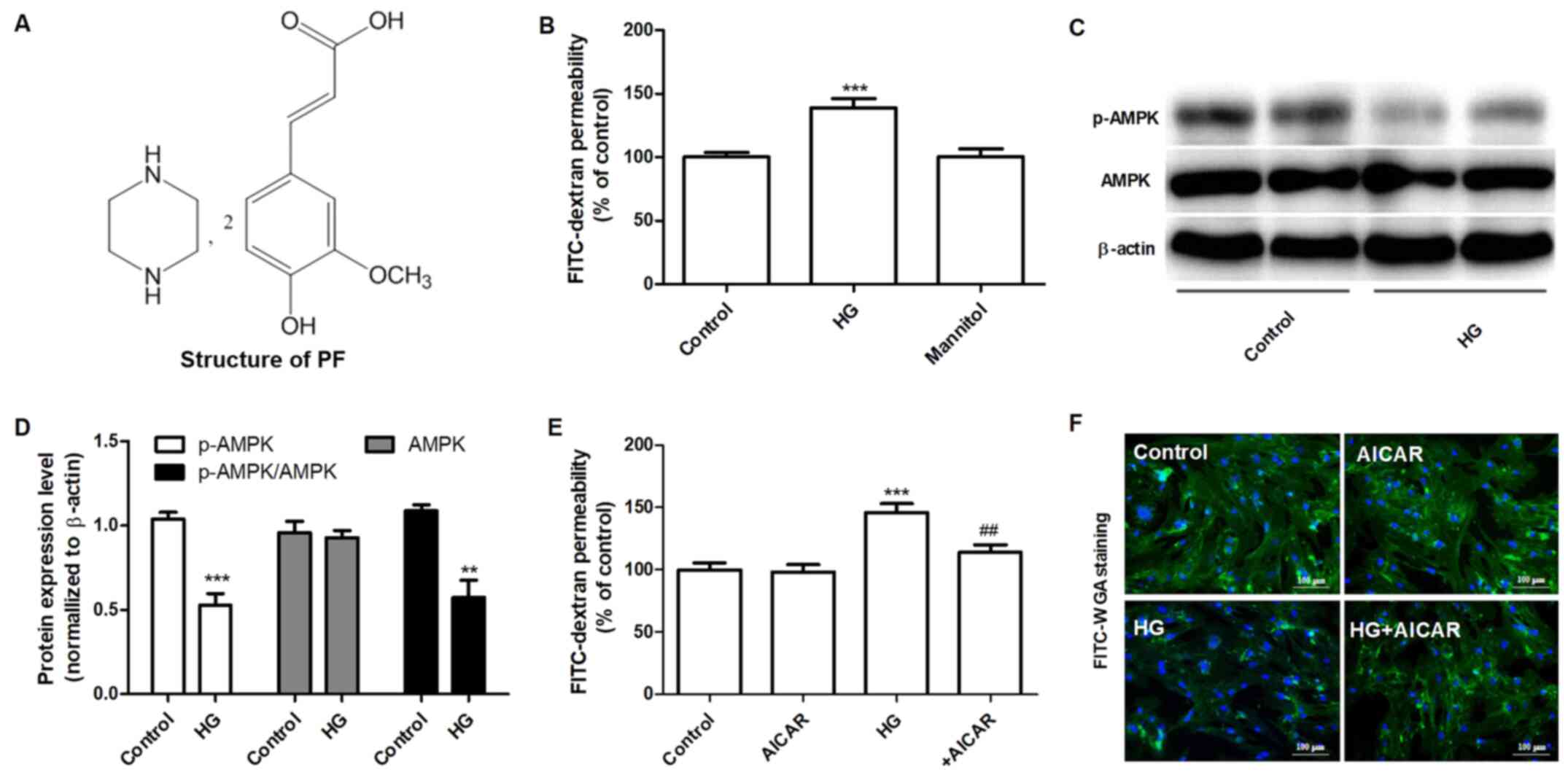
Introduction
Diabetic nephropathy (DN) is associated with damage
to the glomerular filtration barrier (GFB), which consists of the
three following layers: The endothelial layer, the glomerular
basement membrane and the podocytes (1,2). GFB
is responsible for filtering water and small molecules from
circulating plasma. Accumulating evidence has demonstrated that
glomerular endothelial cells (GEnCs) act as the first barrier to
prevent macromolecules from passing the GFB (2,3).
Glycocalyx is the main component of the endothelial surface layer,
which constitutes a molecular size- and charge-specific selective
barrier together with the fenestrations (4,5). The
degradation of the glycocalyx and the decreased number of
fenestrations are the most important characteristics of filtration
barrier injury to GEnCs during DN (6). Since glycocalyx dysfunction exerts an
important role in the development of DN, the prevention of
glycocalyx degradation may be an alternative approach to treat
DN.
Adenosine monophosphate-activated protein kinase
(AMPK) is a ubiquitous heterotrimeric protein composed of a
catalytic subunit (α) and two regulatory subunits (β and γ)
(7). A previous study demonstrated
that AMPK is an important molecule that regulates the progression
of DN (8). Renal hypertrophy
(9) and podocyte apoptosis
(10) in diabetes are regulated by
AMPK, whereas metformin, an activator of AMPK, has been
demonstrated to decrease albuminuria in diabetic rats as well as in
patients with type 2 diabetes mellitus (11,12).
Furthermore, AMPK has been shown to mediate low shear
stress-induced glycocalyx impairment (13). This evidence indicates that AMPK may
be a possible drug target to prevent glomerular filtration
injury.
Piperazine ferulate (Piperazine
3-methoxy-4-hydroxycinnamate; PF; Fig.
1A) is a derivative of ferulic acid that exhibits
anti-hypertensive effects (14),
reduces IgA-mediated nephropathy (15) and prevents the development of DN
(16). A systematic meta-analysis
indicated that PF combined with irbesartan could improve the
efficiency of treatment of DN (17). Our previous study also demonstrated
that PF retarded the progression of DN (16) and attenuated the high glucose
(HG)-induced mesangial cell injury (18). However, it remains unclear whether
the mechanism of PF against DN is associated with the regulation of
GEnC filtration barrier injury. In the present study, the effects
of PF on the HG-induced filtration barrier injury of GEnCs were
investigated. Moreover, the potential mechanism of its action was
assessed.
Materials and methods
Materials
PF was provided by Hunan QianJinXiangJiang
Pharmaceutical Industry Co., Ltd. Fluorescein isothiocyanate
(FITC)-dextran and FITC conjugated wheat germ agglutinin (FITC-WGA)
were obtained from Sigma-Aldrich (Merck KGaA). Rabbit polyclonal
anti-occludin-1 (cat. no. DF7504) and anti-heparanase (Hpa-1)
antibodies (cat. no. DF12411) were purchased from Affinity
Biosciences. Rabbit polyclonal anti-AMPK (cat. no. 2532) and
anti-phosphorylated (p)-AMPKα (Thr172) antibodies (cat. no. 2535)
were obtained from Cell Signaling Technology, Inc. Rabbit
polyclonal anti-Zonula occludens-1 (ZO-1; cat. no. PB9234) and
mouse monoclonal anti-β-actin antibody (cat. no. BM0627), as well
as the syndecan-1 (Sdc-1) ELISA kits (cat. no. EK1339 and EK1554)
were purchased from Boster Biological Technology, Ltd.
SYBR® Premix Ex Taq™ and PrimeScript reverse
transcription reagent kit were obtained from Takara Bio, Inc.
TRIzol® was obtained from Thermo Fisher Scientific, Inc.
RIPA lysis buffer, 5-aminoimidizole-4-carboxamide riboside (AICAR),
the bicinchoninic acid (BCA) assay kit and a BeyoECL Plus kit were
obtained from Beyotime Institute of Biotechnology. Glucose and
mannitol were purchased from Guoyao Chemical Reagent Co. Compound c
was obtained from Selleck Chemicals.
Cell culture and treatment
Human GEnCs were purchased from the ScienCell
Research Laboratories, Inc. and cultured under standard cell
culture conditions at 37˚C with 5% CO2 in a humidified
incubator using endothelial cell medium (ECM; Shanghai Zhong Qiao
Xin Zhou Biotechnology Co., Ltd.), supplemented with 100 U/ml
penicillin, 100 µg/ml streptomycin, 10% (v/v) FBS and 1%
endothelial cell growth factor (Zhongqiaoxinzhou Biotech). Cells
between passages 3 and 8 were used for the experiments. To study
the role of AMPK in the protective effect of PF on HG-induced-GEnCs
injury, GEnCs were preincubated with 20 µM Compound c for 1 h and
subsequently treated with HG (30 mM) in the presence or absence of
PF for 48 h. Next, the permeability of GEnCs to FITC-dextran was
determined.
Endothelial permeability assays
FITC-dextran was used to measure permeability of
GEnC monolayers. GEnC monolayers were cultured on the apical
chamber of the Transwell inserts (0.4 µm pore size; Corning, Inc.)
until they reached 100% confluence and were subsequently treated
with 5.5 mM glucose (control), 30 mM HG or mannitol (5.5 mM glucose
+ 24.5 mM mannitol) with or without PF (25-200 µM) for the
indicated time periods. Dextran was dissolved in ECM and added to
the apical chamber at a concentration of 2.0 g/l following
different experimental treatments. Following 1 h of incubation, the
amount of fluorescence of FITC-dextran in the basolateral chamber
was measured using fluorescence spectrophotometry with an
excitation and emission wavelength of 490 and 520 nm, respectively.
The results are presented as fold-change following normalization to
the control group.
Small interfering RNA (siRNA)-mediated
knockdown
Specific and negative control siRNAs were purchased
from Guangzhou RiboBio Co., Ltd. siRNA-AMPK (40 nM; cat. no.
siB1456165958-1-5) was transfected into GEnCs using Dharma FECT
transfection reagent (Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. The negative control group were GEnCs
transfected with scrambled RNA (40 nM, cat. no. siN0000001-1-5).
Following transfection, the cells were incubated for 24 h and used
for further experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from renal tissues using
TRIzol® reagent and cDNA was synthesized using a
PrimeScript reverse transcription reagent kit according to the
manufacturer's protocol. RT-qPCR was performed using a
LightCycler96® (Roche Diagnostics GmbH) and SYBR-Green
master mix. The RT-qPCR cycling conditions were as follows: Initial
denaturation at 95˚C for 30 sec followed by 45 cycles of
denaturation at 95˚C for 5 sec and annealing 60˚C for 30 sec. The
species-specific primer sequences were as follows: Mouse occludin-1
forward, 5'-CTACGGAGGTGGCTATGGAG-3' and reverse,
5'-AAGGAAGCGATGAAGCAGAA-3'; mouse ZO-1 forward,
5'-ATGACTCCTGACGGTTGGTC-3' and reverse, 5'-GGCTCCAACAAGGTAATTCG-3';
and mouse β-actin forward, 5'-ACTGCTCTGGCTCCTAGCAC-3' and reverse,
5'-ACATCTGCTGGAAGGTGGAC-3'. Gene expression was calculated using
the 2-ΔΔCq method (19).
The data are expressed as the fold-change normalized to
β-actin.
Western blot analysis
Total protein was extracted from cells using RIPA
lysis buffer and the protein concentration was determined using the
BCA method according to the manufacturer's protocol. A total of 35
µg protein/lane was loaded onto a SDS-PAGE (10%) and separated at
120 V. The proteins were transferred to PVDF membranes and blocked
in 5% non-fat milk for 1 h at room temperature. Subsequently, the
blots were incubated with primary antibodies against ZO-1 (1:500),
occludin-1 (1:500), AMPK (1:500), hpa-1 (1:500), p-AMPKα (Thr172,
1:300) and β-actin (1:500) at 4˚C overnight. The blots were
incubated with HRP-conjugated goat anti-mouse (Boster Biological
Technology; cat. no. BA1050) or HRP-conjugated goat anti-rabbit
(Boster Biological Technology; cat. no. BA1054) diluted with
secondary antibody dilution buffer (Beyotime Institute of
Biotechnology, cat. no. P0023D) for 1 h at room temperature. The
bands were visualized using a BeyoECL Plus kit. Densitometry
analysis was performed using ImageJ version 1.45s (National
Institutes of Health). The protein expression levels were
normalized to those of β-actin.
Animal model
The animal experimental protocol was approved by the
Ethics Committee of Animal Experiments of the Central South
University and was performed in accordance with the Guidelines for
the Care and Use of Laboratory Animals (20). Male C57BL/6J mice, aged 8 weeks
(weight, 19-22 g), were purchased from the Experimental Animal
Center of SiLaiKeJingDa. The animals were maintained under a
constant 12 h light/dark cycle at 22±2˚C and allowed free access to
food and water at least for 1 week. Type 1 diabetic mice were
established using a streptozotocin (STZ) injection as previously
described (16). Briefly, the mice
were intraperitoneally injected for 5 consecutive days with STZ (65
mg/kg body weight). After 2 days, the levels of blood glucose were
measured by the mouse tail snipping method using a glucometer
(Sannuo Biotech Ltd.). Mice with blood glucose levels ≥16.7 mmol/l
were considered diabetic. Normal control mice were injected with
citrate buffer solution (0.1 M, pH 4.0). Then, 1 week after blood
glucose stabilization, diabetic mice were divided into the two
following groups (n=15 per group): The diabetic mice group were
treated with vehicle (DM) and the PF group of mice treated with PF
(100 mg/kg, DM+PF). PF was provided by intragastric administration
daily for 12 weeks. All mice were anaesthetized using sodium
pentobarbital (50 mg/kg body weight) and blood samples (1-1.2 ml)
were collected from the retro-orbital sinus after eyeball removal.
Finally, the mice were sacrificed by exsanguination under
anesthesia (50 mg/kg sodium pentobarbital) and the kidney tissues
were collected. Collected blood was left to stand at room
temperature for 1 h, and then centrifuged at 2,000 x g at 4˚C for
15 min. The supernatant was transferred to a new tube and stored at
-80˚C.
Immunofluorescence analysis
Renal tissues were fixed in 4% paraformaldehyde at
4˚C overnight, embedded in paraffin and cut into 5 µm-thick slices.
The slices were processed by dewaxing, gradient alcohol
dehydration, antigen repair with 10 mM sodium citrate (pH 6.0,
microwaved for 10 min) and finally washed in PBS three times. The
sections were incubated with FITC-WGA (10 µg/ml) at room
temperature for 2 h, stained with DAPI at room temperature for 10
min and examined using fluorescence microscopy (Zeiss AG;
magnification, x400). Immunofluorescence was performed by
incubating the slides with ZO-1 (1:200)/CD31 (1:200) or Sdc-1
(1:200)/CD31 (1:200) overnight at 4˚C. The slides were incubated
with the corresponding anti-FITC-conjugated secondary antibody
(Beyotime Institute of Biotechnology; cat. no. A0562; 1:500) or
Cy3-conjugated secondary antibody (Beyotime Institute of
Biotechnology; cat. no. A0516; 1:500) diluted with secondary
antibody dilution buffer for 1 h at room temperature in the dark.
The cell nuclei were stained with DAPI at room temperature for 5
min. Finally, the images were captured using fluorescence
microscopy (magnification, x400).
Immunohistochemical analysis
The renal tissue slices were incubated overnight
with anti-p-AMPKα (Thr-172) or Hpa-1 antibodies at 4˚C overnight,
followed by incubation with the corresponding secondary antibody
conjugated with horseradish peroxidase (Boster Biological
Technology; cat. no. BM3894; diluted with secondary antibody
dilution buffer, 1:1,000) and 3,3'-diaminobenzidine peroxidase
substrate. Finally, the sections were stained with hematoxylin at
room temperature for 1 min and evaluated using conventional light
microscopy (Olympus Corporation; magnification, x400).
Transmission electron microscopy
The kidney tissues were fixed in 2.5% glutaraldehyde
at 4˚C overnight and post-fixed in 1% osmium tetroxide at 4˚C for 2
h. Following dehydration, the samples were embedded in EMbed 812
resin (Thermo Fisher Scientific, Inc.) and polymerized at 60˚C for
48 h. The ultrathin sections (60-80 nm) were cut and stained with
1% uranyl acetate at room temperature for 10 min, followed by 2%
lead citrate buffer for 2 min at 37˚C. The structure of the
glomerulus was observed using a Hitachi High-Tech 7700 electron
microscope (magnification, x5,000).
Determination of the expression levels
of soluble Sdc-1
The levels of soluble Sdc-1 in the cell culture
supernatant or serum were measured using the Sdc-1 ELISA kits
according to the manufacturer's protocols.
Determination of glycocalyx via
endothelial surface analysis
Following treatment, GEnCs were washed with PBS and
fixed in 4% paraformaldehyde at room temperature for 10 min.
Subsequently, the cells were incubated with FITC-WGA(10 µg/ml) at
room temperature for 30 min. Finally, the images were captured
using fluorescence microscopy (magnification, x400).
Statistical analysis
Data are presented as the mean ± standard error of
at least three independent experiments. Data were analyzed using
SPSS software (version 17.0; SPSS, Inc.). A one-way ANOVA followed
by a post-hoc Tukey's test was performed to compare difference
between multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
AMPK mediates HG-induced filtration
barrier injury of GEnCs
The effects of HG on the permeability of the GEnC
barrier to FITC-dextran were examined. HG (30 mM) increased the
permeability of FITC-dextran after 48 h, whereas mannitol did not
have an effect for 48 h (Fig. 1B).
Subsequently, the effects of HG on the expression levels of AMPK
and p-AMPKα (Thr-172) were measured by western blot analysis. GEnCs
treated with HG exhibited decreased levels of p-AMPKα (Thr-172),
although the total expression levels of AMPK were not altered
notably (Fig. 1C and D). The effects of AMPK on the permeability
of GEnC to FITC-dextran were further examined using AICAR, a
selective agonist of AMPK. As shown in Fig. 1E, GEnCs pretreated with AICAR (1 mM)
decreased the HG-induced permeability of GEnC to FITC-dextran.
Similarly, the results of FITC-WGA staining demonstrated that AICAR
significantly inhibited the decrease of the glycocalyx content in
GEnC induced by HG (Fig. 1F). These
data suggested that AMPK mediated HG-induced filtration barrier
injury of GEnCs.
AMPK regulates the expression of Hpa-1
and tight junction (TJ) proteins
Sdc-1 is the main constituent of the glycocalyx.
This protein can be cleaved by Hpa-1, an enzyme capable of
degrading polymeric heparan sulfate molecules of Sdc-1 into shorter
chain length oligosaccharides (21). HG increased the expression levels of
Hpa-1 without affecting the expression levels of Sdc-1 in GEnCs
(Fig. 2A). Therefore, the
expression levels of Sdc-1 in the cell culture supernatant were
determined. The results demonstrated that the expression levels of
Sdc-1 were increased in the HG group. These changes were
significantly inhibited by co-incubation of the cells with AICAR
(Fig. 2A-D). Furthermore, the
effects of AICAR on the expression of TJ proteins, ZO-1 and
occludin-1 were investigated. AICAR significantly inhibited the
HG-induced downregulation of the expression of occludin-1 and ZO-1
at the protein level (Fig. 2A,
E and F). These data suggested that AMPK mediated
the HG-induced filtration barrier injury of GEnCs by regulation of
the expression levels of Hpa-1 and TJ proteins.
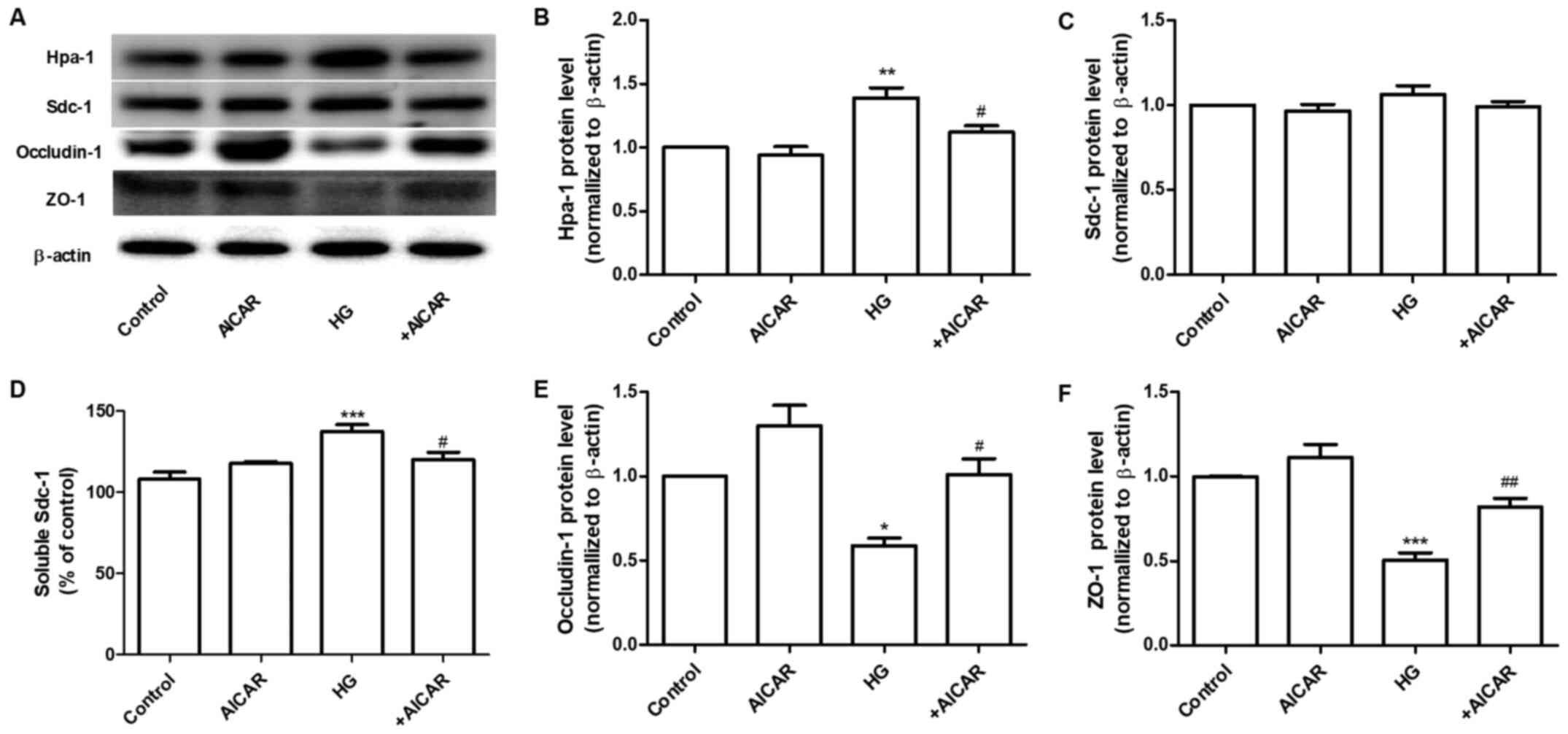 | Figure 2AMPK regulates the expression of
Hpa-1 and TJ proteins. (A) Western blot analysis of Hpa-1, Sdc-1,
occludin-1 and ZO-1 expression. Densitometric analysis of (B) Hpa-1
and (C) Sdc-1 expression. (D) Expression levels of Sdc-1 in the
cell culture supernatant. (E) Densitometric analysis of occludin-1
and (F) ZO-1 expression. Data are presented as the mean ± standard
deviation of three repeats. *P<0.05,
**P<0.01, ***P<0.001 vs. the control
group; #P<0.05, ##P<0.01 vs. HG. AMPK,
adenosine monophosphate-activated protein kinase; Hpa-1,
heparanase-1; TJ, tight junction; Sdc-1, syndecan-1; ZO-1, Zonula
occludens-1; HG, high glucose. |
PF attenuates HG-induced filtration
barrier damage
The effects of PF on the permeability defects of
GEnCs induced by HG were assessed. Incubation of GEnCs with PF (100
and 200 µM) significantly inhibited the increase noted in the
HG-induced permeability of GEnCs to FITC-dextran (Fig. 3A). The specific dose of PF used for
further investigations was 100 µM. Preincubation of GEnCs with PF
attenuated the increase of soluble Sdc-1 expression levels in the
cell culture supernatant (Fig. 3B).
Moreover, it increased the glycoclayx content of GEnCs (Fig. 3C and D). PF ameliorated the HG-induced increase
in the expression levels of Hpa-1 protein in GEnCs (Fig. 3E and F). Similarly, PF inhibited the decrease in
the expression levels of ZO-1 and occludin-1 in GEnCs exposed to HG
(Fig. 3E, G and H).
GEnCs incubated with PF exhibited a significant increase in the
p-AMPK/AMPK ratio compared with those noted in the HG group
(Fig. 3E and I). These finding suggested that PF
inhibited HG-induced filtration barrier injury in GEnCs.
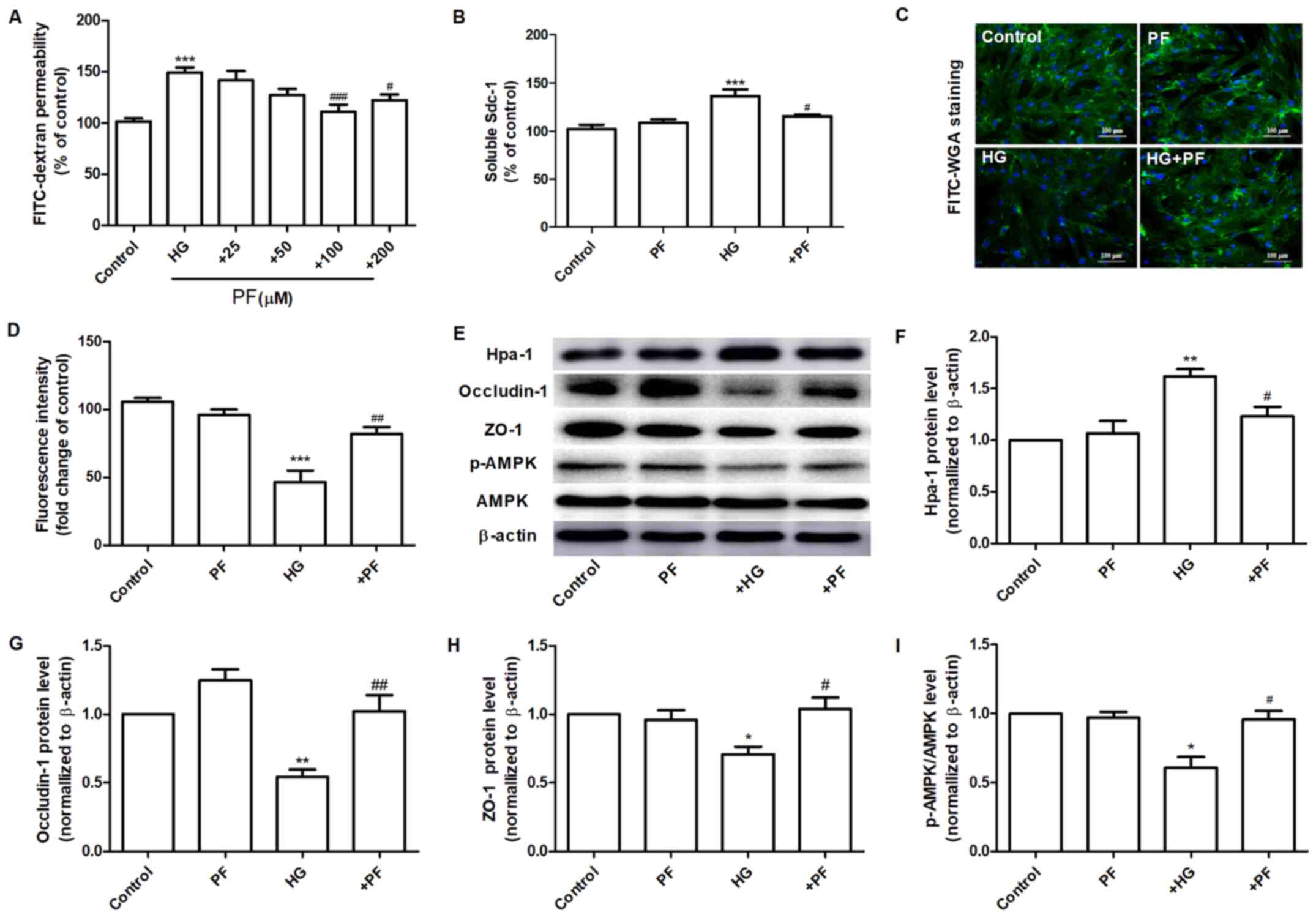 | Figure 3PF attenuates HG-induced filtration
barrier damage. (A) Effects of various concentrations of PF on
HG-induced permeability of GEnCs to FITC-dextran. (B) Expression
levels of Sdc-1 in the cell culture supernatant. (C) Representative
images of glycocalyx stained with FITC-WGA. (D) Quantification
analysis of FITC-WGA staining. (E) Western blot analysis of Hpa-1,
occludin-1, ZO-1, p-AMPK and total AMPK. Densitometry analysis of
(F) Hpa-1, (G) occludin-1, (H) ZO-1 and (I) the p-AMPK/AMPK ratio.
Data are presented as the mean ± standard deviation of three
repeats. *P<0.05, **P<0.01,
***P<0.001 vs. control group; #P<0.05,
##P<0.01, ###P<0.001 vs. HG group. PF,
piperazine ferulate; HG, high glucose; GEnCs, glomerular
endothelial cells; FITC, fluorescein isothiocyanate; WGA, wheat
germ agglutinin; Hpa-1, heparanase-1; ZO-1, Zonula occludens-1; p-,
phosphorylated-; AMPK, adenosine monophosphate-activated protein
kinase. |
PF alleviates HG-induced filtration
barrier injury via AMPK
The regulatory mechanism of PF on HG-induced
filtration barrier injury was further investigated. GEnCs were
preincubated with Compound c (AMPK inhibitor, 20 µM) for 1 h and
subsequently treated with HG (30 mM) in the presence or absence of
PF for 48 h. Compound c abolished the protective effects of PF on
the HG-induced increased of the permeability of GEnCs to
FITC-dextran and increased the Sdc-1 levels in the cell culture
supernatant (Fig. 4A and B). Following knockdown of AMPK in GEnCs by
siRNA (Fig. S1), the effects of PF
on the HG-induced upregulation of Hpa-1 expression and the
downregulation of ZO-1 and occludin-1 were inhibited (Fig. 4C-F). Taken together, the data
revealed that AMPK mediated the protective effects of PF on the
HG-induced filtration barrier injury.
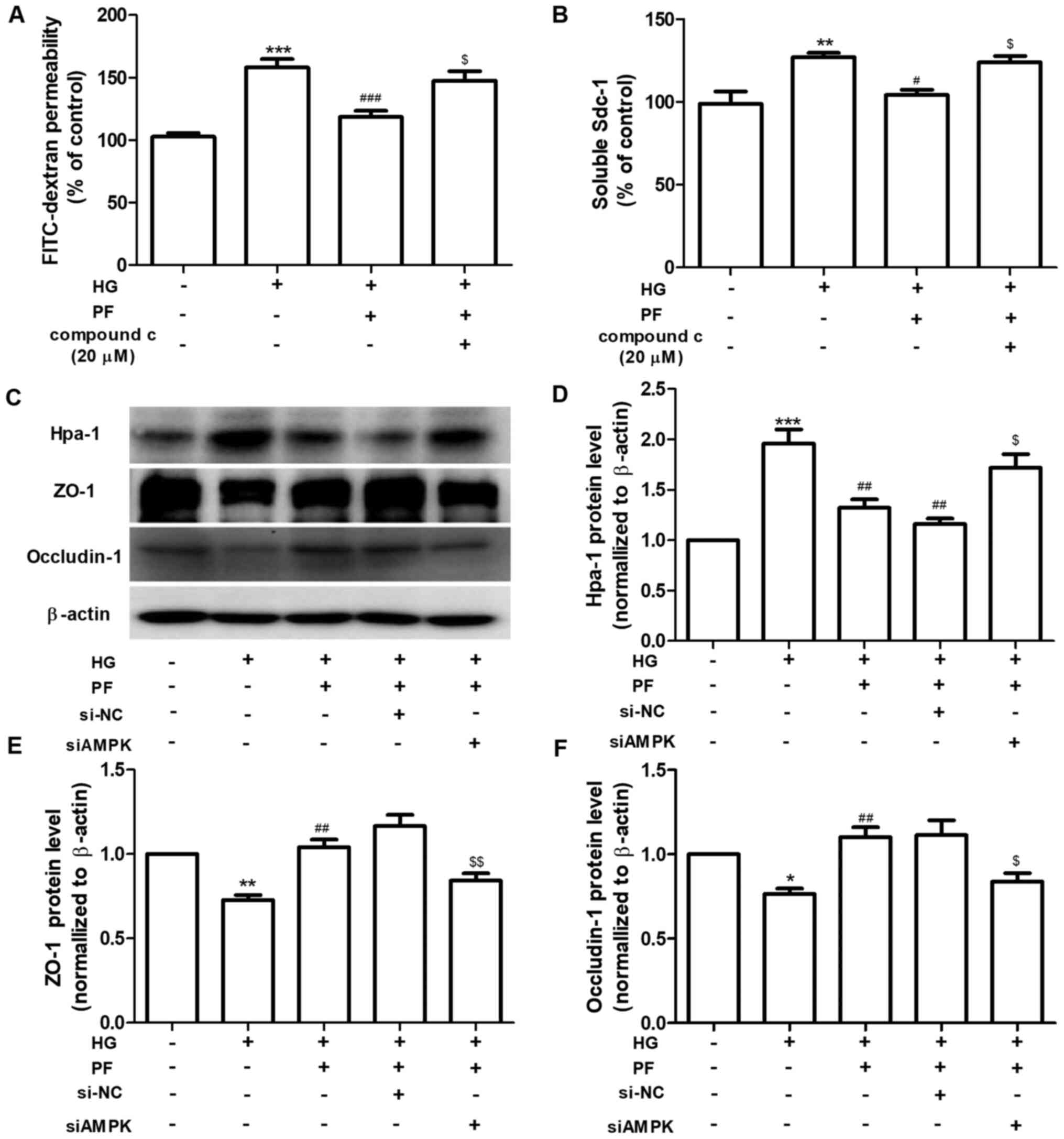 | Figure 4PF alleviates HG-induced filtration
barrier injury via the AMPK pathway. (A) Permeability of GEnCs to
FITC-dextran. (B) Expression levels of Sdc-1 in the cell culture
supernatant. (C) Western blot analysis of Hpa-1, ZO-1 and
occludin-1. Densitometry analysis of (D) Hpa-1, (E) ZO-1 and (F)
occludin-1. Data are presented as the mean ± standard deviation of
three repeats. *P<0.05, **P<0.01,
***P<0.001 vs. control. #P<0.05,
##P<0.01, ###P<0.001 vs. HG;
$P<0.05, $$P<0.01 vs. HG + PF or siCon
+ HG + PF. PF, piperazine ferulate; HG, high glucose; AMPK,
adenosine monophosphate-activated protein kinase; GEnCs, glomerular
endothelial cells; FITC, fluorescein isothiocyanate; Sdc-1,
syndecan-1; Hpa-1, heparanase-1; ZO-1, Zonula occludens-1; si,
small interfering; NC, negative control. |
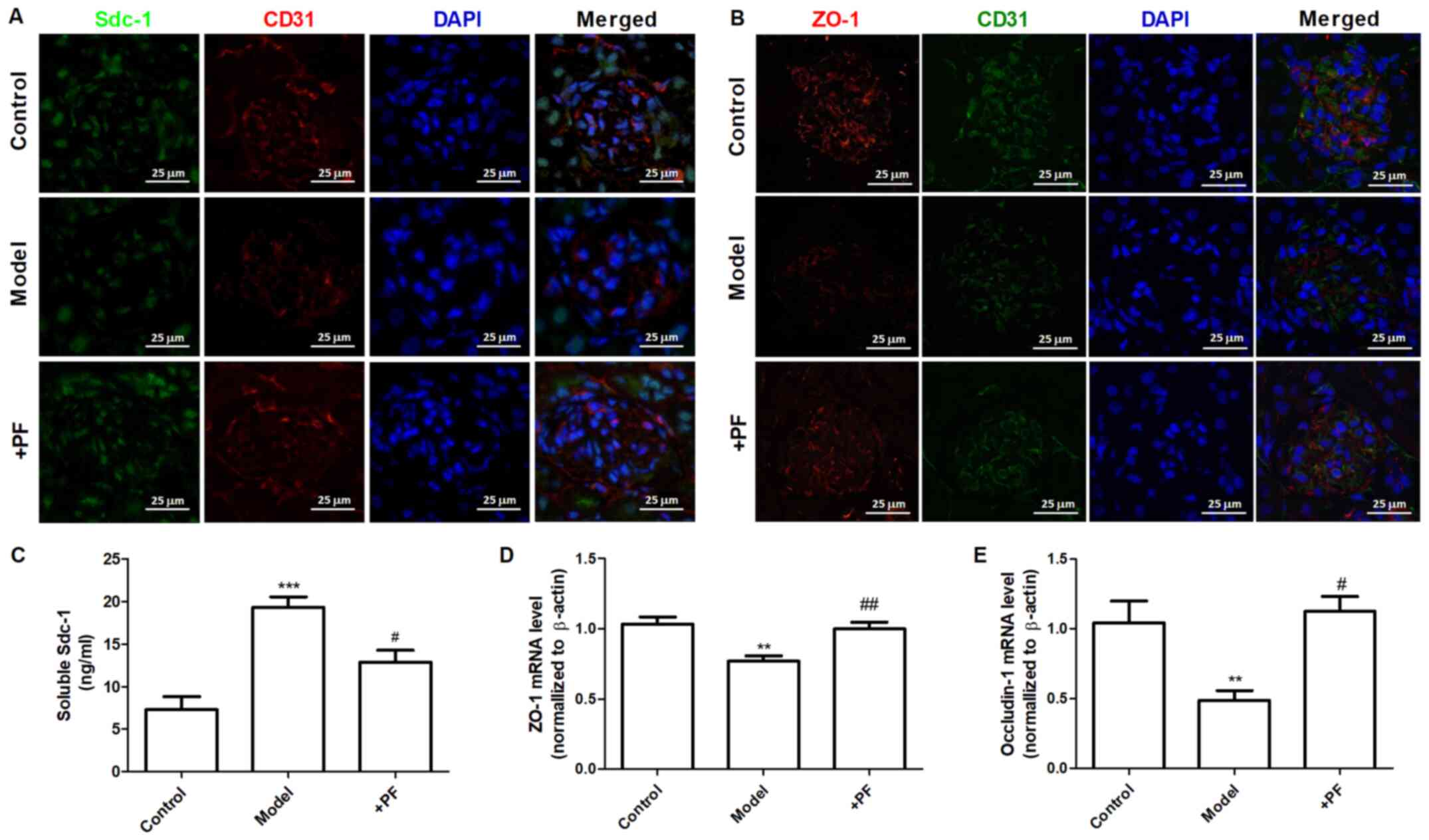
PF alleviates endothelial glycocalyx
injury in vivo
In vitro experiments demonstrated that
diabetic mice treated with PF for 12 weeks exhibited an increase in
the number of GEnC fenestrations compared with that of the model
group (Fig. 5A and B). Immunofluorescence analysis indicated
that the glycocalyx content was increased in the glomerular cells
of the PF-treated group compared with that of the diabetic mice
(Fig. 5C and D). Immunohistochemical analysis indicated
that PF significantly increased p-AMPKα (Thr-172) protein
expression (Fig. 5E and F) and inhibited the increase in Hpa-1
protein levels in the glomerular cells of diabetic mice (Fig. 5G and H). Immunofluorescence analysis indicated
that the expression levels of Sdc-1 (Fig. 6A) and ZO-1 (Fig. 6B) in GEnCs were increased in
PF-treated mice compared with those noted in diabetic mice. In
addition, treatment of diabetic mice with PF inhibited the increase
noted in the expression levels of Sdc-1 in the serum (Fig. 6C). It concomitantly increased the
expression levels of ZO-1 and occludin-1 in kidney tissues
(Fig. 6D and E). Taken together, these finding indicated
that PF reduced GFB damage.
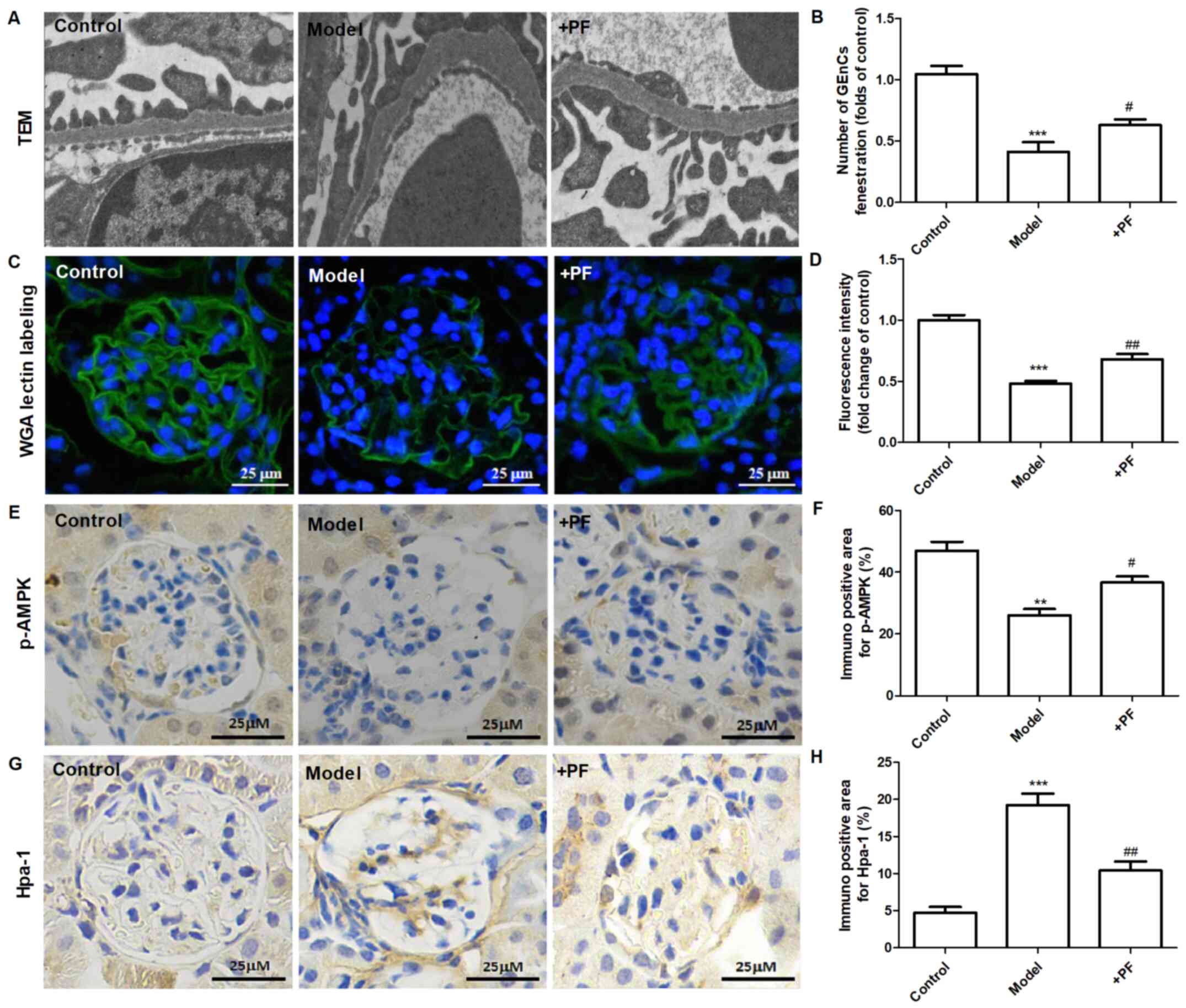 | Figure 5PF alleviates endothelial glycocalyx
injury in vivo. (A) Transmission electron microscopy of
glomeruli extracted from different groups of animals. (B) The
number of glomerular endothelial fenestrations. (C) FITC-WGA
staining for endothelial glycocalyx in the glomerulus.
Magnification, x400. (D) Quantification analysis of FITC-WGA
staining. (E) Immunohistochemical images of p-AMPKα (Thr-172).
Magnification, x400. (F) Quantification analysis of
immunohistochemical staining of p-AMPKα (Thr-172). (G)
Immunohistochemical staining of Hpa-1. Magnification, x400. (H)
Quantification analysis of immunohistochemical staining of Hpa-1.
Data are presented as the mean ± standard deviation of three
repeats. **P<0.01, ***P<0.01 vs. the
control group; #P<0.05, ##P<0.01 vs.
the model group. PF, piperazine ferulate; FITC, fluorescein
isothiocyanate; WGA, wheat germ agglutinin; p-, phosphorylated-;
AMPK, adenosine monophosphate-activated protein kinase; Hpa-1,
heparanase-1. |
Discussion
Endothelial barrier injury is characterized by
increased endothelial permeability and is associated with acute and
chronic nephropathy (22). Emerging
evidence has revealed that damage to the GEnC barrier plays an
important role in the onset and progression of DN development
(4). Our previous study
demonstrated that PF attenuated the levels of 24 h-albuminuria,
blood urea nitrogen and serum creatinine in STZ-induced DN mice
(16), and it also reduced
HG-induced mesangial cell injury (18). The results of the present study
indicated that PF exerted a protective effect on GFB injury. The
effects of PF were investigated on HG-induced filtration barrier
injury of GEnCs, and the results suggested that this compound could
reduce the damage to the glycocalyx degradation noted in GEnCs.
These findings highlight a novel mechanism by which PF retards the
progression of DN. This mechanism of action may restore endothelial
filtration barrier injury.
Endothelial glycocalyx maintains the vascular
function and plays an important role in vascular permeability,
leucocyte, platelet adhesion as well as transduction of fluid
shearing forces (23,24). FITC-WGA has been shown to bind to
the glycoprotein of the glycocalyx (25). This marker was used to evaluate the
effects of PF on glomerular endothelial glycocalyx. The data
indicated that PF prevented HG-induced glycocalyx degradation in
vivo and in vitro. Hpa-1 is an endo-β-D-glucuronidase
that releases 5-7 kDa fragments of heparan sulfate from intact
heparan sulfate chains of proteoglycans (26). In addition, Hpa-1 regulates the
expression levels and turnover of syndecans (27). The results of the present study
showed that HG increased the expression levels of Hpa-1 in
vitro and in vivo. PF attenuated the increased
expression of Hpa-1 in HG-induced GEnCs as well as in the glomeruli
of diabetic mice. However, the expression levels of Sdc-1 did not
decrease in HG-stimulated GEnCs. The effect of HG on the expression
of Sdc-1is similar to those of Singh et al (28). Immunofluorescence analysis indicated
that Sdc-1 levels in the glomeruli of diabetic mice were decreased,
whereas PF inhibited these changes. The possible reason for the
discrepancy noted between the in vivo and in vitro
experiments is the inconsistent exposure factors used for the
different models. The degradation of glycocalyx in vivo may
be affected by blood glucose levels, inflammatory factors and the
course of diabetes (29). The
exposure of GEnCs to HG involved a short term period (24 h), while
the course of diabetes was completed following 12 weeks.
In the present study, the effects of PF on the TJ of
GEnCs were also evaluated. The expression levels of ZO-1 were
decreased in GEnCs of STZ-induced diabetic mice and in HG-treated
GEnCs, respectively. PF treatment increased the expression levels
of ZO-1. TJs between endothelial cells regulate the passage of ions
and small molecules through the paracellular pathway and are
involved in the pathogenesis of DN (30). The integrity of glomerular GFB is
also maintained by TJ proteins that consist of at least 40
different proteins including transmembrane proteins, cytoplasm
accessory ZO proteins and cytoskeletal proteins (30). HG influences the expression levels
and/or redistribution of ZO-1, which leads to TJ destruction and
filtration barrier injury (31,32).
Thus, cell junctions may serve as a therapeutic target of DN.
AMPK is an important energy-sensing enzyme that
controls cellular energy metabolism and is considered a potential
pharmacological target for the treatment of DN (33,34).
Phosphorylation of the α subunit at Thr-172 is essential for AMPK
activity (35). Pharmacological
activation of AMPK by AICAR results in restoration of the
permeability of podocytes to albumin (36). It also increases the epithelial TJ
assembly following calcium switch (37). In addition, ampkinone is an AMPK
activator, which can be used to attenuate endothelial glycocalyx
impairment induced by low shear stress (13). The results of the present study
demonstrated that AMPK mediated HG-induced glycocalyx impairment
and TJ injury in GEnCs. Based on western blotting, p-AMPK
expression was decreased under HG conditions; however AMPK
expression was not altered; whereas, PF increased the
phosphorylation of AMPK following HG treatment. It is worth noting
that treatment of GEnCs with the AMPK siRNA or inhibitor (compound
c) abolished the protective effects of PF on the HG-induced
filtration barrier injury. Specific molecular mechanisms may be
involved in the phosphorylation of AMPK regulated by PF. A previous
report indicated that ferulic acid, which is one of the components
of PF, regulated the phosphorylation of liver kinase B1 and AMPK in
C2C12 myoblasts via activation of sirtuin 1 (Sirt1) (38). An additional study demonstrated that
ferulic acid stimulated the Sirt1/p-AMPK pathway in human cervical
carcinoma cells (39). Therefore,
PF may regulate AMPK phosphorylation via the Sirt1/LKB1
pathway.
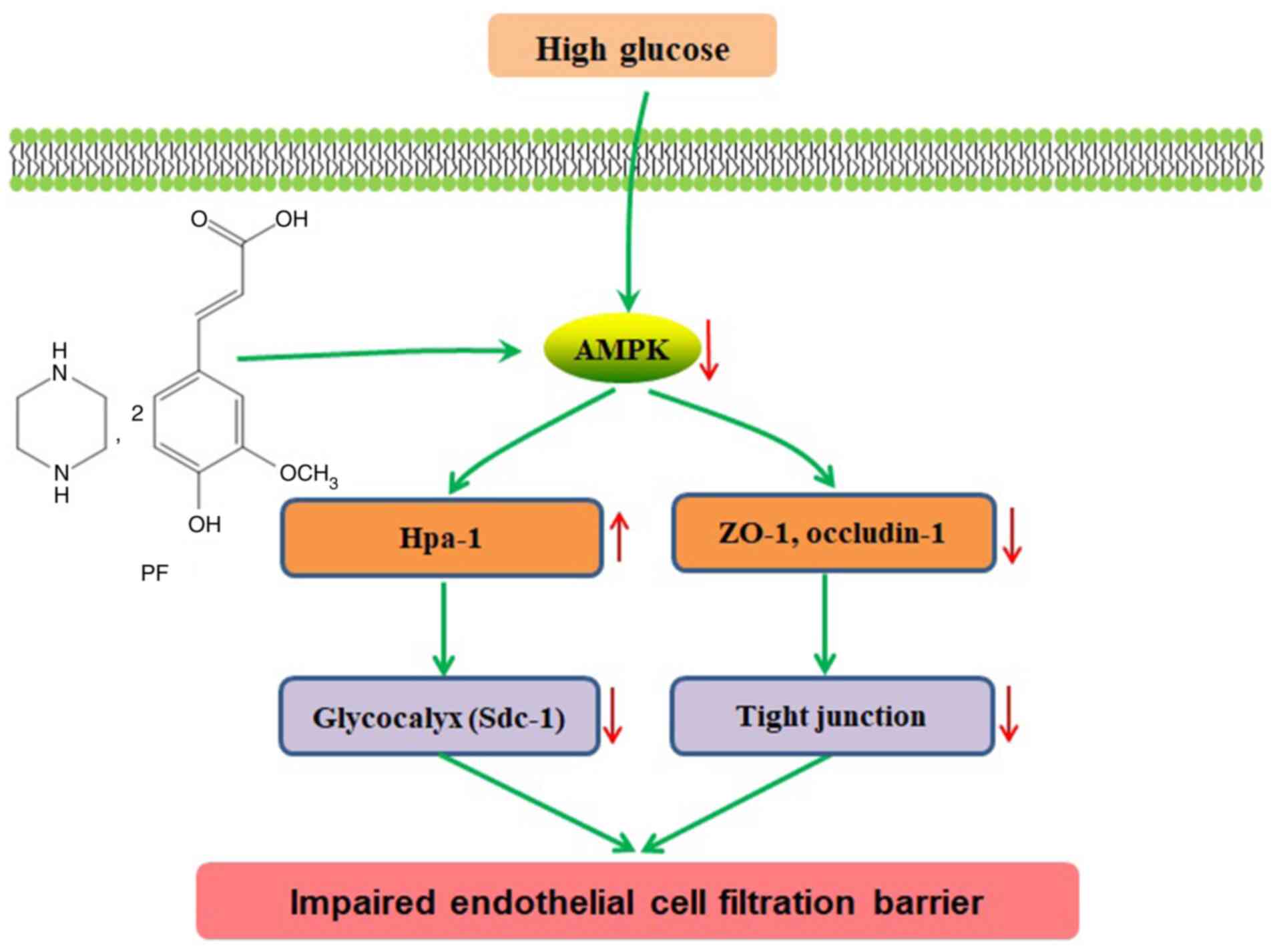
In conclusion, the present study indicated that PF
improved filtration barrier injury of GEnCs in a model of early DN,
and that this action was mediated by AMPK (Fig. 7). Therefore, restoration of GFB
injury via modulation of AMPK activity may serve an alternative
approach for future treatment of DN.
Supplementary Material
Western blotting to confirm
transfection efficiency of si-AMPK-mediated knockdown. Western blot
of AMPK and β-actin expression following transfection with si-AMPK
or the si-NC. si, small interfering; NC, negative control; AMPK,
adenosine monophosphate-activated protein kinase.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported financially by the Hunan
Provincial Natural Scientific Foundation (grant no. 2020JJ5841),
Hunan Traditional Chinese Medicine Science and Technology Project
(grant no. 2021061), National Science and Technology Major Project
Grant (grant no. 2020ZX09201-28), and Open Sharing Fund for the
Large-scale Instruments of Central South University (grant no.
CSUZC202056).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LYY and DXX designed the study. YYY, ZC and XDY
performed the experiments. RRD and LXS contributed to data
analysis. YYY wrote and revised the manuscript. All authors have
read and approved the final manuscript. YYY and LXS confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The animal experimental protocol used in the present
study was approved by the Ethics Committee of Animal Experiments of
the Central South University and was performed in accordance with
the Guidelines for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Boels MGS, Koudijs A, Avramut MC, Sol
WMPJ, Wang G, van Oeveren-Rietdijk AM, van Zonneveld AJ, de Boer
HC, van der Vlag J, van Kooten C, et al: Systemic monocyte
chemotactic protein-1 inhibition modifies renal macrophages and
restores glomerular endothelial glycocalyx and barrier function in
diabetic nephropathy. Am J Pathol. 187:2430–2440. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Song K, Fu J, Song J, Herzog BH, Bergstrom
K, Kondo Y, McDaniel JM, McGee S, Silasi-Mansat R, Lupu F, et al:
Loss of mucin-type O-glycans impairs the integrity of the
glomerular filtration barrier in the mouse kidney. J Biol Chem.
292:16491–16497. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Qi H, Casalena G, Shi S, Yu L, Ebefors K,
Sun Y, Zhang W, D'Agati V, Schlondorff D, Haraldsson B, et al:
Glomerular endothelial mitochondrial dysfunction is essential and
characteristic of diabetic kidney disease susceptibility. Diabetes.
66:763–778. 2017.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Kusano T, Takano H, Kang D, Nagahama K,
Aoki M, Morita M, Kaneko T, Tsuruoka S and Shimizu A: Endothelial
cell injury in acute and chronic glomerular lesions in patients
with IgA nephropathy. Hum Pathol. 49:135–144. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Salmon AH and Satchell SC: Endothelial
glycocalyx dysfunction in disease: Albuminuria and increased
microvascular permeability. J Pathol. 226:562–574. 2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Lavoz C, Matus YS, Orejudo M, Carpio JD,
Droguett A, Egido J, Mezzano S and Ruiz-Ortega M: Interleukin-17A
blockade reduces albuminuria and kidney injury in an accelerated
model of diabetic nephropathy. Kidney Int. 95:1418–1432.
2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Dial AG, Ng SY, Manta A and Ljubicic V:
The role of AMPK in neuromuscular biology and disease. Trends
Endocrinol Metab. 29:300–312. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Packer M: Interplay of adenosine
monophosphate-activated protein kinase/sirtuin-1 activation and
sodium influx inhibition mediates the renal benefits of
sodium-glucose co-transporter-2 inhibitors in type 2 diabetes: A
novel conceptual framework. Diabetes Obes Metab. 22:734–742.
2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lee MJ, Feliers D, Mariappan MM,
Sataranatarajan K, Mahimainathan L, Musi N, Foretz M, Viollet B,
Weinberg JM, Choudhury GG and Kasinath BS: A role for AMP-activated
protein kinase in diabetes-induced renal hypertrophy. Am J Physiol
Renal Physiol. 292:F617–F627. 2007.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Eid AA, Ford BM, Block K, Kasinath BS,
Gorin Y, Ghosh-Choudhury G, Barnes JL and Abboud HE: AMP-activated
protein kinase (AMPK) negatively regulates Nox4-dependent
activation of p53 and epithelial cell apoptosis in diabetes. J Biol
Chem. 285:37503–37512. 2010.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Eisenreich A and Leppert U: Update on the
protective renal effects of metformin in diabetic nephropathy. Curr
Med Chem. 24:3397–3412. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Alhaider AA, Korashy HM, Sayed-Ahmed MM,
Mobark M, Kfoury H and Mansour MA: Metformin attenuates
streptozotocin-induced diabetic nephropathy in rats through
modulation of oxidative stress genes expression. Chem Biol
Interact. 192:233–242. 2011.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang J, Kong X, Wang Z, Gao X, Ge Z, Gu
Y, Ye P, Chao Y, Zhu L, Li X and Chen S: AMP-activated protein
kinase regulates glycocalyx impairment and macrophage recruitment
in response to low shear stress. FASEB J. 33:7202–7212.
2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jianzhi S, Qizeng W, Bin L, Wenhui L,
Yunpeng C, Chenrong F, Lin Z and Huiting C: Piperazine ferulate
exerts antihypertensive effect and improves endothelial function in
vitro and in vivo via the activation of endothelial nitric oxide
synthase. Cell Mol Biol (Noisy-le-grand). 65:119–124.
2019.PubMed/NCBI
|
|
15
|
Liu Z, Pan J, Sun C, Zhou J and Li NA:
Clinical effects of perazine ferulate tablets combined with
eucalyptol limonene pinene enteric soft capsules for treatment of
children with IgA nephropathy. Exp Ther Med. 12:169–172.
2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yang YY, Shi LX, Li JH, Yao LY and Xiang
DX: Piperazine ferulate ameliorates the development of diabetic
nephropathy by regulating endothelial nitric oxide synthase. Mol
Med Rep. 19:2245–2253. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li D, Li B, Peng LX, Liu R and Zeng N:
Therapeutic efficacy of piperazine ferulate combined with
irbesartan in diabetic nephropathy: A systematic review and
meta-analysis. Clin Ther. 42:2196–2212. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang YY, Deng RR, Chen Z, Yao LY, Yang XD
and Xiang DX: Piperazine ferulate attenuates high glucose-induced
mesangial cell injury via the regulation of p66Shc. Mol
Med Rep. 23(374)2021.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Jones-Bolin S: Guidelines for the care and
use of laboratory animals in biomedical research. Curr Protoc
Pharmacol Appendix. 4(Appendix4B)2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yu S, Lv H, Zhang H, Jiang Y, Hong Y, Xia
R, Zhang Q, Ju W, Jiang L, Ou G, et al: Heparanase-1-induced
shedding of heparan sulfate from syndecan-1 in hepatocarcinoma cell
facilitates lymphatic endothelial cell proliferation via VEGF-C/ERK
pathway. Biochem Biophys Res Commun. 485:432–439. 2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zafrani L and Ince C: Microcirculation in
acute and chronic kidney diseases. Am J Kidney Dis. 66:1083–1094.
2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Okada H, Yoshida S, Hara A, Ogura S and
Tomita H: Vascular endothelial injury exacerbates coronavirus
disease 2019: The role of endothelial glycocalyx protection.
Microcirculation. 28(e12654)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhu T, Wang H, Wang L, Zhong X, Huang W,
Deng X, Guo H, Xiong J, Xu Y and Fan J: Ginsenoside Rg1 attenuates
high glucose-induced endothelial barrier dysfunction in human
umbilical vein endothelial cells by protecting the endothelial
glycocalyx. Exp Ther Med. 17:3727–3733. 2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Fukui M, Yamada M, Akune Y, Shigeyasu C
and Tsubota K: Fluorophotometric analysis of the ocular surface
glycocalyx in soft contact lens wearers. Curr Eye Res. 41:9–14.
2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang Y, Macleod V, Miao HQ, Theus A, Zhan
F, Shaughnessy JD Jr, Sawyer J, Li JP, Zcharia E, Vlodavsky I and
Sanderson RD: Heparanase enhances syndecan-1 shedding: A novel
mechanism for stimulation of tumor growth and metastasis. J Biol
Chem. 282:13326–13333. 2007.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Rangarajan S, Richter JR, Richter RP,
Bandari SK, Tripathi K, Vlodavsky I and Sanderson RD:
Heparanase-enhanced shedding of syndecan-1 and its role in driving
disease pathogenesis and progression. J Histochem Cytochem.
68:823–840. 2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Singh A, Friden V, Dasgupta I, Foster RR,
Welsh GI, Tooke JE, Haraldsson B, Mathieson PW and Satchell SC:
High glucose causes dysfunction of the human glomerular endothelial
glycocalyx. Am J Physiol Renal Physiol. 300:F40–F48.
2011.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Reine TM, Lanzalaco F, Kristiansen O,
Enget AR, Satchell S, Jenssen TG and Kolset SO: Matrix
metalloproteinase-9 mediated shedding of syndecan-4 in glomerular
endothelial cells. Microcirculation: Jan 31, 2019 (Epub ahead of
print).
|
|
30
|
Eftekhari A, Vahed SZ, Kavetskyy T,
Rameshrad M, Jafari S, Chodari L, Hosseiniyan SM, Derakhshankhah H,
Ahmadian E and Ardalan M: Cell junction proteins: Crossing the
glomerular filtration barrier in diabetic nephropathy. Int J Biol
Macromol. 148:475–482. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Rincon-Choles H, Vasylyeva TL, Pergola PE,
Bhandari B, Bhandari K, Zhang JH, Wang W, Gorin Y, Barnes JL and
Abboud HE: ZO-1 expression and phosphorylation in diabetic
nephropathy. Diabetes. 55:894–900. 2006.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ha TS, Choi JY, Park HY and Lee JS:
Ginseng total saponin improves podocyte hyperpermeability induced
by high glucose and advanced glycosylation endproducts. J Korean
Med Sci. 26:1316–1321. 2011.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Kim Y and Park CW: New therapeutic agents
in diabetic nephropathy. Korean J Intern Med. 32:11–25.
2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kim Y and Park CW: Adenosine
monophosphate-activated protein kinase in diabetic nephropathy.
Kidney Res Clin Pract. 35:69–77. 2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Morales-Alamo D and Calbet JA: AMPK
signaling in skeletal muscle during exercise: Role of reactive
oxygen and nitrogen species. Free Radic Biol Med. 98:68–77.
2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Sharma K, Ramachandrarao S, Qiu G, Usui
HK, Zhu Y, Dunn SR, Ouedraogo R, Hough K, McCue P, Chan L, et al:
Adiponectin regulates albuminuria and podocyte function in mice. J
Clin Invest. 118:1645–1656. 2008.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zheng B and Cantley LC: Regulation of
epithelial tight junction assembly and disassembly by AMP-activated
protein kinase. Proc Natl Acad Sci USA. 104:819–822.
2007.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chen X, Guo Y, Jia G, Zhao H, Liu G and
Huang Z: Ferulic acid regulates muscle fiber type formation through
the Sirt1/AMPK signaling pathway. Food Funct. 10:259–265.
2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Giovannini L and Bianchi S: Role of
nutraceutical SIRT1 modulators in AMPK and mTOR pathway: Evidence
of a synergistic effect. Nutrition. 34:82–96. 2017.PubMed/NCBI View Article : Google Scholar
|