Introduction
Neuropathic pain is caused by a lesion or disease of
the somatosensory system (1) and
characterized by positive and negative sensory symptoms, such as
the gain and loss of somatosensory function, which occurs alone or
in different combinations (2).
Neuropathic pain results from etiologically diverse disorders
affecting the peripheral or central nervous system or neurological
conditions of unknown etiology, and it either persists continuously
or presents as recurrent painful episodes (3). Chronic neuropathic pain may complicate
neuropathic symptoms and treatment decisions, leading to poor
outcomes and impaired patient quality of life (4). The diagnosis of neuropathic pain is
also complex due to diverse clinical features that are categorized
based on anatomical (such as peripheral and central) and
etiological (such as degenerative, traumatic, infectious metabolic
and toxic) factors (5). Further
research and clinical trials should be implemented to improve the
diagnosis and treatment of neuropathic pain. The present study
aimed to validate the anti-inflammatory and analgesic effects of
dexmedetomidine (DEX) on neuropathic pain in a rat model of chronic
constriction injury (CCI).
DEX is a potent and selective α2-adrenoceptor
agonist with sedative, anxiolytic, sympatholytic and analgesic
effects (6). This drug is used to
induce short- and long-term sedation in patients in intensive care
units and can effectively suppress delirium (7). DEX has been reported to have the
potential to prevent acute pain in adults who undergo abdominal
surgery (8). Moreover, a steadily
increasing number of studies have investigated the inhibitory
effect of DEX on neuropathic pain. For example, DEX suppressed
neuropathic pain by inhibiting P2X purinoceptor 7 through ERK
regulation in a rat model of CCI (9). DEX can protect the nervous system via
anti-inflammatory, anti-excitotoxic, antiapoptotic, antioxidative
and other mechanisms (10).
NLR family pyrin domain containing 3 (NLRP3), which
belongs to the nucleotide-binding oligomerization domain-like
receptor family of proteins, is a pattern recognition receptor that
contributes to the formation of inflammasomes (11). Among all inflammasomes, the NLRP3
inflammasome has been extensively investigated and verified to
serve a critical role in innate immunity and the pathology of human
diseases (12). NLRP3 promotes
inflammation through the cleavage of the proinflammatory cytokines
IL-1β and IL-18 by stimulating caspase-1(13). Grace et al (14) revealed that morphine promoted the
intensity and duration of neuropathic pain by activating the NLRP3
inflammasome. DEX has been indicated to modulate the NLRP3
inflammasome in animal models of acute lung and kidney injury
(15,16). However, whether NLRP3 is implicated
in the DEX-induced reduction in inflammatory responses and
neuropathic pain has not been yet fully elucidated.
Nuclear factor-erythroid 2-related factor 2 (Nrf2)
is a multifunctional protein that modulates antioxidant and other
cytoprotective genes (such as GPX2 and GCLC) and is involved in
inflammatory processes (17,18).
Plumbagin, a naphthoquinone derived from the plant Plumbago, has
been indicated to alleviate CCI-induced neuropathic pain by
upregulating Nrf2(19). In
addition, the new biphenyl diester derivative AB-38b has been
demonstrated to attenuate diabetic nephropathy by suppressing NLRP3
via Nrf2 activation (20). However,
the Nrf2 signaling-mediated regulation of NLRP3 in neuropathic pain
has rarely been investigated. A previous study revealed that DEX
exerted neuroprotective effects via the Nrf2 signaling pathway
(21). The present study aimed to
investigate whether DEX-induced activation of Nrf2 further
regulated NLRP3 to ameliorate neuropathic pain.
Materials and methods
Animal experiments
Male Sprague-Dawley (SD) rats (n=138; weight,
200-220 g; age, 2 months) were purchased from Hunan SJA Laboratory
Animal Co., Ltd. The rats were subjected to a 12/12 h light/dark
cycle at room temperature (23±1˚C) and 60-65% humidity and were
provided standard food and water for at least 1 week. All animal
experiments were approved by the Animal Protection and Use
Committee of Hunan Provincial People's Hospital (Changsha, China)
and strictly followed the ‘Guidelines for the Use and Management of
Laboratory Animals’ issued by the National Institutes of Health and
Animal Research: Reporting In Vivo Experiments Guidelines
(22). Adult male and female rats
respond differentially to harmful stimuli. Among rodents, females
are more sensitive compared with males to noxious stimuli, and
males generally exhibit stronger stress-induced analgesia (23). Therefore, male rats were used for
the experiments in the present study.
A total of 138 SD rats were divided into different
groups in a randomized manner (6 rats per group). Rats in control
groups (n=6x2) were left untreated. Rats in sham groups (n=6x3)
underwent surgery, during which no nerve ligation was performed.
Rats in the model group were subjected to sciatic nerve ligation
for induction of CCI after being anesthetized with pentobarbital
sodium (40 mg/kg). The exposed sciatic nerves were separated and
ligated with four 4-0 chromic gut suture (Ethicon, Inc.).
After the model establishment, the rats in each
group were subjected to the corresponding treatment. The groups
were as follows: Model group (n=6x3; untreated model rats), DEX+1
group (n=6; intraperitoneally injected with 1 µg/kg DEX every day
for 7 days after the operation; Jiangsu Nhwa Pharmaceutical Co.,
Ltd.), DEX+2 (n=6; intraperitoneally injected with 2 µg/kg DEX for
7 days post-operation), DEX+5 group (n=6; intraperitoneally
injected with 5 µg/kg DEX for 7 days post-operation), DEX group
(n=6x2; intraperitoneally injected with 5 µg/kg DEX for 7 days
post-operation), MCC950 group [n=6x2; intraperitoneally injected
with 50 µg/kg MCC950 (NLRP3 antagonist) for 7 days post-operation;
MedChemExpress], nigericin group [n=6x2; intraperitoneally injected
with 1 mg/kg nigericin (NLRP3 activator) for 7 days post-operation;
MedChemExpress], DEX + ML385 group [n=6x2; intraperitoneally
injected with 5 µg/kg DEX and 30 mg/kg ML385 (Nrf2 inhibitor) for 7
days post-operation; Selleck Chemicals], DEX + nigericin group
(n=6x2; intraperitoneally injected with 5 µg/kg DEX and 1 mg/kg
nigericin for 7 days post-operation) and MCC950 + ML385 group
(n=6x2; intraperitoneally injected with 30 mg/kg ML385 and 50 µg/kg
MCC950 for 7 days post-operation). The rats were deeply
anesthetized with an intraperitoneal injection of 50 mg/kg
thiopental sodium and euthanatized via cervical dislocation.
Subsequently, ~1 ml of blood was collected from each rat and
centrifuged at 1,000 x g, 4˚C for 10 min. The blood samples were
stored at -20˚C. The L4-L6 spinal cords were collected from the
rats and immediately stored in an ultra-low temperature
refrigerator (-80˚C). The experimental timetable is presented in
Fig. 1.
 | Figure 1Experimental schedule. A total of 138
rats were included in the present study. A total of 84 rats
underwent behavioral tests on day 0, 1, 3, 7 and 14 after the
operation, including the DEX+1, DEX+2 and DEX+5 groups, two sham
groups, two model groups and one control, DEX, MCC950, nigericin,
DEX + ML385, MCC950 + ML385 and DEX + nigericin groups (n=6 rats
per group). A total of 54 rats were sacrificed for spinal cord
collection 7 days after the drug administration, including one
control, sham, model, DEX, MCC950, nigericin, DEX + ML385, MCC950 +
ML385 and DEX + nigericin groups (n=6 rats per group). |
Animal behavioral analysis
The rats were subjected to behavioral tests on day
0, 1, 3, 7 and 14 post-operatively. These rats were randomly
assigned to the control, sham, model, DEX+1, DEX+2, DEX+5, DEX,
MCC950, nigericin, DEX + ML385, MCC950 + ML385 and DEX + nigericin
groups and were treated as aforementioned (n=6 per group). The
behavioral tests were conducted between 9:00 and 12:00 a.m. in a
quiet environment. The rats were placed in wire cages for 30 min
before the experiment. The mechanical withdrawal threshold (MWT) of
the rats was measured three times at an interval of 5 min using an
automated dynamic plantar aesthesiometer (Ugo Basile S.R.L.). The
minimum retraction force (G) of the rats' right hind paw was
recorded, and the average value of the power that induced a
reliable retreat was recorded as the threshold. Quick withdrawal or
licking of the paws in response to stimuli was considered a
positive reaction.
Hematoxylin and eosin (H&E)
staining
L4-L6 spinal cords were collected from rats in the
control, sham, model, DEX, MCC950, nigericin, DEX + ML385, MCC950 +
ML385 and DEX + nigericin groups (n=6 per group) 7 days
post-operation. The spinal cords were fixed in 4% paraformaldehyde
at 4˚C for 48 h and then dehydrated in 30% sucrose at 4˚C
overnight. The spinal cords were embedded in paraffin and cut into
5-µm sections for H&E staining. At room temperature, the
sections were sequentially immersed in xylene for 5 min, 100%
ethanol for 10 min, 95% ethanol for 5 min and 75% ethanol for 5 min
and finally washed with distilled water. The deparaffinized
sections were stained with hematoxylin at room temperature for 5
min, soaked in deionized water for 5 min, then stained with eosin
for 1 min. Finally, the spinal cord sections were observed using a
light microscope (ABX50; Olympus Corporation). The scoring criteria
of spinal cord injury were as follows: 0=No lesion; 1=gray matter
containing 1-5 eosinophilic neurons; 2=gray matter containing 6-10
eosinophilic neurons; 3=gray matter containing >10 eosinophilic
neurons; 4=infarction of <1/3 of the gray matter area;
5=infarction of 1/3-1/2 of the gray matter area; and 6=infarction
of >1/2 of the gray matter area.
TUNEL staining
L4-L6 spinal cords were collected from rats in the
control, sham, model, DEX, MCC950, nigericin, DEX + ML385, MCC950 +
ML385 and DEX + nigericin groups (n=6 per group) 7 days
post-operation. The tissues were fixed, dehydrated, paraffin
embedded and sliced as aforementioned. Paraffin sections of the
spinal cords were dewaxed with xylene and hydrated using alcohol
gradients (100% ethanol, 10 min; 95%, 5 min; 75%, 5 min; distilled
water). Subsequently, the spinal cord sections were treated with
proteinase K at room temperature for 30 min and immersed in
enhanced endogenous peroxidase blocking solution (100%; cat. no.
P0100B; Beyotime Institute of Biotechnology) at room temperature
for 10 min. The tissues were incubated with TUNEL detection
solution (cat. no. C1091; Beyotime Institute of Biotechnology) for
60 min at 37˚C in the dark. Finally, the spinal cord sections were
washed with PBS three times, mounted with neutral balsam and
observed using a light microscope. A total of three non-overlapping
fields were randomly selected from each slice. The apoptosis rate
(%) was determined as the number of brown cells/total number of
cells x100%.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from the L4-L6 spinal cords
of rats in the control, sham, model, DEX and DEX + ML385 groups
(n=6 per group) 7 days post-operation using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's instructions. RNA was reverse transcribed into
cDNA using a Universal cDNA Synthesis kit (Promega Corporation)
according to the manufacturer's instructions. Gene expression was
analyzed using a LightCycler 480 (Roche Diagnostics) in accordance
with the instructions of the fluorescence quantitative PCR kit
(SYBR Green PCR kit; Takara Bio, Inc.). The reaction conditions
were as follows: 5 min of pre-denaturation at 95˚C, followed by 40
cycles of 10 sec of denaturation at 95˚C, 10 sec of annealing at
60˚C and 20 sec of extension at 72˚C. Relative quantification was
performed using the comparative 2-∆∆Cq method (24), with GAPDH as an internal reference.
The primer sequences used are presented in Table I.
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Primer name | Sequence,
5'-3' |
|---|
| Nrf2 forward |
AGGTTGCCCACATTCCCAAA |
| Nrf2 reverse |
AGTGACTGAAACGTAGCCGA |
| GAPDH forward |
TTCTGGGATACACGGAGCAC |
| GAPDH reverse |
TACCAGCACCAGCGTCAAAG |
Western blotting
The L4-L6 segments of the spinal cords were
collected from rats in the control, sham, model, DEX, MCC950,
nigericin, DEX + ML385, MCC950 + ML385 and DEX + nigericin groups
(n=6 per group) 7 days post-operation and mixed with RIPA buffer
(Beyotime Institute of Biotechnology) containing protease and
phosphatase inhibitors. Total protein was extracted from the
lysates after centrifugation at 8,759 x g for 30 min at 4˚C, or
nucleoproteins were extracted using a nuclear protein extraction
kit (cat. no. R0050, Beijing Solarbio Science & Technology Co.,
Ltd.) according to the manufacturer's instructions. Protein
concentration was measured using a BCA kit. The proteins (100
µg/well) were separated using 8% SDS-PAGE and transferred onto a
PVDF membrane. The membrane was immersed in 5% non-fat milk at room
temperature for 1 h and then incubated with primary antibodies
against GAPDH (1:10,000; cat. no. ab181602), Nrf2 (1:100; cat. no.
ab137550) and NLRP3 (1:500; cat. no. ab214185) (all Abcam) at 4˚C
overnight. After being washed with 0.1% PBS-Tween-20 (TBST) three
times, the membrane was incubated with horseradish
peroxidase-labeled goat anti-rabbit IgG secondary antibody
(1:5,000; cat. no. CW0103; CoWin Biosciences) at room temperature
for 30 min. Finally, the membrane was washed with TBST four times
and subjected to color development with ECL (Proanti Biotechnology
Development Co., Ltd.). The blots were detected by a
chemiluminescence imaging system (GE Healthcare). The greyscale of
blots was analyzed by ImageJ (version 1.46; National Institutes of
Health).
ELISA
Whole blood of rats in the control, sham, model,
DEX, MCC950, nigericin, DEX + ML385, MCC950 + ML385 and DEX +
nigericin groups (n=6 per group) was mixed with EDTA and then
centrifuged at 1,000 x g for 10 min at 4˚C to collect the
supernatant. The expression levels of TNF-α (cat. no. RTA00), IL-1β
(cat. no. RLB00), IL-6 (cat. no. R6000B) and IL-10 (cat. no. R1000)
were measured using commercial ELISA kits (R&D Systems, Inc.)
in accordance with the manufacturer's instructions.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 7.0 (GraphPad Software, Inc.). Histopathological scores are
presented as the median ± interquartile range and were analyzed
using Kruskal-Wallis followed by Dunn's post hoc test. All other
data were normally distributed and are presented as the mean ±
standard deviation. Unpaired Student's t-test was used to compare
differences between two groups. One-way ANOVA followed by Tukey's
post hoc test was used for multigroup comparisons. P<0.05 was
considered to indicate a statistically significant difference.
Results
DEX reduces neuropathic pain in CCI
rats
CCI rats were injected with different concentrations
of DEX (1, 2 and 5 µg/kg) to determine the optimal dose for
subsequent experiments. MWT was measured to assess the pain
sensitivity of the rats. Compared with the sham group, the model
group exhibited a significant decrease in the MWT (Fig. 2A; P<0.01). Compared with the
model group, the DEX+1 group demonstrated no significant change in
the MWT, while the DEX+2 group and DEX+5 group exhibited
significantly increased MWTs (Fig.
2A; both P<0.05). Among the three groups, the DEX+5 group
demonstrated the highest increase in the MWT. Therefore, 5 µg/kg
DEX was used for subsequent experiments.
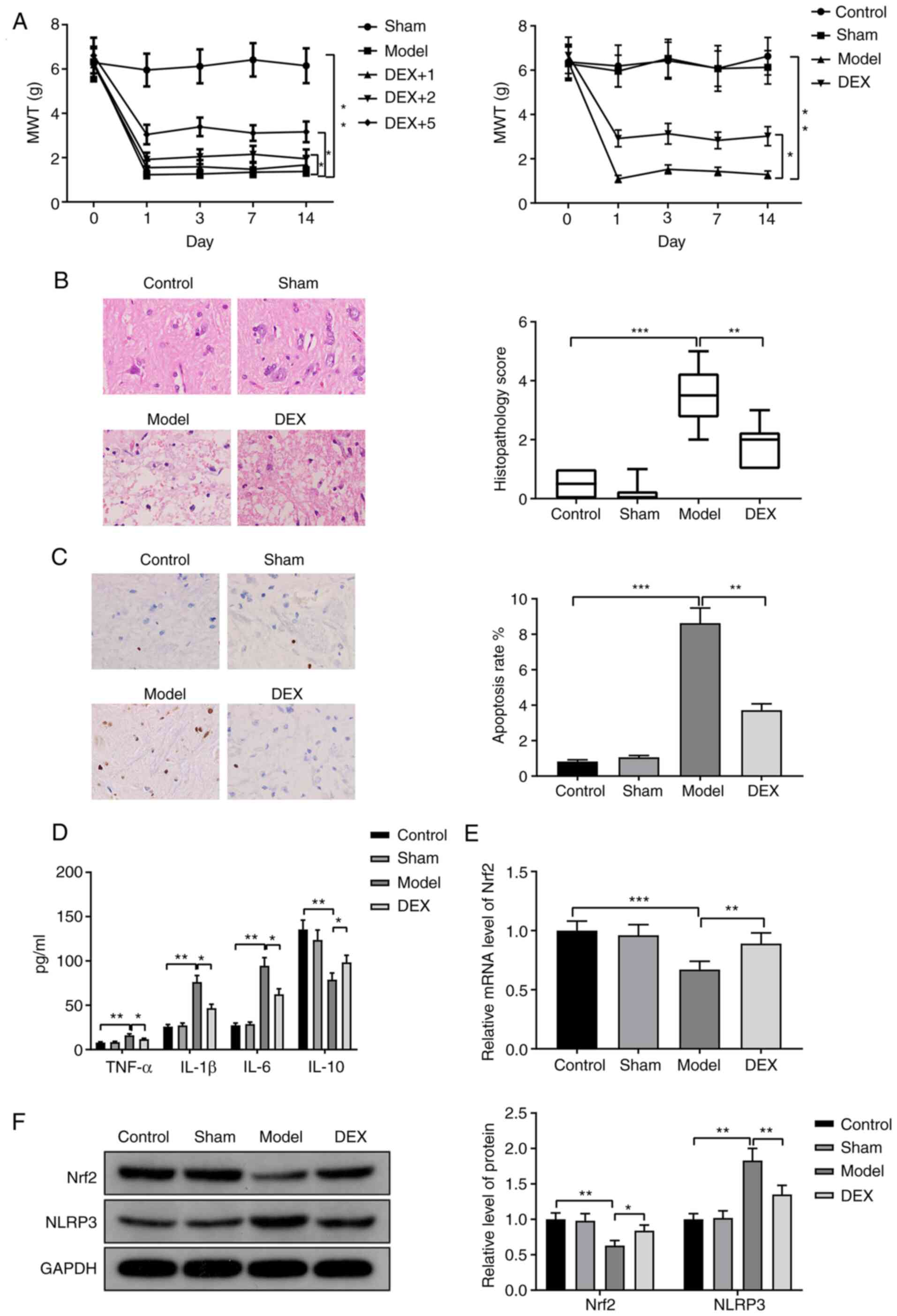 | Figure 2DEX reduces neuropathic pain in CCI
rats. (A) CCI rats were injected with 1, 2 or 5 µg/kg DEX before
MWT of the rats was measured. (B) CCI rats were injected with 5
µg/kg DEX and hematoxylin and eosin staining revealed the spinal
cord injury (magnification, x400). Each set of data was arranged
from X1 to Xn in ascending order. The median values were equal
(Xn/2+Xn/2+1)/2 when ‘n’ was even. The number of rats in each group
was even (n=6), so the median values were the average of X3+X4. (C)
TUNEL staining was performed to examine the apoptosis in the spinal
cords (magnification, x400). (D) The expression levels of
inflammatory factors were measured via ELISA. (E) Reverse
transcription-quantitative PCR was performed to detect the
expression of Nrf2. (F) Western blotting was performed to detect
the expression levels of Nrf2 and NLRP3. n=6 per group.
*P<0.05; **P<0.01;
***P<0.001. DEX, dexmedetomidine; CCI, chronic
constriction injury; MWT, mechanical withdrawal threshold; Nrf2,
nuclear factor erythroid 2-related factor 2; NLRP3, NLR family
pyrin domain containing 3; g, pressure unit. |
CCI rats were injected with 5 µg/kg DEX. As
demonstrated in Fig. 2A, the MWT
was decreased in the model group compared with the control group
(P<0.01), but increased in the DEX group compared with the model
group (P<0.05). This result indicated that the rats in the model
group had the highest pain sensitivity, whereas DEX reduced the
pain sensitivity of CCI rats.
H&E staining demonstrated that spinal injury was
increased in the model group compared with the control group
(P<0.001) but was reduced in the DEX group compared with the
model group (P<0.01; Fig. 2B).
The number of TUNEL-positive cells in the spinal cord was increased
in the model group compared with the control group (P<0.001) but
was decreased in the DEX group compared with the model group
(P<0.01) (Fig. 2C). TNF-α, IL-1β
and IL-6 expression levels were increased in the model group
compared with the control group (P<0.01), but were decreased in
the DEX group compared with the model group (P<0.05) (Fig. 2D). The expression patterns of IL-10
were the opposite of those of TNF-α, IL-1β and IL-6 in the model
and DEX groups (Fig. 2D; P<0.01
vs. the control group and P<0.05 vs. the model group). RT-qPCR
and western blotting results demonstrated that the expression
levels of Nrf2 mRNA and protein were downregulated in the model
group compared with the control group (Fig. 2E, P<0.001; Fig. 2F, P<0.01), but were upregulated
in the DEX group compared with the model group (Fig. 2E, P<0.01; Fig. 2F, P<0.05). Moreover, the protein
expression level of NLRP3 was upregulated in the model group
compared with the control group, but was downregulated in the DEX
group compared with the model group (Fig. 2F; both P<0.01). There was no
significant difference between the sham group and the control
group. These results demonstrated that DEX suppressed inflammation
and apoptosis in the spinal cord to reduce neuropathic pain in CCI
rats.
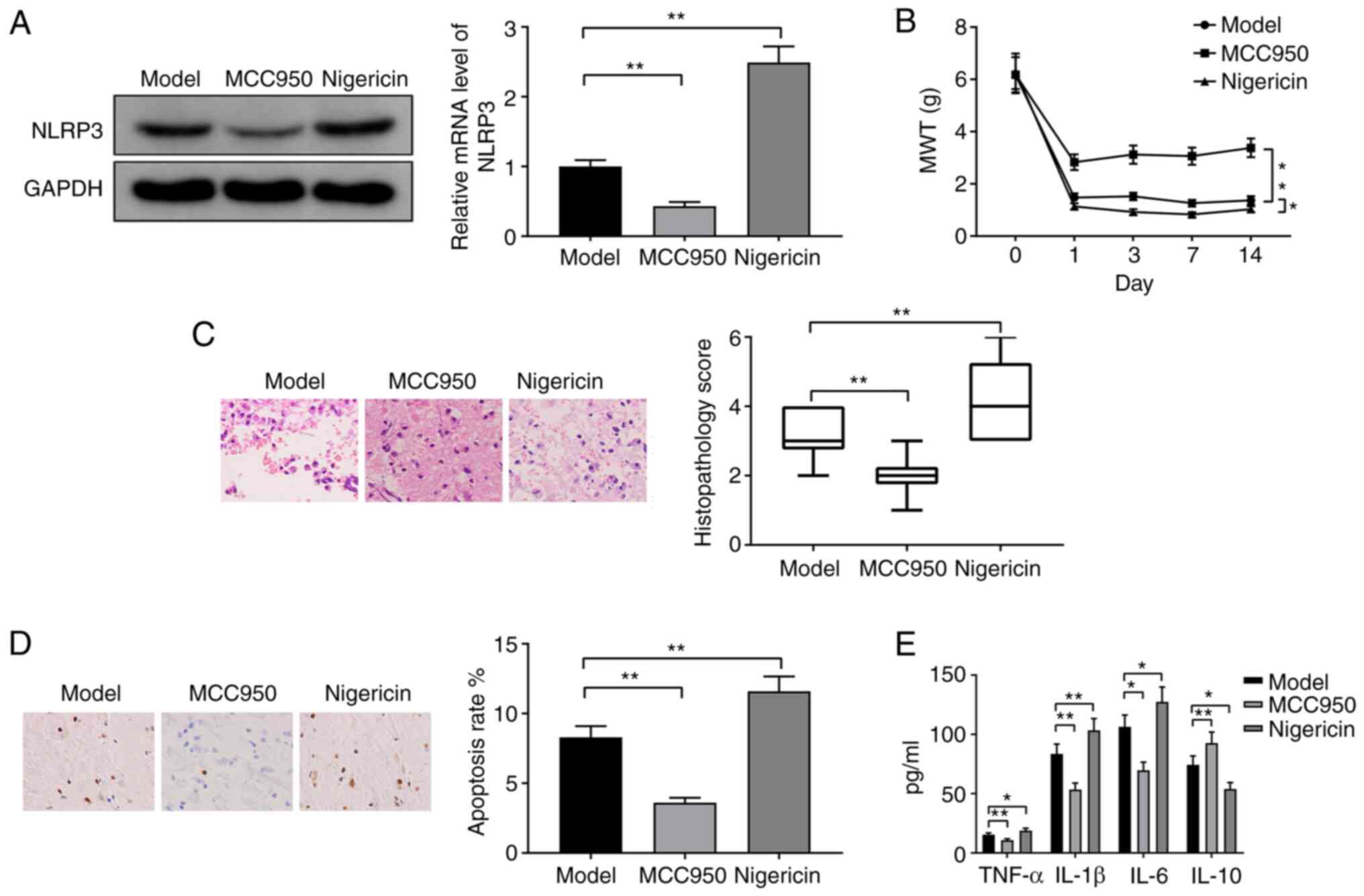
NLRP3 increases neuropathic pain in
CCI rats
CCI rats were injected with the NLRP3 antagonist
MCC950 or the NLRP3 activator nigericin. Western blotting revealed
downregulation of NLRP3 in the MCC950 group and upregulation of
NLRP3 in the nigericin group compared with the model group
(Fig. 3A; both P<0.01). These
results demonstrated the successful suppression or activation of
NLRP3 in CCI rats. MWT was significantly increased in the MCC950
group (Fig. 3B; P<0.01), but was
significantly decreased in the nigericin group compared with the
model group (Fig. 3B; P<0.05).
H&E staining indicated that spinal injury was reduced in the
MCC950 group but was increased in the nigericin group compared with
the model group (Fig. 3C; both
P<0.01). The TUNEL staining results indicated that, compared
with the model group, the apoptosis rate was significantly
decreased in the MCC950 group, but was increased in the nigericin
group (Fig. 3D; both P<0.01).
Decreases in the expression levels of TNF-α, IL-1β and IL-6 and an
increase in IL-10 were observed in the MCC950 group compared with
the model group, while the inverse expression patterns of these
cytokines were detected in the nigericin group (Fig. 3E; P<0.05). These results
suggested that NLRP3 could promote neuropathic pain in CCI
rats.
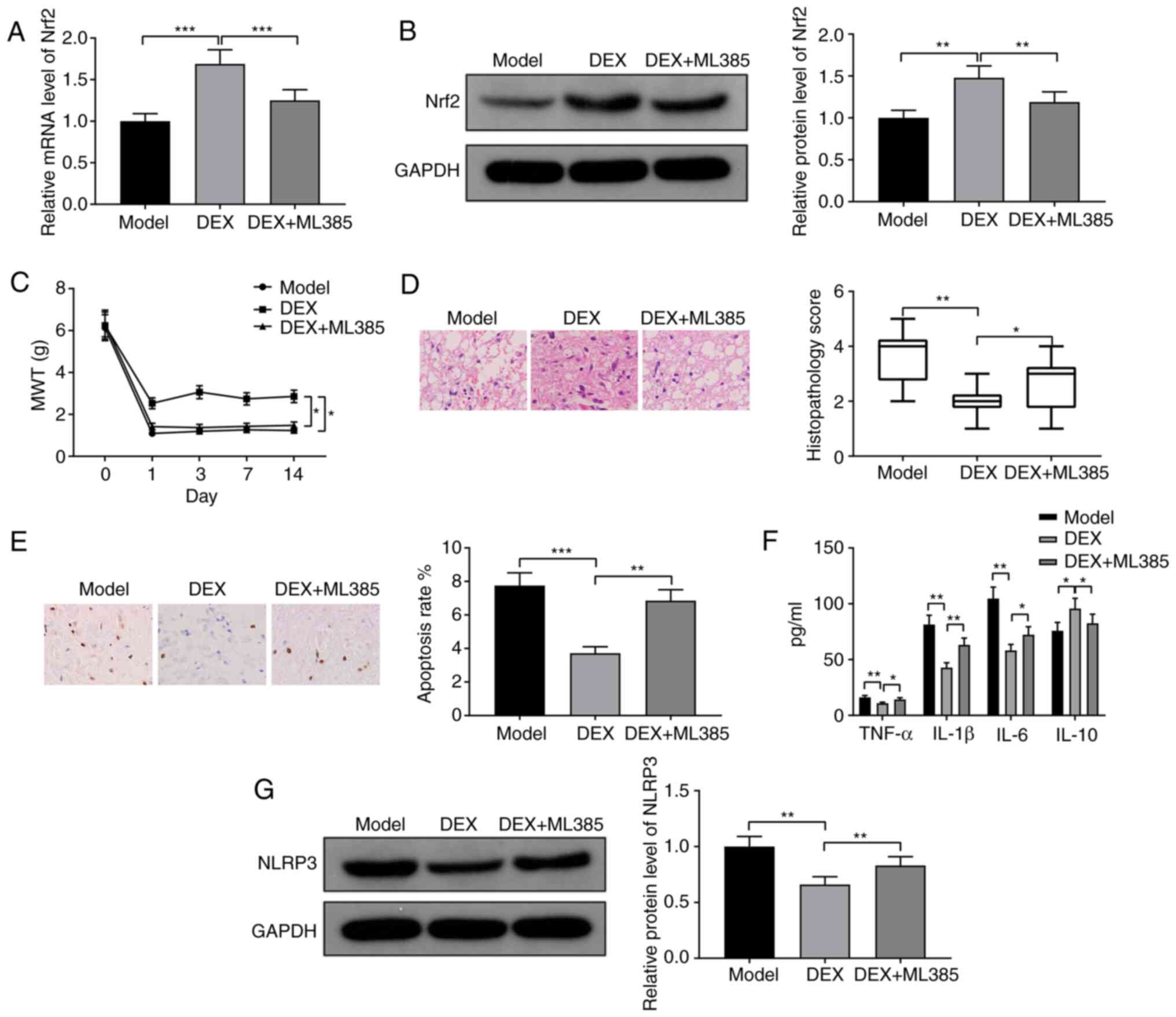
DEX reduces neuropathic pain by
activating Nrf2 in CCI rats
CCI rats were injected with the Nrf2 inhibitor ML385
together with DEX. RT-qPCR and western blotting demonstrated that
Nrf2 was downregulated in the DEX + ML385 group compared with the
DEX group, indicating the effectiveness of ML385 (Fig. 4A, P<0.001; Fig. 4B, P<0.01). The DEX + ML385 group
exhibited significantly decreased MWT compared with the DEX group
(Fig. 4C; P<0.05). H&E and
TUNEL staining demonstrated that spinal injury and apoptosis rate
were exacerbated in the DEX + ML385 group compared with the DEX
group (Fig. 4D, P<0.05; Fig. 4E, P<0.01). The expression levels
of TNF-α, IL-1β and IL-6 were increased, while the expression of
IL-10 was decreased in the DEX + ML385 group compared with the DEX
group (Fig. 4F; all P<0.05).
Moreover, the expression level of NLRP3 was also upregulated in the
DEX + ML385 group compared with the DEX group (Fig. 4G; P<0.01). In conclusion, DEX
promoted the expression of Nrf2 to alleviate neuropathic pain in
CCI rats.
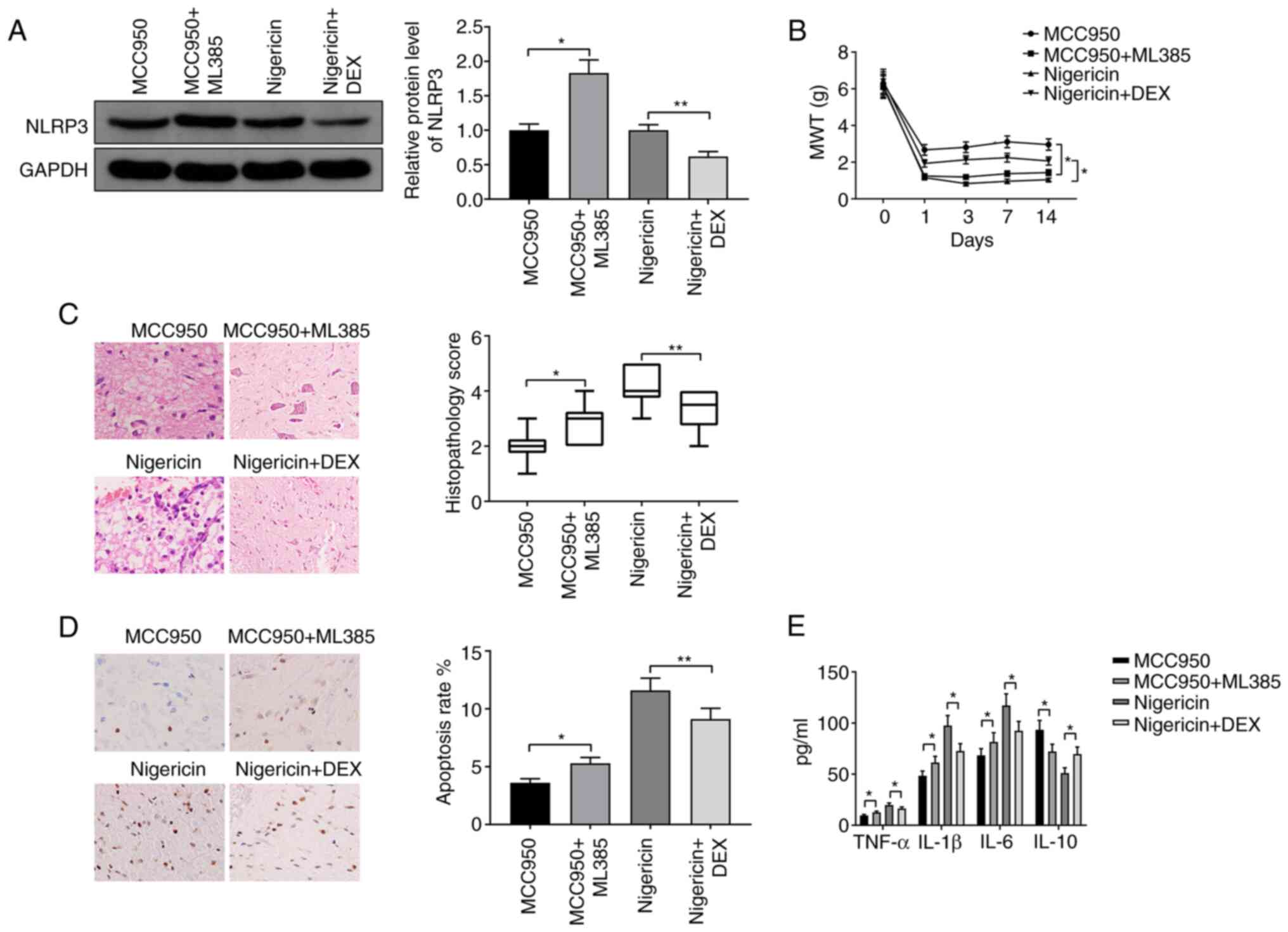
DEX alleviates neuropathic pain in CCI
rats by suppressing NLRP3 via the activation of Nrf2
To investigate the mechanism by which DEX alleviated
pain, CCI rats were injected with MCC950 + ML385 or nigericin +
DEX. NLRP3 was significantly upregulated in the MCC950 + ML385
group compared with the MCC950 group (Fig. 5A; P<0.05) and downregulated in
the nigericin + DEX group compared with the nigericin group
(Fig. 5A; P<0.01). MWT was
decreased in the MCC950 + ML385 group and increased in the
nigericin + DEX group compared with the MCC950 group and nigericin
group, respectively (Fig. 5B; both
P<0.05). Spinal cord injury and apoptosis rate were
significantly increased in the MCC950 + ML385 group, but were
suppressed in the nigericin + DEX group compared with the MCC950
group (P<0.05) and nigericin group (P<0.01), respectively
(Fig. 5C and D). Moreover, increases in the expression
levels of TNF-α, IL-1β and IL-6 and a decrease in the expression
level of IL-10 were observed in the MCC950 + ML385 group compared
with the MCC950 group, whereas inverse expression patterns of these
cytokines were detected in the nigericin + DEX group compared with
the nigericin group (Fig. 5E; all
P<0.05). Taken together, these results suggested that DEX
inhibited NLRP3 by activating Nrf2 to suppress neuropathic pain in
CCI rats.
Discussion
The diagnosis and treatment of neuropathic pain
remain challenging due to the diverse etiologies and clinical
manifestations of this disease (25). Over the past decades,
pharmacotherapy has been recommended for the management of
neuropathic pain, but only a minority of patients exhibit an
adequate response (26). The
present study aimed to discover the mechanism by which DEX relieved
neuropathic pain, and the results suggested that DEX inhibited
NLRP3 by stimulating Nrf2 to suppress neuropathic pain in a rat
model of CCI.
Firstly, the neuroprotective role of DEX in a rat
model of neuropathic pain was demonstrated. DEX reduced the
expression levels of TNF-α, IL-1β and IL-6 while increasing the
expression level of IL-10 in CCI rats. Moreover, downregulation of
NLRP3 and upregulation of Nrf2 were also detected in the spinal
cord. TNF-α, IL-1β and IL-6 have been identified as proinflammatory
factors, and IL-10 has been identified as an anti-inflammatory
factor in neuropathic pain (27).
DEX was revealed to attenuate neuropathic pain by promoting
anti-inflammatory activity in CCI (28). Consistent with the results of the
current study, Farghaly et al (29) demonstrated that DEX decreased TNF-α
and IL-6 to alleviate neuropathic pain.
In the present study, NLRP3 promoted neuropathic
pain and inflammatory responses in CCI rats. Despite the lack of
research on the interaction between DEX and NLRP3, the involvement
of NLRP3 in neuropathic pain has been intensively investigated. For
example, microRNA-223 has been indicated to ameliorate
morphine-induced analgesic tolerance to neuropathic pain by
downregulating NLRP3(30). Peptide
5 has been demonstrated to induce analgesia by inhibiting NLRP3 to
protect rats from peripheral nerve injury (31). Furthermore, paclitaxel has been
revealed to induce neuropathic pain by stimulating the NLRP3
inflammasome and the proinflammatory factor IL-1β (32).
The present study demonstrated that DEX relieved
neuropathic pain via the activation of Nrf2. CCI rats that were
treated with DEX and ML385 exhibited exacerbated neuropathic pain
and upregulated NLRP3 expression. A previous study showed that the
expression of Nrf2 was reduced in a rat model of chronic
neuropathic pain (33). Nrf2 may
alleviate oxaliplatin-induced peripheral neuropathy by maintaining
mitochondrial homeostasis and suppressing oxidative stress
(34). Moreover, rutin protects
against diabetic neuropathy by inhibiting plasma glucose levels and
neuroinflammation through the activation of Nrf2 signaling
(35). However, to the best of our
knowledge, the regulation of Nrf2 in DEX-induced neuroprotection is
largely unknown.
The NLRP3/Nrf2 pathway has been extensively
investigated in numerous pharmacological and pathological
processes. For example, mangiferin has been indicated to reduce
lipopolysaccharide- and D-galactosamine-induced liver injury by
inhibiting inflammation and oxidative stress via suppression of
NLRP3 and activation of Nrf2(36).
Dimethyl fumarate has been revealed to alleviate dextran sulfate
sodium-induced colitis by stimulating Nrf2-mediated suppression of
the NLRP3 inflammasome (37).
Sulforaphane has been demonstrated to protect acinar cells from
acute pancreatitis by regulating Nrf2-mediated oxidative stress and
the NLRP3 inflammasome (38). In
the present study, NLRP3 was upregulated in MCC950-treated CCI rats
in response to ML385 injection, indicating the suppressive effect
of Nrf2 on NLRP3 expression. Furthermore, DEX was indicated to
downregulate NLRP3 in nigericin-treated CCI rats, resulting in a
reduction in the MWT, as well as spinal cord injury and
inflammatory responses. Based on the aforementioned protective
effect of Nrf2, it was concluded that DEX relieved neuropathic pain
in CCI rats by suppressing NLRP3 via the activation of Nrf2.
Additionally, Nrf2 has been revealed to modulate HO-1 in dietary
oleuropein- and peracetylated oleuropein-mediated suppression of
murine lupus nephritis (39).
Further studies should be performed to explore whether a downstream
target of Nrf2 is involved in the management of neuropathic
pain.
The present study had several limitations that
should be mentioned. Firstly, only the mechanism underlying the
effect of DEX in animal models was examined, and this mechanism
should be further clarified in vitro and clinically. In the
present study, DEX relieved neuropathic pain by activating Nrf2 to
inhibit NLRP3-mediated neuroinflammation. Targeting the Nrf2/NLRP3
signaling pathway may be a novel intervention method for
neuropathic pain treatment. However, it has been demonstrated that
DEX exerts neuroprotective and analgesic effects via the
microRNA-101/E2F2/toll-like receptor 4/NF-κB axis (40). Whether this signaling pathway is
targeted in addition to the Nrf2/NLRP3 signaling pathway in the
context of DEX treatment requires further elucidation.
Additionally, the analgesic effect of DEX should be clinically
evaluated through preliminary clinical pharmacological and safety
analyses and pharmacokinetic tests.
In summary, DEX ameliorated neuropathic pain in a
rat model of CCI. The NLRP3/Nrf2 pathway was verified to modulate
inflammatory responses in neuropathic pain. DEX inhibited
neuropathic pain by activating Nrf2 through the suppression of
NLRP3. Understanding the mechanism of DEX may contribute to the
alleviation of neuropathic pain in patients and improve the
pharmacotherapeutic treatment of neuropathy.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WS and JL conceived the study design. WS and JL
designed the experiments. XL performed the experiments. YT analyzed
the data. JL and WS provided critical materials. WS and XL wrote
the manuscript. JL supervised the study. WS and JL confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Protection and Use Committee of Hunan Provincial People's Hospital
(Changsha, China) and strictly followed the ‘Guidelines for the Use
and Management of Laboratory Animals’ issued by the National
Institutes of Health and Animal Research: Reporting In Vivo
Experiments Guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bouhassira D and Attal N: Translational
neuropathic pain research: A clinical perspective. Neuroscience.
338:27–35. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Gierthmuhlen J and Baron R: Neuropathic
pain. Semin Neurol. 36:462–468. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Scholz J, Finnerup NB, Attal N, Aziz Q,
Baron R, Bennett MI, Benoliel R, Cohen M, Cruccu G, Davis KD, et
al: The IASP classification of chronic pain for ICD-11: Chronic
neuropathic pain. Pain. 160:53–59. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Colloca L, Ludman T, Bouhassira D, Baron
R, Dickenson AH, Yarnitsky D, Freeman R, Truini A, Attal N,
Finnerup NB, et al: Neuropathic pain. Nat Rev Dis Primers.
3(17002)2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Gilron I, Baron R and Jensen T:
Neuropathic pain: Principles of diagnosis and treatment. Mayo Clin
Proc. 90:532–545. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Weerink MAS, Struys MMRF, Hannivoort LN,
Barends CRM, Absalom AR and Colin P: Clinical pharmacokinetics and
pharmacodynamics of dexmedetomidine. Clin Pharmacokinet.
56:893–913. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Keating GM: Dexmedetomidine: A review of
its use for sedation in the intensive care setting. Drugs.
75:1119–1130. 2015.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jessen Lundorf L, Korvenius Nedergaard H
and Moller AM: Perioperative dexmedetomidine for acute pain after
abdominal surgery in adults. Cochrane Database Syst Rev.
2(CD010358)2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lin JP, Chen CQ, Huang LE, Li NN, Yang Y,
Zhu SM and Yao YX: Dexmedetomidine attenuates neuropathic pain by
inhibiting P2X7R expression and ERK phosphorylation in rats. Exp
Neurobiol. 27:267–276. 2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhao Y, He J, Yu N, Jia C and Wang S:
Mechanisms of dexmedetomidine in neuropathic pain. Front Neurosci.
14(330)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jo EK, Kim JK, Shin DM and Sasakawa C:
Molecular mechanisms regulating NLRP3 inflammasome activation. Cell
Mol Immunol. 13:148–159. 2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
He Y, Hara H and Nunez G: Mechanism and
regulation of NLRP3 inflammasome activation. Trends Biochem Sci.
41:1012–1021. 2016.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Coll RC, Robertson AA, Chae JJ, Higgins
SC, Munoz-Planillo R, Inserra MC, Vetter I, Dungan LS, Monks BG,
Stutz A, et al: A small-molecule inhibitor of the NLRP3
inflammasome for the treatment of inflammatory diseases. Nat Med.
21:248–255. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Grace PM, Strand KA, Galer EL, Urban DJ,
Wang X, Baratta MV, Fabisiak TJ, Anderson ND, Cheng K, Greene LI,
et al: Morphine paradoxically prolongs neuropathic pain in rats by
amplifying spinal NLRP3 inflammasome activation. Proc Natl Acad Sci
USA. 113:E3441–E3450. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ming T, Yuan M, Kong Q, Huang Q, Xia Z and
Wu X: Dexmedetomidine alleviates blunt chest trauma and hemorrhagic
shock-resuscitation-induced acute lung injury through inhibiting
the NLRP3 inflammasome. Mol Med Rep. 22:2507–2515. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yang T, Feng X, Zhao Y, Zhang H, Cui H,
Wei M, Yang H and Fan H: Dexmedetomidine enhances autophagy via
α2-AR/AMPK/mTOR pathway to inhibit the activation of NLRP3
inflammasome and subsequently alleviates lipopolysaccharide-induced
acute kidney injury. Front Pharmacol. 11(790)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Ahmed SM, Luo L, Namani A, Wang XJ and
Tang X: Nrf2 signaling pathway: Pivotal roles in inflammation.
Biochim Biophys Acta Mol Basis Dis. 1863:585–597. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Thimmulappa RK, Lee H, Rangasamy T, Reddy
SP, Yamamoto M, Kensler TW and Biswal S: Nrf2 is a critical
regulator of the innate immune response and survival during
experimental sepsis. J Clin Invest. 116:984–995. 2006.PubMed/NCBI View
Article : Google Scholar
|
|
19
|
Arruri V, Komirishetty P, Areti A,
Dungavath SK and Kumar A: Nrf2 and NF-κB modulation by Plumbagin
attenuates functional, behavioural and biochemical deficits in rat
model of neuropathic pain. Pharmacol Rep. 69:625–632.
2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Du L, Wang J, Chen Y, Li X, Wang L, Li Y,
Jin X, Gu X, Hao M, Zhu X, et al: Novel biphenyl diester derivative
AB-38b inhibits NLRP3 inflammasome through Nrf2 activation in
diabetic nephropathy. Cell Biol Toxicol. 36:243–260.
2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li F, Wang X, Zhang Z, Zhang X and Gao P:
Dexmedetomidine attenuates neuroinflammatory-induced apoptosis
after traumatic brain injury via Nrf2 signaling pathway. Ann Clin
Transl Neurol. 6:1825–1835. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kilkenny C, Browne W, Cuthill IC, Emerson
M and Altman DG: NC3Rs Reporting Guidelines Working Group. Animal
research: Reporting in vivo experiments: The ARRIVE guidelines. J
Gene Med. 12:561–563. 2010.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wiesenfeld-Hallin Z: Sex differences in
pain perception. Gend Med. 2:137–145. 2005.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cruccu G and Truini A: A review of
neuropathic pain: From guidelines to clinical practice. Pain Ther.
6 (Suppl 1):S35–S42. 2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Attal N and Bouhassira D: Pharmacotherapy
of neuropathic pain: Which drugs, which treatment algorithms? Pain.
156 (Suppl 1):S104–S114. 2015.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lees JG, Fivelman B, Duffy SS, Makker PG,
Perera CJ and Moalem-Taylor G: Cytokines in neuropathic pain and
associated depression. Mod Trends Pharmacopsychiatry. 30:51–66.
2015.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liang F, Liu M, Fu X, Zhou X, Chen P and
Han F: Dexmedetomidine attenuates neuropathic pain in chronic
constriction injury by suppressing NR2B, NF-κB, and iNOS
activation. Saudi Pharm J. 25:649–654. 2017.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Farghaly HS, Mahmoud AM and Abdel-Sater
KA: Effect of dexmedetomidine and cold stress in a rat model of
neuropathic pain: Role of interleukin-6 and tumor necrosis
factor-α. Eur J Pharmacol. 776:139–145. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xie XJ, Ma LG, Xi K, Fan DM, Li JG, Zhang
Q and Zhang W: Effects of microRNA-223 on morphine analgesic
tolerance by targeting NLRP3 in a rat model of neuropathic pain.
Mol Pain. 13(1744806917706582)2017.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Tonkin RS, Bowles C, Perera CJ, Keating
BA, Makker PG, Duffy SS, Lees JG, Tran C, Don AS, Fath T, et al:
Attenuation of mechanical pain hypersensitivity by treatment with
Peptide5, a connexin-43 mimetic peptide, involves inhibition of
NLRP3 inflammasome in nerve-injured mice. Exp Neurol. 300:1–12.
2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Jia M, Wu C, Gao F, Xiang H, Sun N, Peng
P, Li J, Yuan X, Li H, Meng X, et al: Activation of NLRP3
inflammasome in peripheral nerve contributes to paclitaxel-induced
neuropathic pain. Mol Pain. 13(1744806917719804)2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Li S, Yang C, Fang X, Zhan G, Huang N, Gao
J, Xu H, Hashimoto K and Luo A: Role of Keap1-Nrf2 signaling in
anhedonia symptoms in a rat model of chronic neuropathic pain:
Improvement with sulforaphane. Front Pharmacol.
9(887)2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Yang Y, Luo L, Cai X, Fang Y, Wang J, Chen
G, Yang J, Zhou Q, Sun X, Cheng X, et al: Nrf2 inhibits
oxaliplatin-induced peripheral neuropathy via protection of
mitochondrial function. Free Radic Biol Med. 120:13–24.
2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Tian R, Yang W, Xue Q, Gao L, Huo J, Ren D
and Chen X: Rutin ameliorates diabetic neuropathy by lowering
plasma glucose and decreasing oxidative stress via Nrf2 signaling
pathway in rats. Eur J Pharmacol. 771:84–92. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Pan CW, Pan ZZ, Hu JJ, Chen WL, Zhou GY,
Lin W, Jin LX and Xu CL: Mangiferin alleviates lipopolysaccharide
and D-galactosamine-induced acute liver injury by activating the
Nrf2 pathway and inhibiting NLRP3 inflammasome activation. Eur J
Pharmacol. 770:85–91. 2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Liu X, Zhou W, Zhang X, Lu P, Du Q, Tao L,
Ding Y, Wang Y and Hu R: Dimethyl fumarate ameliorates dextran
sulfate sodium-induced murine experimental colitis by activating
Nrf2 and suppressing NLRP3 inflammasome activation. Biochem
Pharmacol. 112:37–49. 2016.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Dong Z, Shang H, Chen YQ, Pan LL, Bhatia M
and Sun J: Sulforaphane protects pancreatic acinar cell injury by
modulating Nrf2-Mediated oxidative stress and NLRP3 inflammatory
pathway. Oxid Med Cell Longev. 2016(7864150)2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Castejon ML, Sanchez-Hidalgo M,
Aparicio-Soto M, Montoya T, Martin-LaCave I, Fernandez-Bolanos JG
and Alarcon-de-la-Lastra C: Dietary oleuropein and its new
acyl-derivate attenuate murine lupus nephritis through HO-1/Nrf2
activation and suppressing JAK/STAT, NF-κB, MAPK and NLRP3
inflammasome signaling pathways. J Nutr Biochem.
74(108229)2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhang W, Yu T, Cui X, Yu H and Li X:
Analgesic effect of dexmedetomidine in rats after chronic
constriction injury by mediating microRNA-101 expression and the
E2F2-TLR4-NF-κB axis. Exp Physiol. 105:1588–1597. 2020.PubMed/NCBI View Article : Google Scholar
|