti-rabbit IgG (H+L)
(1:5,000; cat. no. AS014; ABclonal Biotech Co., Ltd.) or
HRP-conjugated goat anti-mouse IgG (H+L) secondary antibody
(1:5,000; cat. no. AS003; ABclonal Biotech Co., Ltd.) for 1 h at
room temperature. The bands were detected using a bioimaging system
(Bio-Rad Laboratories, Inc.) using ECL reagent (Merck KGaA). The
bands were analyzed using ImageJ 2x software (National Institutes
of Health). β-actin was used to normalize the relative expression
of proteins.
Statistical analysis
Data are presented as the mean ± SEM. Statistical
analysis was performed using one-way ANOVA followed by Tukey's
multiple comparison test with GraphPad Prism 5.0 software (GraphPad
Software Inc.). The number of biological replicates was three, and
the experiments were performed at least three times. P<0.05 was
considered to indicate a statistically significant difference.
Results
Culture and identification of BMSCs
and ADSCs
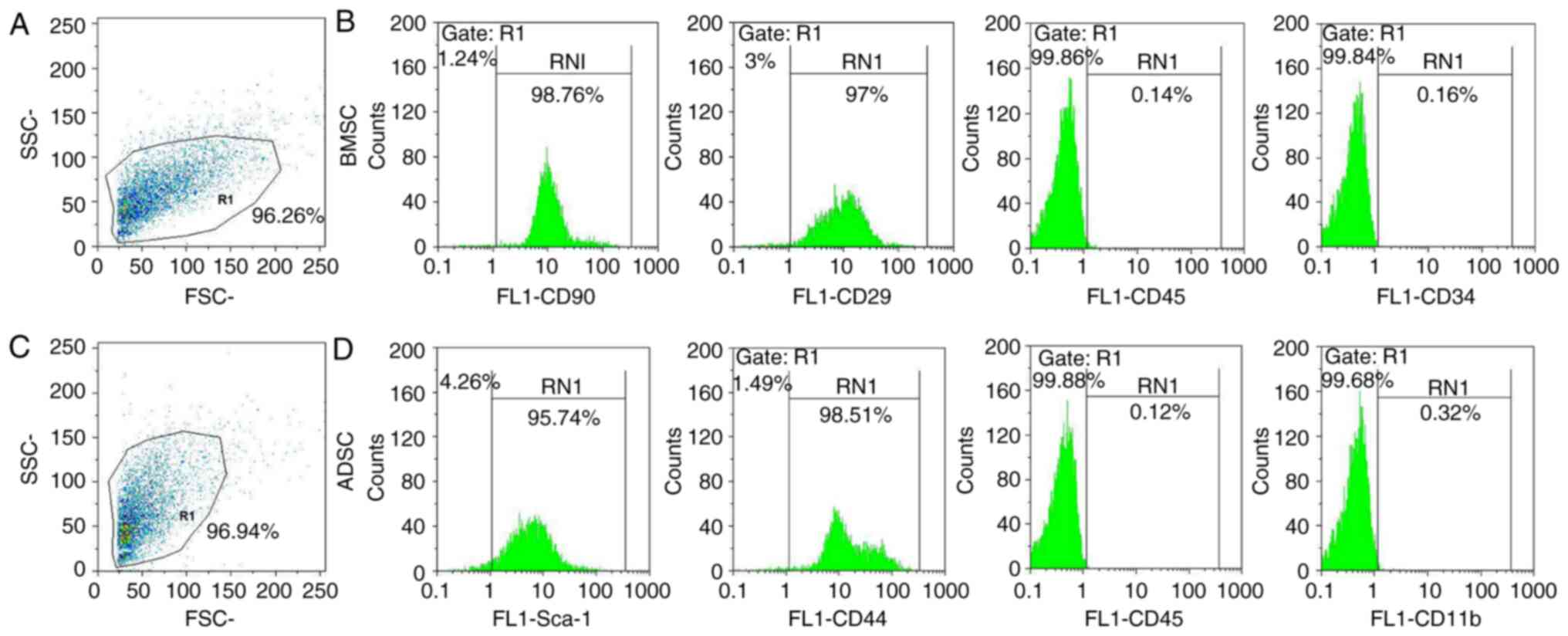
BMSCs and ADSCs were examined via flow cytometry.
The present results demonstrated that rat BMSCs were positive for
CD29 and CD90 and negative for CD45 and CD34(32) (Fig.
1A and B). Rat ADSCs expressed
high levels of a number of stem cell-specific markers (Sca-1 and
CD44) and were negative for CD45 and CD11b (26,33)
(Fig. 1C and D). Therefore, the present results
confirmed that rat ADSCs and BMSCs were successfully obtained and
cultured.
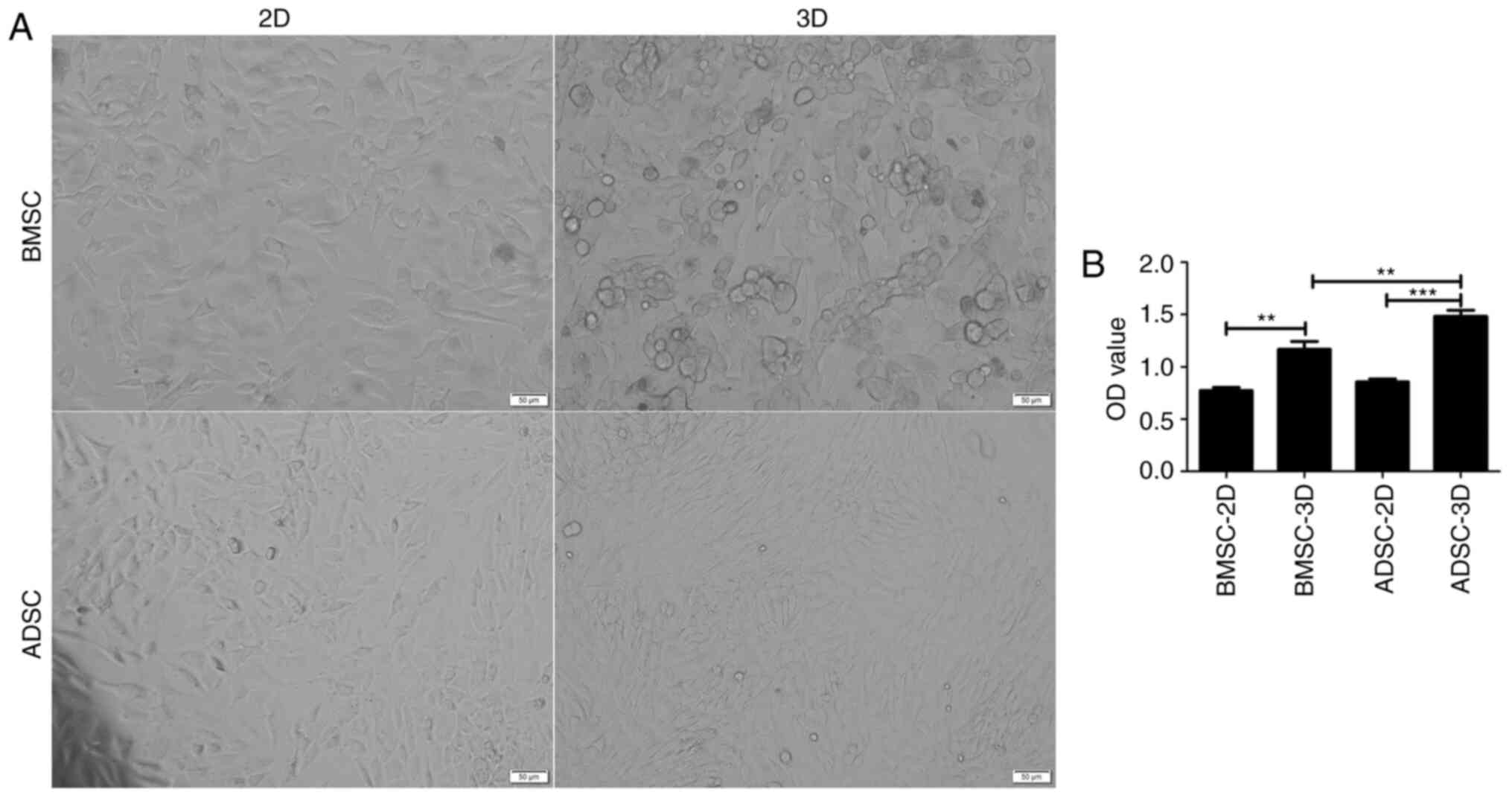
Viability of BMSCs and ADSCs
differentiated toward NPC-like cells
After 7 days of induction, the shape of BMSCs and
ADSCs cultured in 3D gradually changed from a long spindle-like
morphology to a round or triangular morphology. In addition, the
cells became shorter, indicating an alteration toward NPC-like
cells (10,34) (Fig.
2A). It was subsequently assessed whether these two cell types
also exhibited differences in cell viability. CCK-8 assay results
indicated that the OD values were higher in 3D culture (Fig. 2B) and that ADSCs in 3D culture
presented higher OD values compared with BMSCs. The current results
suggested that BMSCs and ADSCs exhibited an increased growth
potential in 3D than in 2D culture. Moreover, ADSCs were indicated
to present a greater potential for differentiation into NPC-like
cells compared with BMSCs.
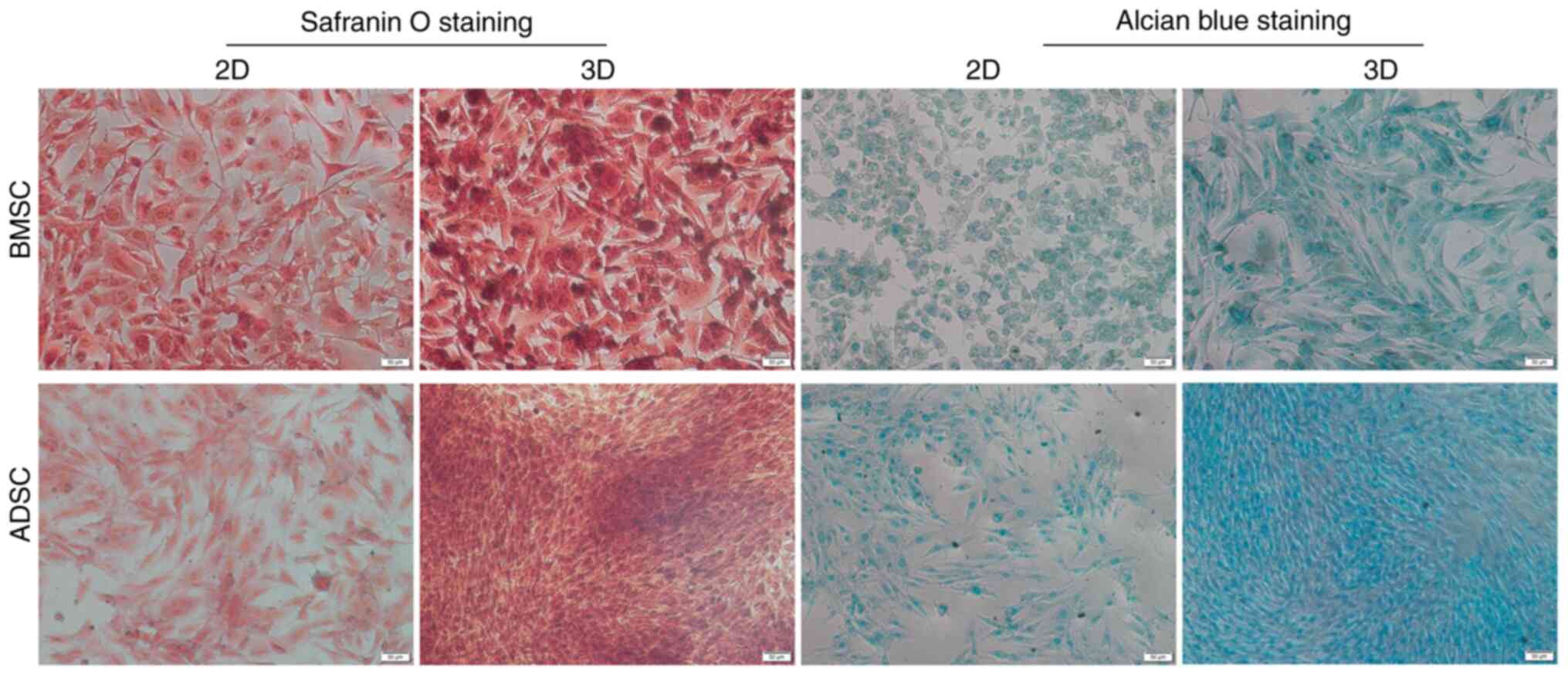
NPC-like differentiation of BMSCs and
ADSCs
To further examine the potential of BMSCs and ADSCs
to differentiate toward an NPC-like phenotype in 3D and 2D culture
under hypoxia, the secretion of GAGs was examined. GAGs are the
main components of ECM, and can be visualized via Alcian blue
staining. Proteoglycan formation was also examined in all groups
using safranin O staining. The present results indicated that BMSCs
and ADSCs in 3D culture exhibited higher levels of GAGs and
proteoglycans than those in the control groups; moreover, ADSCs in
3D culture exhibited higher levels of GAGs and proteoglycans than
BMSCs in 3D culture, but there was no notable difference between
the two groups in 2D culture (Fig.
3).
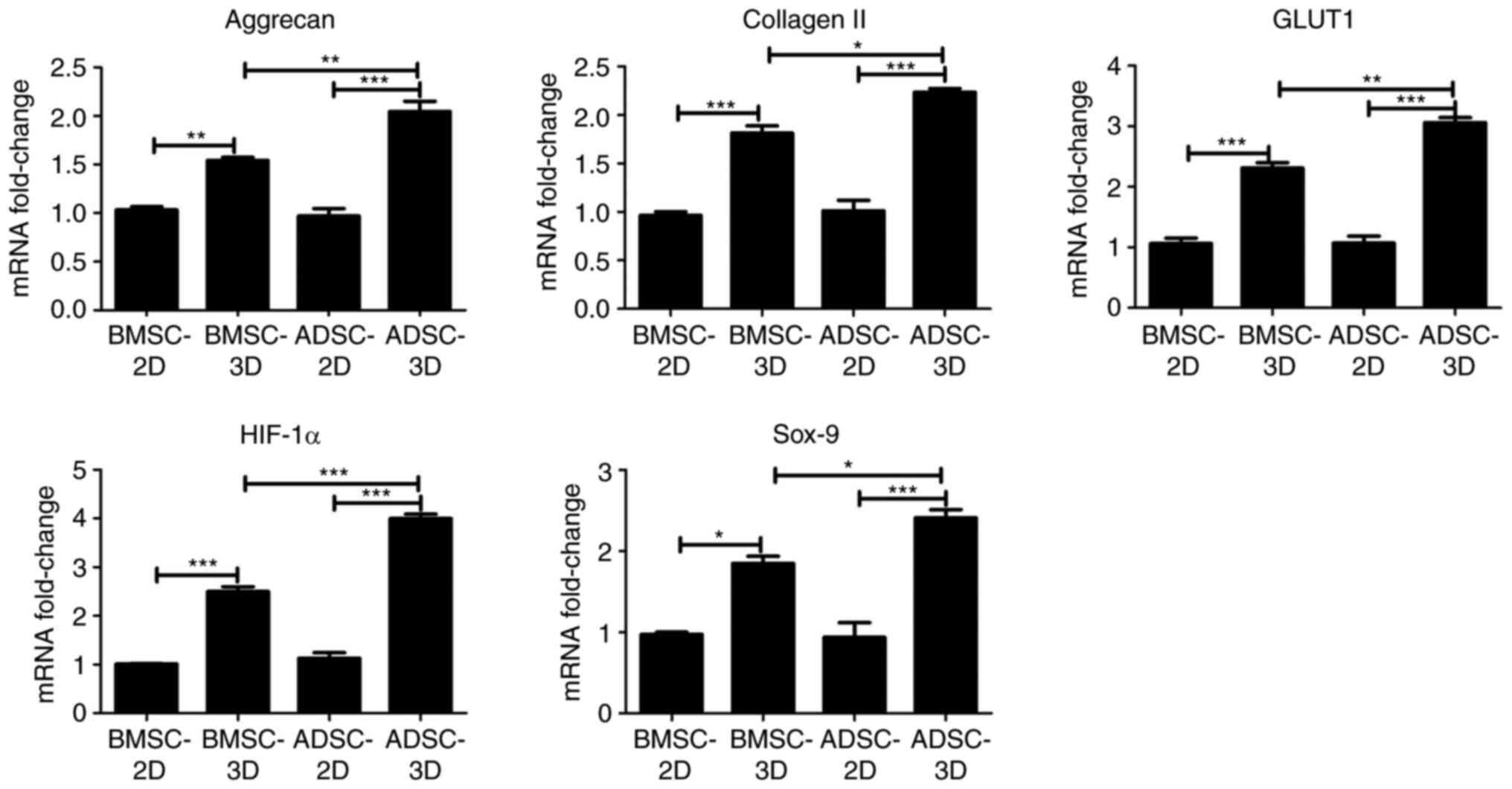
Gene expression analysis of BMSCs and
ADSCs differentiated toward NPC-like cells
To further investigate the difference between BMSCs
and ADSCs in their ability to differentiate toward NPC-like cells,
RT-qPCR and western blot assays were performed to examine
differences in the expression of NPC marker genes (HIF-1α and
GLUT1) and chondrocyte-specific genes (Sox-9, aggrecan and type II
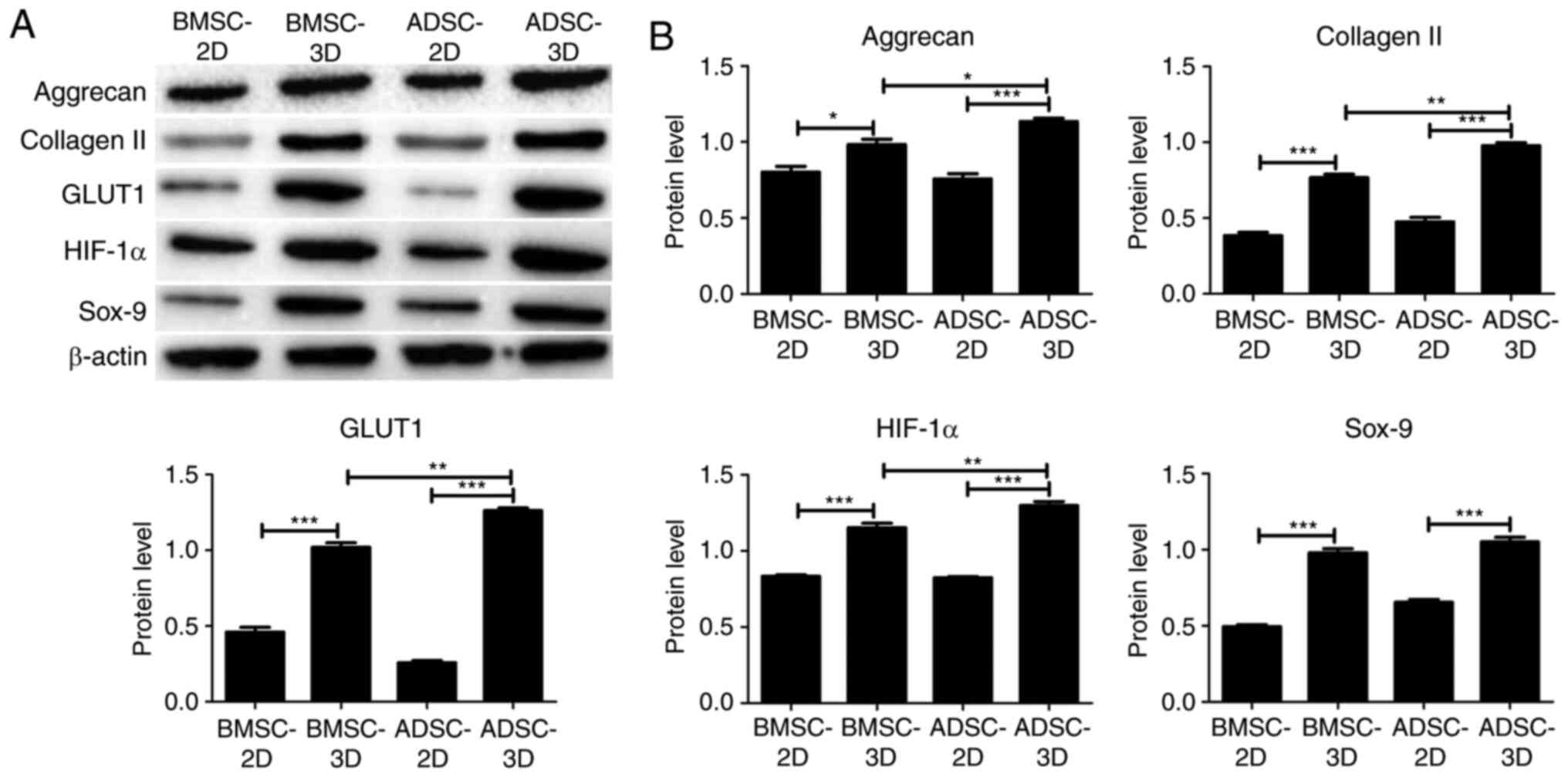
collagen) among the groups. As presented in Fig. 4, there was a significant increase in
the mRNA expression of HIF-1α, GLUT1, Sox-9, aggrecan and collagen
II in BMSCs and ADSCs in 3D culture compared with BMSCs and ADSCs
in 2D culture. In addition, the mRNA expression of HIF-1α, GLUT1,
Sox-9, aggrecan and collagen II was significantly higher in the
ADSC compared with the BMSC 3D culture group (Fig. 4). Similar results were obtained at
the protein expression level via western blotting, as presented in
Fig. 5. However, Sox-9 protein
expression was not significantly higher in the ADSC 3D culture
group when compared with the BMSC 3D culture group. The present
data indicated that ADSCs were exhibited a higher tendency to
differentiate into NPC-like cells compared with BMSCs upon
differentiation induction.
Discussion
IDD is one of the main clinical causes of neck pain
and low back pain (35). From a
physiological structure perspective, the intervertebral disc is in
a microenvironment with high mechanical strength, high osmotic
pressure and low nutrition and oxygen levels, and is prone to
degeneration with increasing age (36,37).
During the degenerative process, intervertebral disc cell apoptosis
increases and the matrix composition changes; in particular, there
is gradual loss of collagen and proteoglycans, resulting in a
deterioration of the local environment, which in turn accelerates
cell apoptosis (38). At the same
time, the number of intervertebral disc cells is limited, and the
regenerative ability of these cells is weak (36). Once degeneration occurs, it is
usually irreversible in the natural state (36). Although a number of surgical
procedures have been applied in an attempt to relieve clinical
symptoms, they all damage the structure of the intervertebral disc
and accelerate IDD to a certain extent (39,40).
The past decade has witnessed an increase in
regenerative medicine research and tissue engineering, with
exciting results for disc regeneration, such as new biomaterials
and improved cellular and molecular solutions (41,42).
Previous studies have addressed IDD with biological solutions,
including growth factors and cytokines, gene therapy, tissue
engineering and cell therapy based on the transplantation
technology (41,42). Cell therapy is a novel therapeutic
approach that aims to delay or even reverse IDD by replenishing the
body with stroma-rich cells to compensate for the lack of stromal
components (especially collagen and proteoglycans) (43,44).
Several types of cell transplantation therapies have entered the
rapid research stage (43,44).
Chen et al (45) and Liu et al (46) uncovered through in vitro
culture and iPSC identification that iPSCs exhibited the potential
to differentiate into myeloid nucleoid cells. Ni et al
(47) successfully isolated
embryonic-derived MSCs and revealed that the expression of nucleoid
cell markers increased significantly when they were cultured under
hypoxia. Jin et al (33)
induced adipogenic MSCs with TGF-b3 treatment and observed that the
expression of intervertebral disc-like cell markers and ECM
components was increased compared with adipogenic MSCs alone. Cao
et al (48) demonstrated
that BMSC can increase TGF-b and decrease the NF-κB pathway
activity to promote proteoglycan, predominately type II collagen
and Sox-9 gene expression, delaying the degeneration of
intervertebral discs. Therefore, BMSCs and ADSCs can be
differentiated into NPC-like cells, which may then be used for the
treatment of IDD.
The aim of the present investigation was to compare
the potential of these two types of cells to be induced to
differentiate into NPC-like cells and serve as seed cells for cell
therapy in the treatment of IDD. CCK-8 assay results indicated that
cell viability was increased in the 3D compared with the 2D
culture, and that ADSCs exhibited a higher viability than BMSCs,
which suggested that the two stem cell types were suitable for
growth in 3D culture, while ADSCs exhibited a greater growth
potential in 3D culture. The same number of cells were inoculated
into the plates, the final cell density in 3D culture was observed
to be higher compared with the 2D culture. Collagen II, Sox-9 and
aggrecan are chondrocyte-specific (49), while HIF-1α and GLUT1 are two NPC
markers (50,51); however, NPCs can also be regarded as
chondrocyte-like cells because of their expression of
chondrocyte-specific genes (52).
Risbud et al (53) suggested
that under hypoxic conditions (2% O2), rat BMSCs can be
differentiated toward an NPC-like phenotype in chondrogenic medium
within alginate beads. In the present study, BMSCs and ADSCs
treated with differentiating medium exhibited a significant
increase in the expression of NPC marker genes (HIF-1α, GLUT1) and
chondrocyte-specific genes (Sox-9, aggrecan and type II collagen)
in 3D culture compared with 2D culture. Furthermore, ADSCs
exhibited higher expression of these genes compared with BMSCs in
3D culture. In NPCs and chondrocytes, characteristic markers,
including type II collagen, aggrecan and Sox-9 are expressed
(33). HIF-1α is a key
transcription factor, which can be used as a phenotypic marker to
distinguish NPCs from chondrocytes (54). The present results demonstrated
ADSCs had a greater ability to differentiate into NPC-like cells
than BMSCs when 3D cultured.
Rat ADSCs can be differentiated toward an NPC-like
phenotype in 3D alginate hydrogels and cultured in an induction
medium containing TGF-β1 under hypoxic conditions (26). GAGs are the main components of ECM
and were produced by the differentiated BMSCs and ADSCs cultured in
differentiation medium, as detected by Alcian blue staining. Feng
et al (55) also indicated
that hypoxia markedly enhanced NPC phenotypes, resulting in a
greater production of collagen type II and GAGs in nanofibrous
scaffolds. Safranin O is a cationic basic dye (56) that can bind to 6-chondroitin sulfate
or keratinic sulfate, but not to collagen (57). Proteoglycans in NPCs mainly include
chondroitin sulfate or keratinic sulfate, and the content and
distribution of chondroitin sulfate or keratinic sulfate in NPCs
can be indirectly measured by safranin O staining (56,57).
In the present study, higher GAG and proteoglycan levels were
detected in 3D culture than in 2D culture; moreover, in 3D culture,
ADSCs produced higher levels of GAGs and proteoglycans compared
with BMSCs. The expression of NPC marker genes and
chondrocyte-specific genes was detected using RT-qPCR and western
blot assays, confirming that BMSCs and ADSCs produced higher
proteoglycan levels under the 3D compared with the 2D culture
condition, suggesting that the differentiation into NPC-like cells
under the 3D culture condition was enhanced. The present results
indicated that BMSCs and ADSCs differentiated toward an NPC-like
phenotype, and that ADSCs had a greater ability to differentiate
into NPC-like cells than BMSCs. Previous studies demonstrated that
both BMSCs (53,58) and ADSCs (27) could differentiate toward NPC-like
cells under the same conditions with a differentiating medium.
Although NPCs have been used to regenerate the disc
tissue in previous studies (8,9),
access to healthy NPCs, especially autologous cells, is very
limited in clinical settings. BMSCs and ADSCs are multi-potential
stem cells that can differentiate along several lineages,
potentially providing a suitable autologous stem cell source for
nucleus pulposus tissue regeneration (14). BMSCs or ADSCs are two of the most
widely studied types of MSCs and can be derived from a wide range
of sources, including bone and fatty tissue (13). Moreover, ADSCs are easier to obtain
than BMSCs as adipose tissue is more readily available and widely
distributed than bone in animals. Thus, these cell sources could be
candidate cells for the treatment of IDD-related diseases.
The current study presents a number of limitations.
Animal implantation experiments should be conducted to confirm that
hypoxic induction can contribute to maintaining the NPC phenotype,
and that BMSCs or ADSCs can be used to treat IDD-related diseases
in vivo. In future studies, the therapeutic function of BMSC
or ADSC-3D scaffold implantation will be explored on a mouse IDD
model.
It has been reported that BMSCs and ADSCs exhibited
an equal potential to be differentiated into osteocytes, adipocytes
and chondrocytes (59-61).
However, research comparing the differentiation of these two types
of stem cells into NPC-like cells in the same study has not been
previously reported. The present results are the first to indicate
that ADSCs have a greater ability to differentiate into NPC-like
cells than BMSCs when cultured in 3D hydrogels with differentiation
medium under hypoxic conditions, to the best of our knowledge.
Furthermore, while obtaining BMSCs represents a difficult
procedure, ADSCs can be easily harvested from patients via a simple
and minimally invasive approach. Thus, the present findings
provided a reference for the selection of candidate cells for the
treatment of IDD-related diseases.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by Yunnan Provincial
Science and Technology Department-Kunming Medical University
applied basic research joint special fund project (grant no.
201501UH00216).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XD and BL conceived and designed the study. XD, YG,
ZZ, YX and CL performed the experiments. XD, YG and HL analyzed the
data. XD and BL wrote the manuscript. XD and BL reviewed and edited
the manuscript. XD and BL confirm the authenticity of all the raw
data. All authors read and approved the manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Experiment and Ethics Committee of Kunming Medical University
(Kunming, China; approval no. KM20190301).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Maher C, Underwood M and Buchbinder R:
Non-specific low back pain. Lancet. 389:736–747. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wang MY, Vasudevan R and Mindea SA:
Minimally invasive lateral interbody fusion for the treatment of
rostral adjacent-segment lumbar degenerative stenosis without
supplemental pedicle screw fixation. J Neurosurg Spine. 21:861–866.
2014.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Colombini A, Lombardi G, Corsi MM and
Banfi G: Pathophysiology of the human intervertebral disc. Int J
Biochem Cell Biol. 40:837–842. 2008.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ghannam M, Jumah F, Mansour S, Samara A,
Alkhdour S, Alzuabi MA, Aker L, Adeeb N, Massengale J, Oskouian RJ
and Tubbs RS: Surgical anatomy, radiological features, and
molecular biology of the lumbar intervertebral discs. Clin Anat.
30:251–266. 2017.PubMed/NCBI View
Article : Google Scholar
|
|
5
|
Sakai D and Grad S: Advancing the cellular
and molecular therapy for intervertebral disc disease. Adv Drug
Deliv Rev. 84:159–171. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Luoma K, Riihimaki H, Luukkonen R,
Raininko R, Viikari-Juntura E and Lamminen A: Low back pain in
relation to lumbar disc degeneration. Spine (Phila Pa 1976).
25:487–492. 2000.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Buckwalter JA: Aging and degeneration of
the human intervertebral disc. Spine (Phila Pa 1976). 20:1307–1314.
1995.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Nishimura K and Mochida J: Percutaneous
reinsertion of the nucleus pulposus. An experimental study. Spine
(Phila Pa 1976). 23:1531–1538; discussion 1539. 1998.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Okuma M, Mochida J, Nishimura K, Sakabe K
and Seiki K: Reinsertion of stimulated nucleus pulposus cells
retards intervertebral disc degeneration: An in vitro and in vivo
experimental study. J Orthop Res. 18:988–997. 2000.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Feng G, Jin X, Hu J, Ma H, Gupte MJ, Liu H
and Ma PX: Effects of hypoxias and scaffold architecture on rabbit
mesenchymal stem cell differentiation towards a nucleus
pulposus-like phenotype. Biomaterials. 32:8182–8189.
2011.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yuan D, Chen Z, Xiang X, Deng S, Liu K,
Xiao D, Deng L and Feng G: The establishment and biological
assessment of a whole tissue-engineered intervertebral disc with
PBST fibers and a chitosan hydrogel in vitro and in vivo. J Biomed
Mater Res B Appl Biomater. 107:2305–2316. 2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Binch ALA, Fitzgerald JC, Growney EA and
Barry F: Cell-based strategies for IVD repair: Clinical progress
and translational obstacles. Nat Rev Rheumatol. 17:158–175.
2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Nan L, FX , Zhang L, Liu Y, Wang F
and Zhou SF: Research progresses of stem cell in the treatment of
intervertebral disc degenerative disease. Chinese Journal of Injury
Repair and Wound Healing (Electronic Edition). 13:134–138. 2018.(in
Chinese).
|
|
14
|
Strioga M, Viswanathan S, Darinskas A,
Slaby O and Michalek J: Same or not the same? Comparison of adipose
tissue-derived versus bone marrow-derived mesenchymal stem and
stromal cells. Stem Cells Dev. 21:2724–2752. 2012.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Han C, Jiang C, Yu C and Shen H:
Differentiation of transforming growth factor β1-induced
mesenchymal stem cells into nucleus pulposus-like cells under
simulated microgravity conditions. Cell Mol Biol (Noisy-le-grand).
61:50–55. 2015.PubMed/NCBI
|
|
16
|
Xu J, E XQ, Wang NX, Wang MN, Xie HX, Cao
YH, Sun LH, Tian J, Chen HJ and Yan JL: BMP7 enhances the effect of
BMSCs on extracellular matrix remodeling in a rabbit model of
intervertebral disc degeneration. FEBS J. 283:1689–1700.
2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Elabd C, Centeno CJ, Schultz JR, Lutz G,
Ichim T and Silva FJ: Intra-discal injection of autologous, hypoxic
cultured bone marrow-derived mesenchymal stem cells in five
patients with chronic lower back pain: A long-term safety and
feasibility study. J Transl Med. 14(253)2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Noriega DC, Ardura F, Hernández-Ramajo R,
Martín-Ferrero MÁ, Sánchez-Lite I, Toribio B, Alberca M, García V,
Moraleda JM, Sánchez A and García-Sancho J: Intervertebral disc
repair by allogeneic mesenchymal bone marrow cells: A randomized
controlled trial. Transplantation. 101:1945–1951. 2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Xu J, Qi DL, Pang XJ and Jing CW: Rabbit
nucleus pulposus cells facilitate differentiation of
adipose-derived stem cells into nucleus pulposus-like cells. Indian
J Cancer. 52 (Suppl 1):e17–e21. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Clarke LE, McConnell JC, Sherratt MJ,
Derby B, Richardson SM and Hoyland JA: Growth differentiation
factor 6 and transforming growth factor-beta differentially mediate
mesenchymal stem cell differentiation, composition, and
micromechanical properties of nucleus pulposus constructs.
Arthritis Res Ther. 16(R67)2014.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Roughley P, Hoemann C, DesRosiers E, Mwale
F, Antoniou J and Alini M: The potential of chitosan-based gels
containing intervertebral disc cells for nucleus pulposus
supplementation. Biomaterials. 27:388–396. 2006.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sakai D, Mochida J, Yamamoto Y, Nomura T,
Okuma M, Nishimura K, Nakai T, Ando K and Hotta T: Transplantation
of mesenchymal stem cells embedded in Atelocollagen gel to the
intervertebral disc: A potential therapeutic model for disc
degeneration. Biomaterials. 24:3531–3541. 2003.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Simona BR, Hirt L, Demkó L, Zambelli T,
Vörös J, Ehrbar M and Milleret V: Density gradients at hydrogel
interfaces for enhanced cell penetration. Biomater Sci. 3:586–591.
2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Loessner D, Stok KS, Lutolf MP, Hutmacher
DW, Clements JA and Rizzi SC: Bioengineered 3D platform to explore
cell-ECM interactions and drug resistance of epithelial ovarian
cancer cells. Biomaterials. 31:8494–8506. 2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li X, Wu A, Han C, Chen C, Zhou T, Zhang
K, Yang X, Chen Z, Qin A, Tian H and Zhao J: Bone marrow-derived
mesenchymal stem cells in three-dimensional co-culture attenuate
degeneration of nucleus pulposus cells. Aging (Albany NY).
11:9167–9187. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Xie LW, Fang H, Chen AM and Li F:
Differentiation of rat adipose tissue-derived mesenchymal stem
cells towards a nucleus pulposus-like phenotype in vitro. Chin J
Traumatol. 12:98–103. 2009.PubMed/NCBI
|
|
27
|
Hanson K, Isder C, Shogren K, Mikula AL,
Lu L, Yaszemski MJ and Elder BD: The inhibitory effects of
vancomycin on rat bone marrow-derived mesenchymal stem cell
differentiation. J Neurosurg Spine. 1–5. 2021.PubMed/NCBI View Article : Google Scholar : (Epub ahead of
print).
|
|
28
|
Pieróg J, Tamo L, Fakin R, Kocher G,
Gugger M, Grodzki T, Geiser T, Gazdhar A and Schmid RA: Bone marrow
stem cells modified with human interleukin 10 attenuate acute
rejection in rat lung allotransplantation. Eur J Cardiothorac Surg.
53:194–200. 2018.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Kinebuchi Y, Aizawa N, Imamura T, Ishizuka
O, Igawa Y and Nishizawa O: Autologous bone-marrow-derived
mesenchymal stem cell transplantation into injured rat urethral
sphincter. Int J Urol. 17:359–368. 2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Riahi M, Parivar K, Baharara J and Zandi
R: Evaluation of the repair of diaphyseal fracture of femoral bone
using bone marrow mesenchymal stem cells in nicotine-bearing rat.
Bratisl Lek Listy. 120:434–442. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Hu J, Deng G, Tian Y, Pu Y, Cao P and Yuan
W: An in vitro investigation into the role of bone marrow-derived
mesenchymal stem cells in the control of disc degeneration. Mol Med
Rep. 12:5701–5708. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Jin ES, Min J, Jeon SR, Choi KH and Jeong
JH: Analysis of molecular expression in adipose tissue-derived
mesenchymal stem cells: Prospects for use in the treatment of
intervertebral disc degeneration. J Korean Neurosurg Soc.
53:207–212. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wei G HY, Qin W, Liao C and Lin Z:
Differentiation of bone marrow mesenchymal stem cells into nucleus
pulposus-like cells after co-culture with nucleus pulposus cells.
Chin J Tissue Engineering Res. 17:7834–7839. 2013.
|
|
35
|
Vos T, Flaxman AD, Naghavi M, Lozano R,
Michaud C, Ezzati M, Shibuya K, Salomon JA, Abdalla S, Aboyans V,
et al: Years lived with disability (YLDs) for 1160 sequelae of 289
diseases and injuries 1990-2010: A systematic analysis for the
Global Burden of Disease Study 2010. Lancet. 380:2163–2196.
2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Vergroesen PP, Kingma I, Emanuel KS,
Hoogendoorn RJ, Welting TJ, van Royen BJ, van Dieën JH and Smit TH:
Mechanics and biology in intervertebral disc degeneration: A
vicious circle. Osteoarthritis Cartilage. 23:1057–1070.
2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chan S, Walser J, Käppeli P, Shamsollahi
M, Ferguson S and Gantenbein-Ritter B: Region specific response of
intervertebral disc cells to complex dynamic loading: An organ
culture study using a dynamic torsion-compression bioreactor. PLoS
One. 8(e72489)2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Anderson DG and Tannoury C: Molecular
pathogenic factors in symptomatic disc degeneration. Spine J. 5
(Suppl 6):260S–266S. 2005.PubMed/NCBI View Article : Google Scholar
|
|
39
|
van den Eerenbeemt KD, Ostelo RW, van
Royen BJ, Peul WC and van Tulder MW: Total disc replacement surgery
for symptomatic degenerative lumbar disc disease: A systematic
review of the literature. Eur Spine J. 19:1262–1280.
2010.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Miller LE and Block JE: Safety and
effectiveness of bone allografts in anterior cervical discectomy
and fusion surgery. Spine (Phila Pa 1976). 36:2045–2050.
2011.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Alini M, Roughley P, Antoniou J, Stoll T
and Aebi M: A biological approach to treating disc degeneration:
Not for today, but maybe for tomorrow. Eur Spine J 11 Suppl. 2
(Suppl 2):S215–S220. 2002.PubMed/NCBI View Article : Google Scholar
|
|
42
|
An HS, Thonar EJ and Masuda K: Biological
repair of intervertebral disc. Spine (Phila Pa 1976). 28 (Suppl
15):S86–S92. 2003.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hohaus C, Ganey TM, Minkus Y and Meisel
HJ: Cell transplantation in lumbar spine disc degeneration disease.
Eur Spine J 17 Suppl. 4 (Suppl 4):492–503. 2008.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Meisel HJ, Siodla V, Ganey T, Minkus Y,
Hutton WC and Alasevic OJ: Clinical experience in cell-based
therapeutics: Disc chondrocyte transplantation A treatment for
degenerated or damaged intervertebral disc. Biomol Eng. 24:5–21.
2007.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chen J, Lee EJ, Jing L, Christoforou N,
Leong KW and Setton LA: Differentiation of mouse induced
pluripotent stem cells (iPSCs) into nucleus pulposus-like cells in
vitro. PLoS One. 8(e75548)2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Liu K, Chen Z, Luo XW, Song GQ, Wang P, Li
XD, Zhao M, Han XW, Bai YG, Yang ZL and Feng G: Determination of
the potential of induced pluripotent stem cells to differentiate
into mouse nucleus pulposus cells in vitro. Genet Mol Res.
14:12394–12405. 2015.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Ni L, Liu X, Sochacki KR, Ebraheim M,
Fahrenkopf M, Shi Q, Liu J and Yang H: Effects of hypoxia on
differentiation from human placenta-derived mesenchymal stem cells
to nucleus pulposus-like cells. Spine J. 14:2451–2458.
2014.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Cao C, Zou J, Liu X, Shapiro A, Moral M,
Luo Z, Shi Q, Liu J, Yang H and Ebraheim N: Bone marrow mesenchymal
stem cells slow intervertebral disc degeneration through the NF-κB
pathway. Spine J. 15:530–538. 2015.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Sive JI, Baird P, Jeziorsk M, Watkins A,
Hoyland JA and Freemont AJ: Expression of chondrocyte markers by
cells of normal and degenerate intervertebral discs. Mol Pathol.
55:91–97. 2002.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Rajpurohit R, Risbud MV, Ducheyne P,
Vresilovic EJ and Shapiro IM: Phenotypic characteristics of the
nucleus pulposus: Expression of hypoxia inducing factor-1, glucose
transporter-1 and MMP-2. Cell Tissue Res. 308:401–407.
2002.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Richardson SM, Knowles R, Tyler J,
Mobasheri A and Hoyland JA: Expression of glucose transporters
GLUT-1, GLUT-3, GLUT-9 and HIF-1alpha in normal and degenerate
human intervertebral disc. Histochem Cell Biol. 129:503–511.
2008.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Cui X, Liu M, Wang J, Zhou Y and Xiang Q:
Electrospun scaffold containing TGF-β1 promotes human mesenchymal
stem cell differentiation towards a nucleus pulposus-like phenotype
under hypoxia. IET Nanobiotechnol. 9:76–84. 2015.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Risbud MV, Albert TJ, Guttapalli A,
Vresilovic EJ, Hillibrand AS, Vaccaro AR and Shapiro IM:
Differentiation of mesenchymal stem cells towards a nucleus
pulposus-like phenotype in vitro: Implications for cell-based
transplantation therapy. Spine (Phila Pa 1976). 29:2627–2632.
2004.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Risbud MV, Guttapalli A, Stokes DG,
Hawkins D, Danielson KG, Schaer TP, Albert TJ and Shapiro IM:
Nucleus pulposus cells express HIF-1 alpha under normoxic culture
conditions: A metabolic adaptation to the intervertebral disc
microenvironment. J Cell Biochem. 98:152–159. 2006.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Feng G, Li L, Liu H, Song Y, Huang F, Tu
C, Shen B, Gong Q, Li T, Liu L, et al: Hypoxia differentially
regulates human nucleus pulposus and annulus fibrosus cell
extracellular matrix production in 3D scaffolds. Osteoarthritis
Cartilage. 21:582–588. 2013.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Aaron RK, Jolly G, Ciombor DM and Barrach
HJ: A histochemical method for the demonstration of calcifying
cartilage. Calcif Tissue Int. 43:244–249. 1988.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Rosenberg L: Chemical basis for the
histological use of safranin O in the study of articular cartilage.
J Bone Joint Surg Am. 53:69–82. 1971.PubMed/NCBI
|
|
58
|
Fang Z, Yang Q, Luo W, Li GH, Xiao J, Li F
and Xiong W: Differentiation of GFP-Bcl-2-engineered mesenchymal
stem cells towards a nucleus pulposus-like phenotype under hypoxia
in vitro. Biochem Biophys Res Commun. 432:444–450. 2013.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Yoshimura H, Muneta T, Nimura A, Yokoyama
A, Koga H and Sekiya I: Comparison of rat mesenchymal stem cells
derived from bone marrow, synovium, periosteum, adipose tissue, and
muscle. Cell Tissue Res. 327:449–462. 2007.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Anokhina EB and Buravkova LB:
Heterogeneity of stromal precursor cells isolated from rat bone
marrow. Tsitologiia. 49:40–47. 2007.PubMed/NCBI
|
|
61
|
Keyser KA, Beagles KE and Kiem HP:
Comparison of mesenchymal stem cells from different tissues to
suppress T-cell activation. Cell Transplant. 16:555–562.
2007.PubMed/NCBI View Article : Google Scholar
|