Introduction
Radiotherapy has been widely used for the treatment
of several types of cancer, including prostate cancer (1), gynecological cancer (2), lung cancer (3) and pituitary adenomas (4). It is estimated that 80% of cancers may
regress following radiation treatment (5). However, radiotherapy may result in a
series of long-term systemic and local side effects, such as
secondary intracranial tumors (6),
pituitary dysfunction (7) and
stroke (8). When radiotherapy is
used to treat pituitary adenomas, it may increase the risk of
ischemic stroke, and neuroinflammation is the main pathological
process. Therefore, it is crucial to prevent and/or reduce the
impact of such complications on patients.
Astrocytes, the most widely distributed cells in the
central nervous system (CNS), play key roles in providing
structural support for nerve cells, and participate in the
formation of the blood-brain barrier (9). Doron et al (10) confirmed that inflammatory activation
of astrocytes promoted melanoma brain tropism via the C-X-C motif
chemokine ligand 10/C-X-C motif chemokine receptor 3 signaling
axis. However, Tsunemi et al (11) reported that astrocytes protected
human dopaminergic neurons from α-synuclein accumulation and
propagation (11). Therefore,
astrocytes may act as a double-edged sword in disease, and it is
crucial to identify effective therapies for reducing the harmful
effects and enhancing the neuroprotective function of
astrocytes.
Connexin (Cx) is widely distributed in the body,
such as in astrocytes and hepatocytes; however, the distribution of
Cx varies among different cell types (12,13).
Cx43 is the main gap junction protein in cardiomyocytes and is also
the main Cx protein found between ventricular myocytes (14). Abnormal expression levels of Cx43
have been associated with tumor occurrence and development.
Previous reports have confirmed that the decreased level or lack of
Cx43 expression may result in abnormal intercellular communication,
weakened ability of the body to monitor and regulate cells, and
excessive clonal cell growth (15).
In addition, Cx43 plays an important role in neuroinflammation,
including promoting the assembly of gap junctions and increasing
the intercellular signal exchange (16-18).
Thus, elucidating the association between the immune and nervous
systems may be important for preventing CNS disease.
MicroRNA(miR)-206 has been shown to serve an
important role in a variety of tumor types, including glioma
(19-21).
Duan et al (22) reported
that miR-206 modulated lipopolysaccharide-mediated inflammatory
cytokine production in human astrocytes. Furthermore, miR-206 has
been found to alleviate the sevoflurane-induced inhibition of
hippocampal astrocyte activation in aged rats, by targeting IGT-1
via suppression of the PI3K/AKT/CREB signaling pathway (23). However, the role of miR-206 in
irradiation-induced neuroinflammation remains unknown. Notably,
Cx43 was found to be a direct target gene of miR-206(24). Therefore, we hypothesized that
miR-206 may play an important role in irradiation-induced
neuroinflammation by regulating Cx43 expression.
Therefore, the present study was designed to
investigate the roles of microRNA (miR)-206/Cx43 in an
irradiation-induced neuroinflammation cell model.
Materials and methods
Cell culture and inflammation model
establishment
The HA-1800 normal astrocyte cell line was purchased
from Shang Hai Ze Ye Biotechnology Co., Ltd. (cat. no. AC340443;
http://www.shzysw.cn/) and cultured in DMEM
(Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Invitrogen; Thermo fisher Scientific, Inc.) and 1%
penicillin-streptomycin, and maintained at 37˚C in a humidified
incubator with 5% CO2. The HA-1800 cells were treated
with 20 Gy γ-radiation for 12, 24 and 48 h to establish the in
vitro inflammation model (25).
Cell transfection
A total of 100 nM mimics control (sense,
5'-UUCUCCGAACGUGUCACGUTT-3' and antisense, 5'-ACG
UGACACGUUCGGAGAATT-3'; GenePharma Co., Ltd.), 100 nM miR-206 mimics
(sense, 5'-UGGAAGUAAGGAAGU GUG UGG-3' and antisense,
5'-ACACACUUCCUUACAUUCC AUU-3'; GenePharma Co., Ltd.), 1 µg
Cx43-plasmid (cat. no. sc-400241-ACT; Santa Cruz Biotechnology,
Inc.) and 1 µg Control CRISPR Activation Plasmid (control-plasmid;
cat. no. sc-437275; Santa Cruz Biotechnology, Inc.) were
transfected into the HA-1800 cells using Lipofectamine®
2000 (Thermo Fisher Scientific, Inc.) at 37˚C for 24 h, following
the manufacturer's protocol. Reverse transcription-quantitative PCR
(RT-qPCR) and western blot analysis were used to detect the
expression levels of the related genes and proteins, respectively.
Subsequent experiments were performed 24 h after transfection.
Dual-luciferase reporter assay
TargetScan bioinformatics software (http://www.targetscan.org/vert_72/) was used to
predict the binding sites between miR-206 and Cx43. The
3'-untranslated region of Cx43, which contains the miR-206 binding
site or mutated target site, were synthesized using genomic PCR and
cloned into pMIR vectors (Ambion; Thermo Fisher Scientific, Inc.)
to construct the reporter vector Cx43 wild-type (Cx43-WT) or Cx43
mutated-type (Cx43-MUT). The 293 cells (American Type Culture
Collection) were transfected with Cx43 WT or MUT combined with
miR-206 mimics or mimics control using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) and incubated for
48 h, according to the manufacturer's protocol. A Dual-Luciferase
Reporter Assay system (Promega Corporation) was used to assess the
luciferase activity, and the data were normalized to Renilla
luciferase activity.
MTT assay
HA-1800 cells were treated with 20 Gy γ-radiation
for 12, 24 and 48 h, or transfected with mimic control, miR-206
mimic, miR-206 mimic+control-plasmid, or miR-206 mimic+Cx43-plasmid
for 24 h and then treated with 20 Gy γ-radiation for 24 h. Then,
the HA-1800 cells (104 cells per well) were cultured in
96-well plates (BD Bioscience) and incubated for 24 h at 37˚C.
Then, 10 µl MTT (5 mg/ml) solution was added to the cells and
continuously incubated for a further 4 h. Subsequently, the
solution was removed and 100 µl DMSO was added to dissolve the
formazan product. Finally, the optical density at 570 nm was
assessed using a multifunctional plate reader (BioTek Instruments,
Inc.), after 15 min of vibration mixing according to the
manufacturer's protocol.
Flow cytometry analysis
HA-1800 cells were treated as aforementioned, then
harvested by trypsinization and centrifugation (1,000 x g, 5 min,
4˚C). HA-1800 cell apoptosis was determined using an Annexin
V-FITC/PI apoptosis detection kit (BD Biosciences), according to
the manufacturer's protocol. Finally, the apoptotic cells were
quantified using a BD FACSCalibur flow cytometer (Becton-Dickinson
and Company) and analyzed using CellQuest software (version 5.1; BD
Biosciences).
Western blot analysis
Total protein was extracted from the HA-1800 cells
with RIPA lysis buffer (Beyotime Institute of Biotechnology) and
the concentration was determined using a BCA Protein Assay kit
(Invitrogen; Thermo Fisher Scientific, Inc.). Then, equal amounts
of protein (40 µg/lane) were separated using 10% SDS-PAGE and
transferred to a PVDF membrane. After blocking with 5% skimmed milk
in PBS-0.1% Tween-20 at room temperature for 1.5 h, the membranes
were incubated with primary antibodies against β-actin (cat. no.
ab8227; dilution, 1:1,000; Abcam), cleaved caspase-3 (cat. no.
ab32042; dilution, 1:1,000; Abcam), caspase-3 (cat. no. ab32351;
dilution, 1:1,000; Abcam) and Cx43 (cat. no. ab235585; dilution,
1:1,000; Abcam) overnight at 4˚C. Subsequently, the membranes were
washed and incubated with an anti-rabbit IgG horseradish
peroxidase-linked antibody (cat. no. 7074; dilution, 1:2,000; Cell
Signaling Technology, Inc.) at room temperature for 1.5 h. Finally,
the proteins were assessed using an ECL detection system
(MilliporeSigma) in accordance with the manufacturer's
instructions.
ELISA
Inflammatory factors (TNF-α, cat. no. 430201; IL-β,
cat. no. 437007; IL-6, cat. no. 430507; and IFN-γ, cat. no. 430107)
were evaluated using ELISA kits (BioLegend, Inc.) according to the
manufacturer's instructions. After the HA-1800 cells were sonicated
in 2% sulfuric acid, followed by three freeze/thaw cycles, they
then centrifuged for 10 min at 4˚C at 10,000 x g. Subsequently, the
concentrations of TNF-α, IL-β, IL-6 and IFN-γ in the supernatant
were detected using ELISA. The OD of each well at 450 nm was
measured using a microplate reader (Bio-Tek Instruments, Inc.), in
accordance with the manufacturer's instructions.
RT-qPCR analysis
Following treatment, total RNA was extracted from
the HA-1800 cells using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) in accordance with the
manufacturer's instructions. miScript Reverse Transcription kit
(Qiagen, Inc.) was used to transcribe total RNA into cDNA. The
reverse transcription reaction conditions were as following: 25˚C
for 5 min, 42˚C for 60 min and 80˚C for 2 min. An ABI 7000
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) with the SYBR-Green PCR Master Mix kit (Takara Biotechnology
Co., Ltd.) was used to examine the mRNA expression levels of
miR-206, Cx43 and GAPDH. The amplification conditions were as
follows: Pre-denaturation at 95˚C for 10 min; followed by 37 cycles
of denaturation at 95˚C for 10 sec, annealing at 60˚C for 20 sec
and extension at 72˚C for 34 sec. Primers were purchased from
Sangon Biotech Co., Ltd. The primer sequences were as follows:
GAPDH, forward, 5'-CTTTGGTATCGTGGAAGGACTC-3' and reverse, 5'-GTA
GAGGCAGGGATGATGTTCT-3'; Cx43, forward, 5'- TGCTTG
GGATAGCTGGGCGGA-3' and reverse, 5'-TGGGGGCAGA GAGAGAAAGCCC-3'; U6,
forward, 5'-GCT TCGGCAGCACA TATACTAAAAT-3' and reverse,
5'-CGCTTCACGAATTTG CGTGTCAT-3'; and miR-206, forward,
5'-GGCGGTGGAAT GTAAGGAAG-3' and reverse, 5'-GGCTGTCGTGGACT GCG-3'.
Target gene expression level was determined using the
2-ΔΔCq method (26).
Statistical analysis
Statistical analysis was performed using SPSS v20.0
(IBM Corp). All the results are presented as the mean ± SD from
three independent experiments. The mean differences among groups
were calculated using either one-way ANOVA followed by Tukey's post
hoc tests or unpaired Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
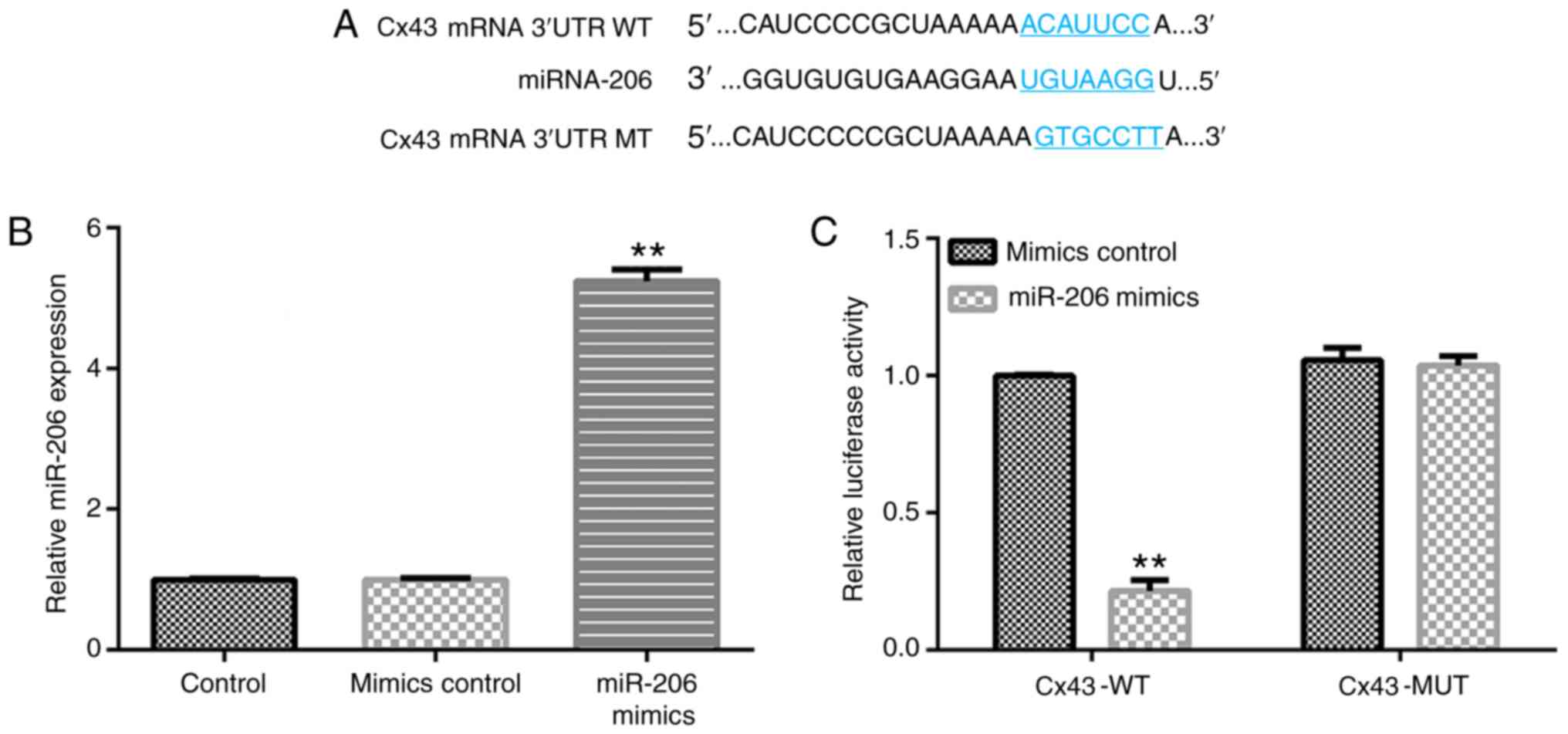
Cx43 directly interacts with
miR-206
First, the association between Cx43 and miR-206 was
investigated. As presented in Fig.
1A, Cx43 was a potential target of miR-206. RT-qPCR analysis
confirmed that the miR-206 mRNA expression was significantly higher
in 293 cells transfected with miR-206 mimics compared with that in
the mimics control group (Fig. 1B).
Furthermore, the dual-luciferase reporter system confirmed that
miR-206 mimics notably suppressed Cx43-WT luciferase activity,
while there were no significant changes in Cx43-MUT luciferase
activity (Fig. 1C), suggesting that
the effects of miR-206 on neuroinflammation may be mediated by
targeting Cx43.
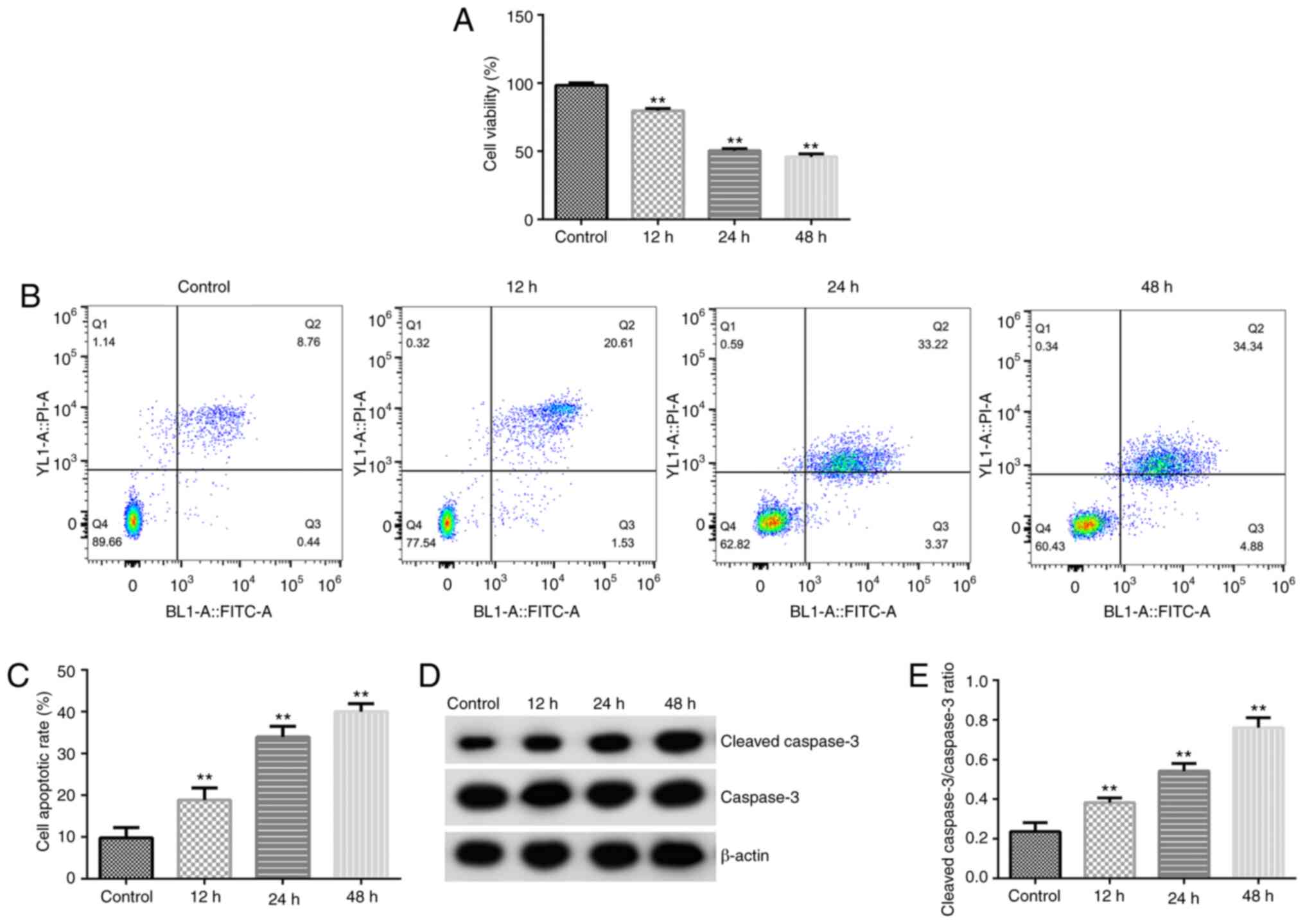
Irradiation suppresses proliferation
and induces apoptosis in the HA-1800 cells
Next, the HA-1800 cells were treated with 20 Gy
γ-radiation for 12, 24 and 48 h to establish an in vitro
inflammatory model. Analysis using MTT assay and flow cytometry
suggested that irradiation significantly inhibited HA-1800 cell
proliferation (Fig. 2A) and
increased the number of apoptotic cells (Fig. 2B and C). In addition, the expression levels of
apoptosis-related proteins were investigated using western blot
analysis. As displayed in Fig. 2D
and E, irradiation markedly
enhanced cleaved caspase-3 protein expression (Fig. 2D) and increased the cleaved
caspase-3/caspase-3 ratio (Fig.
2E). The aforementioned data demonstrated that irradiation
could inhibit the proliferation of HA-1800 cells.
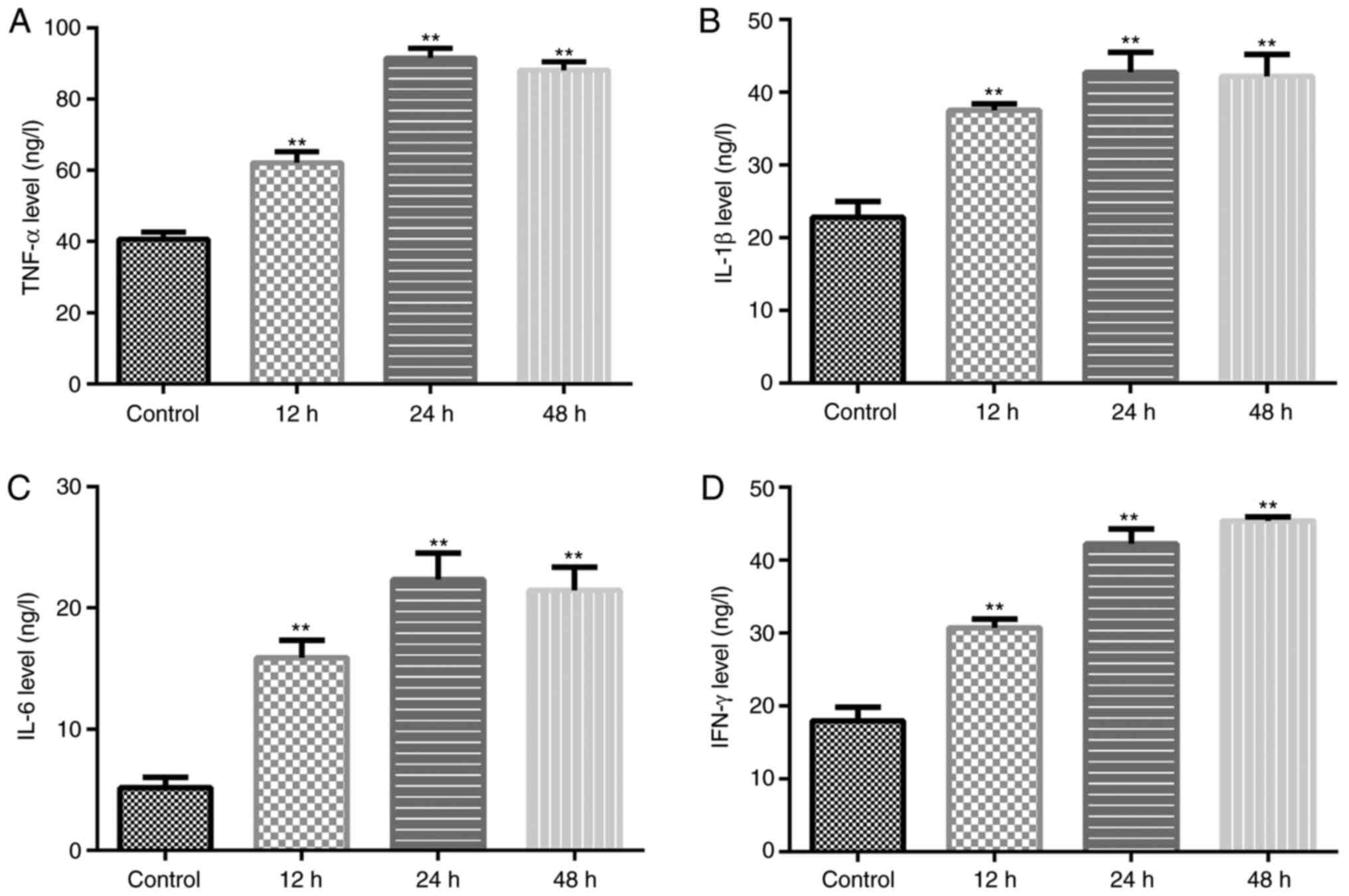
Irradiation promotes inflammatory
factor secretion in HA-1800 cells
Subsequently, the concentration of inflammatory
factors (TNF-α, IL-β, IL-6 and IFN-γ) in the supernatant of the
HA-1800 cells was investigated. Analysis using ELISA revealed that
irradiation markedly elevated the secretion of TNF-α, IL-β, IL-6
and IFN-γ (Fig. 3A-D) compared with
the control group. These findings demonstrated that the
inflammatory model in the HA-1800 cells induced by γ-radiation was
successfully established, and that γ-radiation may promote the
detrimental effects of astrocytes.
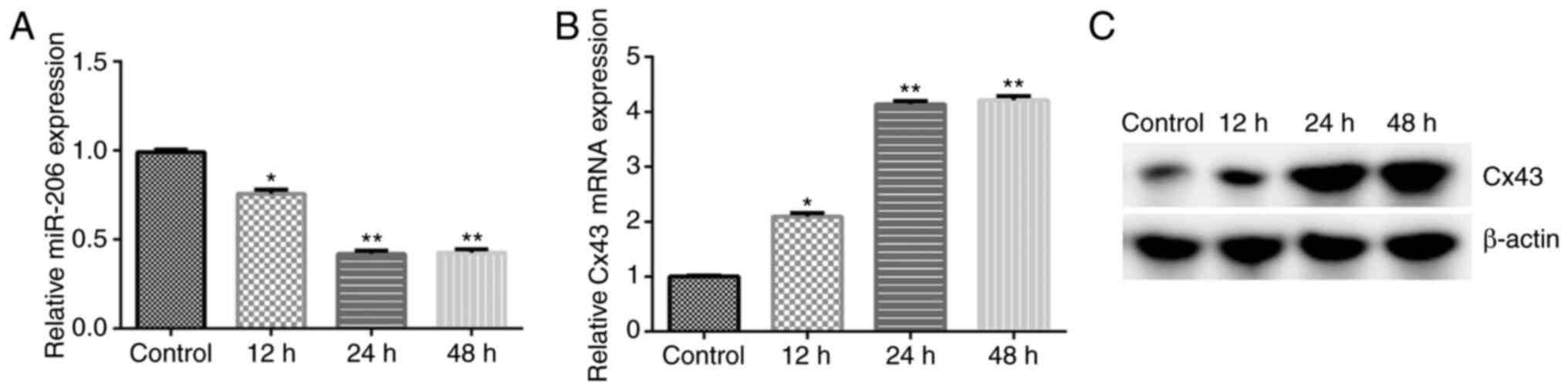
miR-206 expression is notably
downregulated and Cx43 expression is upregulated in
irradiation-induced HA-1800 cells
Subsequently, the mRNA and protein expression levels
of miR-206 and Cx43 were investigated in irradiation-induced
HA-1800 cells using RT-qPCR and western blot analyses,
respectively. It was found that the expression level of miR-206 was
significantly downregulated (Fig.
4A) and the expression level of Cx43 was notably upregulated in
irradiation-induced HA-1800 cells compared with the control group
(Fig. 4B and C). These findings revealed that the
miR-206/Cx43 axis may play a protective role in irradiation-induced
neuroinflammation.
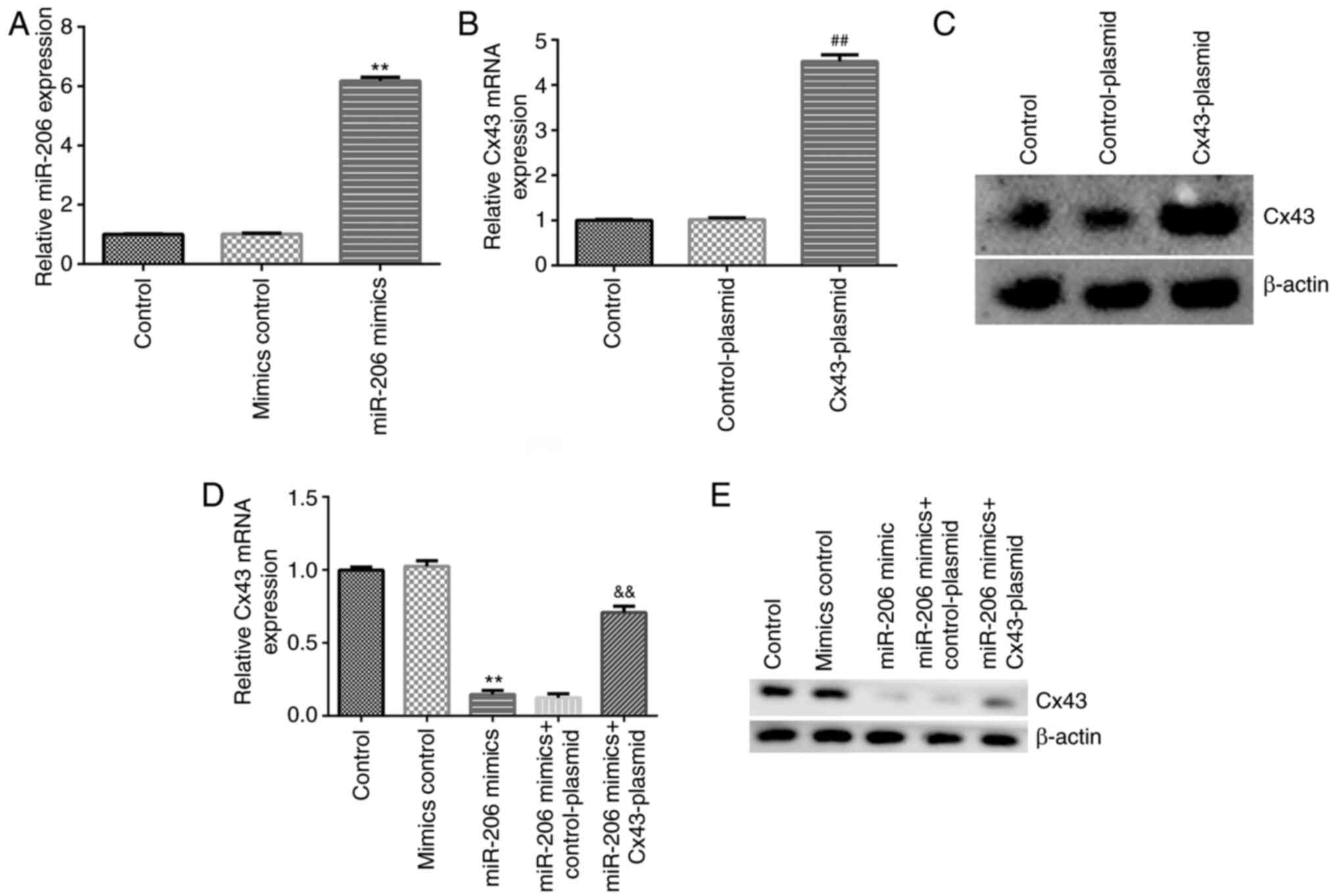
Cx43-plasmid reverses the effect of
miR-206 mimics on Cx43 expression level in HA-1800 cells
To further understand the regulatory association
between miR-206 and Cx43 in HA-1800 cells, mimics control, miR-206
mimics, Cx43-plasmid or control-plasmid were transfected into
HA-1800 cells for 24 h and the transfection efficiency was
determined using RT-qPCR and western blot analyses. As shown in
Fig. 5A, the expression of miR-206
was upregulated in HA-1800 cells transfected miR-206 mimics. In
addition, compared with that in the control-plasmid group, the
Cx43-plasmid markedly enhanced Cx43 expression in HA-1800 cells
(Fig. 5B and C). However, the expression level of Cx43
was lower in HA-1800 cells transfected with miR-206 mimics compared
with that in the mimics control group, and this inhibition was
increased in miR-206 mimics + Cx43-plasmid co-transfected cells
(Fig. 5D and E), indicating that miR-206 could regulate
Cx43 expression level in HA-1800 cells.
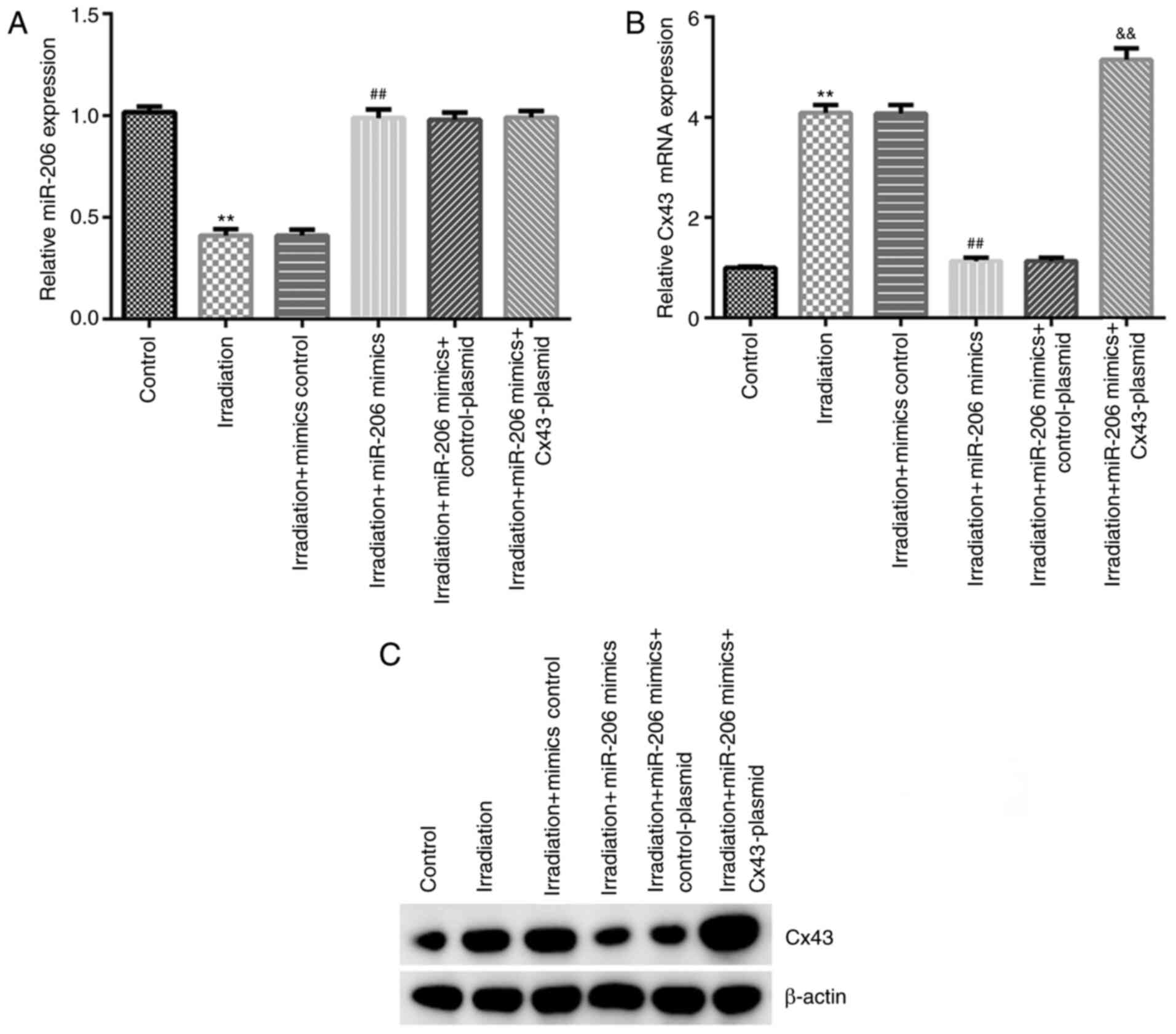
miR-206 negatively regulates the
expression level of Cxn-43 in γ-radiation-induced HA-1800
cells
Previous reports have confirmed that astrocytes play
different roles in the CNS (9,27,28);
therefore, the roles of miR-206 and Cx43 in neuroinflammation were
investigated in the present study. HA-1800 cells were transfected
with mimics control, miR-206 mimics, control-plasmid or
Cx43-plasmid for 24 h, and induced with 20 Gy γ-radiation for
another 24 h. Subsequently, the expression levels of miR-206 and
Cx43 mRNA and protein levels were assessed using RT-qPCR and
western blot analyses, respectively. The results shown in Fig. 6A-C revealed that miR-206 expression
was downregulated and Cx43 was upregulated in γ-radiation-induced
HA-1800 cells compared with the control group, while miR-206 mimics
notably increased miR-206 expression and decreased Cx43 expression.
In addition, there was no change in the miR-206 expression in the
irradiation + miR-206 mimics + Cx43-plasmid group compared with
that in the irradiation + miR-206 mimics + control-plasmid group,
while the expression level of Cx43 was higher in the irradiation +
miR-206 mimics + Cx43-plasmid group compared with that in the
irradiation + miR-206 mimics + control-plasmid group. These data
revealed that miR-206 negatively regulated Cx43 expression in
γ-radiation-induced HA-1800 cells.
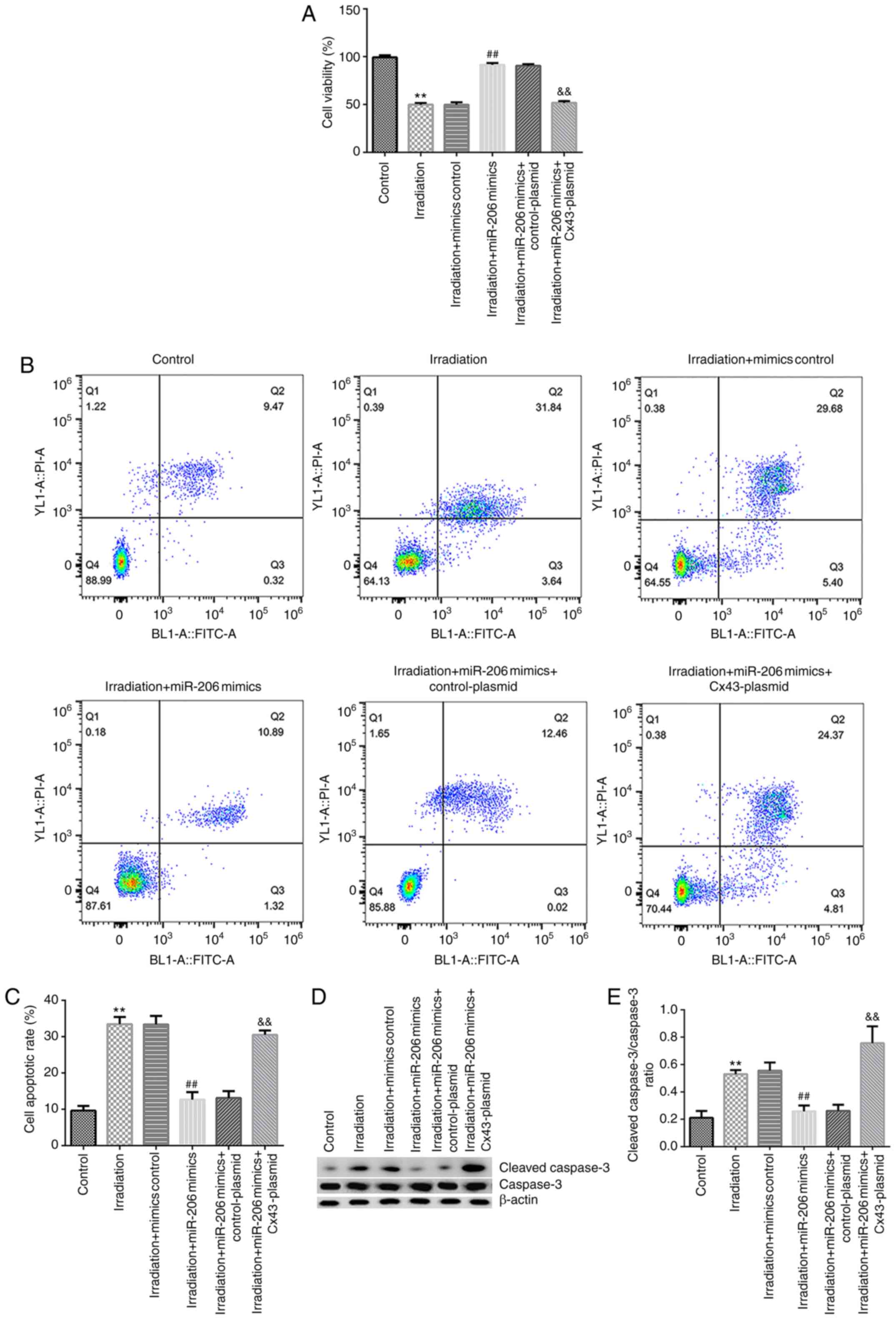
miR-206 alleviates γ-radiation-induced
HA-1800 cell apoptosis by downregulating CX43 expression
To further elucidate the role of miR-206 in
γ-radiation-induced neuroinflammation, HA-1800 cells were
transfected with mimics control, miR-206 mimics, control-plasmid or
Cx43-plasmid for 24 h, then induced with 20 Gy γ-radiation for
another 24 h. Subsequently, the effects of miR-206 mimics on cell
viability and apoptosis were investigated. The results of the MTT
assay and flow cytometry suggested that γ-radiation markedly
inhibited HA-1800 cell viability (Fig.
7A) and increased the number of apoptotic cells (Fig. 7B and C).
Cell apoptosis is typically regulated via
apoptosis-specific genes; therefore, the expression level of
apoptosis-associated proteins was determined in the present study.
As shown in Fig. 7D and E, γ-radiation increased cleaved caspase-3
expression (Fig. 7D) and the
cleaved caspase-3/caspase-3 ratio (Fig.
7E) compared with the control group. In addition, the opposite
effects were observed in the HA-1800 cells transfected with miR-206
mimics and induced by irradiation, whereas these effects were
reversed by transfection with Cx43-plasmid, indicating that miR-206
protected the cells against γ-radiation-stimulated
neuroinflammation by targeting Cx43.
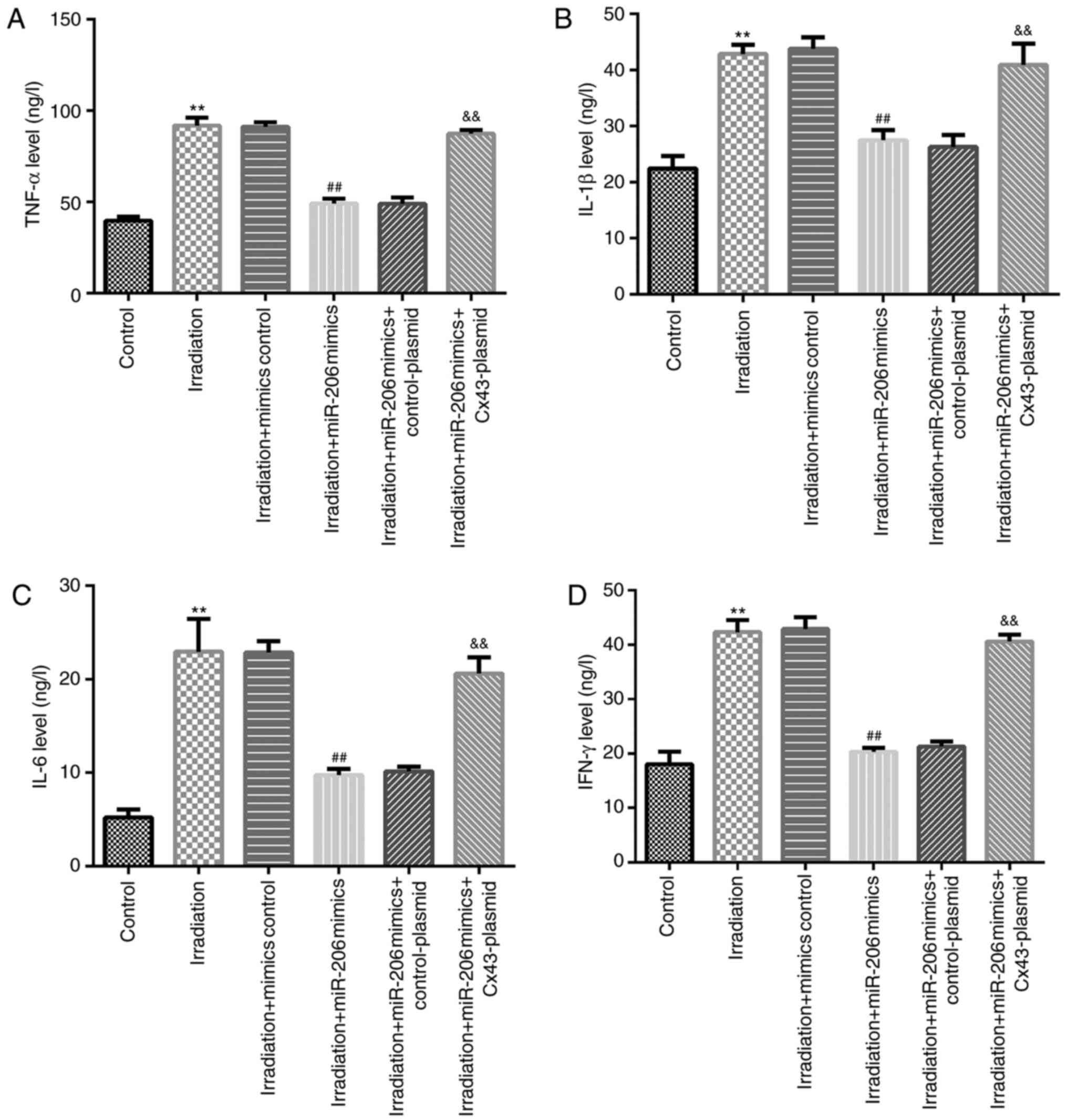
miR-206 attenuates the inflammatory
response in γ-radiation-induced HA-1800 cells by downregulating
Cx43 expression
To investigate the molecular mechanism underlying
the role of miR-206 in γ-radiation-induced HA-1800 cells, the
release of the inflammatory cytokines TNF-α, IL-β, IL-6 and IFN-γ
was investigated using ELISA. As shown in Fig. 8A-D, γ-radiation notably promoted the
release of these inflammatory factors, while γ-radiation-induced
HA-1800 cells transfected with miR-206 mimics secreted lower levels
of inflammatory cytokines compared with those in the irradiation
alone group. However, these findings were abolished following
transfection with Cx43-plasmid. Taken together, the aforementioned
results indicated that miR-206 may reduce the inflammatory response
by negatively regulating Cx43 expression in HA-1800 cells.
Discussion
Radiation therapy has been widely used to treat
several types of cancer, including anal cancer and pituitary
adenomas (29,30). Numerous reports have shown that
radiation therapy could effectively inhibit tumor growth, with
neuroinflammation being the main pathological process. Chang
(31) revealed the effect of
far-infrared radiation therapy on inflammation regulation in
lipopolysaccharide-induced peritonitis in mice. Astrocytes, the
most abundant cell type in the CNS, are functionally and
structurally associated with numerous pathological responses to
disease (32). It has also been
reported that astrocytes aggravated ischemic stroke injury;
however, they also exert neuroprotective effects by releasing
neurotrophic proteins (33,34). At present, certain astrocyte-based
neuroprotective treatment strategies include treatment for
astrocyte protection (35),
astrocyte anti-excitotoxicity and antioxidation (36), as well as astrocyte-derived growth
factors (37).
Cxs are transmembrane proteins, which are
responsible for intercellular communication (38). Cx43, a member of the Cx family, has
been identified as the main component of the gap junction channels
in astrocytes. Furthermore, regulating the opening of the Cx43
hemichannel may protect from tissue injury due to excessive
activation of the inflammatory response (39). Investigations on the association
among neurological diseases, the inflammatory response and Cx
channel disorders are currently at the primary stage. Therefore,
the present study was designed to establish effective therapy
strategies to promote the neuroprotective effects and abolish the
detrimental effects of astrocytes.
It has been reported that the upregulation of Cx43
is crucial for radiation-induced neuroinflammation (25). In addition, Cx43 was found to be a
direct target gene of miR-206(24).
Thus, in the present study, it was hypothesized that miR-206 may
play a protective role in radiation-induced neuroinflammation by
regulating Cx43 expression. First, the TargetScan database was used
and it was found that Cx43 was a potential target of miR-206. To
further investigate the roles of miR-206 in astrocytes, mimics
control or miR-206 mimics were transfected into HA-1800 cells.
RT-qPCR analysis revealed that the miR-206 expression was
significantly higher in HA-1800 cells transfected with miR-206
mimics compared with that in the mimics control group.
Dual-luciferase reporter analysis further confirmed that Cx43
directly interacted with miR-206, indicating that the effects of
miR-206 on inflammation in astrocytes may be mediated by targeting
Cx43.
Astrocytes may exert both harmful and beneficial
effects (40). In the present
study, the type of astrocyte function induced by γ-radiation was
investigated. The HA-1800 cell line was used as a model and cells
were induced with 20 Gy γ-radiation for 12, 24 and 48 h to generate
an in vitro inflammatory model. Then, the effects of
γ-radiation on HA-1800 cell viability, apoptosis and inflammatory
factor release were analyzed. The results indicated that
γ-radiation notably suppressed HA-1800 cell viability, increased
the number of apoptotic cells and upregulated release of TNF-α,
IL-β, IL-6 and IFN-γ. In addition, the expression of the
apoptosis-related proteins, cleaved caspase-3 and caspase-3, was
also examined in irradiation-induced HA-1800 cells and the results
demonstrated that cleaved caspase-3 protein expression and the
cleaved caspase-3/caspase-3 ratio were increased following
irradiation. All these findings demonstrated that γ-radiation may
promote the detrimental effects of astrocytes. Furthermore, the
expression levels of miR-206 and Cx43 in irradiation-treated
HA-1800 cells were analyzed. miR-206 was notably downregulated in
irradiation-induced HA-1800 cells and the expression level of Cx43
was markedly higher in response to irradiation, compared with that
in the control group.
Yang et al (41) reported that inhibition of Cx43 and
phosphorylated-nuclear receptor subfamily 2 group B member 1 in
spinal astrocytes impaired bone cancer pain in mice. Subsequently,
it was investigated whether controlling the expression level of
Cx43 in astrocytes could eliminate the detrimental effects of
irradiation. miR-206 and Cx43 were overexpressed in HA-1800 cells
using miR-206 mimics and Cx43-plasmid, respectively, and
transfection efficacy was confirmed using RT-qPCR and western blot
analyses. Both Cx43 mRNA and protein expression levels were found
to be increased in the Cx43-plasmid transfected cells, while
miR-206 mimics increased miR-206 expression and reduced Cx43
expression, indicating that miR-206 regulated Cx43 expression in
the HA-1800 cells. Research has confirmed that astrocytes play
vital roles in the CNS (42);
therefore, the roles of miR-206 and Cx43 in γ-radiation-induced
HA-1800 cells was investigated. HA-1800 cells were transfected with
mimics control, miR-206 mimics, control-plasmid or Cx43-plasmid for
24 h, then induced with 20 Gy γ-radiation for another 24 h. The
expression levels of Cx43 were decreased in cells transfected with
miR-206 mimics, which indicated that miR-206 could abolish the
upregulated Cx43 expression stimulated by γ-radiation. Furthermore,
the expression level of Cx43 was higher in the irradiation +
miR-206 mimics + Cx43-plasmid group compared with that in the
irradiation + miR-206 mimics + control-plasmid group, whereas the
miR-206 expression level in the irradiation + miR-206 mimics +
Cx43-plasmid group did not change, suggesting that miR-206
negatively regulated Cx43 expression in γ-radiation-induced HA-1800
cells.
To further elucidate the regulatory functions of
miR-206 and Cx43 in γ-radiation-induced HA-1800 cells, the
viability and apoptosis of astrocytes following irradiation
treatment was investigated. It was found that miR-206 mimics
relieved irradiation-induced neuroinflammation, which was confirmed
by increased cell proliferation, inhibition of apoptosis, and
reduced cleaved caspase-3 expression and inflammatory cytokine
secretion. These effects were also found to be reversed by
Cx43-plasmid, demonstrating that miR-206 protected against
γ-radiation-stimulated neuroinflammation by regulating Cx43
expression. Radiotherapy may cause several long-term complications,
such as secondary intracranial tumors, hypopituitarism and stroke
(6-8).
Although radiation therapy is one of the causes of ischemic stroke
among patients with pituitary adenoma, the underlying mechanism
remains unclear. The findings of the present study indicated that
the miR-206/Cx43 axis may be involved in the occurrence and
development of ischemic stroke induced by radiotherapy in patients
with pituitary adenoma, providing a theoretical basis for the
clinical treatment of such patients. Moreover, miR-206 may serve as
a novel potential diagnostic biomarker for ischemic stroke among
patients with pituitary adenoma, as this can be measured in the
blood or cerebrospinal fluid, which will be further investigated in
our future research.
However, this study was only a preliminary in
vitro study on the role of miR-206 in irradiation-induced
neuroinflammation. In order to further verify the role of miR-206
in irradiation-induced neuroinflammation, certain issues must be
further explored in depth. For example, it is unclear whether only
γ-ray-treated astrocytes can accurately mimic the conditions in the
CNS. Furthermore, the interaction between astrocytes and the
surrounding microenvironment under the effect of γ-rays must be
further examined. In addition, the role of miR-206 in
irradiation-induced neuroinflammation should be investigated using
animal models. We plan to address these issues in future
studies.
In summary, the results of the present study
revealed that miR-206 relieved irradiation-induced neural injury by
regulating Cx43 expression in astrocytes, suggesting that the
miR-206/Cx43 axis may serve as a novel potential diagnostic
biomarker for the clinical treatment of inflammation-associated
neural injury in radiation-induced diseases.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WZ contributed to the conception and design of the
study, data acquisition, analysis and interpretation, and also
drafted and critically revised the manuscript. LF contributed to
data collection and statistical analysis. HX contributed to data
collection, statistical analysis and manuscript preparation. WZ and
HX confirmed the authenticity of all the raw data. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wisniewski T, Winiecki J, Makarewicz R and
Zekanowska E: The effect of radiotherapy and hormone therapy on
osteopontin concentrations in prostate cancer patients. J BUON.
25:527–530. 2020.PubMed/NCBI
|
|
2
|
Alcântara-Silva TR, de Freitas-Junior R,
Freitas NM, de Paula Junior W, da Silva DJ, Machado GD, Ribeiro MK,
Carneiro JP and Soares LR: Music therapy reduces
radiotherapy-induced fatigue in patients with breast or
gynecological cancer: A Randomized Trial. Integr Cancer Ther.
17:628–635. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Agrawal V, Benjamin KT and Ko EC:
Radiotherapy and Immunotherapy Combinations for Lung Cancer. Curr
Oncol Rep. 23(4)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Brada M and Jankowska P: Radiotherapy for
pituitary adenomas. Endocrinol Metab Clin North Am. 37:263–275.
2008.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Dahl O, Dale JE and Brydøy M: Rationale
for combination of radiation therapy and immune checkpoint blockers
to improve cancer treatment. Acta Oncol. 58:9–20. 2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Yamanaka R, Abe E, Sato T, Hayano A and
Takashima Y: Secondary intracranial tumors following radiotherapy
for pituitary adenomas: A systematic review. Cancers (Basel).
9(E103)2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Xiang B, Zhu X, He M, Wu W, Pang H, Zhang
Z, Yang Y, Li Y, Wang Y, Wang Y, et al: Pituitary dysfunction in
patients with intracranial germ cell tumors treated with
radiotherapy. Endocr Pract. 26:1458–1468. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kuan FC, Lee KD, Huang SF, Chen PT, Huang
CE, Wang TY and Chen MC: Radiotherapy is associated with an
accelerated risk of ischemic stroke in oral cavity cancer survivors
after primary surgery. Cancers (Basel). 12(E616)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Schultz C, Dehghani F, Hubbard GB, Thal
DR, Struckhoff G, Braak E and Braak H: Filamentous tau pathology in
nerve cells, astrocytes, and oligodendrocytes of aged baboons. J
Neuropathol Exp Neurol. 59:39–52. 2000.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Doron H, Amer M, Ershaid N, Blazquez R,
Shani O, Lahav TG, Cohen N, Adler O, Hakim Z, Pozzi S, et al:
Inflammatory activation of astrocytes facilitates melanoma brain
tropism via the CXCL10-CXCR3 signaling axis. Cell Rep.
28:1785–1798.e6. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Tsunemi T, Ishiguro Y, Yoroisaka A, Valdez
C, Miyamoto K, Ishikawa K, Saiki S, Akamatsu W, Hattori N and
Krainc D: Astrocytes protect human dopaminergic neurons from
alpha-synuclein accumulation and propagation. J Neurosci.
40:8618–8628. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Nielsen BS, Hansen DB, Ransom BR, Nielsen
MS and MacAulay N: Connexin hemichannels in astrocytes: An
assessment of controversies regarding their functional
characteristics. Neurochem Res. 42:2537–2550. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Pei H, Zhai C, Li H, Yan F, Qin J, Yuan H,
Zhang R, Wang S, Zhang W, Chang M, et al: Connexin 32 and connexin
43 are involved in lineage restriction of hepatic progenitor cells
to hepatocytes. Stem Cell Res Ther. 8(252)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Biendarra-Tiegs SM, Clemens DJ, Secreto FJ
and Nelson TJ: Human induced pluripotent stem cell-derived
non-cardiomyocytes modulate cardiac electrophysiological maturation
through Connexin 43-mediated cell-cell interactions. Stem Cells
Dev. 29:75–89. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ma JW, Ji DD, Li QQ, Zhang T and Luo L:
Inhibition of connexin 43 attenuates oxidative stress and apoptosis
in human umbilical vein endothelial cells. BMC Pulm Med.
20(19)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wang X, Feng L, Xin M, Hao Y, Wang X,
Shang P, Zhao M, Hou S, Zhang Y, Xiao Y, et al: Mechanisms
underlying astrocytic connexin-43 autophagy degradation during
cerebral ischemia injury and the effect on neuroinflammation and
cell apoptosis. Biomed Pharmacother. 127(110125)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yin X, Feng L, Ma D, Yin P, Wang X, Hou S,
Hao Y, Zhang J, Xin M and Feng J: Roles of astrocytic connexin-43,
hemichannels, and gap junctions in oxygen-glucose
deprivation/reperfusion injury induced neuroinflammation and the
possible regulatory mechanisms of salvianolic acid B and
carbenoxolone. J Neuroinflammation. 15(97)2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Vignal N, Boulay AC, San C, Cohen-Salmon
M, Rizzo-Padoin N, Sarda-Mantel L, Declèves X, Cisternino S and
Hosten B: Astroglial Connexin 43 deficiency protects against
LPS-induced neuroinflammation: A TSPO Brain µPET Study with
[18F]FEPPA. Cells. 9(389)2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhou F, Cao W, Xu R, Zhang J, Yu T, Xu X,
Zhi T, Yin J, Cao S, Liu N, et al: MicroRNA-206 attenuates glioma
cell proliferation, migration, and invasion by blocking the
WNT/β-catenin pathway via direct targeting of Frizzled 7 mRNA. Am J
Transl Res. 11:4584–4601. 2019.PubMed/NCBI
|
|
20
|
Jiao D, Chen J, Li Y, Tang X, Wang J, Xu
W, Song J, Li Y, Tao H and Chen Q: miR-1-3p and miR-206 sensitizes
HGF-induced gefitinib-resistant human lung cancer cells through
inhibition of c-Met signalling and EMT. J Cell Mol Med.
22:3526–3536. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Liu C, Li J, Wang W, Zhong X, Xu F and Lu
J: miR-206 inhibits liver cancer stem cell expansion by regulating
EGFR expression. Cell Cycle. 19:1077–1088. 2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Duan X, Zohaib A, Li Y, Zhu B, Ye J, Wan
S, Xu Q, Song Y, Chen H and Cao S: miR-206 modulates
lipopolysaccharide-mediated inflammatory cytokine production in
human astrocytes. Cell Signal. 27:61–68. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Liu TJ, Wang B, Li QX, Dong XL, Han XL and
Zhang SB: Effects of microRNA-206 and its target gene IGF-1 on
sevoflurane-induced activation of hippocampal astrocytes in aged
rats through the PI3K/AKT/CREB signaling pathway. J Cell Physiol.
233:4294–4306. 2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Li H, Xiang Y, Fan LJ, Zhang XY, Li JP, Yu
CX, Bao LY, Cao DS, Xing WB, Liao XH, et al: Myocardin inhibited
the gap protein connexin 43 via promoted miR-206 to regulate
vascular smooth muscle cell phenotypic switch. Gene. 616:22–30.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Chen W, Tong W, Guo Y, He B, Chen L, Yang
W, Wu C, Ren D, Zheng P and Feng J: Up-regulation of Connexin-43 is
critical for irradiation-induced neuroinflammation. CNS Neurol
Disord Drug Targets. 17:539–546. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-ΔΔC(T)) method. Methods. 25:402–408. 2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Nutma E, van Gent D, Amor S and Peferoen
LAN: Astrocyte and oligodendrocyte cross-talk in the central
nervous system. Cells. 9(600)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sofroniew MV and Vinters HV: Astrocytes:
Biology and pathology. Acta Neuropathol. 119:7–35. 2010.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pawlowski J and Jones WE III: Radiation
Therapy For Anal Cancer. StatPearls Publishing, Treasure Island,
FL, 2021.
|
|
30
|
Wormhoudt TL, Boss MK, Lunn K, Griffin L,
Leary D, Dowers K, Rao S and LaRue SM: Stereotactic radiation
therapy for the treatment of functional pituitary adenomas
associated with feline acromegaly. J Vet Intern Med. 32:1383–1391.
2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chang Y: The effect of far infrared
radiation therapy on inflammation regulation in
lipopolysaccharide-induced peritonitis in mice. SAGE Open Med: Sep
10, 2018 (Epub ahead of print). doi: 10.1177/2050312118798941.
|
|
32
|
Li S, Zhou C, Zhu Y, Chao Z, Sheng Z,
Zhang Y and Zhao Y: Ferrostatin-1 alleviates angiotensin II (Ang
II)- induced inflammation and ferroptosis in astrocytes. Int
Immunopharmacol. 90(107179)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Sun JB, Li Y, Cai YF, Huang Y, Liu S,
Yeung PK, Deng MZ, Sun GS, Zilundu PL, Hu QS, et al: Scutellarin
protects oxygen/glucose-deprived astrocytes and reduces focal
cerebral ischemic injury. Neural Regen Res. 13:1396–1407.
2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ishihara Y, Itoh K, Oguro A, Chiba Y, Ueno
M, Tsuji M, Vogel CF and Yamazaki T: Neuroprotective activation of
astrocytes by methylmercury exposure in the inferior colliculus.
Sci Rep. 9(13899)2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Cabezas R, Baez-Jurado E, Hidalgo-Lanussa
O, Echeverria V, Ashrad GM, Sahebkar A and Barreto GE: Growth
factors and neuroglobin in astrocyte protection against
neurodegeneration and oxidative stress. Mol Neurobiol.
56:2339–2351. 2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
McBean GJ: Astrocyte antioxidant systems.
Antioxidants (Basel). 7(112)2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Karki P, Smith K, Johnson J Jr and Lee E:
Astrocyte-derived growth factors and estrogen neuroprotection: Role
of transforming growth factor-α in estrogen-induced upregulation of
glutamate transporters in astrocytes. Mol Cell Endocrinol.
389:58–64. 2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Nalewajska M, Marchelek-Myśliwiec M,
Opara-Bajerowicz M, Dziedziejko V and Pawlik A:
Connexins-therapeutic targets in cancers. Int J Mol Sci.
21(E9119)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li T, Niu J, Yu G, Ezan P, Yi C, Wang X,
Koulakoff A, Gao X, Chen X, Sáez JC, et al: Connexin 43 deletion in
astrocytes promotes CNS remyelination by modulating local
inflammation. Glia. 68:1201–1212. 2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Sanchez MC, Benitez A, Ortloff L and Green
LM: Alterations in glutamate uptake in NT2-derived neurons and
astrocytes after exposure to gamma radiation. Radiat Res.
171:41–52. 2009.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Yang H, Yan H, Li X, Liu J, Cao S, Huang
B, Huang D and Wu L: Inhibition of Connexin 43 and phosphorylated
NR2B in spinal astrocytes attenuates bone cancer pain in mice.
Front Cell Neurosci. 12(129)2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Bylicky MA, Mueller GP and Day RM:
Radiation resistance of normal human astrocytes: The role of
non-homologous end joining DNA repair activity. J Radiat Res
(Tokyo). 60:37–50. 2019.PubMed/NCBI View Article : Google Scholar
|