Introduction
Cirrhosis is induced by several types of chronic or
repeated hepatic damages and its incidence is increasing worldwide
(1). Although numerous attempts
have been made to prevent and/or reverse liver fibrosis in the past
decade, liver transplantation still remains the only effective
therapy (2). The effect of
progressive inflammation on hepatic fibrosis has been widely
investigated in previous years, and this not only induces chronic
hepatocyte injury, but also increases the activation and
proliferation of hepatic stellate cells (HSCs) (3). HSCs can transform into myofibroblasts
(MFBs) to synthesize fibers, resulting in hepatic fibrosis and even
cirrhosis (4). In addition, the
epithelial-mesenchymal transition (EMT) of hepatocytes can be
induced by inflammation via the upregulation of growth factors,
particularly TGF, thus resulting in the transformation of
hepatocytes into MFBs and progression of liver fibrosis (5). Although hepatocyte EMT has been
refuted by a series of lineage tracing experiments (6,7), our
previous study demonstrated in vivo that during hepatic
fibrosis the epithelial biomarkers on hepatocytes were lost and the
mesenchymal biomarkers were increased (8), indicating that hepatocytes could be an
important source of fibers during hepatic fibrosis. It has also
been reported that autophagy is involved in liver fibrosis and
facilitates its progress (9,10). On
the other hand, prostaglandins can exacerbate inflammation
(11), whereas cyclooxygenase-2
(COX-2) acts as a key rate-limiting enzyme to catalyze them
(12). The high expression levels
of COX-2 in fibrotic livers indicated that COX-2 could be
considered as a key regulator in accelerating hepatitis (12).
In the present study, thioacetamide (TAA), a classic
drug against liver damage, was used to induce hepatic fibrosis in
rats (13). Subsequently, rats with
hepatic fibrosis were co-treated with celecoxib and octreotide to
investigate their effects on hepatocyte EMT, apoptosis and
autophagy. Additionally, the hepatic fibrosis process was monitored
to evaluate the effects of celecoxib and octreotide on protection
of hepatocytes during fibrosis. Somatostatin (SST) exerts
immunomodulatory, antisecretory and antiproliferative functions via
binding to SST receptors (SSTR1-5), while octreotide is an analogue
of SST (14-16).
The present study hypothesized that the combination of octreotide
with celecoxib could be considered as an efficient approach for
protection of hepatocytes and attenuation of hepatic fibrosis.
Materials and methods
Animal studies
A total of 36 male Sprague-Dawley (SD) rats (age, 12
weeks; weight, 200-230 g) were purchased from West China Medical
Experimental Animal Center, Sichuan University (Chengdu, Sichuan,
China). All animals were housed in a specific pathogen-free
facility (temperature, 25±2˚C; humidity, 50±10%) with a 12/12-h
light/dark cycle and free access to chow and water. The animal
procedures were approved by the Animal Use and Care Committee of
the Sichuan University (approval no. K2015004; Chengdu, Sichuan,
China).
Liver fibrosis model was induced by intraperitoneal
injection (i.p.) of 200 mg/kg TAA (40 mg/ml dissolved in sterile
saline; Sigma-Aldrich; Merck KGaA) every 3 days for 16 weeks
(17). Animals were randomized into
control, TAA and TAA + celecoxib + octreotide (TAA + C) groups,
with 12 animals in each group. Rats in the control group were
treated with sterile saline (1 ml; i.p.) every 3 days as a vehicle
control. Rats in the TAA group were treated with TAA (200 mg/kg
dissolved in sterile saline; i.p.) every 3 days and sterile saline
daily (1 ml/kg; s.c.; twice per day), while those in the TAA + C
group were given a combination of TAA, celecoxib (gastric gavage;
20 mg/kg/day; Pfizer, Inc.) and octreotide (s.c.; 50 µg/kg/day;
Novartis International AG). In the TAA + C group, celecoxib and
octreotide were given to rats together with TAA synchronously from
the beginning of the experiment. During the modeling, one rat died
in each of the TAA and TAA + C groups, while no death was observed
in the control group. All animals were sacrificed by cardiac
exsanguination under general anesthesia with sodium pentobarbital
(30 mg/kg; i.p.).
Morphology detection of liver
tissues
The gross morphology of the liver was examined to
evaluate hepatic fibrosis. Liver tissues were fixed in 4% neutral
buffered paraformaldehyde for 48 h at 4˚C. Tissues were dehydrated
using an ascending alcohol series at 70, 80, 90, 95 and 100%.
Degreasing was carried out using xylene for 45 min at room
temperature. Subsequently, samples were infiltrated in paraffin for
30 min at 65˚C three times. Finally, tissues were embedded in
paraffin wax and sectioned into 4-µm thick sections.
Following dewaxing and rehydration with xylene, and
rehydration with a descending alcohol series, liver slides were
stained with hematoxylin for 5 min at room temperature. Slides were
subsequently washed with acid alcohol (1% hydrochloric acid:70%
alcohol) for 15 sec at room temperature and immerged in ammonia
solution for 10 sec at room temperature. Slides were stained with
1% eosin for 2 min at room temperature. The slides were
subsequently dehydrated using an ascending alcohol series at 90, 95
and 100% for 2 min at room temperature at each stage, followed by
incubation in xylene twice for 3 min each. Sections were sealed
using synthetic resin and observed using a light microscope
(Eclipse Ti2-A; Nikon Corporation).
To evaluate hepatic nodules and fibrotic septa,
sections were stained with Masson's Trichrome and Sirius Red. For
Masson's Trichrome, following deparaffinization and hydration in
water, slides were subsequently treated with Trichrome Stain kit
(cat. no. ab150686; Abcam) Briefly, slides were incubated in
Bouin's Fluid for 60 min at 60˚C followed by a 10-min cooling
period. Slides were washed in water until sections were completely
clear, and slides were stained with working Weigert's Iron
Hematoxylin for 5 min, followed by washing in water for 2 min.
Biebrich Scarlet and Acid Fuchsin solution was added to slides for
15 min, followed by washing in water. Slides were differentiated in
Phosphomolybdic and Phosphotungstic Acid solution for 15 min.
Applied Aniline Blue solution was added to slides for 10 min
followed by washing in distilled water. 1% Applied Acetic Acid
solution was added to slides for 3-5 min, followed by dehydration
in an ascending alcohol series. Slides were incubated in xylene and
sealed in synthetic resin. Subsequently, five images per section
were randomly captured using a light microscope (Eclipse Ti2-A;
Nikon Corporation; magnification, x100) from each liver tissue. The
area of fibers was evaluated using Image-Pro Plus 6.0 software
(CAD/CAM Services, Inc.) and fibrosis was assessed using the Ishak
scoring system (scores, 0-6; higher score denotes more severe
fibrosis) (18). The standards of
Ishak's Score were as follows: 0, no fibrosis; 1, fibrous expansion
of some portal areas, with or without short fibrous septa; 2,
fibrous expansion of most portal areas, with or without short
fibrous septa; 3, fibrous expansion of most portal areas with
occasional portal to portal bridging; 4, fibrous expansion of
portal areas with marked portal to portal bridging as well as
portal to central bridging; 5, marked bridging with occasional
nodules; 6, cirrhosis, probable or definite.
Sirius Red staining was applied to verify the
results of the Masson's Trichrome staining. Following
deparaffinization and hydration in water, slides were subsequently
treated as follows: Stained in 0.1% Sirius red and saturated picric
acid for 10 min at room temperature, followed by washing in water.
Slides were dehydrated using an ascending alcohol series, incubated
in Xylene and sealed in synthetic resin. Finally, images of
sections were randomly captured using a light microscope as
previously described.
For electron microscopy detection, animals were
transcardially perfused with 0.01 M PBS (pH 7.4), anesthetized with
sodium pentobarbital at 30 mg/kg i.p., and transcardially perfused
with 2% paraformaldehyde and 2.5% glutaraldehyde mixed solution.
The next steps differed between distinct types of electron
microscopy. Therefore, for scanning electron microscopy (SEM),
liver tissues were immediately frozen with liquid nitrogen for 20
min at -196˚C, smashed into pieces and fixed in 2.5% glutaraldehyde
overnight at 4˚C. After dehydration using an ascending alcohol
series and sputter coating using gold, the surface ultrastructure
of liver tissues was observed under a SEM system (JSM-7500F; JEOL
Ltd.). For transmission electron microscopy (TEM), liver tissues
were cut into cubes of ~1 mm3 and fixed with 2.5%
glutaraldehyde at 4˚C overnight, and fixed again in 1%
phosphate-buffered osmium tetroxide (0.1 M; pH 7.2) for 1 h at room
temperature. Sections were dehydrated in a graded alcohol solution,
and embedded with Epon 812 resin overnight at 35˚C, 12 h at 45˚C
and 24 h at 60˚C, in sequence. Tissues were cut into 70-nm sections
followed by staining with uranyl acetate for 10 min at room
temperature and lead citrate for 5 min at room temperature.
Sections were observed using a TEM system (H-600IV; Hitachi, Ltd.)
and images were obtained using Gatan Microscopy Suite 64-bit 3.2
(Gatan, Inc.).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from hepatic tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. The purity and
concentration of RNA was measured using the OD at 260 nm and the
260/280 nm ratio was calculated. RNA with a 260/280 ratio between
1.8-2.0 was used for subsequent experiments. cDNA was synthesized
using a cDNA Reverse Transcription kit (Fermentas; Thermo Fisher
Scientific, Inc.) under the following conditions: 42˚C for 60 min,
70˚C for 5 min and 4˚C for 30 min. Subsequently, qPCR was performed
using a SYBR® Green qPCR supermix (Bio-Rad Laboratories,
Inc.) on a CFX96 qPCR system (Bio-Rad Laboratories, Inc.). The
following thermocycling conditions were used: Initial denaturation
at 95˚C for 3 min; followed by 40 cycles of denaturation at 95˚C
for 30 sec, and annealing at 60˚C for 30 sec. The primer sequences
used are listed in Table SI. The
relative mRNA expression levels of the target genes were calculated
using the 2-ΔΔCq method (19) and normalized to that of GAPDH.
Immunohistochemistry (IHC)
assessment
After deparaffinization and rehydration of liver
slides, antigen retrieval was performed using citrate buffer
solution (0.01 M; pH 6.0) in an electric oven ~95˚C for 15 min.
After rinsing with PBS, the slides were incubated in 3%
H2O2 for 10 min at room temperature. The
slides were subsequently blocked with 5% goat serum (cat. no.
ZLI-9021; OriGene Technologies, Inc.) for 15 min at 37˚C. Following
incubation with the primary antibody overnight at 4˚C, a Secondary
Antibody kit was applied according to the manufacturer's
instructions (cat. nos. SP-9001; SP-9002; PV-9003; all purchased
from ZSGB-BIO, Co., Ltd.) Sections were incubated with
biotin-conjugated secondary antibody for 30 min at 37˚C followed by
incubation with HRP-conjugated avidin for 30 min at 37˚C. IHC
reactions were developed using DAB Horseradish Peroxidase Color
Development kit (cat. no. ZLI-9018; ZSGB-BIO, Co., Ltd.), and
nuclei were counterstained with hematoxylin for 3 min at room
temperature. The slides were dehydrated using an ascending alcohol
series at 90, 95 and 100% for 2 min at each stage, followed by
incubation in xylene twice for 3 min each. Finally, tissue sections
were sealed with neutral resins. The images of the sections were
captured using a light microscope as previously described.
The primary antibodies used for IHC analysis were
listed in Table SII. The cell
number of positive PCNA was counted in five randomly captured
images per slide (magnification, x400), while the optical density
(OD) of SST and cleaved caspase-3 was measured using the Image-Pro
Plus v 6.0 analysis software (Media Cybernetics, Inc.). The
relative OD value of the control group was considered to be 1, and
the relative OD values of the other two groups were obtained based
on the ratios of the original OD values of the other two groups to
that of the control group, separately.
Western blot analysis
Liver tissues were homogenized in liquid nitrogen,
and whole proteins were extracted using ice-cold RIPA buffer
(Beyotime Institute of Biotechnology). Protein concentrations were
quantified using a BCA assay (cat. no. PA115-01; Tiangen Biotech
Co, Ltd.). Protein samples were added to 5X loading buffer, heated
in boiling water for 5 min, and then equal amounts of proteins (30
µg) from each sample were resolved by 12 or 15% SDS-PAGE, and
transferred to PVDF membranes (MilliporeSigma). Membranes were
blocked with 5% non-fat powdered milk in TBS (20 mM Tris-HCl pH
7.5, 150 mM NaCl) with 0.1% Tween-20 for 1 h at room temperature.
The membranes were subsequently incubated with primary antibodies
at 4˚C overnight followed by secondary antibody incubation for 1 h
at 37˚C. The targeting proteins were detected by chemiluminescence
using an ECL kit (Beyotime Institute of Biotechnology). Protein
expression levels were normalized to GAPDH and analyzed using
Quantity_One_v462_PC (Bio-Rad Laboratories, Inc.). The antibodies
used for western blot analysis were listed in Table SII.
ELISA for serum prostaglandin E2
(PGE2)
Blood was collected from the right femoral artery of
rats and centrifuged at 1,000 x g for 10 min at room temperature to
obtain blood serum. The concentration of serum PGE2 was quantified
using an ELISA Kit for PGE2 (cat. no. CEA538Ge; Wuhan USCN Business
Co., Ltd.) according to the manufacturer's instructions. Plates
were read using a Thermo microplate reader (Thermo Fisher
Scientific, Inc.) at a wavelength of 450 nm.
TUNEL assay
TUNEL staining was applied using 4-µm thick paraffin
sections of liver tissues, obtained as previously described. This
test was performed using an In Situ Cell Death Detection kit
(cat. no. 11684817910; Roche Diagnostics GmbH) according to the
manufacturer's instructions. Following deparaffinization and
rehydration, the sections incubated in permeabilization solution
containing 0.1% Triton X-100 and 0.1% sodium citrate for 20 min at
room temperature, followed by washing with PBS. Sections were
incubated with 3% hydrogen peroxide in PBS for 5 min to inhibit
endogenous peroxidase activity and rinsed with PBS. Sections were
subsequently incubated with a TUNEL reaction mixture for 1 h at
37˚C, rinsed with PBS, and incubated with converter-POD for 30 min
at 37˚C. Sections were washed with PBS and stained with DAB (cat.
no. ZLI-9018; Origene Technologies, Inc.) and rinsed with PBS.
Nuclei were stained with hematoxylin staining solution for 5 min at
room temperature, dehydrated using an ascending alcohol series and
incubated with xylene. Finally, tissue sections were sealed with
neutral resins. The sections were observed using a light microscope
and the TUNEL-positive cells were counted using Image-Pro Plus v
6.0 analysis software, as previously described.
Statistical analysis
All data were presented as the mean ± standard
deviation. The results were analyzed using SPSS 19.0 software (IBM
Corp.), and one-way ANOVA was applied to compare differences among
multiple groups followed by Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Celecoxib and octreotide mitigate
hepatic inflammation
Compared with those in the control group, the
protein expression levels of COX-2, IL-6 and TNF-α were increased
in the TAA group (Fig. 1E).
However, intrahepatic inflammation was attenuated in the TAA + C
group compared with the TAA group, as demonstrated by the
downregulation of COX-2, IL-6 and TNF-α. In addition, the OD value
of SST in liver tissues and the serum levels of PGE2 in the TAA + C
group were notably decreased compared with those in the TAA group
(Fig. 1C and D). Moreover, the mRNA expression level of
COX-2 was decreased in the TAA + C group compared with the TAA
group, but this result was not significant.
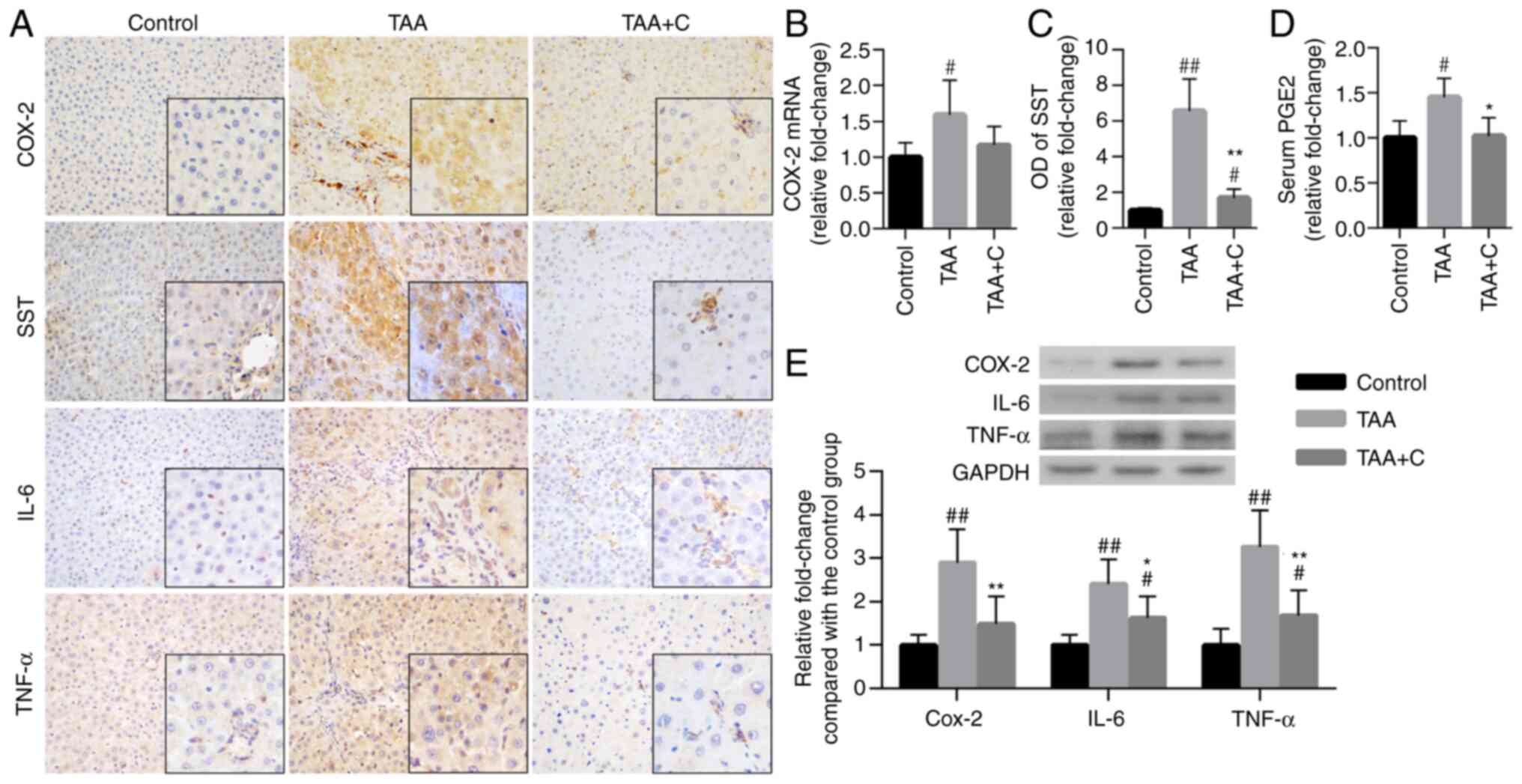 | Figure 1Treatment with celecoxib and
octreotide attenuates hepatic inflammation. (A) Immunohistochemical
staining of COX-2, SST, IL-6 and TNF-α in liver sections
(magnification, x400; insert, x1,000). (B) COX-2 mRNA expression
was measured in hepatic tissues by reverse
transcription-quantitative PCR (n=11/group). (C) OD values of SST
(n=6). (D) Serum secretion levels of PGE2 were detected by ELISA
(n=11). (E) Relative protein expression levels of COX-2, IL-6 and
TNF-α were determined in liver tissues by western blot analysis
(n=6). #P<0.05 and ##P<0.01 vs. control
group; *P<0.05 and **P<0.01 vs. TAA
group. COX-2, cyclooxygenase-2; OD, optical density; PGE2,
prostaglandin E2; SST, somatostatin; TAA, thioacetamide. |
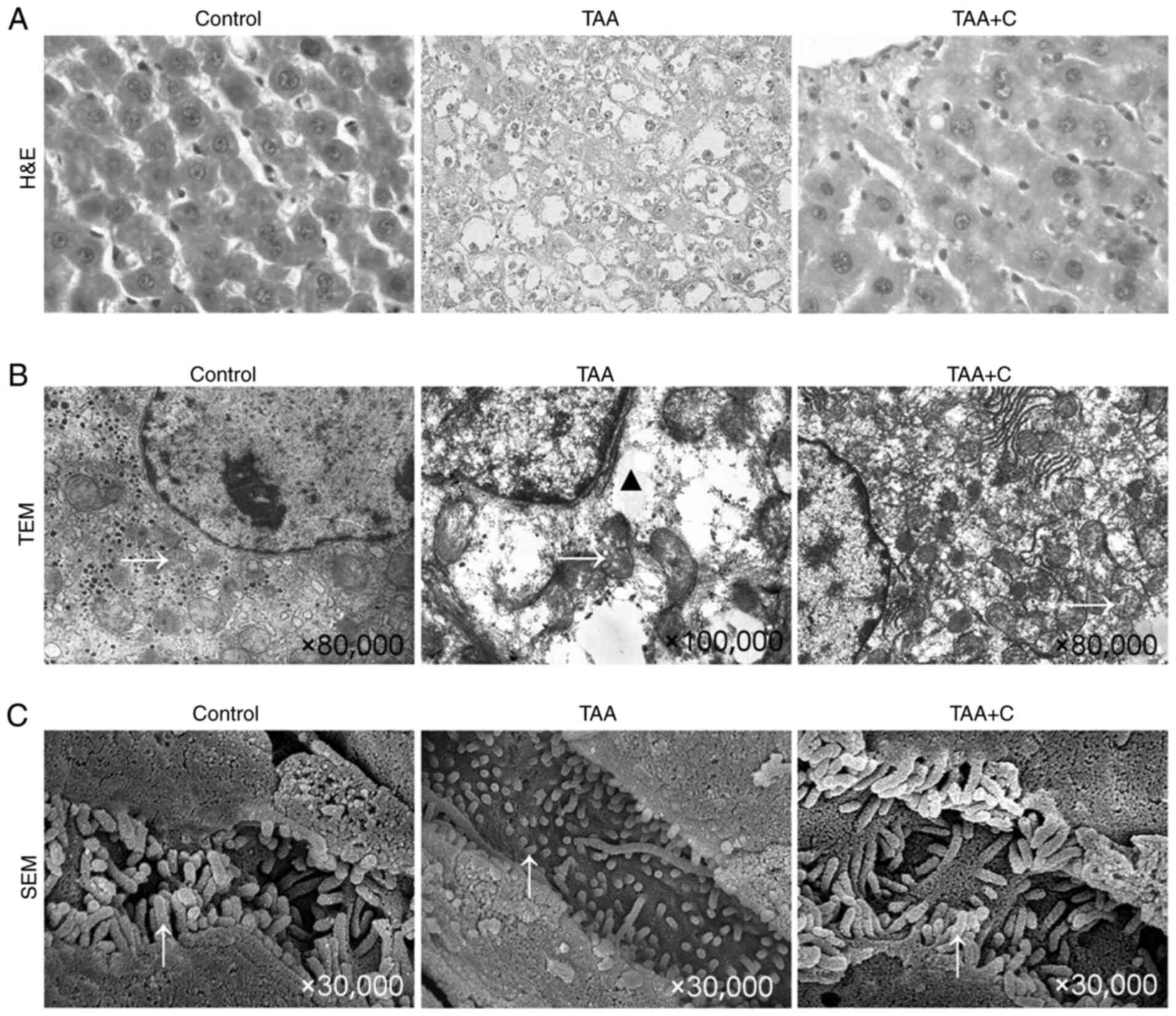
Celecoxib and octreotide protect the
morphology of hepatocytes
Cytoplasmic vacuolization and nucleus pyknosis in
hepatocytes, irregular arranged hepatic cords and collapsed hepatic
sinusoids were observed in the TAA group. However, these changes
were not apparent in the TAA + C group (Fig. 2A).
TEM images further revealed numerous irregular, even
lysed, mitochondria, as well as an increased number of vesicles in
the TAA group. However, in the TAA + C group, restoration of the
aforementioned findings was observed, although partially broken and
incomplete mitochondria could still be found (Fig. 2B). Additionally, observation by SEM
revealed that microvilli in the TAA + C group were similar to those
in the control group. However, microvilli in the TAA group were
shorter, thinner and sparser, suggesting more severe hepatocyte
damage.
Co-treatment with celecoxib and
octreotide attenuates hepatocyte EMT
The expression levels of EMT-related markers,
including the TGF-β1/Smad signaling pathway factors, TGF-β1,
phosphorylated (p)-Smad2, Smad2, 3, 4, and 7, Snail1, E-cadherin
and N-cadherin, were determined in liver tissues by RT-qPCR, IHC
and western blot analysis (Fig. 3).
The mRNA expression levels of Smad3, Smad4 and Smad7 were not
significantly different among the three groups (P>0.05; Fig. 3B). Compared with those in the
control group, the mRNA levels of the pro-EMT markers, TGF-β1,
Smad2 and Snail1, were notably increased in the TAA group, and the
mRNA level of E-cadherin was significantly decreased in the TAA
group (Fig. 3B). Furthermore, the
levels of TGF-β1, Smad2 and Snail1 were markedly reduced in the TAA
+ C group compared with the TAA group. Finally, E-cadherin was
upregulated in the TAA + C group compared with the TAA group.
 | Figure 3Treatment with celecoxib and
octreotide attenuates epithelial-mesenchymal transition. (A)
Immunohistochemical staining of TGF-β1, p-Smad2/3, Snail1,
E-cadherin and N-cadherin in liver sections (magnification, x400;
insert, x1,000). (B) Relative TGF-β1, Smad2, Smad3, Smad4, Smad7,
Snail1 and E-cadherin mRNA expression was quantified by reverse
transcription-quantitative PCR (n=11). (C) Relative protein
expression levels of TGF-β1, Snail1, E-cadherin and N-cadherin, and
the ratio of p-Smad2/Smad2 in liver tissues were determined by
western blot analysis (n=6). #P<0.05 and
##P<0.01 vs. control group; *P<0.05 and
**P<0.01 vs. TAA group. p-Smad2, phosphorylated
Smad2; TAA, thioacetamide. |
The western blot analyses results revealed that the
levels of the pro-EMT markers, including TGF-β1, the ratio of
p-Smad2 to/Smad2, Snail1 and N-cadherin in hepatocytes of the TAA
group were markedly higher compared with those in the control group
(Fig. 3C). Similar results were
revealed by IHC (Fig. 3A). On the
contrary, the expression levels of the epithelial marker E-cadherin
in the TAA group were decreased compared with those in the control
group (P<0.01). Compared with the TAA group, TGF-β1, the ratio
of p-Smad2/Smad2, Snail1 and N-cadherin were markedly downregulated
and E-cadherin was notably upregulated in the TAA + C group
(P<0.05 or P<0.01).
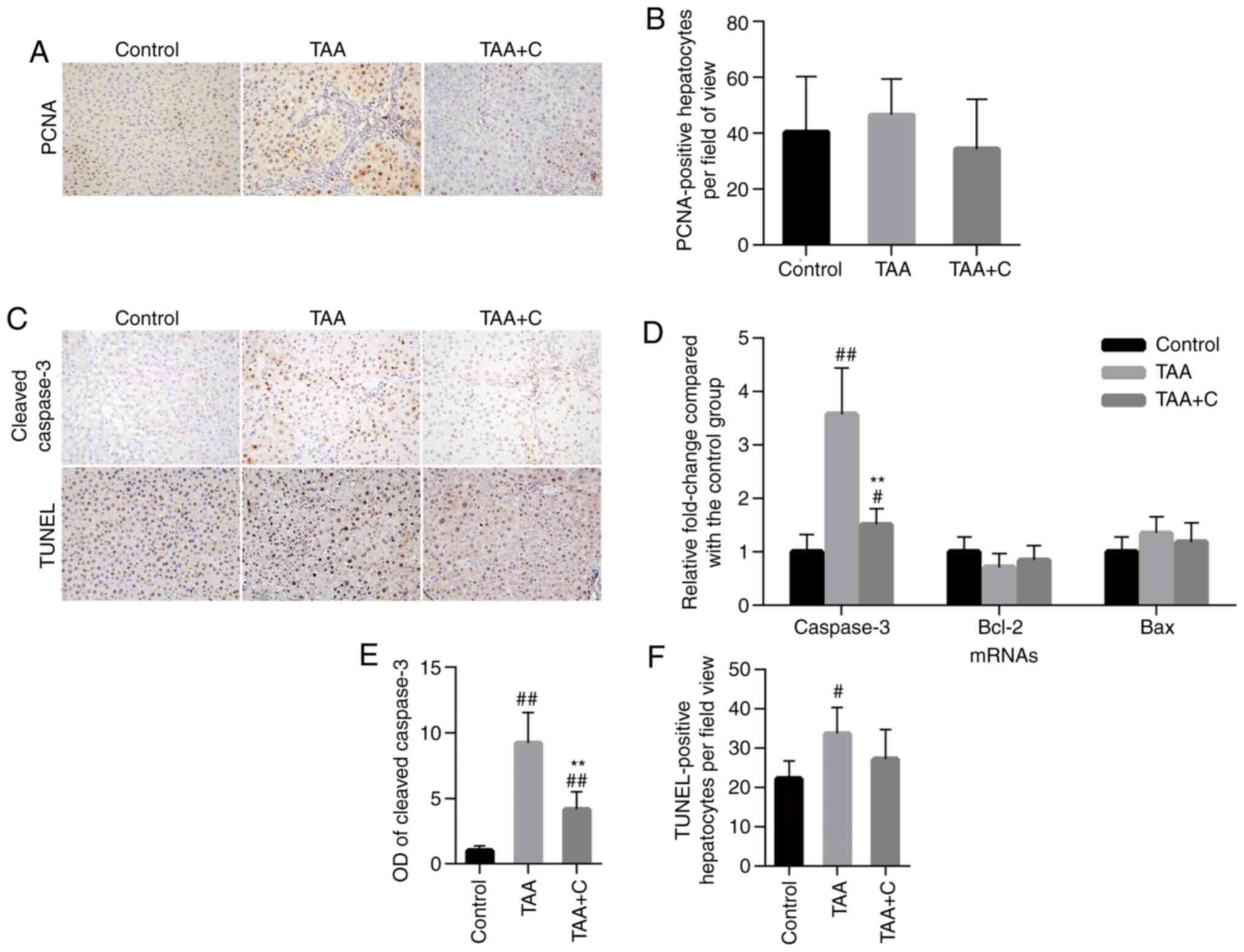
Celecoxib and octreotide have no
effect on hepatocyte proliferation
The protein expression of proliferating cell nuclear
antigen (PCNA) was evaluated by IHC staining by measuring the
PCNA-positive cell number. Hepatocytes were identified basing on
their morphology and the PCNA-positive hepatocytes were counted
(Fig. 4A and B). There were no significant difference in
the total number of PCNA-positive hepatocytes among the control,
TAA and TAA + C groups.
Celecoxib and octreotide affect
hepatocyte apoptosis
The mRNA expression levels of the apoptosis-related
factors, bax, caspase-3 and bcl-2 were measured by RT-qPCR to
evaluate the intrahepatic apoptosis status (Fig. 4D). Among all experimental groups, no
significant differences were observed in the mRNA expression levels
of bax and bcl-2. However, the mRNA expression levels of caspase-3
in the control group were significantly decreased compared with
those in the other two groups. The highest levels of caspase-3 were
observed in TAA group, which were also significantly higher
compared with those in the TAA + C group. To confirm the
aforementioned findings, the protein expression level of cleaved
caspase-3 was evaluated via IHC on slides. The results were
consistent with those observed in the RT-qPCR analysis (Fig. 4C and E).
The TUNEL staining results demonstrated that the
number of TUNEL-positive hepatocytes was statistically higher in
the TAA group compared with the control group (33.71±6.74 vs.
22.69±4,55; Fig. 4C and F). In addition, the number of
TUNEL-positive cells was reduced in the TAA + C group (27.23±7.45)
compared with the TAA group (33.71±6.74); however, the difference
was not statistically significant.
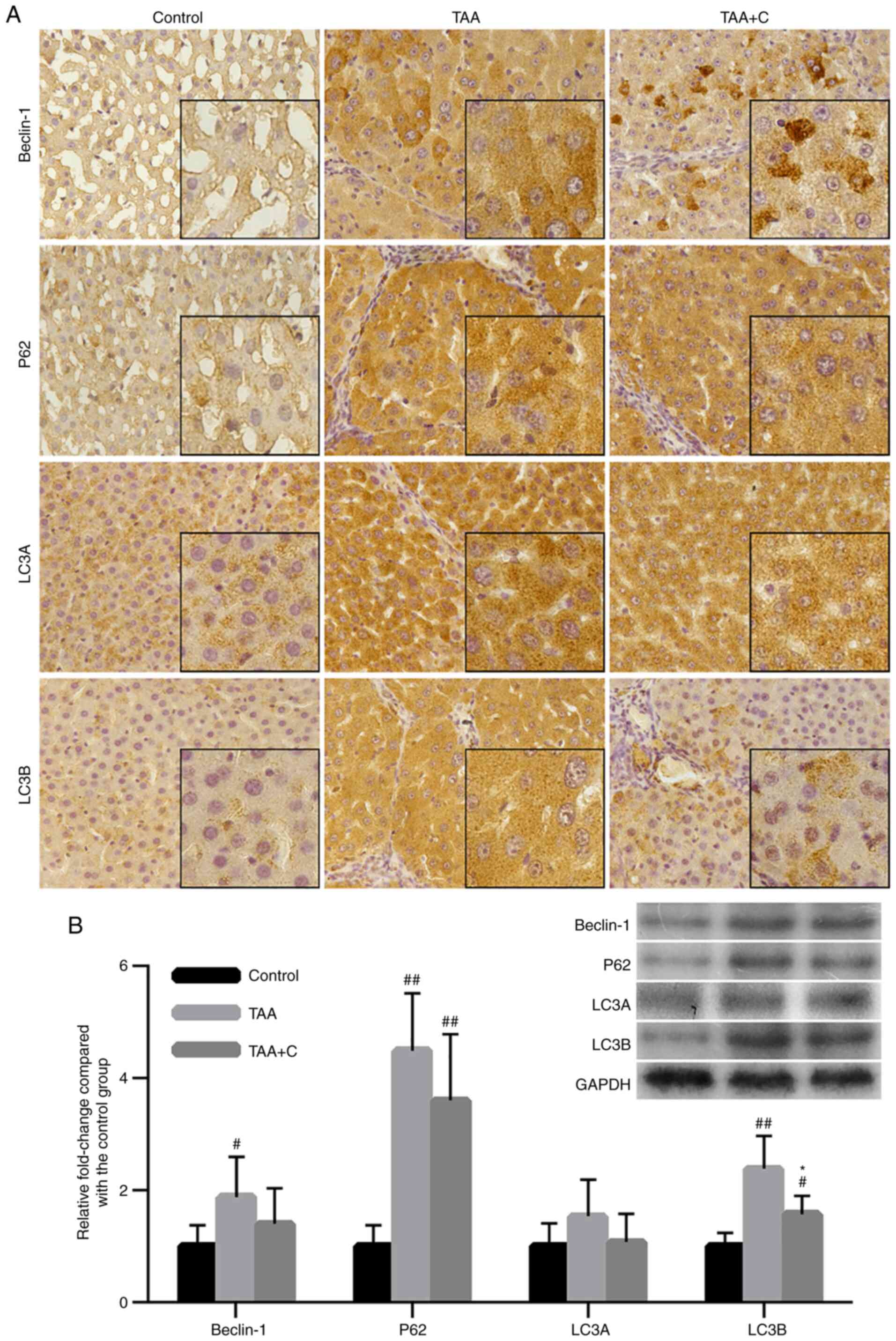
Co-treatment with celecoxib and
octreotide slightly attenuates hepatocyte autophagy
To investigate the effect of the combined treatment
with celecoxib and octreotide on inhibiting hepatocyte autophagy,
the expression levels of the autophagy-related markers beclin-1,
P62, LC3A and LC3B were assessed by IHC and western blot analysis
(Fig. 5). The IHC assay results
demonstrated that positive signal was mainly observed in
hepatocytes, while the mesenchyme was almost negative. All factors
tested were upregulated in the TAA and TAA + C groups compared with
the control group (Fig. 5A).
Furthermore, compared with the control group, the western blotting
results demonstrated that beclin-1, P62 and LC3B were notably
upregulated in the TAA group, P62 and LC3B were notably upregulated
in the TAA + C group, while the TAA group exhibited increased
expression levels compared with the TAA + C group. However, a
statistically significant difference between these groups was only
observed for LC3B expression (Fig.
5B).
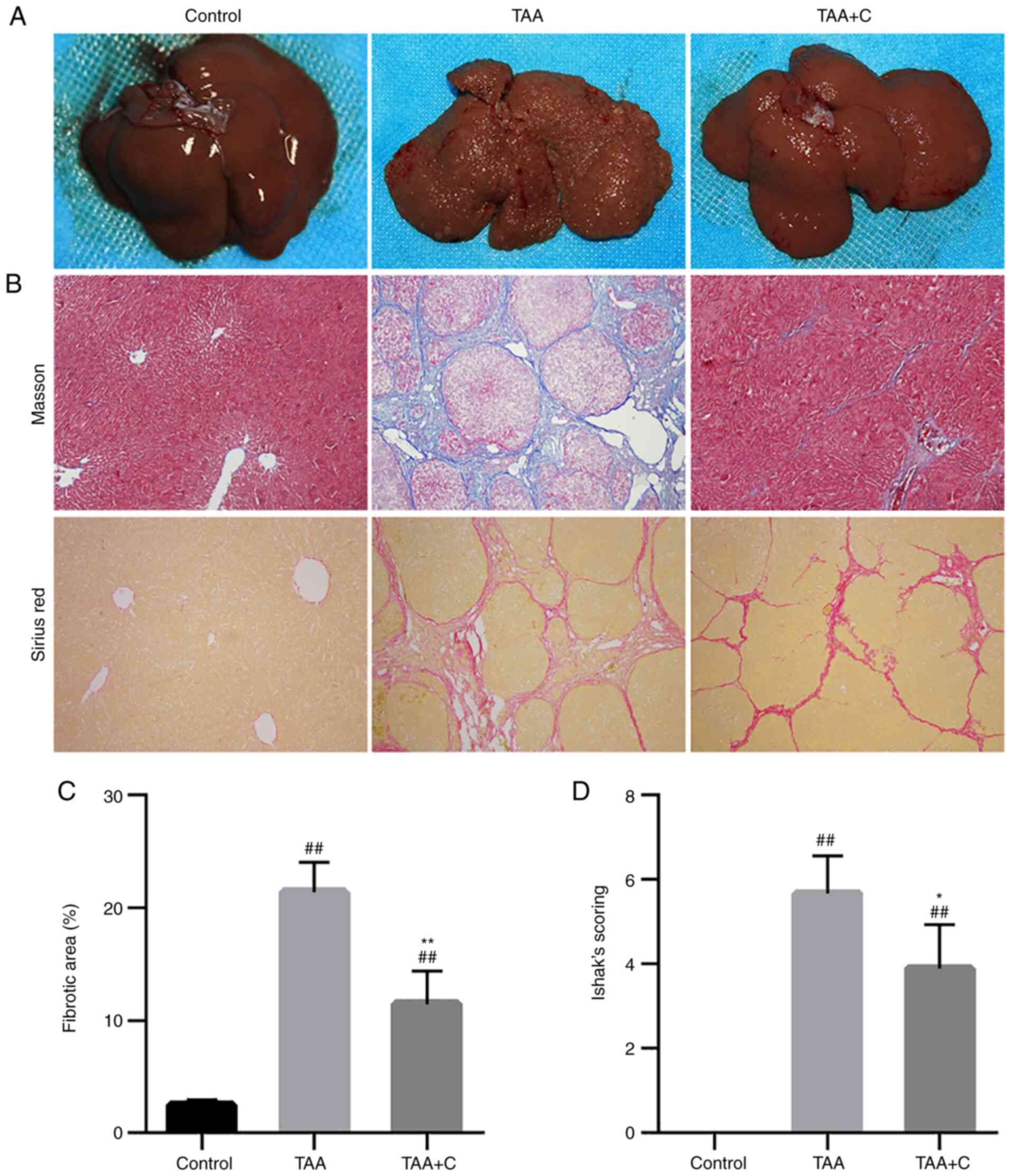
Celecoxib and octreotide attenuate
liver fibrosis
Based on the gross features of liver and
histological staining, the TAA group exerted the typical
morphological characteristics of fibrosis, with large nodules on
the surface and complete interlobular fibrotic septa (Fig. 6). In the TAA + C group, the size of
nodules on the surface was much smaller and the interlobular
fibrotic septa were incomplete (Fig.
6A and B). The statistical
analysis revealed that the fibrotic area and the Ishak's score in
the TAA and TAA + C groups were significantly higher compared with
those in the control group (P<0.01), and those in the TAA group
were markedly higher than those in the TAA + C group (P<0.05 or
P<0.01; Fig. 6C and D).
Discussion
In the present study, a hepatic fibrosis rat model
induced by TAA was established to investigate the effects of
celecoxib and octreotide on hepatic cells and liver fibrogenesis.
Co-treatment with these two drugs exhibited marked
anti-inflammatory effects, thereby strongly attenuating
intrahepatic inflammation. Furthermore, the drugs alleviated the
pathological injury and apoptosis of hepatocytes, and inhibited EMT
and hepatocyte autophagy, thus resulting in the inhibition of
hepatic fibrosis.
It is widely accepted that persistent inflammation
serves an important role in triggering liver fibrosis (20). In addition, the eruption of chronic
inflammation always induces liver fibrosis (1). Therefore, anti-inflammatory reactions
are considered as an efficient approach for inhibiting fibrogenesis
(21). Our previous study
demonstrated that celecoxib could improve the integrity of the
intestinal epithelial barrier, block the inflammatory transport
from the gut into the liver, and ameliorate the progress of hepatic
fibrosis (22). However, other
studies reported different results regarding the treatment of liver
fibrosis with celecoxib or SST analogues (23-26).
A number of studies failed to alleviate hepatic fibrosis induced by
CCl4 (25,26); however, these findings could be due
to treatment with extremely high doses of hepatoxin drugs, which
could mediate more severe damages, and low doses of celecoxib,
which could be insufficient to inhibit the excessive inflammatory
responses in the liver (25,26).
In addition, both celecoxib and SST analogues failed to treat liver
fibrosis induced by bile duct ligation (23,24).
In the present study, liver fibrosis in rats was slowly induced,
and the doses of celecoxib and octreotide were sufficient to
attenuate the expression of inflammatory factors, such as IL-6 and
TNF-α, thus providing strong anti-inflammatory and anti-fibrosis
effects.
Several studies have reported that EMT usually
occurs in the liver during liver fibrosis (27,28).
In the inflammatory microenvironment, the TGF-β1/Smads signaling
pathway is upregulated and activated, and is generally considered
as one of the crucial regulators of EMT (29). The Snail1 transcriptional factor
serves a key role in the control of EMT and fibroblast activation
(30). In the present study, along
with the upregulation of the expression levels of TGF-β1 and
p-Smad2, the pro-EMT TGF-β1/Smads signaling pathway was activated
in the TAA group compared with the control group. Snail1 was also
markedly upregulated in the TAA group compared with the control
group. Therefore, the changes in the expression levels of
EMT-related biomarkers, such as the highly expressed Snail1 and
N-cadherin, and the markedly downregulated E-cadherin, also
supported the occurrence of hepatocyte EMT in fibrotic liver.
However, studies based on genetic labeling techniques could not
identify any MFBs originating from either hepatocytes or
cholangiocytes (7,31). Apart from the shortcomings of the
genetic labeling technique itself, it should be considered almost
certain that even if hepatocytes could not be completely
transformed to MFBs, they could at least lose the epithelial
cell-related biomarkers and express those associated with
mesenchymal cells, thus contributing to hepatic fibrosis (32). In the present study, the combination
of celecoxib and octreotide reduced the activation of the
TGF-β1/Smads signaling pathway, downregulated the expression levels
of Snail1, and effectively alleviated hepatocyte EMT.
During the development of liver fibrosis, chronic
parenchymal damages, especially hepatocyte injury, are one of the
main features of fibrosis (1). On
the contrary, alleviating hepatocyte injury can attenuate the
progression of liver fibrosis (33). According to the histopathological
results, the shape, mitochondria and microvilli on the surface of
hepatocytes were well protected following co-treatment with
celecoxib and octreotide, indicating that the combination of these
drugs could attenuate hepatocyte injury, thus maintaining
hepatocyte function and attenuating the progress of liver
fibrosis.
Chronic inflammation in the liver may cause
hepatocyte death, leading to the release of apoptotic bodies and
other cellular debris, which are phagocytosed by HSCs, thus
resulting in their activation, proliferation, differentiation and
matrix deposition (34).
Furthermore, high levels of TGF-β1 promote hepatocyte cell death,
which contributes to liver fibrosis and later cirrhosis (35). An in vitro study has
demonstrated that an SST analogue, sorafenib, could reduce
apoptosis of murine hepatocytes (36). In the present study, the combined
treatment with celecoxib and octreotide protected hepatocytes from
apoptosis. This finding could be associated with the inhibition of
inflammation and the TGF-β1 signaling pathway.
Autophagy is a highly conserved eukaryotic cellular
self-eating catabolic pathway, which finally leads to the formation
of lysosomes (37). Autophagy
contributes to liver homeostasis through its role in energy balance
and in the quality control of the cytoplasm, by removing misfolded
proteins, damaged organelles and lipid droplets (38). The deregulation of autophagy has
been associated with several liver diseases and its regulation has
been recognized as a potential novel treatment strategy (39). However, the results of studies
investigating the effects of autophagy on liver fibrosis or
cirrhosis are contradictory (40-44).
A number of studies have suggested that autophagy prevents the
development of liver fibrosis (41,44).
On the other hand, other studies have demonstrated that autophagy
promotes the progression of hepatic fibrosis (42,43).
The aforementioned studies suggested that autophagy could regulate
multiple biological processes that affect the onset hepatic
fibrosis. In the present study, the expression levels of
autophagy-related markers beclin-1, P62 and LC3A/B were increased
in hepatocytes in the TAA group. Among these factors, LC3B is
considered to be the most typical indicator for the evaluation of
cellular autophagy (45). The
results of the present study demonstrated that the expression
levels of LC3B in the TAA + C group were notably decreased compared
those in the TAA group, indicating that the level of autophagy in
the TAA + C group was also lower than that in the TAA group. The
present study hypothesized that co-treatment with celecoxib and
octreotide could prevent intrahepatic inflammation-mediated damage
to hepatocytes.
At present, etiological and symptomatic therapy are
the main treatment approaches for liver cirrhosis in clinical
settings (46). For example,
antiviral therapy is used to treat hepatitis virus-related
cirrhosis, endoscopic variceal ligation is used for treating
gastrointestinal bleeding, while transjugular intrahepatic
portosystemic stent-shunt is applied for treating refractory
ascites and portal hypertension (47). However, the treatment for protection
of hepatocytes remains under investigation. In conclusion, the
present study demonstrated that inhibition of intrahepatic
inflammation by co-treatment with celecoxib and octreotide could
alleviate TAA-induced hepatic fibrosis in rats by protecting
hepatocytes. Several mechanisms could be involved in this process,
such as attenuation of pathological injury, and inhibition of
hepatocyte apoptosis, EMT and autophagy. Therefore, the combined
treatment with celecoxib and octreotide could reduce the
progression of hepatic fibrosis and may be considered as a
potential therapy approach.
Supplementary Material
Primers used for quantitative PCR
analysis.
Antibodies used for
immunohistochemistry and western blotting.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National Natural
Science Foundation of China (grant no. 81500468) and Open Subject
of Provincial Key Laboratory of Preclinical Medicine in 2020 (grant
no. JCKF2020005).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SLW and HYZ conceived and designed the present
study. SF and HT performed the experiments. JHG and GMW analyzed
the data. SHT and WJY contributed to interpretation of data. SF
wrote the manuscript and SLW edited the paper. SLW and HYZ confirm
the authenticity of all the raw data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The experiments were approved by the Animal Use and
Care Committee of Sichuan University (approval no. K2015004;
Chengdu, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interest.
References
|
1
|
Parola M and Pinzani M: Liver fibrosis:
Pathophysiology, pathogenetic targets and clinical issues. Mol
Aspects Med. 65:37–55. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bodzin AS and Baker TB: Liver
transplantation today: Where we are now and where we are going.
Liver Transpl. 24:1470–1475. 2018.PubMed/NCBI View
Article : Google Scholar
|
|
3
|
Shu Y, Liu X, Huang H, Wen Q and Shu J:
Research progress of natural compounds in anti-liver fibrosis by
affecting autophagy of hepatic stellate cells. Mol Biol Rep.
48:1915–1924. 2021.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Campana L and Iredale JP: Regression of
liver fibrosis. Semin Liver Dis. 37:1–10. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yu K, Li Q, Shi G and Li N: Involvement of
epithelial-mesenchymal transition in liver fibrosis. Saudi J
Gastroenterol. 24:5–11. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Xie G and Diehl AM: Evidence for and
against epithelial-to-mesenchymal transition in the liver. Am J
Physiol Gastrointest Liver Physiol. 305:G881–G890. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Taura K, Miura K, Iwaisako K, Osterreicher
CH, Kodama Y, Penz-Osterreicher M and Brenner DA: Hepatocytes do
not undergo epithelial-mesenchymal transition in liver fibrosis in
mice. Hepatology. 51:1027–1036. 2010.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wen SL, Gao JH, Yang WJ, Lu YY, Tong H,
Huang ZY, Liu ZX and Tang CW: Celecoxib attenuates hepatic
cirrhosis through inhibition of epithelial-to-mesenchymal
transition of hepatocytes. J Gastroenterol Hepatol. 29:1932–1942.
2014.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Mallat A, Lodder J, Teixeira-Clerc F,
Moreau R, Codogno P and Lotersztajn S: Autophagy: A multifaceted
partner in liver fibrosis. Biomed Res Int.
2014(869390)2014.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Lee YA, Wallace MC and Friedman SL:
Pathobiology of liver fibrosis: A translational success story. Gut.
64:830–841. 2015.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Rouzer CA and Marnett LJ: Cyclooxygenases:
Structural and functional insights. J Lipid Res. 50
(Suppl):S29–S34. 2009.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Holt AP and Adams DH: Complex roles of
cyclo-oxygenase 2 in hepatitis. Gut. 56:903–904. 2007.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gissen P and Arias IM: Structural and
functional hepatocyte polarity and liver disease. J Hepatol.
63:1023–1037. 2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Tulipano G and Schulz S: Novel insights in
somatostatin receptor physiology. Eur J Endocrinol. 156:S3–S11.
2007.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Barnett P: Somatostatin and somatostatin
receptor physiology. Endocrine. 20:255–264. 2003.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Sun L and Coy DH: Somatostatin and its
Analogs. Curr Drug Targets. 17:529–537. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Li X, Benjamin IS and Alexander B:
Reproducible production of thioacetamide-induced macronodular
cirrhosis in the rat with no mortality. J Hepatol. 36:488–493.
2002.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Ishak K, Baptista A, Bianchi L, Callea F,
De Groote J, Gudat F, Denk H, Desmet V, Korb G, MacSween RN, et al:
Histological grading and staging of chronic hepatitis. J Hepatol.
22:696–699. 1995.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hellemans J, Mortier G, De Paepe A,
Speleman F and Vandesompele J: qBase relative quantification
framework and software for management and automated analysis of
real-time quantitative PCR data. Genome Biol. 8(R19)2007.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kartasheva-Ebertz DM, Pol S and Lagaye S:
Retinoic acid: A new old friend of IL-17A in the immune pathogeny
of liver fibrosis. Front Immunol. 12(691073)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Alegre F, Pelegrin P and Feldstein AE:
Inflammasomes in liver fibrosis. Semin Liver Dis. 37:119–127.
2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Gao JH, Wen SL, Tong H, Wang CH, Yang WJ,
Tang SH, Yan ZP, Tai Y, Ye C, Liu R, et al: Inhibition of
cyclooxygenase-2 alleviates liver cirrhosis via improvement of the
dysfunctional gut-liver axis in rats. Am J Physiol Gastrointest
Liver Physiol. 310:G962–G972. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yu J, Hui AY, Chu ES, Go MY, Cheung KF, Wu
CW, Chan HL and Sung JJ: The anti-inflammatory effect of celecoxib
does not prevent liver fibrosis in bile duct-ligated rats. Liver
Int. 29:25–36. 2009.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Tahan G, Eren F, Tarçin O, Akin H, Tahan
V, Şahın H, Özdoğan O, İmeryüz N, Çelıkel Ç, Avşar E and Tözün N:
Effects of a long-acting somatostatin analogue, lanreotide, on bile
duct ligation-induced liver fibrosis in rats. Turk J Gastroenterol.
21:287–292. 2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hui AY, Leung WK, Chan HL, Chan FK, Go MY,
Chan KK, Tang BD, Chu ES and Sung JJ: Effect of celecoxib on
experimental liver fibrosis in rat. Liver Int. 26:125–136.
2006.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Harris TR, Kodani S, Rand AA, Yang J, Imai
DM, Hwang SH and Hammock BD: Celecoxib does not protect against
fibrosis and inflammation in a carbon tetrachloride-induced model
of liver injury. Mol Pharmacol. 94:834–841. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Pinzani M: Epithelial-mesenchymal
transition in chronic liver disease: Fibrogenesis or escape from
death? J Hepatol. 55:459–465. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lee SJ, Kim KH and Park KK: Mechanisms of
fibrogenesis in liver cirrhosis: The molecular aspects of
epithelial-mesenchymal transition. World J Hepatol. 6:207–216.
2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Tsubakihara Y and Moustakas A:
Epithelial-mesenchymal transition and metastasis under the control
of transforming growth factor β. Int J Mol Sci.
19(3672)2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Baulida J, Díaz VM and Herreros AG:
Snail1: A transcriptional factor controlled at multiple levels. J
Clin Med. 8(757)2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Scholten D, Osterreicher CH, Scholten A,
Iwaisako K, Gu G, Brenner DA and Kisseleva T: Genetic labeling does
not detect epithelial-to-mesenchymal transition of cholangiocytes
in liver fibrosis in mice. Gastroenterology. 139:987–998.
2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Taura K, Iwaisako K, Hatano E and Uemoto
S: Controversies over the epithelial-to-mesenchymal transition in
liver fibrosis. J Clin Med. 5(9)2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Pinheiro D, Dias I, Ribeiro Silva K,
Stumbo AC, Thole A, Cortez E, de Carvalho L, Weiskirchen R and
Carvalho S: Mechanisms underlying cell therapy in liver fibrosis:
An overview. Cells. 8(1339)2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tsuchida T and Friedman SL: Mechanisms of
hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol.
14:397–411. 2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Fabregat I, Moreno-Càceres J, Sánchez A,
Dooley S, Dewidar B, Giannelli G and Ten Dijke P: IT-LIVER
Consortium. TGF-β signalling and liver disease. FEBS J.
283:2219–2232. 2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Chen YL, Lv J, Ye XL, Sun MY, Xu Q, Liu
CH, Min LH, Li HP, Liu P and Ding X: Sorafenib inhibits
transforming growth factor 1-mediated epithelial-mesenchymal
transition and apoptosis in mouse hepatocytes. Hepatology.
53:1708–1718. 2011.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Song Y, Zhao Y, Wang F, Tao L, Xiao J and
Yang C: Autophagy in hepatic fibrosis. BioMed research
international. 2014(436242)2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Boya P, Reggiori F and Codogno P: Emerging
regulation and functions of autophagy. Nat Cell Biol. 15:713–720.
2013.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Allaire M, Rautou PE, Codogno P and
Lotersztajn S: Autophagy in liver diseases: Time for translation? J
Hepatol. 70:985–998. 2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Ke PY: Diverse functions of autophagy in
liver physiology and liver diseases. Int J Mol Sci.
20(300)2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Tong M, Zheng Q, Liu M, Chen L, Lin YH,
Tang SG and Zhu YM: 5-methoxytryptophan alleviates liver fibrosis
by modulating FOXO3a/miR-21/ATG5 signaling pathway mediated
autophagy. Cell Cycle. 20:676–688. 2021.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Kuscuoglu D, Bewersdorf L, Wenzel K, Gross
A, Kobazi Ensari G, Luo Y, Kilic K, Hittatiya K, Golob-Schwarzl N,
Leube RE, et al: Dual proteotoxic stress accelerates liver injury
via activation of p62-Nrf2. J Pathol. 254:80–91. 2021.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Tan S, Liu X, Chen L, Wu X, Tao L, Pan X,
Tan S, Liu H, Jiang J and Wu B: Fas/FasL mediates
NF-κBp65/PUMA-modulated hepatocytes apoptosis via autophagy to
drive liver fibrosis. Cell Death Dis. 12(474)2021.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Song M, Zhang H, Chen Z, Yang J, Li J,
Shao S and Liu J: Shikonin reduces hepatic fibrosis by inducing
apoptosis and inhibiting autophagy via the platelet-activating
factor-mitogen-activated protein kinase axis. Exp Ther Med.
21(28)2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Dikic I and Elazar Z: Mechanism and
medical implications of mammalian autophagy. Nat Rev Mol Cell Biol.
19:349–364. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Fukui H, Saito H, Ueno Y, Uto H, Obara K,
Sakaida I, Shibuya A, Seike M, Nagoshi S, Segawa M, et al:
Evidence-based clinical practice guidelines for liver cirrhosis
2015. J Gastroenterol. 51:629–650. 2016.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Yoshiji H, Nagoshi S, Akahane T, Asaoka Y,
Ueno Y, Ogawa K, Kawaguchi T, Kurosaki M, Sakaida I, Shimizu M, et
al: Evidence-based clinical practice guidelines for Liver Cirrhosis
2020. J Gastroenterol. 56:593–619. 2021.PubMed/NCBI View Article : Google Scholar
|