Introduction
Oxidative stress is caused by an imbalance between
the production of reactive oxygen species (ROS) and the antioxidant
capacity of cellular antioxidants in biological systems (1). Excessive oxidative stress is related
to the pathogenesis of various diseases, including
neurodegenerative diseases, tumors and inflammation (2,3).
Previous studies have demonstrated that increased levels of ROS
production lead to the development of several chronic intestinal
inflammatory diseases (4-6).
In particular, increased ROS production is typically associated
with the pathogenesis of inflammatory bowel disease, which is
characterized by chronic inflammation in human gastrointestinal
disease (7,8). Therefore, inhibiting oxidative
stress-induced injury may serve as an important therapeutic
strategy.
Shikonin, a natural naphthoquinone extracted from
the roots of the traditional Chinese medicine Lithospermumery
throrhizon, possesses multiple pharmacological properties,
including antioxidant, anti-inflammatory, antiviral, enhanced
immunity, antifertility and antitumor effects (9-13).
Numerous studies have demonstrated that shikonin displayed
efficient antioxidative activities against various types of ROS
(14,15). Guo et al (2) reported that shikonin attenuated
acetaminophen-induced acute liver injury via inhibition of
oxidative stress. Several studies indicated that shikonin displayed
significant protective effects in brain and hepatic
ischemia/reperfusion injury by reducing ROS (16-19).
However, the potential antioxidant mechanism underlying shikonin
activity is not completely understood.
The present study aimed to investigate the effects
of shikonin against H2O2-induced oxidative
stress injury in human intestinal epithelial cells and to explore
the underlying molecular mechanism. In many studies, human colon
cancer cells were used as oxidative damage models (20,21).
Therefore, HT29 human colon cancer cells were selected to construct
oxidative damage models in the present study.
Materials and methods
Cell culture and treatment
HT29 cells were purchased from American Type Culture
Collection. The cell line was established at the Memorial Sloan
Kettering Cancer Center and was authenticated using STR profiling.
Cells were cultured in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin-streptomycin (Gibco; Thermo
Fisher Scientific, Inc.) in a humidified atmosphere with 5%
CO2 at 37˚C. Cells (5x104-105
cells/ml) were harvested and used for subsequent experiments. Cells
were divided into six groups for treatment: A control group, cells
cultured in medium without H2O2 and shikonin;
DMSO group, cells cultured in medium with 0.1% DMSO; and
H2O2 group, cells cultured in medium with 800
µM H2O2 (Sigma-Aldrich; Merck KGaA) all of
which were cultured for 24 h at 37˚C; and 3 shikonin groups, cells
pretreated with 2.5, 5 or 10 µg/ml shikonin (Shanghai Yuan Ye
Biotechnology Co., Ltd.; purity ≥98%) for 6 h at 37˚C, as
previously described (22,23), and then co-treated with 800 µM
H2O2 for 24 h at 37˚C.
MTT assay
For the MTT assay, HT29 cells (5x103)
were seeded in 96-well plates and cultured for 24 h at 37˚C. cells
were treated with H2O2 (25-1,600 µM) for 4,
8, 12 or 24 h at 37˚C, or treated with shikonin (2.5, 5, 10, 25,
50, 100 and 200 µg/ml) for 24 h at 37˚C. Subsequently, cells were
incubated with 0.5 mg/ml MTT (Sigma-Aldrich; Merck K Ga A) for an
additional 4 h at 37˚C. The supernatant was discarded and 100 µl
DMSO was added to each well to dissolve the formazan crystals.
Absorbance was measured at a wavelength of 490 nm using a
microplate reader. The results are presented as a percentage of the
control.
Early and late apoptosis detection
assay
Early and late apoptosis was measured using a
FITC-conjugated annexin V and propidium iodide kit (BD
Biosciences). Cells (1x106) were trypsinized, washed
with PBS and resuspended in 1Xbinding buffer. Subsequently, 5 µl
FITC-conjugated annexin V and 5 µl propidium iodide were added to
100 µl cell suspension. Following incubation for 15 min at room
temperature in the dark, apoptosis was analyzed via flow cytometry
(EXPO32 ADC; Epics XL-MCL; Beckman Coulter, Inc.).
Cell cycle assay
Cells (1x106) were trypsinized with 0.25%
trypsin-EDTA at room temperature for 24 h and washed three times in
PBS. Cell cycle distribution was detected using a Cycle test Plus
DNA reagent kit (BD Biosciences). The percentage of cells in each
cell cycle phase (G0/G1, S and
G2/M) was calculated via flow cytometry (Epics XL-MCL;
Beckman Coulter, Inc.). The software used was the inbuilt software
provided with the flow cytometer. Proliferation index [PI;PI (%)=S
phase (%) + G2/M phase (%)].
ROS measurement
Intracellular ROS levels were measured using a ROS
assay kit (cat. no. S0033M; Beyotime Institute of Biotechnology)
according to the manufacturer's instructions. Briefly, cells
(1x106) were collected and incubated for 20 min in 500
µl 2',7'-dichlorofluorescin diacetate fluorescence (DCFH-DA)
solution (10 µM) at 37˚C in the dark. Following washing with PBS,
cells were resuspended in 500 µl PBS and analyzed via flow
cytometry (EXPO32 ADC, Epics XL-MCL; Beckman Coulter, Inc.).
Levels of malondialdehyde (MDA) and
superoxide dismutase (SOD) assays
Cells (1x106) were collected and
centrifuged at 10,000 x g for 10 min at 4˚C. The levels of SOD and
MDA in the supernatant were measured using SOD (cat. no. S0101S)
and MDA (cat. no. S0131S) assay kits (both Beyotime Institute of
Biotechnology), respectively, according to the manufacturer's
protocol.
Lactate dehydrogenase (LDH) activity
assay
Cell membrane integrity was determined using an LDH
assay. LDH levels in the cell medium from treated cells were
determined using an LDH assay kit (cat. no. C0016; Beyotime
Institute of Biotechnology) according to the manufacturer's
protocol. Absorbance was measured at a wavelength of 490 nm using a
microplate reader. LDH levels were calculated according to the
following formula: LDH (%)=(sample-blank)/(control-blank) x100.
Mitochondrial membrane potential
assay
To assess mitochondrial integrity, the mitochondrial
membrane potential assay kit with JC-1 (cat. no. C2006; Beyotime
Institute of Biotechnology) was used. Cells (1x106) were
resuspended in 500 µl medium, followed by addition of 500 µl JC-1
dye for 20 min at 37˚C. Cells were rinsed twice with JC-1 dye
buffer. The fluorescent signal in cells was calculated by
performing flow cytometry (EXPO32 ADC; Epics XL-MCL; Beckman
Coulter, Inc.).
Caspase-3 and caspase-9 activity
assays
Cells (1x106) were digested with trypsin
and harvested by centrifugation at 1,000 x g for 5 min at 4˚C.
Caspase-3 and caspase-9 activities were measured using caspase-3
(cat. no. BC3830) and caspase-9 (cat. no. BC3890) activity
detection kits (Beijing Solarbio Science & Technology Co.,
Ltd.) according to the manufacturer's protocol. Caspase-3 and
caspase-9 activities are presented as U/mg protein.
Western blotting
Total protein was extracted from cells using RIPA
buffer (Beyotime Institute of Biotechnology) containing 1 mM PMSF
(Beyotime Institute of Biotechnology) and phosphatase inhibitor for
30 min on ice. Total protein was quantified using a bicinchoninic
acid protein assay kit (Beyotime Institute of Biotechnology). Equal
amounts of protein (20 µg) were separated via 10% SDS-PAGE and
transferred to 0.45 µm PVDF membranes. After 5% nonfat milk
blocking at room temperature for 2 h, the membranes were incubated
with primary antibodies targeted against: Cytochrome c (cat.
no. 4280; Cell Signaling Technology, Inc.; 1:1,000), Bax (cat. no.
5023; Cell Signaling Technology, Inc.; 1:1,000), Bcl-2 (cat. no.
3498; Cell Signaling Technology, Inc.; 1:1,000) and β-actin (cat.
no. 4970; Cell Signaling Technology, Inc.; 1:1,000) at 4˚C
overnight. Following primary incubation, the membranes were
incubated with a horseradish peroxidase-conjugated polymer-tagged
secondary antibody (cat. no. 7074; Cell Signaling Technology, Inc.;
1:5,000) for 2 h at room temperature. Protein bands were visualized
using ECL reagent (Thermo Fisher Scientific, Inc.) and protein
expression was semi-quantified using Image J software (National
Institutes of Health, version 1.8.0) with β-actin as the loading
control.
Statistical analysis
Data are presented as the mean ± SD. All experiments
were performed in triplicate. Statistical analyses were performed
using SPSS software (version 19.0; IBM Corp.). Comparisons among
multiple groups were analyzed using the one-way analysis of
variance (ANOVA) test and Dunnett's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Shikonin attenuates
H2O2-induced decreases in HT29 cell
viability
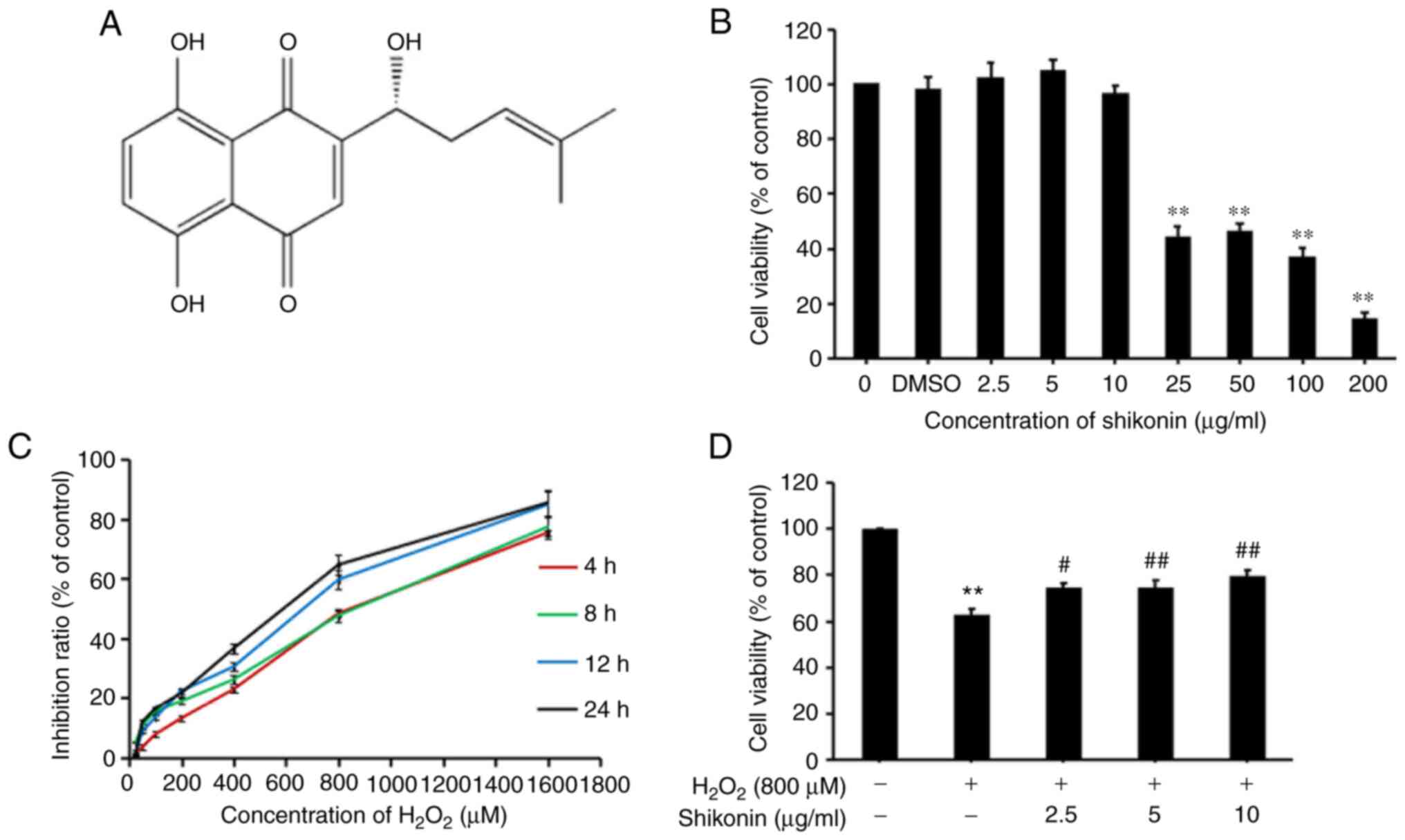
To identify suitable H2O2 and
shikonin concentrations and durations of action, an MTT assay was
performed to evaluate cell cytotoxicity. The results indicated that
H2O2 caused marked cytotoxicity in HT29 cells
at 800 µM compared with 0 µM, and when cells treated with 800 µM
H2O2, the cell inhibition rates at 4, 8, 12
and 24 h were 22.91, 26.12, 30.30 and 36.64%, respectively
(Fig. 1C). Therefore, cells treated
with 800 µM H2O2 for 24 h were selected for
subsequent experiments. Subsequently, the cytotoxic effect of
shikonin at different concentrations was investigated by performing
an MTT assay. Compared with the 0 µg/ml shikonin, no significant
cytotoxic effect was observed in cells treated with 2.5, 5 and 10
µg/ml shikonin alone. Therefore, three concentrations of shikonin
were used in subsequent experiments (Fig. 1B). Finally, to assess the effects of
shikonin against H2O2-induced cytotoxicity,
HT29 cells were pretreated with different concentrations of
shikonin (2.5, 5 and 10 µg/ml) for 6 h, and then co-treated with
800 µM H2O2 for 24 h. The results indicated
that the cell viability of the shikonin group gradually increased
compared with the H2O2 group in a
dose-dependent manner, suggesting that shikonin reversed
H2O2-induced decreases in cell viability
(Fig. 1D).
Shikonin attenuates
H2O2-induced HT29 cell apoptosis
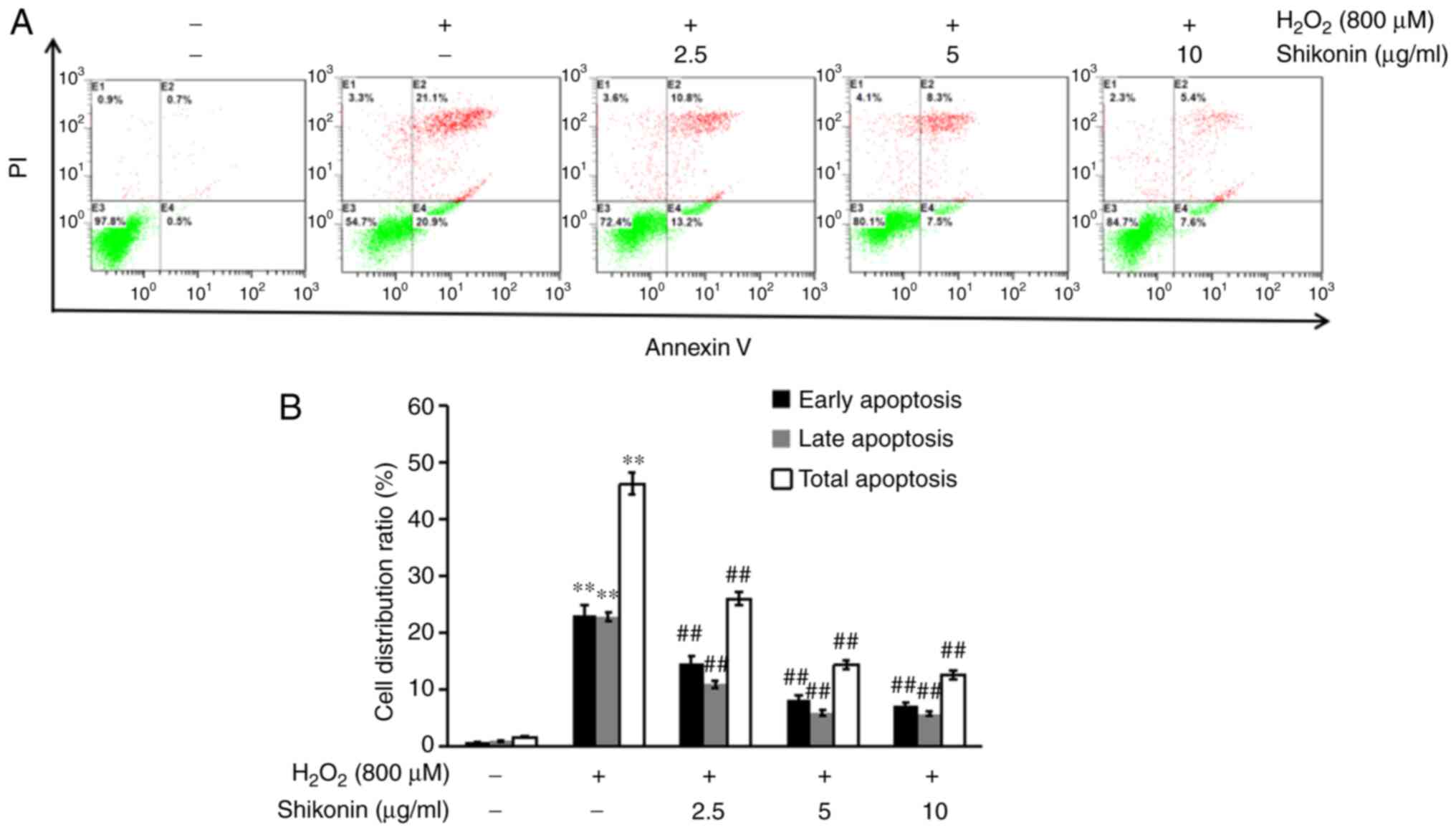
To assess the protective effects of shikonin against
H2O2-induced cell apoptosis, flow cytometry
was performed to detect apoptosis following Annexin V and propidium
iodide staining (Fig. 2). Compared
with the control group, the percentage of total apoptotic cells was
significantly increased in the H2O2 group. By
contrast, the shikonin groups displayed a significantly decreased
percentage of apoptotic cells compared with the
H2O2 group, indicating that shikonin
inhibited H2O2-induced cell apoptosis.
Shikonin suppresses
H2O2-induced G0/G1 cell
cycle arrest in HT29 cells
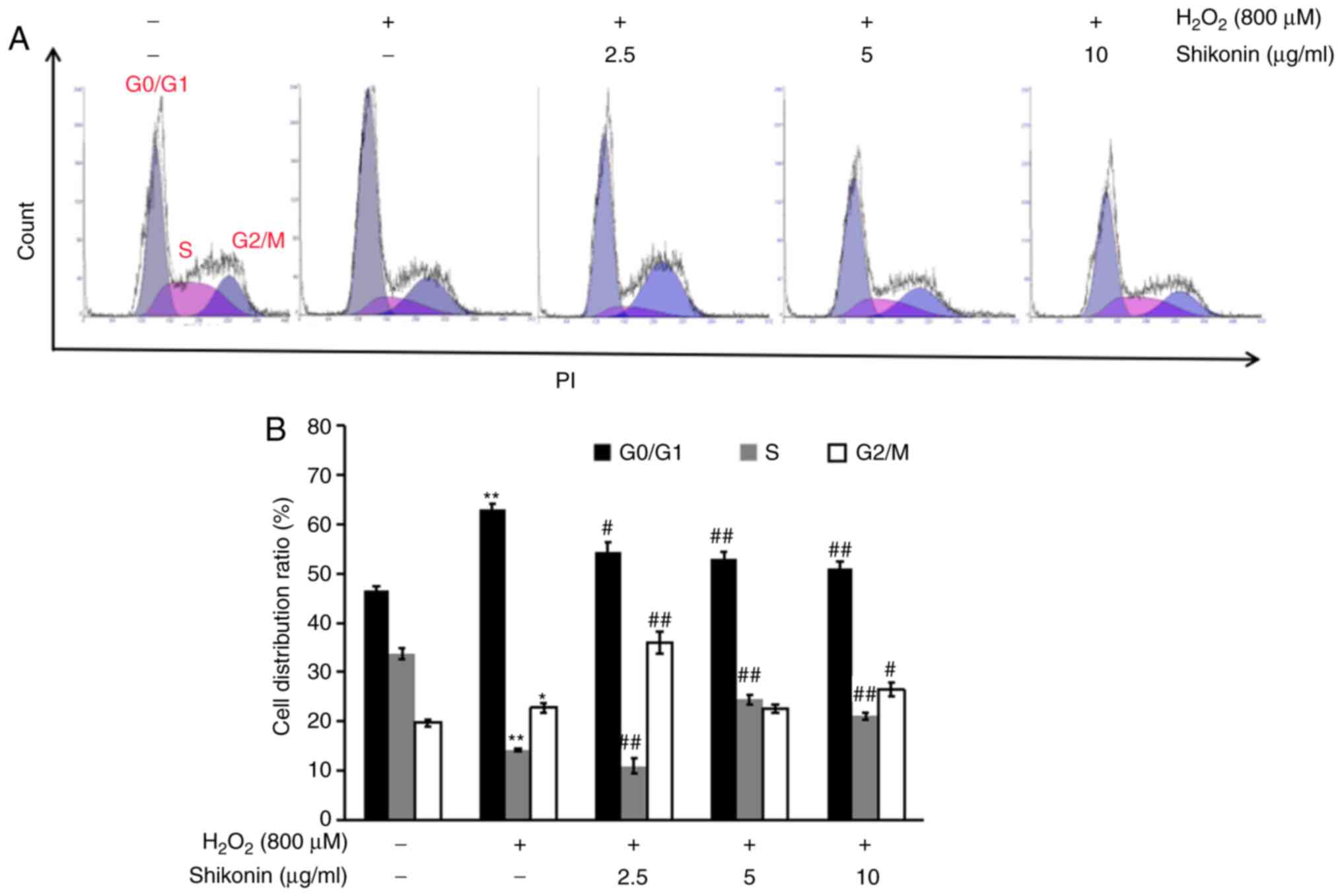
To investigate the mechanism underlying the effects
of shikonin on H2O2-mediated inhibition of
cell proliferation, the effects of shikonin on regulating the cell
cycle in H2O2-treated HT29 cells were
evaluated by conducting flow cytometry. Compared with the control
group, an increased percentage of G0/G1 cells
and a decreased proliferation index (PI) [PI;PI (%)=S phase (%) +
G2/M phase (%)] were observed in the
H2O2 group (Fig.
3). The PI gradually increased in a dose-dependent manner in
the shikonin groups compared with the H2O2
group. The results suggested that shikonin suppressed
H2O2-induced G0/G1 cell
cycle arrest.
Shikonin inhibits
H2O2-induced increases in ROS and MDA levels
in HT29 cells
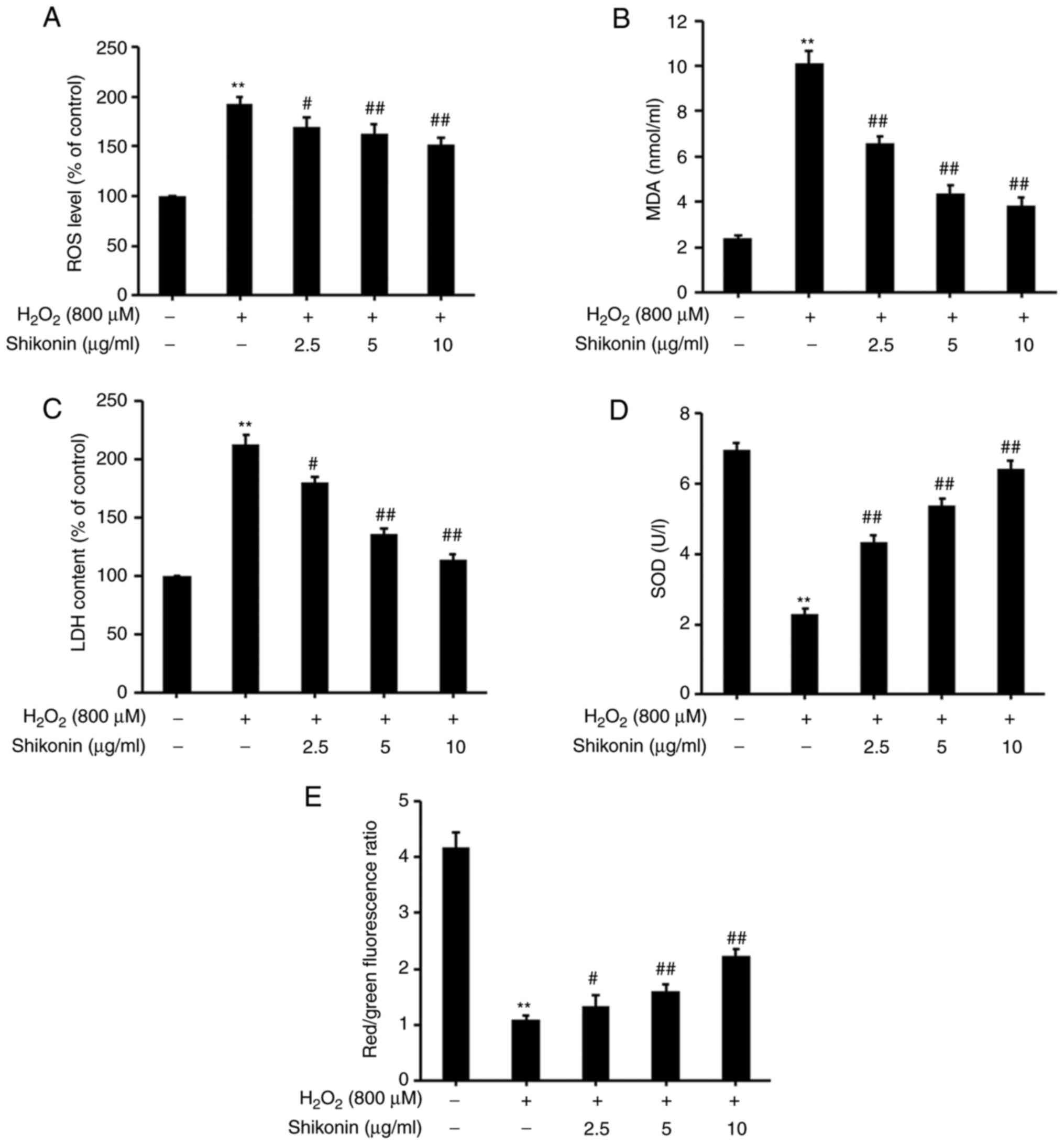
The levels of intracellular ROS as a major oxidant
were assessed by conducting a DCFH-DA assay (Fig. 4A). Following exposure to
H2O2, the levels of intracellular ROS were
significantly increased compared with the control group. However,
significantly lower levels of intracellular ROS were observed in
the shikonin groups compared with the H2O2
group.
MDA is a biomarker of oxidative stress (24), and MDA activity was measured using
an MDA assay kit (Fig. 4B). Upon
H2O2 exposure, MDA levels were significantly
increased compared with the control group. MDA levels in the
shikonin groups gradually decreased in a dose-dependent manner
compared with the H2O2 group. The results
suggested that shikonin inhibited
H2O2-induced intracellular ROS and MDA
accumulation.
Shikonin prevents
H2O2-induced LDH release in HT29 cells
LDH, a glycolytic enzyme, has been suggested to be a
key indicator of cell membrane integrity (12). The effects of shikonin on
extracellular LDH levels were measured using an LDH assay kit
(Fig. 4C). LDH levels were
significantly increased in H2O2-treated cells
compared with the control group. LDH levels in the shikonin groups
were significantly lower compared with the
H2O2 group. The results indicated that
shikonin prevented H2O2-induced LDH
release.
Shikonin inhibits
H2O2-induced decreases in SOD levels in HT29
cells
SOD is an important antioxidant enzyme in the
mitochondria and serves as a defense against oxidative stress
(25). SOD activity was measured
using a SOD assay kit (Fig. 4D).
SOD levels were significantly decreased in the
H2O2 group compared with the control group.
Compared with the H2O2 group, SOD levels
gradually increased with increasing shikonin concentrations,
suggesting that shikonin prevented
H2O2-mediated decreases in SOD levels.
Shikonin reverses
H2O2-induced decreases in mitochondrial
membrane potential in HT29 cells
Mitochondrial dysfunction has been reported to be
associated with H2O2-inducedcell apoptosis
(26). To further investigate
whether shikonin was associated with
H2O2-induced mitochondrial dysfunction, the
mitochondrial membrane potential in HT29 cells was investigated by
performing a JC-1 dye assay (Fig.
4E). Mitochondrial membrane potential levels were significantly
decreased in HT29 cells exposed to H2O2
compared with the control group. However, a gradual increase in
mitochondrial membrane potential levels was observed in cells
pretreated with 2.5, 5 or 10 µg/ml shikonin compared with the
H2O2 group. Therefore, the results indicated
that shikonin reversed H2O2-mediated
decreases in mitochondrial membrane potential.
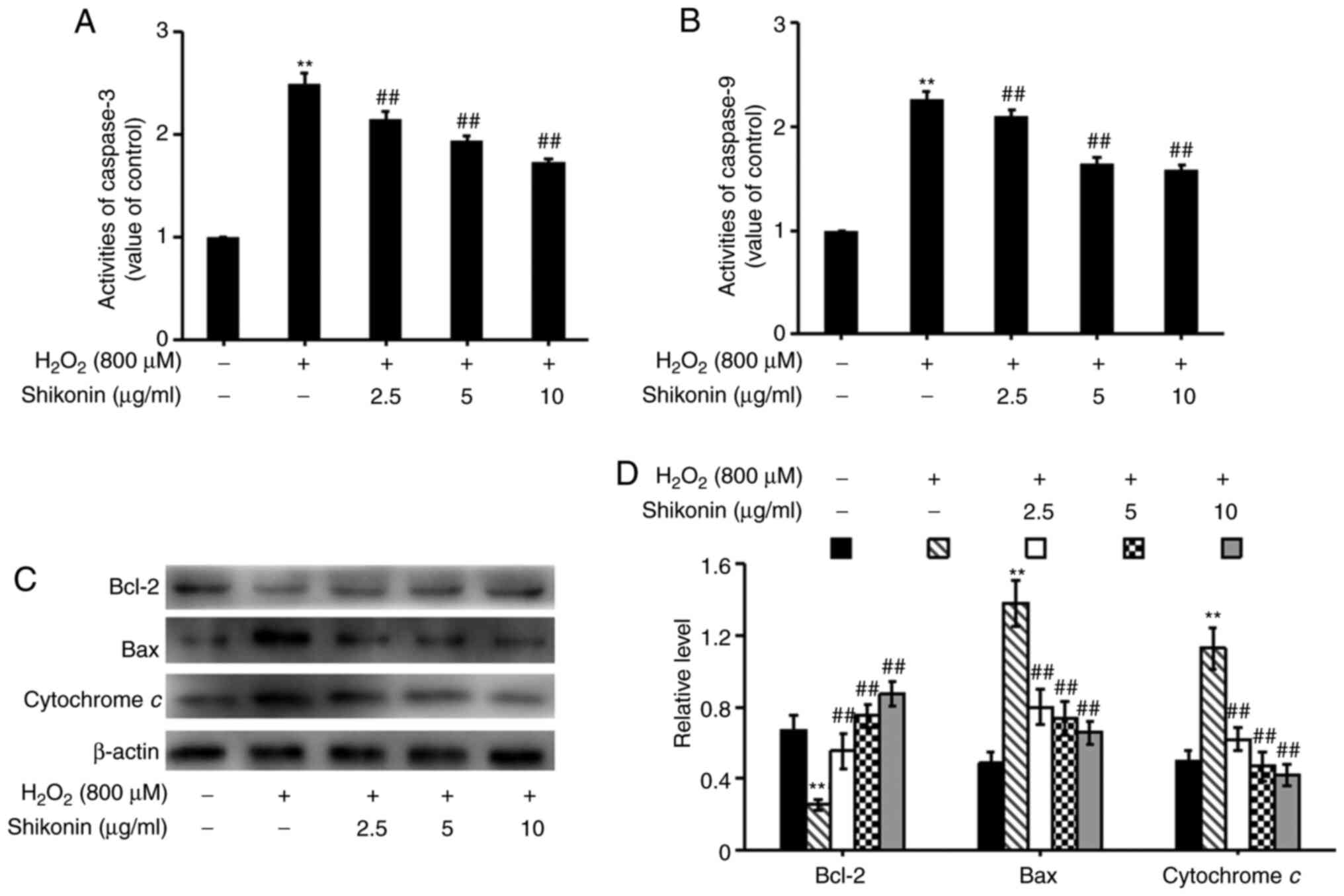
Shikonin protects HT29 cells against
oxidative stress via inhibition of the mitochondrial apoptosis
pathway
To assess whether the mitochondrial apoptosis
pathway was involved in promoting the effects of shikonin on cell
apoptosis, caspase-3 and caspase-9 levels were measured by
performing caspase assays (Fig. 5A
and B), and the expression levels
of Bcl-2, Bax and cytochrome c were determined via western
blotting (Fig. 5C and D). Following exposure to
H2O2, Bcl-2 expression levels were
significantly decreased, whereas the activity levels of caspase-3
and caspase-9, as well as the expression levels of Bax and
cytochrome c were significantly increased compared with the
control group. The opposite effects were observed in
H2O2-treated cells pretreated with shikonin
(2.5, 5 and 10 µg/ml). Therefore, the results indicated that
shikonin protected HT29 cells against oxidative stress via
inhibiting the mitochondrial apoptosis pathway in a
concentration-dependent manner.
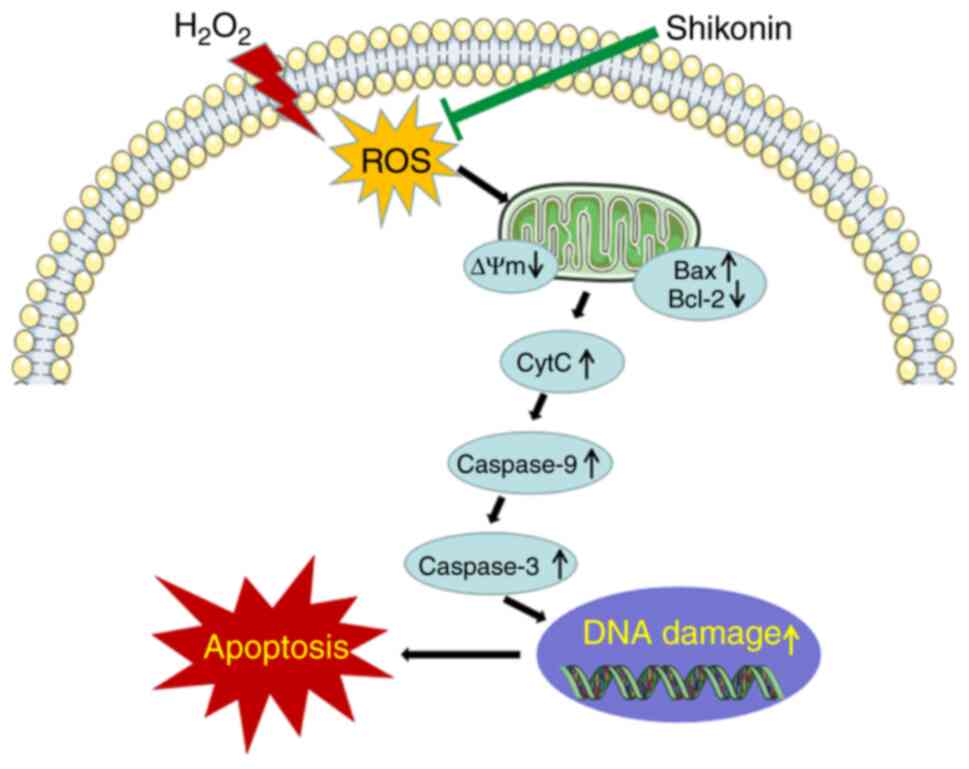
The possible mechanism of
shikonin
The potential cytoprotective mechanism underlying
the effects of shikonin against H2O2-induced
oxidative injury via elimination of ROS, attenuation of DNA damage
and inhibition of mitochondria mediated apoptosis (Fig. 6).
Discussion
The present study investigated the protective
effects of shikonin on H2O2-induced oxidative
stress in HT29 cells and explored the mechanism underlying the
antioxidative effects of shikonin. Shikonin protected against
H2O2-induced injury in HT29 cells. Consistent
with previous reports, the results of the MTT assay indicated that
H2O2 displayed cytotoxicity in HT29 cells in
a concentration-dependent manner (23,27).
Moreover, the flow cytometry results demonstrated that
H2O2 decreased the PI and increased apoptosis
in HT29 cells compared with the control group.
H2O2-mediated decreases in HT29 cell
viability were significantly reversed following pretreatment with
shikonin. In addition, shikonin also increased the PI and
attenuated apoptosis in H2O2-treated HT29
cells. The results suggested that shikonin exerted a protective
effect against H2O2-induced oxidative stress
in HT29 cells.
ROS is an important antioxidant enzyme in the
mitochondria, which has been demonstrated to serve a critical role
in DNA oxidative damage and is also a major oxidant in vivo
(28-32).
Under normal physiological conditions, SOD and other antioxidants
can scavenge ROS, resulting in a dynamic equilibrium between the
generation and removal of ROS (33,34).
MDA is a biomarker of oxidative stress that reflects ROS-induced
membrane lipid peroxidation (35,36).
As cells are damaged, high amounts of ROS accumulate and induce
lipid peroxidation on the membrane to produce MDA. MDA can damage
the membrane structure, eventually leading to enhanced membrane
permeability, increased generation of intracellular enzymes and the
release of LDH (37). The results
of the present study indicated that shikonin significantly
decreased the levels of intracellular ROS and LDH, decreased MDA
levels and restored SOD activity in
H2O2-treated cells. Collectively, the results
indicated that shikonin reduced oxidative stress, at least in part
via its antioxidant activity and ROS elimination.
Mitochondria are the major physiological sources of
ROS, and H2O2 may increase oxidative damage
by inducing mitochondrial dysfunction, resulting in increased ROS
production and induction of mitochondrial membrane potential loss
(38,39). In the present study, shikonin
significantly inhibited H2O2-induced
mitochondrial membrane potential loss, indicating that shikonin may
display a protective effect against
H2O2-induced oxidative damage via the
mitochondrial pathway.
The mitochondrial, death receptor and endoplasmic
reticulum signaling pathways are three major apoptosis signaling
pathways that are dependent on caspases (40). In the mitochondria-mediated
apoptosis pathway, oxidative stress induces the opening of
mitochondrial permeability transition pores. Mitochondria release
the apoptotic promoter and cytochrome c, which activates the
caspase cascade and induces apoptosis (40). The Bcl-2 family, including
antiapoptotic regulator Bcl-2, proapoptotic regulator Bax and death
proteins, is a major regulator of the mitochondrial apoptotic
pathway (41). The antiapoptotic
mechanism underlying Bcl-2 is direct antioxidation, whereas Bax is
the primary mediator of the mitochondrial apoptosis pathway
(42). Activated Bax leads to the
release of cytochrome c and mediates apoptosis induced by
the mitochondrial pathway (43). It
has been reported that Bcl-2 upregulation and Bax downregulation
can alleviate the occurrence of apoptosis (44). The results of animal experiments
also demonstrated that ischemia-reperfusion injury and heart
failure can cause Bcl-2 downregulation and increase apoptosis,
whereas Bcl-2 expression in myocardial cells is upregulated
following ischemic preconditioning treatment (45). Consistent with previous reports, the
present study also suggested that shikonin upregulated Bcl-2
expression and downregulated Bax expression in
H2O2-induced HT29 cells.
Caspases are a family of proteases that serve
important roles in the process of apoptosis (46). Caspase-9 is upstream of the
apoptotic signal transduction process and activates
caspase-3(40). Caspase-3-mediated
protein cleavage is an important component of the molecular
mechanism underlying apoptosis. In addition, caspase-3 serves a key
role in nuclear apoptosis, including chromatin condensation and DNA
fragmentation (40,47). The present study examined the
activities of caspase-3 and caspase-9, and the results indicated
that the shikonin group displayed significant downregulation of
caspase-3 and caspase-9 activities compared with the
H2O2 group, suggesting that shikonin
protected against H2O2-induced oxidative
damage of HT29 cells from mitochondrial machinery mediated by the
apoptotic pathway.
In conclusion, the present study indicated that the
protective effects of shikonin against
H2O2-induced oxidative stress injury were
activated at least in part via removing ROS, ameliorating
mitochondrial dysfunction, attenuating DNA oxidative damage and
inhibiting mitochondrial pathway-mediated apoptosis. The results
suggested a potential mechanism underlying the antioxidant role of
shikonin and a new perspective for the rational use of shikonin for
the treatment of oxidation damage-associated diseases in the
future.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 21375029), the Medical
Research Foundation of Guangdong Province (grant no. B2017044), the
Research Foundation of Guangdong Medical University (grant no.
M2016021) and the Traditional Chinese Medicine Research Foundation
of Guangdong Provincial Bureau (grant no. 20182072).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
YZ and AQ designed the study, acquired the data and
drafted the manuscript. QH performed the statistical analysis and
drafted the manuscript. JZ and LL performed the statistical
analysis. JZ revised the manuscript for intellectual content. KC
acquired the data. CC designed the study and drafted the
manuscript. All authors have read and approved the final
manuscript. CC and YZ confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sinha N and Dabla PK: Oxidative stress and
antioxidants in hypertension-a current review. Curr Hypertens Rev.
11:132–142. 2015.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Guo H, Sun J, Li D, Hu Y, Yu X, Hua H,
Jing X, Chen F, Jia Z and Xu J: Shikonin attenuates
acetaminophen-induced acute liver injury via inhibition of
oxidative stress and inflammation. Biomed Pharmacother.
112(108704)2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Pistollato F, Iglesias RC, Ruiz R,
Aparicio S, Crespo J, Lopez LD, Manna PP, Giampieri F and Battino
M: Nutritional patterns associated with the maintenance of
neurocognitive functions and the risk of dementia and Alzheimer's
disease: A focus on human studies. Pharmacol Res. 131:32–43.
2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Yan H, Wang H, Zhang X, Li X and Yu J:
Ascorbic acid ameliorates oxidative stress and inflammation in
dextran sulfate sodium-induced ulcerative colitis in mice. Int J
Clin Exp Med. 8:20245–20253. 2015.PubMed/NCBI
|
|
5
|
Mrowicka M, Mrowicki J, Mik M, Wojtczak R,
Dziki L, Dziki A and Majsterek I: Association between SOD1, CAT,
GSHPX1 polymorphisms and the risk of inflammatory bowel disease in
the Polish population. Oncotarget. 8:109332–109339. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Shalkami AS, HAssan M and Bakr AG:
Anti-inflammatory, antioxidant and anti-apoptotic activity of
diosmin in acetic acid-induced ulcerative colitis. Hum Exp Toxicol.
37:78–86. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kruidenier L and Verspaget HW: Review
article: Oxidative stress as a pathogenic factor in inflammatory
bowel disease-radicals or ridiculous? Aliment Pharmacol Ther.
16:1997–2015. 2002.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zhu H and Li YR: Oxidative stress and
redox signaling mechanisms of inflammatory bowel disease: Updated
experimental and clinical evidence. Exp Biol Med (Maywood).
237:474–480. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Sakthivel KM and Guruvayoorappan C:
Amentoflavone inhibits iNOS, COX-2 expression and modulates
cytokine profile, NF-κB signal transduction pathways in rats with
ulcerative colitis. Int Immunopharmacol. 17:907–916.
2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhong Y, Yu W, Feng J, Fan Z and Li J:
Curcumin suppresses tumor necrosis factor-alpha-induced matrix
metalloproteinase-2 expression and activity in rat vascular smooth
muscle cells via the NF-κB pathway. Exp Ther Med. 7:1653–1658.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
An S, Park YD, Paik YK, Jeong TS and Lee
WS: Human ACAT inhibitory effects of shikonin derivatives from
Lithospermum erythrorhizon. Bioorg Med Chem Lett. 17:1112–1116.
2007.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang Z, Yang L, Wang B, Zhang L, Zhang Q,
Li D, Zhang S, Gao H and Wang X: Protective role of liriodendrin in
mice with dextran sulphate sodium-induced ulcerative colitis. Int
Immunopharmacol. 52:203–210. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Liang W, Cai A, Chen G, Xi H, Wu X, Cui J,
Zhang K, Zhao X, Yu J, Wei B and Chen L: Shikonin induces
mitochondria-mediated apoptosis and enhances chemotherapeutic
sensitivity of gastric cancer through reactive oxygen species. Sci
Rep. 6(38267)2016.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Gao D, Kakuma M, Oka S, Sugino K and
Sakurai H: Reaction of beta-alkannin (shikonin) with reactive
oxygen species: Detection of beta-alkannin free radicals. Bioorg
Med Chem. 8:2561–2569. 2000.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Tong Y, Bai L, Gong R, Chuan J, Duan X and
Zhu Y: Shikonin protects PC12 cells against β-amyloid
peptide-induced cell injury through antioxidant and antiapoptotic
activities. Sci Rep. 8(26)2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Esmaeilzadeh E, gardaneh M, Gharib E and
Sabouni F: Shikonin protects dopaminergic cell line PC12 against
6-hydroxydopamine-mediated neurotoxicity via both
glutathione-dependent and independent pathways and by inhibiting
apoptosis. Neurochem Res. 38:1590–1604. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tong Y, Chuan J, Bai L, Shi J, Zhong L,
Duan X and Zhu Y: The protective effect of shikonin on renal
tubular epithelial cell injury induced by high glucose. Biomed
Pharmacother. 98:701–708. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liu T, Zhang Q, Mo W, Yu Q, Xu S, Li J, Li
S, Feng J, Wu L, Lu X, et al: The protective effects of shikonin on
hepatic ischemia/reperfusion injury are mediated by the activation
of the PI3K/Akt pathway. Sci Rep. 7(44785)2017.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang Z, Liu T, Gan L, Wang T, Yuan X,
Zhang B, Chen H and Zheng Q: Shikonin protects mouse brain against
cerebral ischemia/reperfusion injury through its antioxidant
activity. Eur J Pharmacol. 643:211–217. 2010.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Moore LD, Le T and Fan G: DNA methylation
and its basic function. Neuropsychopharmacology. 38:23–38.
2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Sena P, Mancini S, Benincasa M, Mariani F,
Palumbo C and Roncucci L: Metformin induces apoptosis and alters
cellular responses to oxidative stress in Ht29 colon cancer cells:
Preliminary findings. Int J Mol Sci. 19(1478)2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Bai J, Yu J, Wang J, Xue B, He N, Tian Y,
Yang L, Wang Y, Wang Y and Tang Q: DNA methylation of miR-122
aggravates oxidative stress in colitis targeting SELENBP1 partially
by p65NF-κ B signaling. Oxid Med Cell Longev.
2019(5294105)2019.
|
|
23
|
Zhao X, Fang J, Li S, Gaur U, Xing X, Wang
H and Zheng W: Artemisinin attenuated hydrogen peroxide
(H2O2)-induced oxidative injury in SH-SY5Y and hippocampal neurons
via the activation of AMPK pathway. Int J Mol Sci.
20(2680)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Akbay E, Arbag H, Uyar Y and Ozturk K:
Oxidative stress and antioxidant factors in pathophysiology of
allergic rhinitis. Kulak Burun Bogaz Ihtis Derg. 17:189–1896.
2007.PubMed/NCBI(In Turkish).
|
|
25
|
Bresciani G, DA CI and González-Gallego J:
Manganese superoxide dismutase and oxidative stress modulation. Adv
Clin Chem. 68:87–130. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Jablonski RP, Kim SJ, Cheresh P, Williams
DB, Morales-Nebreda L, Cheng Y, Yeldandi A, Bhorade S, Pardo A,
Selman M, et al: SIRT3 deficiency promotes lung fibrosis by
augmenting alveolar epithelial cell mitochondrial DNA damage and
apoptosis. FASEB J. 31:2520–2532. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chen XH, Zhou X, Yang XY, Zhou ZB, Lu DH,
Tang Y, Ling ZM, Zhou LH and Feng X: Propofol protects against
H2O2-induced oxidative injury in differentiated PC12 cells via
inhibition of Ca(2+)-dependent NADPH oxidase. Cell Mol Neurobiol.
36:541–551. 2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wagener FA, Dekker D, Berden JH,
Scharstuhl A and van der Vlag J: The role of reactive oxygen
species in apoptosis of the diabetic kidney. Apoptosis.
14:1451–1458. 2009.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yang H, Villani RM, Wang H, Simpson MJ,
Roberts MS, Tang M and Liang X: The role of cellular reactive
oxygen species in cancer chemotherapy. J Exp Clin Cancer Res.
37(266)2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Cavallucci V, D'amelio M and Cecconi F:
Abeta toxicity in Alzheimer's disease. Mol Neurobiol. 45:366–378.
2012.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li C, Jiang W, Liu ZG, Liang PQ and Hu R:
Role of reactive oxygen species in GDC-0152-induced apoptosis and
autophagy of NB4 cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi.
27:1786–1793. 2019.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
32
|
Prasad S, Gupta SC and Tyagi AK: Reactive
oxygen species (ROS) and cancer: Role of antioxidative
nutraceuticals. Cancer Lett. 387:95–105. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kehrer JP and Klotz LO: Free radicals and
related reactive species as mediators of tissue injury and disease:
Implications for Health. Crit Rev Toxicol. 45:765–798.
2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
He L, He T, Farrar S, Ji L, Liu T and Ma
X: Antioxidants maintain cellular redox homeostasis by elimination
of reactive oxygen species. Cell Physiol Biochem. 44:532–553.
2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Tsikas D: Assessment of lipid peroxidation
by measuring malondialdehyde (MDA) and relatives in biological
samples: Analytical and biological challenges. Anal Biochem.
524:13–30. 2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Del RD, Stewart AJ and Pellegrini N: A
review of recent studies on malondialdehyde as toxic molecule and
biological marker of oxidative stress. Nutr Metab Cardiovasc Dis.
15:316–328. 2005.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Gallo M, Sapio L, Spina A, Naviglio D,
Calogero A and Naviglio S: Lactic dehydrogenase and cancer: An
overview. Front Biosci (Landmark Ed). 20:1234–1249. 2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chong CM and Zheng W: Artemisinin protects
human retinal pigment epithelial cells from hydrogen
peroxide-induced oxidative damage through activation of ERK/CREB
signaling. Redox Biol. 9:50–56. 2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li S, Chaudhary SC, Zhao X, Gaur U, Fang
J, Yan F and Zheng W: Artemisinin protects human retinal pigmented
epithelial cells against hydrogen peroxide-induced oxidative damage
by enhancing the activation of AMP-active protein kinase. Int J
Biol Sci. 15:2016–2028. 2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Green DR and Llambi F: Cell death
signaling. Cold Spring Harb Perspect Biol.
7(a006080)2015.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Maddika S, Ande SR, Panigrahi S,
Paranjothy T, Weglarczyk K, Zuse A, Eshraghi M, Manda KD, Wiechec E
and Los M: Cell survival, cell death and cell cycle pathways are
interconnected: Implications for cancer therapy. Drug Resist Updat.
10:13–29. 2007.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
43
|
Lin HH, Chen JH, Huang CC and Wang CJ:
Apoptotic effect of 3,4-dihydroxybenzoic acid on human gastric
carcinoma cells involving JNK/p38 MAPK signaling activation. Int J
Cancer. 120:2306–2316. 2007.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–169. 2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Maulik N, Goswami S, Galang N and Das DK:
Differential regulation of Bcl-2, AP-1 and NF-kappaB on
cardiomyocyte apoptosis during myocardial ischemic stress
adaptation. FEBS Lett. 443:331–336. 1999.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Khalilzadeh B, Shadjou N, Kanberoglu GS,
Afsharan H, de la Guardia M, Charoudeh HN, Ostadrahimi A and
Rashidi MR: Advances in nanomaterial based optical biosensing and
bioimaging of apoptosis via caspase-3 activity: A review. Mikrochim
Acta. 185(434)2018.PubMed/NCBI View Article : Google Scholar
|