Kidney disease, which is one of the fastest-growing
causes of mortality worldwide, includes acute and chronic types
that damage the structure and function of renal tissue (1). Acute kidney injury (AKI) and chronic
kidney disease (CKD) are associated with increased risk of adverse
cardiovascular events, as well as increased morbidity and mortality
(2,3). These conditions can eventually develop
into end-stage renal disease (ESRD) that requires dialysis or
kidney replacement (4). The global
economic cost of kidney disease is vast. In 2007, the United States
Medicare expenditures on CKD exceeded $60,000,000 USD, accounting
for 27% of the total Medicare budget (5), while expenditures on AKI exceed
$10,000,000 USD per year (6).
Therefore, elucidating the pathophysiological mechanisms of kidney
disease will lead to the development of promising therapeutics for
multiple renal diseases, which is essential to public health.
Reversible phosphorylation and dephosphorylation of
proteins serve important roles in regulating protein activities and
constitute the central mechanism of signal transduction (7). Protein phosphatase 2A (PP2A) enzymes
are the main members of the serine/threonine (Ser/Thr) phosphatase
family and are involved in the majority of total
cellular-phosphatase activities (7). PP2A, which participates in a number of
key pathways during all stages of the cell cycle, has been
extensively studied in relation to tumor suppression (8). A growing body of literature suggests
that PP2A is crucial in a number of aspects of renal-disease
pathogenesis including AKI (9), CKD
(10), aging (11) and tumorigenic processes (12). In the current review, the unique
structure and function of PP2A is described and its interactions
with associated signaling pathways in kidney disease are discussed.
Therapeutic targeting of PP2A has demonstrated promise in enhancing
the efficacy of treatments for patients with kidney disease.
Protein phosphorylation and dephosphorylation are
components of a vital mechanism used in the modulation of key
cellular processes associated with cell proliferation,
differentiation, migration and other biological behaviors involved
in signaling regulation (13,14).
Phosphatases are divided into four large families: i) Protein
serine/threonine phosphatases [including the protein phosphatases
(PPPs) and metal-dependent protein phosphatases], (2) protein tyrosine phosphatases (2), dual-specificity phosphatases (DUSP)
and histidine phosphatases (3,4). As
the largest of the Ser/Thr family, PPP is further subdivided into
PPl, PP2A, PP2B, PP4, PPS and PP6 according to the sensitivity and
specificity of the substrate to inhibitors or effectors (15,16). A
number of extracellular stimuli affect the level of protein
phosphorylation in the modulation of cell processes (17). These processes are involved in
cellular proliferation, migration, growth, differentiation,
metabolism, the immune system, cytoskeletal reorganization and
muscle contraction (17). Over 50%
of proteins in human cells are capable of undergoing reversible
phosphorylation; Therefore, phosphorylation pathways exhibit
promising potential applications in the treatment for a variety of
different diseases (18).
The kidneys receive ~20% of cardiac output and
consume 10% of the bodies oxygen to perform their main function of
adjusting body-fluid constituents by glomerular filtration and
renal tubular reabsorption (4).
Each glomerulus consists of three main classes of cells forming the
filtration layer: i) fenestrated-endothelium cells located inside
the capillary, ii) mesangial cells located outside the capillary
and iii) specialized epithelial cells called podocytes (19). Phosphotyrosine phosphatase is
hyperactive in endothelial cells of the glomerular-mesangial-cell
layer in human tissues (20).
Furthermore, PTP serves a central role in podocyte homeostasis, and
this has been demonstrated in a study in which treatment of
podocytes with a nonspecific PTP inhibitor induced drastic
morphological alterations in their actin-cytoskeleton network
(21). Svennilson et al
(22) demonstrated that the
expression of PP1 and PP2A is ubiquitous in the early metanephric
kidney. The importance of kinases in kidney disease is well
established; However, the roles of phosphatases are yet to be fully
elucidated (10). To the best of
our knowledge, there has been little research on phosphatase in the
kidneys. Accumulating evidence has indicated that phosphatases are
involved in normal nephron growth and renal pathological processes,
and may be a promising target for therapies in patients with renal
diseases (10,23).
In eukaryotic cells, at least 99% of protein
phosphorylation is associated with Ser/Thr residues (24). PP2A, which is one of the four major
cytoplasmic Ser/Thr phosphatases, acts on a number of different
components within various key signal-transduction pathways
(25). The function and structure
of PP2A is conserved in organisms ranging from yeasts to mammals,
in which PP2A regulates many important cellular functions (26). The PP2A enzyme core is made up of a
65-kDa scaffolding subunit ‘A’, which modulates its enzymatic
properties, and a 36-kDa catalytic subunit ‘C’ (27). These subunits bind to the regulatory
subunit ‘B’ (PP2A-B) to form the various heterotrimeric complexes
(28). The A and C subunits have
two isoforms that are encoded by genes α and β (29). While these isoforms are strongly
homologous for each other, the majority of cell types predominantly
express the α isoform (29). There
are four categories of the B subunit, including the PR55 (B), PR61
(B'), PR72/130 (B') and the striatin family (B'). The B subunit is
contained in multiple isoforms (α up to ε) that are encoded by
different genes. Because the structure of the B subunit varies
dramatically, PP2A targets an extensive array of components
involved in critical signal-transduction pathways that regulate
cellular functions (29). Although
the A and C subunits are widely expressed, the cellular
localization and expression of the B subunit vary greatly across
cell types and tissues (30-35).
PP2A dephosphorylates a number of key cellular
molecules including Akt, MEK, MAPK, c-Myc, p53 and β-catenin
(36). Furthermore, PP2A regulates
a number of different cellular processes such as proliferation,
metabolism and apoptosis (36).
Aberrant PP2A regulation is commonly observed in a range of
diseases such as cancer, cardiovascular pathologies and
neurodegenerative disorders (29).
Evidence has indicated that PP2A can act as a tumor suppressor
(37). PP2A can suppress
tumorigenesis via downregulation of the Akt/(TSC1/TSC1)/Rheb/mTOR
signaling pathway, inactivation of c-Myc and antagonism of the
Wnt/β-catenin pathway (38).
Previous studies have also revealed that PP2A is essential in
kidney organogenesis and developmental processes, and participates
in kidney diseases via one of several complex mechanisms (39,40).
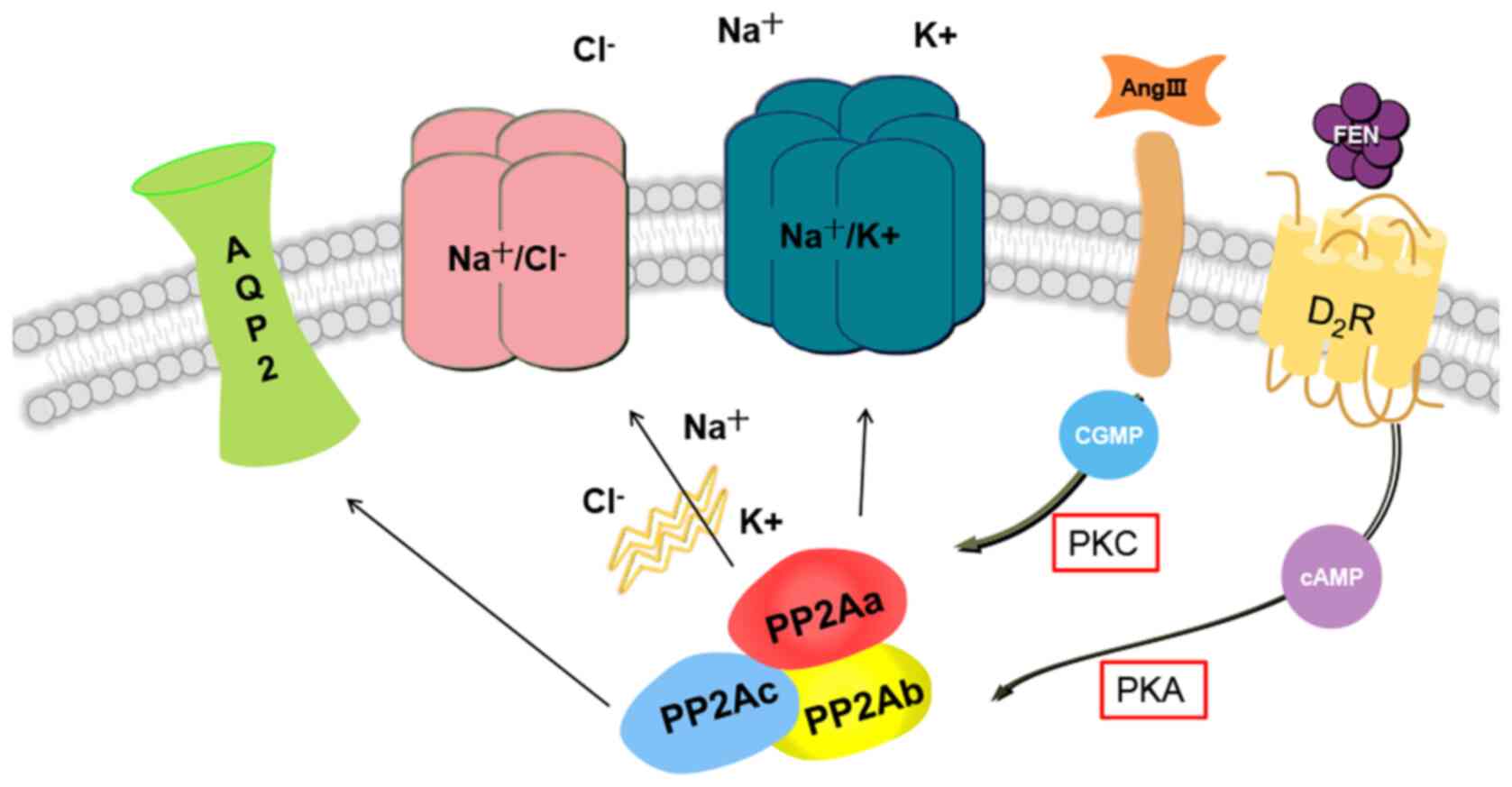
The absorption of ions in the kidneys is regulated
and controlled by multiple physiological mechanisms (41). Previous studies have indicated PP2A
is responsible for the maintenance of ion channels and homeostasis
(42-44).
PP1 and PP2A serve an important role in the phosphorylation,
surface distribution and function of
Na+/Cl--dependent choline transporters
(43). Ang II is an important
regulator of the ouabain-resistant Na+-ATPase from renal
proximal tubule cells, which are involved in the signal cascade of
Ang II receptors, protein kinase (PK)A and PKC (42,44-46).
Chronic malnutrition can stimulate the activity of renal
tubulointerstitial Ang II and damage the regulation of
phosphorylation that is mediated by PP2A (47). These events result in excessive
reabsorption of Na+ in proximal renal tubuloepithelial
cells (47). Gildea et al
(48) demonstrated that PP2A
mediates increased activity of Ang II, dopamine receptors and cAMP
in proximal tubules. This aforementioned study also demonstrated
that abnormal control of these systems leads to an increase in
sodium reabsorption. Furthermore, cell-permeable second-messenger
analogs cAMP and GMP can activate PP2A, which is important for
plasma membrane recruitment of dopamine D1 receptor (D1R),
angiotensin II type-2 receptor and inhibition of luminal
Na+ transport (48). In
human adenocarcinoma cells, PP2A also participates in stimulation
of D1R, which increases Na+/K+-ATPase
activity by accelerating Na+/K+-ATPase
recruitment from intracellular pools into the basal-lateral
membrane; Meanwhile, okadaic acid (OA), which a selective inhibitor
of PP2A, inhibits the increase in
Na+/K+-ATPase activity (49). Li et al (50) indicated that calyculin, which is a
PP1 and PP2A inhibitor, inhibits
Na+/K+-ATPase activity at a lower dose than
that of OA in renal proximal tubules. In addition, potassium
channels are also regulated by these phosphatases (51). 11,12-epoxyeicosatrienoic acid
dilates preglomerular microvessels via adenosine A2A receptor
mediation and activates conductance across the K+
channels in renal smooth muscles by stimulating PP2A (52). These findings revealed that PP2A
functions cooperatively with this crucial regulatory mechanism in
regulating the dynamics of Na+, K+ and
Cl- flow (Fig. 1).
The targeting of AQP2 on the apical plasma membrane
of renal collecting duct cells, which are used to maintain body
water homeostasis in mammals, is mainly modulated via the activity
of antidiuretic peptide arginine vasopressin (53). AQP2 protein has at least four
vasopressin-regulated phosphorylation sites at serine residues 256,
261, 264 and 269 (54,55). Phosphorylation of these sites
controls the transport of AQP2 in and out of the plasma membrane at
the top of the collecting tube (56-60).
PP1 and/or PP2A act on the internalization of membrane-associated
AQP2 by reducing the phosphorylation of ser256 and ser264 in kidney
inner-medullar tissue (Fig. 1). The
phosphorylation site at serine-264 of AQP2 is also regulated by
PP2B (61). However, PP2B does not
affect the membrane accumulation of AQP2(61). Calyculin-mediated inhibition of PP2A
activity leads to an increase in p256-AQP2 expression and a
decrease in p261-AQP2 expression in MDCK-hAQP2 cells, which
promotes AQP2 transport to the plasma membrane (62). Valenti et al (63) demonstrated that OA increases the
phosphorylation of AQP2 at ser256 by 60% and promotes AQP2 to
target the apical plasma membrane of collecting duct cells.
PP2A serves a crucial role in embryonic development,
and particularly in development and survival (40). Evidence has suggested that PP2A
serves complicated stimulatory and inhibitory roles in growth- and
hormone-factor signaling, especially in the extracellular
signal-regulated ERK/MAPK cascade and in the activity of ubiquitous
intermediary messengers during mitosis (64). PP2A appears to be involved in
controlling the activity of maturation-promoting factor (MPF)
(65,66) and the modulation of MAP-ERK kinase
activity (67,68). Normal kidney development, beginning
at embryonic day 12 (E12) in rats and during the 5th gestational
week in humans, is strictly regulated and involves several crucial
steps in order to achieve a predetermined number of functioning
nephrons (39). On day 15 of
embryonic development, the percentage of PP2A in total Ser/Thr
phosphatase activity is 78% in rat kidneys, as evaluated using a
phosphatase activity assay. At the El8 and E21 stages of
nephrogenesis, the expression of PP2A is limited to the nephrogenic
zone, in which it is strongly expressed (39). When nephrogenesis terminates, PP2A
expression is downregulated substantially (39). Svennilson et al (22) demonstrated that PP2A mRNA is
strongly expressed in various cell types during early development
of the kidneys. Additionally, the use of low doses of OA inhibits
early (E13) embryonic kidney growth and disturbs nephron formation
in E15 kidneys. Subsequently, normal PP2A activity is indispensable
to metanephric development, and inhibition of this activity can
induce morphological disorder and apoptosis (22).
Recent studies have highlighted the particular
importance of functional PP2A in Wnt signaling due to the fact
Wnt-4 serves a key role in kidney development (69,70).
Overall, these studies provide novel insights into the importance
of PP2A activity during renal morphogenesis and within
signal-transduction pathways.
Podocytes (or visceral epithelial cells) have
complicated interdigitating foot processes (FPs) that cover the
external surface of the glomerular basement membrane (71). Adjacent FPs from different units are
interconnected by a continuous membrane structure called the slit
diaphragm (19). The slit diaphragm
consists of membrane and cytoskeletal proteins, such as
synaptopodin, nephrin, podocin, α-actinin-4, podoplanin and CD2
Associated Protein, as well as signaling molecules, all of which
serve important roles in maintaining the basic function of the
glomerular filtration barrier (GFB) (72,73).
Impairments in pathways and molecular processes that regulate the
function of the GFB may lead to CKD (74). Congenital or acquired podocyte
damage can cause podocytes to lose certain specific markers,
causing disappearance of FPs, detachment and proteinuria (75). Additionally, podocyte injury is
closely associated with a number of renal diseases, including
diabetic nephropathy, membrane nephritis, IgA nephropathy and focal
segmental glomerulosclerosis (75).
Kumar and Tikoo (20) demonstrated
that selective inhibition of PP2A activity to restore insulin
levels can induce the phosphorylation of Sirtuin 1 and Forkhead Box
O1 and increase the activity of AKT, causing degradation of p53 and
podocyte death. Kobayashi et al (76) also indicated that utilizing OA to
inhibit PP2A can suppress microtubule elongation and abolish
process formation in conditionally immortalized mouse podocytes.
Zhu et al (77) demonstrated
that podocyte-specific knockout of PP2A (Pod-PP2A-KO) in mice
causes considerable weight loss, growth retardation, proteinuria,
severe lethargy, and mortality in >70% mice at 15 weeks of age.
Histological examination has indicated severe glomerulopathy and
dramatic loss of FPs, as well as reduced expression of a number of
different slit-diaphragm molecules and impairment of cytoskeletal
rearrangement in podocytes (77).
The highly conserved Y box protein 1 (YB-1) of the
cold-shock protein family has been indicated to be associated with
cellular stress response and renal fibrosis (78). YB-1 is a target molecule in PP2A
dephosphorylation, and fine-tuning YB-1 via post-translational
modification by modulating PP2A activity may serve a role in
maintaining the functional integrity of podocytes and GFB (78). Zhong et al (79) revealed that podocyte-specific PP2A
deficiency aggravates diabetic glomerulopathy and accelerates
diabetic kidney disease. Arctigenin (ATG) is a major component of
Fructus Arctii, a traditional herbal remedy that reduces
proteinuria in patients with diabetes (79). ATG administration has been revealed
to attenuate proteinuria and podocyte injury in mouse models of
diabetes (79). Furthermore,
enhanced PP2A activity occurring via ATG ameliorates podocyte
adhesion partly through T335-mediated phosphorylation of drebrin-1
(DBN1). This aforementioned result reveals a novel mechanism in the
regulation of podocyte cytoskeletal rearrangement (79). Selective inhibition of PP2A can
improve insulin resistance, restore AKT levels, induce FOXO1
phosphorylation and rescue podocytes from cell death (20). These aforementioned data indicated
that PP2A may be a potential drug target for the prevention of
podocyte injury.
Renal cell carcinoma (RCC) is one of the most common
renal malignancies and accounts for ~2.4% of all cancers and 1.7%
of total cancer-associated deaths worldwide (80). To date, therapeutics for patients
with metastatic renal cancer remain limited in effectiveness and
specificity (81). A number of
human cancers are associated with PP2A dysfunction, such as lung
cancer (82), breast cancer
(83) and leukemia (84). Furthermore, downregulation of PP2A
expression and its impact on cellular transformation reveals that
PP2A can function as a tumor-suppressor gene in a number of
different malignant cancers including leukemia, lung, breast,
gastric and colon cancer (85).
Increasing evidence has indicated that PP2A serves a
tumor-suppressive role, but the individual roles of its subunits,
which are deregulated in cancer, remain unknown (80). PP2A expression is decreased in RCC
tissues, and patients with high expression of PP2A in tumor tissues
exhibit improved survival compared with those exhibiting a low
expression of PP2A (12). However,
the mechanism of PP2A deregulation in RCC is yet to be determined.
Evidence has suggested that PP2A inactivation in cancer mainly
occurs via overexpression of suvar/enhancer of zeste/trithorax and
cancerous inhibitor of PP2A, which are both endogenous PP2A
inhibitors (86-89).
MicroRNAs (miRNA/miR) can act as oncogenes or tumor
suppressor genes, depending on the function of target genes in
malignant tumors (87).
Additionally, miR-183 (a member of the miR-183-96-182 cluster) is
expressed at higher levels in two renal cancer cell lines (ACHN and
A498) and can promote the growth of renal cancer cells (87). A study has indicated that miR-183
can directly target the 3'untranslated regions of PP2A-Cα, PP2A-Cβ
mRNA, as well as PP2A-B56-γ protein phosphatase subunits and
inhibit their expression (81).
These results confirm that miR-183 serves an oncogenic role in
renal cancer cells by targeting PP2A directly (81). PP2A, Akt and Mcl-1 are essential in
RCC malignancy and treatment resistance (90). The PP2A/Akt axis is an important
substitute for aspirin-mediated induction of susceptibility to
ABT-737 mediated-apoptosis in RCC cells (90). Styryllactone (R)-goniothalamin and
its enantiomer (S)-goniothalamin cause apoptosis in cancer cells of
human kidneys by reducing Ras expression and PP2A activity
(91).
In human renal-carcinoma Caki cells, downregulation
of PP2A via small-interfering RNA can significantly inhibit the
upregulation of a pro-apoptotic protein Bim via the pharmacological
inhibitor ZFL [cathepsin S inhibitor: Z-FL-COCHO (ZFL)] (92). Downregulation of PP2A decreases
apoptosis and cleavage of poly (ADP-ribose) polymerase in ZFL- and
oxaliplatin-treated Caki cells (92). Combined treatment with Raf
inhibitors sorafenib and GW5074 is used to produce a two-pronged
attack on renal cancer cells (93).
The two inhibitors promote translocation of pC-RafS338 and
pDAPKS308 from mitochondria to cytoplasm, resulting in
mitochondrial dysfunction and ROS generation. Subsequently,
reactive oxygen species (ROS) accelerate the PP2A-mediated
dephosphorylation of pDAPKS308 to DAPK. Finally, PP2A separates
from the C-Raf-DAPK complex and leads to cancer cell death
(93). Luteolin also induces
apoptosis in 786-o cells (94).
This cytotoxicity is caused by the downregulation of Akt and
consequent upregulation of Ask1, p38 and JNK activity, which is
regulated by PP2A activation (94).
In summary, PP2A expression may be a useful tool
that can be used in the prediction of prognosis and therapeutic
outcome in patients with RCC. Further research on the molecular
mechanisms of PP2A in human RCC will facilitate the identification
of novel therapeutics and the development of effective treatments
for patients with renal cancer.
Glomerulosclerosis and tubulointerstitial fibrosis
are the major pathological features of renal fibrosis, which is the
final manifestation of a number of different CKDs (95). As conductors, renal microvascular
endothelial cells (EC) serve important roles in kidney fibrosis
(96). The association between
renal fibrosis and endothelial dysfunction is well established
(97). Endothelial dysfunction
leads to a significant reduction in the number of peritubular
capillaries in the interstitium (97). Chronic ischemia and hypoxia result
in scar formation and remodeling processes in renal tissues
(98). A previous study has
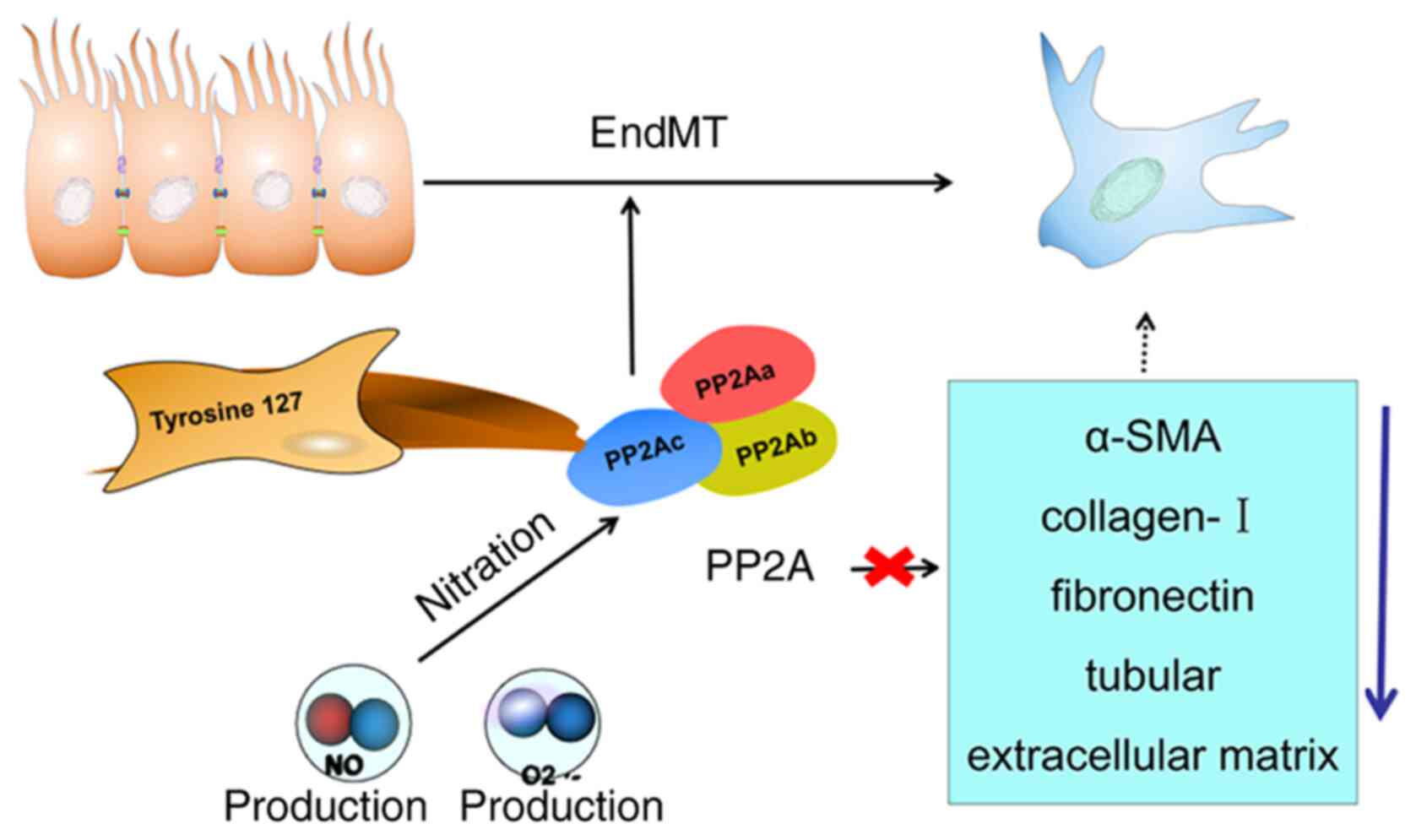
indicated that Tyr nitration in the C subunit of PP2A decreases
PP2Ac tyrosine phosphorylation and increases PP2A activity and
endothelial dysfunction (99).
TGF-β1-induced nitrification accelerates the nitrification of PP2Ac
and increases the activity of PP2A in endothelial cells (10). Okadaic acid inhibits the activity of
PP2A, weakening the effects of PP2A on EC cytoskeletal
rearrangement induced by thrombin or nocodazole (100). This finding indicates that PP2A
activity serves an important role in the maintenance of the EC
cytoskeleton. Endothelial-mesenchymal transition (EndMT) is a major
cellular behavioral mechanism that aims to increase the production
of myofibroblasts (101), which
are involved in the pathogenesis and progression of renal fibrosis
(102-105).
Furthermore, EndMT serves a key role in the development of CKD
(106,107).
PP2A activation occurs in mouse unilateral ureteral
obstruction and TGF-β1-treated human umbilical vein endothelial
cells (HUVECs) in vitro (10). Additionally, OA significantly
inhibits the expression of α-smooth muscle actin (a fibroblast
marker), which is induced by TGF-β1 and maintains the expression of
VE cadherin in HUVECs (10).
Furthermore, TGF-β1 decreases the abundance of phosphorylated
serine and threonine residues in occludin immunoprecipitates, which
is significantly inhibited by pretreatment with OA (10). Erythrocyte sphingosine 1-phosphate
serves a beneficial role in CKD by promoting the activity of
2,3-BPG (an erythrocyte-specific metabolite that negatively
regulates the binding affinity of hemoglobin-O2) and subsequently
triggering O2 delivery to renal cells and tissues
(108). These events counteract
hypoxia-induced kidney damage and slow CKD progression by
inhibiting PP2A activity (108).
PP2Ac serves an important role in the dephosphorylation activity of
PP2A (25), and it has been
demonstrated that inhibition of PP2Ac by OA attenuates renal
fibrosis by suppressing fibronectin and collagen I expression
(109). These events reverse
epithelial-mesenchymal transition in renal tubules, while ectopic
overexpression of PP2Ac accelerates tubular extracellular matrix
(ECM) accumulation in vitro (109) (Fig.
2). Additionally, nitration of the PP2Ac tyrosine is crucial
for PP2A activation during EndMT (10). Wu and Wilson (99) suggested that microvascular
endothelial cells produce peroxynitrite, which nitrates PP2Ac under
proinflammatory stimuli. This nitration enhances the activity of
PP2A in the mediation of endothelial barrier dysfunction.
Inhibition of NO and O2 production by
NG-nitro-L-arginine methyl ester (an inhibitor of NO synthetases)
and apocynin (APO, a specific inhibitor of NADPH oxidase) decreases
the nitrification of PP2Ac and suppresses the process of EndMT via
induction of TGF-β1 activity (99).
Furthermore, tyrosine 127 (Tyr127) is essential for PP2Ac
nitration, while inhibiting PP2Ac Tyr127 nitration with the peptide
TAT-Y127WT effectively decreases PP2Ac nitration and ameliorates
ECM deposition and capillary rarefaction (Fig. 2) (110). These results indicate that PP2Ac
may be a novel drug target that could be used in anti-fibrotic
therapies.
CKD is not only the main risk factor in acute
myocardial infarction (AMI) but also an important factor in the
reduced survival rate of patients with AMI (111-113).
CKD downregulates PP2A-B55α protein expression, resulting in
upregulated Akt-Thr308 phosphorylation (114). Decreased Akt activation, which is
impaired by insufficient phosphorylation of ser473 during
reperfusion, contributes to enlargement of myocardial infarct foci
(114). These results indicate
that PP2A activates the process of EndMT, while blocking PP2A
signaling can inhibit this process. Therefore, inhibition of PP2A
activity to prevent endothelial-mesenchymal transition is a
promising strategy that could be used in anti-CKD therapies.
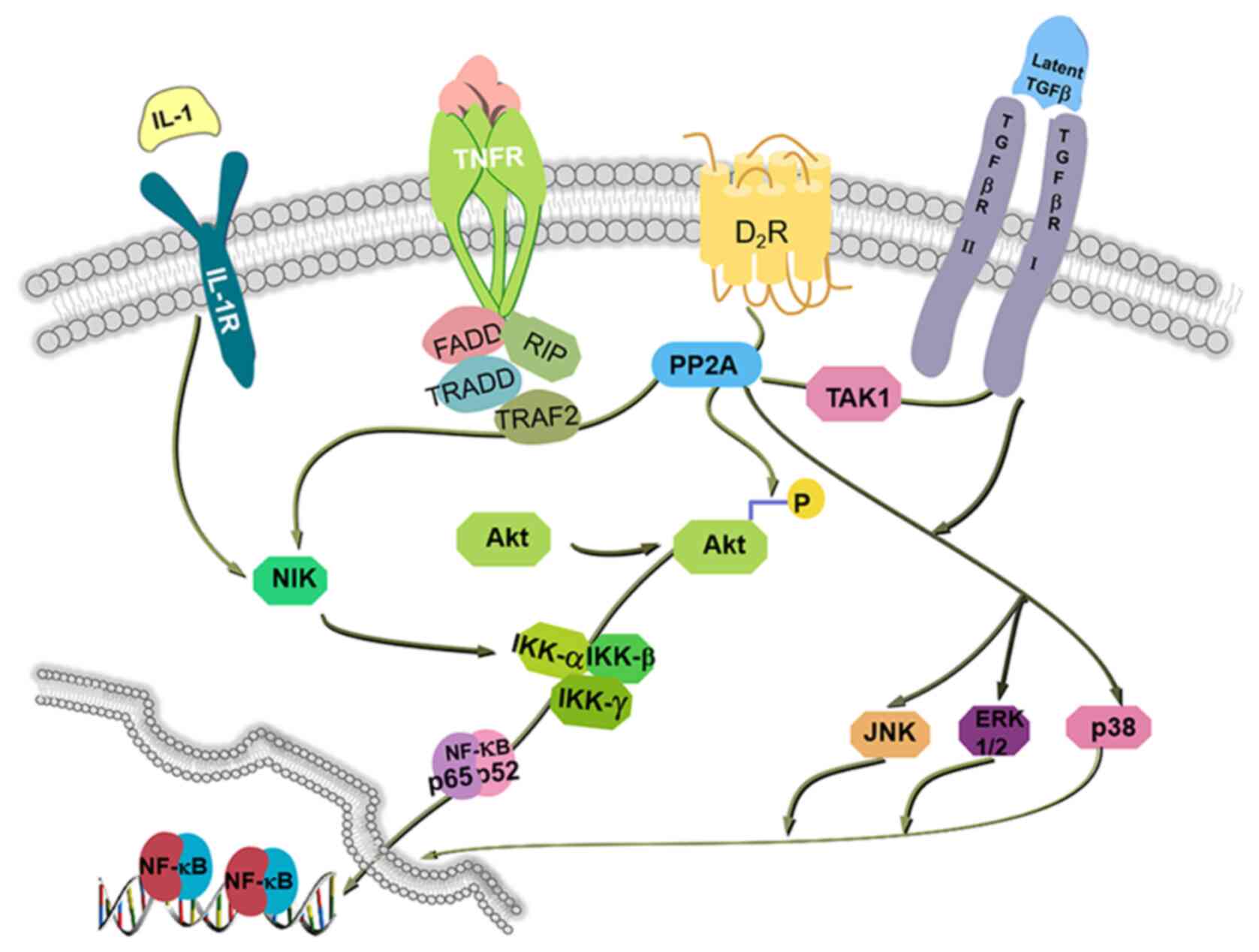
Inflammation, which is the process of detecting and
eliminating harmful pathogens, is the main pathogenic mechanism of
CKD and AKI (115). PP2A has been
identified as an effective negative regulator of a number of
different inflammatory signaling pathways (116,117), including dephosphorylation and
inhibition of p65 NF-κB (118,119). An increase in PP2A activity by ATG
attenuates the effects of NF-κB-mediated inflammation and enhances
the stability of podocyte actin cytoskeleton via DBN1
dephosphorylation (79). The renal
dopaminergic system is involved in the regulation of ROS production
and the inflammatory response (120-125).
A previous study indicated that dopamine D2 receptor (D2R) controls
inflammation in the kidneys by modulating Akt dephosphorylation via
PP2A (126). The increased PP2A
activity inhibits increases in NF-κB activity, which are induced by
D2R silencing (126). PP2A is also
a major cellular phosphatase that can potentially contribute to
NF-κB activity (127). In these
events, PP2A regulates at least three different pathways, including
TNF receptor-associated factor 2, IKK and NF-κB p65 during the
downregulation of NF-κB activity (128). Inactivation of PP2A via a PTK/PTP
imbalance triggered by oxidative stress, causes NF-κB activation,
which contributes to the accumulation of oxidative stress in aged
rat kidneys (11). Inactivation or
knockdown of PP2A triggers activation of NF-κB signaling
components, such as NIK/IKK and MAPKs (ERK, p38, and JNK), leading
to NF-κB activation (11). PP2A is
also a negative regulator of TGF-β1-activated kinase 1 activation
in cultured mesangial cells (129). Studies on senescence marker
protein-30 (SMP30) have indicated that blocking the inactivation of
PP1 and PP2A via oxidative stress leads to the activation of NF-κB
in the kidneys of SMP30Y/-mice (130). Therefore, PP2A regulation of NF-κB
activity may provide a novel therapeutic target for the development
of anti-inflammatory therapies in patients with kidney disease.
Fig. 3 illustrates how PP2A
regulates NF-κB by participating in a range of cellular and
molecular mechanisms involved in renal inflammation.
The current understanding of the structure and
function of PP2A is extensive, and links between PP2A and renal
diseases are becoming increasingly clear. PP2A serves diverse roles
in renal pathophysiology, and normal PP2A activity is required for
regulating ion channels to maintain the homeostasis of
Na+, K+ and Cl- (49). PP2A also regulates the
phosphorylation of AQP2 and its membrane accumulations to adjust
urine concentration (63). Normal
PP2A activity is critical not only in the formation and function of
podocytes (21) but also in renal
morphogenesis and development (40). It should be noted that patients with
RCC who exhibit a higher PP2A expression have a higher chance of
survival (12). PP2A expression may
be useful in predicting patient prognosis and therapeutic outcomes
in patients with RCC. PP2A has also been identified to be an
antifibrotic factor and may be a promising therapeutic target in
the treatment of renal fibrosis (23). In renal inflammation, PP2A is
associated with a number of signaling pathways that regulate NF-κB
activity (11,40,118,128). As PP2A serves numerous versatile
roles in the pathogenic progression of kidney diseases, targeting
PP2A in future therapeutics may improve patient outcomes. However,
future studies should further examine the mechanisms of PP2A in the
progression of kidney diseases.
Not applicable.
Funding: The current study was supported by the National Natural
Science Foundation of China (grant no. 8186030172), the Yunnan
Provincial Science and Technology Department Funds (grant no.
2020J0614), the 2018 TEN-HUNDREND-THOUSAND Project to Reserve
Talent Fund Program in Kunming (grant no. 2018-SW-018) and the
Yunnan Provincial Science and Technology Department Funds (grant
no. 2019Y0360).
Not applicable.
SW, JW and LY were involved in study conception and
interpretation, writing and critically revising the manuscript. LS,
YM and ZH wrote the manuscript. QF was involved with conception and
design of the figures. All authors read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Klein J and Schanstra JP: Implementation
of proteomics biomarkers in nephrology: From animal models to human
application? Proteomics Clin Appl. 13(e1800089)2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Ferenbach DA and Bonventre JV: Acute
kidney injury and chronic kidney disease: From the laboratory to
the clinic. Nephrol Ther. 12 (Suppl 1):S41–S48. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ferenbach DA and Bonventre JV: Mechanisms
of maladaptive repair after AKI leading to accelerated kidney
ageing and CKD. Nat Rev Nephrol. 11:264–276. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Eirin A, Lerman A and Lerman LO: The
emerging role of mitochondrial targeting in kidney disease. Handb
Exp Pharmacol. 240:229–250. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Jha V, Garcia-Garcia G, Iseki K, Li Z,
Naicker S, Plattner B, Saran R, Wang AY and Yang CW: Chronic kidney
disease: Global dimension and perspectives. Lancet. 382:260–272.
2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Chertow GM, Burdick E, Honour M, Bonventre
JV and Bates DW: Acute kidney injury, mortality, length of stay,
and costs in hospitalized patients. J Am Soc Nephrol. 16:3365–3370.
2005.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Mumby MC and Walter G: Protein
serine/threonine phosphatases: Structure, regulation, and functions
in cell growth. Physiol Rev. 73:673–699. 1993.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Eichhorn PJ, Creyghton MP and Bernards R:
Protein phosphatase 2A regulatory subunits and cancer. Biochim
Biophys Acta. 1795:1–15. 2009.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Tsao CC, Nica AF, Kurinna SM, Jiffar T,
Mumby M and Ruvolo PP: Mitochondrial protein phosphatase 2A
regulates cell death induced by simulated ischemia in kidney
NRK-52E cells. Cell Cycle. 6:2377–2385. 2007.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Deng Y, Guo Y, Liu P, Zeng R, Ning Y, Pei
G, Li Y, Chen M, Guo S, Li X, et al: Blocking protein phosphatase
2A signaling prevents endothelial-to-mesenchymal transition and
renal fibrosis: A peptide-based drug therapy. Sci Rep.
6(19821)2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jin Jung K, Hyun Kim D, Kyeong Lee E, Woo
Song C, Pal Yu B and Young Chung H: Oxidative stress induces
inactivation of protein phosphatase 2A, promoting proinflammatory
NF-kappaB in aged rat kidney. Free Radic Biol Med. 61:206–217.
2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Li J, Sheng C, Li W and Zheng JH: Protein
phosphatase-2A is down-regulated in patients within clear cell
renal cell carcinoma. Int J Clin Exp Pathol. 7:1147–1153.
2014.PubMed/NCBI
|
|
13
|
Sen CK: Cellular thiols and
redox-regulated signal transduction. Curr Top Cell Regul. 36:1–30.
2000.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Barik S: Protein phosphorylation and
signal transduction. Subcell Biochem. 26:115–164. 1996.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Shi Y: Serine/threonine phosphatases:
Mechanism through structure. Cell. 139:468–484. 2009.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Barford D, Das AK and Egloff MP: The
structure and mechanism of protein phosphatases: Insights into
catalysis and regulation. Annu Rev Biophys Biomol Struct.
27:133–164. 1998.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Barford D: Colworth medal lecture.
Structural studies of reversible protein phosphorylation and
protein phosphatases. Biochem Soc Trans. 27:751–766.
1999.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Hornbeck PV, Kornhauser JM, Tkachev S,
Zhang B, Skrzypek E, Murray B, Latham V and Sullivan M:
PhosphoSitePlus: A comprehensive resource for investigating the
structure and function of experimentally determined
post-translational modifications in man and mouse. Nucleic Acids
Res. 40:D261–D270. 2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Geraldes P: Protein phosphatases and
podocyte function. Curr Opin Nephrol Hypertens. 27:49–55.
2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kumar S and Tikoo K: Independent role of
PP2A and mTORc1 in palmitate induced podocyte death. Biochimie.
112:73–84. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Reiser J, Pixley FJ, Hug A, Kriz W, Smoyer
WE, Stanley ER and Mundel P: Regulation of mouse podocyte process
dynamics by protein tyrosine phosphatases rapid communication.
Kidney Int. 57:2035–2042. 2000.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Svennilson J, Durbeej M, Celsi G,
Laestadius A, da Cruz e Silva EF, Ekblom P and Aperia A: Evidence
for a role of protein phosphatases 1 and 2A during early
nephrogenesis. Kidney Int. 48:103–110. 1995.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Everett AD, Xue C and Stoops T:
Developmental expression of protein phosphatase 2A in the kidney. J
Am Soc Nephrol. 10:1737–1745. 1999.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Gotz J, Probst A, Mistl C, Nitsch RM and
Ehler E: Distinct role of protein phosphatase 2A subunit Calpha in
the regulation of E-cadherin and beta-catenin during development.
Mech Dev. 93:83–93. 2000.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Mumby M: The 3D structure of protein
phosphatase 2A: New insights into a ubiquitous regulator of cell
signaling. ACS Chem Biol. 2:99–103. 2007.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Janssens V and Goris J: Protein
phosphatase 2A: A highly regulated family of serine/threonine
phosphatases implicated in cell growth and signalling. Biochem J.
353:417–439. 2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Turowski P, Favre B, Campbell KS, Lamb NJ
and Hemmings BA: Modulation of the enzymatic properties of protein
phosphatase 2A catalytic subunit by the recombinant 65-kDa
regulatory subunit PR65alpha. Eur J Biochem. 248:200–208.
1997.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sontag E: Protein phosphatase 2A: The
Trojan horse of cellular signaling. Cell Signal. 13:7–16.
2001.PubMed/NCBI View Article : Google Scholar
|
|
29
|
O'Connor CM, Perl A, Leonard D, Sangodkar
J and Narla G: Therapeutic targeting of PP2A. Int J Biochem Cell
Biol. 96:182–193. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Forester CM, Maddox J, Louis JV, Goris J
and Virshup DM: Control of mitotic exit by PP2A regulation of
Cdc25C and Cdk1. Proc Natl Acad Sci USA. 104:19867–19872.
2007.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Slupe AM, Merrill RA and Strack S:
Determinants for substrate specificity of protein phosphatase 2A.
Enzyme Res. 2011(398751)2011.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Flegg CP, Sharma M, Medina-Palazon C,
Jamieson C, Galea M, Brocardo MG, Mills K and Henderson BR: Nuclear
export and centrosome targeting of the protein phosphatase 2A
subunit B56alpha: Role of B56alpha in nuclear export of the
catalytic subunit. J Biol Chem. 285:18144–18154. 2010.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Bononi A, Agnoletto C, De Marchi E, Marchi
S, Patergnani S, Bonora M, Giorgi C, Missiroli S, Poletti F,
Rimessi A and Pinton P: Protein kinases and phosphatases in the
control of cell fate. Enzyme Res. 2011(329098)2011.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Riedel CG, Katis VL, Katou Y, Mori S, Itoh
T, Helmhart W, Gálová M, Petronczki M, Gregan J and Cetin B:
Protein phosphatase 2A protects centromeric sister chromatid
cohesion during meiosis I. Nature. 441:53–61. 2006.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Jin Z, Shi J, Saraf A, Mei W, Zhu GZ,
Strack S and Yang J: The 48-kDa alternative translation isoform of
PP2A:B56epsilon is required for Wnt signaling during
midbrain-hindbrain boundary formation. J Biol Chem. 284:7190–7200.
2009.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Seshacharyulu P, Pandey P, Datta K and
Batra SK: Phosphatase: PP2A structural importance, regulation and
its aberrant expression in cancer. Cancer Lett. 335:9–18.
2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Kanno T, Tsuchiya A, Shimizu T, Nakao S,
Tanaka A and Nishizaki T: Effects of newly synthesized
DCP-LA-phospholipids on protein kinase C and protein phosphatases.
Cell Physiol Biochem. 31:555–564. 2013.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Kurimchak A and Grana X: PP2A
counterbalances phosphorylation of pRB and mitotic proteins by
multiple CDKs: Potential implications for PP2A disruption in
cancer. Genes Cancer. 3:739–748. 2012.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Svennilson J, Sandberg-Nordqvist A and
Aperia A: Age-dependent expression of protein phosphatase 2A in the
developing rat kidney. Pediatr Nephrol. 13:800–805. 1999.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Yang J, Wu J, Tan C and Klein PS:
PP2A:B56epsilon is required for Wnt/beta-catenin signaling during
embryonic development. Development. 130:5569–5578. 2003.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Jørgensen PL: Sodium and potassium ion
pump in kidney tubules. Physiol Rev. 60:864–917. 1980.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Rangel LB, Lopes AG, Lara LS, Carvalho TL,
Silva IV, Oliveira MM, Einicker-Lamas M, Vieyra A, Nogaroli L and
Caruso-Neves C: PI-PLCbeta is involved in the modulation of the
proximal tubule Na+-ATPase by angiotensin II. Regul Pept.
127:177–182. 2005.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Gates J Jr, Ferguson SM, Blakely RD and
Apparsundaram S: Regulation of choline transporter surface
expression and phosphorylation by protein kinase C and protein
phosphatase 1/2A. J Pharmacol Exp Ther. 310:536–545.
2004.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Vieira-Filho LD, Cabral EV, Farias JS,
Silva PA, Muzi-Filho H, Vieyra A and Paixão AD: Renal molecular
mechanisms underlying altered Na+ handling and genesis of
hypertension during adulthood in prenatally undernourished rats. Br
J Nutr. 111:1932–1944. 2014.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Dias J, Ferrao FM, Axelband F, Carmona AK,
Lara LS and Vieyra A: ANG-(3-4) inhibits renal
Na+-ATPase in hypertensive rats through a mechanism that
involves dissociation of ANG II receptors, heterodimers, and PKA.
Am J Physiol Renal Physiol. 306:F855–F863. 2014.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Vieira-Filho LD, Lara LS, Silva PA, Santos
FT, Luzardo R, Oliveira FS, Paixão AD and Vieyra A: Placental
malnutrition changes the regulatory network of renal Na-ATPase in
adult rat progeny: Reprogramming by maternal α-tocopherol during
lactation. Arch Biochem Biophys. 505:91–97. 2011.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Silva PA, Muzi-Filho H, Pereira-Acacio A,
Dias J, Martins JF, Landim-Vieira M, Verdoorn KS, Lara LS,
Vieira-Filho LD, Cabral EV, et al: Altered signaling pathways
linked to angiotensin II underpin the upregulation of renal
Na(+)-ATPase in chronically undernourished rats. Biochim Biophys
Acta. 1842:2357–2366. 2014.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Gildea JJ, Xu P, Kemp BA, Carey RM, Jose
PA and Felder RA: The dopamine D1 receptor and angiotensin II
type-2 receptor are required for inhibition of sodium transport
through a protein phosphatase 2A pathway. Hypertension.
73:1258–1265. 2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Lecuona E, Garcia A and Sznajder JI: A
novel role for protein phosphatase 2A in the dopaminergic
regulation of Na, K-ATPase. FEBS Lett. 481:217–220. 2000.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Li D, Cheng SX, Fisone G, Caplan MJ,
Ohtomo Y and Aperia A: Effects of okadaic acid, calyculin A, and
PDBu on state of phosphorylation of rat renal
Na+-K+-ATPase. Am J Physiol. 275:F863–F869.
1998.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Tiran Z, Peretz A, Sines T, Shinder V, Sap
J, Attali B and Elson A: Tyrosine phosphatases epsilon and alpha
perform specific and overlapping functions in regulation of
voltage-gated potassium channels in Schwann cells. Mol Biol Cell.
17:4330–4342. 2006.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Capdevila J and Wang W: Role of cytochrome
P450 epoxygenase in regulating renal membrane transport and
hypertension. Curr Opin Nephrol Hypertens. 22:163–169.
2013.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Nielsen S and Agre P: The aquaporin family
of water channels in kidney. Kidney Int. 48:1057–1068.
1995.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Christensen BM, Zelenina M, Aperia A and
Nielsen S: Localization and regulation of PKA-phosphorylated AQP2
in response to V(2)-receptor agonist/antagonist treatment. Am J
Physiol Renal Physiol. 278:F29–F42. 2000.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Hoffert JD, Pisitkun T, Wang G, Shen RF
and Knepper MA: Quantitative phosphoproteomics of
vasopressin-sensitive renal cells: Regulation of aquaporin-2
phosphorylation at two sites. Proc Natl Acad Sci USA.
103:7159–7164. 2006.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Tamma G, Robben JH, Trimpert C, Boone M
and Deen PM: Regulation of AQP2 localization by S256 and S261
phosphorylation and ubiquitination. Am J Physiol Cell Physiol.
300:C636–C646. 2011.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Moeller HB, Knepper MA and Fenton RA:
Serine 269 phosphorylated aquaporin-2 is targeted to the apical
membrane of collecting duct principal cells. Kidney Int.
75:295–303. 2009.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Brown D: The ins and outs of aquaporin-2
trafficking. Am J Physiol Renal Physiol. 284:F893–F901.
2003.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Kamsteeg EJ, Heijnen I, van Os CH and Deen
PM: The subcellular localization of an aquaporin-2 tetramer depends
on the stoichiometry of phosphorylated and nonphosphorylated
monomers. J Cell Biol. 151:919–930. 2000.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Hoffert JD, Fenton RA, Moeller HB, Simons
B, Tchapyjnikov D, McDill BW, Yu MJ, Pisitkun T, Chen F and Knepper
MA: Vasopressin-stimulated increase in phosphorylation at Ser269
potentiates plasma membrane retention of aquaporin-2. J Biol Chem.
283:24617–24627. 2008.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Ren H, Yang B, Ruiz JA, Efe O, Ilori TO,
Sands JM and Klein JD: Phosphatase inhibition increases AQP2
accumulation in the rat IMCD apical plasma membrane. Am J Physiol
Renal Physiol. 311:F1189–F1197. 2016.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Tamma G, Lasorsa D, Trimpert C, Ranieri M,
Di Mise A, Mola MG, Mastrofrancesco L, Devuyst O, Svelto M, Deen PM
and Valenti G: A protein kinase A-independent pathway controlling
aquaporin 2 trafficking as a possible cause for the syndrome of
inappropriate antidiuresis associated with polycystic kidney
disease 1 haploinsufficiency. J Am Soc Nephrol. 25:2241–2253.
2014.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Valenti G, Procino G, Carmosino M, Frigeri
A, Mannucci R, Nicoletti I and Svelto M: The phosphatase inhibitor
okadaic acid induces AQP2 translocation independently from AQP2
phosphorylation in renal collecting duct cells. J Cell Sci.
113:1985–1992. 2000.PubMed/NCBI
|
|
64
|
Millward TA, Zolnierowicz S and Hemmings
BA: Regulation of protein kinase cascades by protein phosphatase
2A. Trends Biochem Sci. 24:186–191. 1999.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Lee TH, Solomon MJ, Mumby MC and Kirschner
MW: INH, a negative regulator of MPF, is a form of protein
phosphatase 2A. Cell. 64:415–423. 1991.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Izumi T, Walker DH and Maller JL: Periodic
changes in phosphorylation of the Xenopus cdc25 phosphatase
regulate its activity. Mol Biol Cell. 3:927–939. 1992.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Haystead TA, Weiel JE, Litchfield DW,
Tsukitani Y, Fischer EH and Krebs EG: Okadaic acid mimics the
action of insulin in stimulating protein kinase activity in
isolated adipocytes. The role of protein phosphatase 2a in
attenuation of the signal. J Biol Chem. 265:16571–16580.
1990.PubMed/NCBI
|
|
68
|
Tsao H and Greene LA: The roles of
macromolecular synthesis and phosphorylation in the regulation of a
protein kinase activity transiently stimulated by nerve growth
factor. J Biol Chem. 266:12981–12988. 1991.PubMed/NCBI
|
|
69
|
Stark K, Vainio S, Vassileva G and McMahon
AP: Epithelial transformation of metanephric mesenchyme in the
developing kidney regulated by Wnt-4. Nature. 372:679–683.
1994.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Zeng L, Fagotto F, Zhang T, Hsu W, Vasicek
TJ, Perry WL III, Lee JJ, Tilghman SM, Gumbiner BM and Costantini
F: The mouse Fused locus encodes Axin, an inhibitor of the Wnt
signaling pathway that regulates embryonic axis formation. Cell.
90:181–192. 1997.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Altintas MM and Reiser J: Bridges to
cross, burn, and mend: Cells of renin lineage as podocyte
progenitors. Am J Physiol Renal Physiol. 309:F499–F500.
2015.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Pagtalunan ME, Miller PL, Jumping-Eagle S,
Nelson RG, Myers BD, Rennke HG, Coplon NS, Sun L and Meyer TW:
Podocyte loss and progressive glomerular injury in type II
diabetes. J Clin Invest. 99:342–348. 1997.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Huber TB and Benzing T: The slit
diaphragm: A signaling platform to regulate podocyte function. Curr
Opin Nephrol Hypertens. 14:211–216. 2005.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Sedor JR, Madhavan SM, Kim JH and
Konieczkowski M: Out on a LIM: Chronic kidney disease, podocyte
phenotype and the Wilm's tumor interacting protein (WTIP). Trans Am
Clin Climatol Assoc. 122:184–197. 2011.PubMed/NCBI
|
|
75
|
Liu M, Liang K, Zhen J, Zhou M, Wang X,
Wang Z, Wei X, Zhang Y, Sun Y, Zhou Z, et al: Sirt6 deficiency
exacerbates podocyte injury and proteinuria through targeting Notch
signaling. Nat Commun. 8(413)2017.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Kobayashi N, Reiser J, Schwarz K, Sakai T,
Kriz W and Mundel P: Process formation of podocytes: Morphogenetic
activity of microtubules and regulation by protein serine/threonine
phosphatase PP2A. Histochem Cell Biol. 115:255–266. 2001.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Zhu X, Ye Y, Xu C, Gao C, Zhang Y, Zhou J,
Lin W and Mao J: Protein phosphatase 2A modulates podocyte
maturation and glomerular functional integrity in mice. Cell Commun
Signal. 17(91)2019.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Hanssen L, Frye BC, Ostendorf T, Alidousty
C, Djudjaj S, Boor P, Rauen T, Floege J, Mertens PR and Raffetseder
U: Y-box binding protein-1 mediates profibrotic effects of
calcineurin inhibitors in the kidney. J Immunol. 187:298–308.
2011.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Zhong Y, Lee K, Deng Y, Ma Y, Chen Y, Li
X, Wei C, Yang S, Wang T, Wong NJ, et al: Arctigenin attenuates
diabetic kidney disease through the activation of PP2A in
podocytes. Nat Commun. 10(4523)2019.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386.
2015.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Qiu M, Liu L, Chen L, Tan G, Liang Z, Wang
K, Liu J and Chen H: MicroRNA-183 plays as oncogenes by increasing
cell proliferation, migration and invasion via targeting protein
phosphatase 2A in renal cancer cells. Biochem Biophys Res Commun.
452:163–169. 2014.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Liu P, Xiang Y, Liu X, Zhang T, Yang R,
Chen S, Xu L, Yu Q, Zhao H, Zhang L, et al: Cucurbitacin B induces
the lysosomal degradation of EGFR and suppresses the CIP2A/PP2A/Akt
signaling axis in gefitinib-resistant non-small cell lung cancer.
Molecules. 24(647)2019.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Cairns J, Ly RC, Niu N, Kalari KR, Carlson
EE and Wang L: CDC25B partners with PP2A to induce AMPK activation
and tumor suppression in triple negative breast cancer. NAR Cancer.
2(zcaa39)2020.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Vicente C, Arriazu E, Martínez-Balsalobre
E, Peris I, Marcotegui N, García-Ramírez P, Pippa R, Rabal O,
Oyarzábal J, Guruceaga E, et al: A novel FTY720 analogue targets
SET-PP2A interaction and inhibits growth of acute myeloid leukemia
cells without inducing cardiac toxicity. Cancer Lett. 468:1–13.
2020.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Westermarck J and Hahn WC: Multiple
pathways regulated by the tumor suppressor PP2A in transformation.
Trends Mol Med. 14:152–160. 2008.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Xing ML, Lu YF, Wang DF, Zou XY, Zhang SX
and Yun Z: Clinical significance of sCIP2A levels in breast cancer.
Eur Rev Med Pharmacol Sci. 20:82–91. 2016.PubMed/NCBI
|
|
87
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 101:2999–3004. 2004.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Kauko O and Westermarck J: Non-genomic
mechanisms of protein phosphatase 2A (PP2A) regulation in cancer.
Int J Biochem Cell Biol. 96:157–164. 2018.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Lambrecht C, Haesen D, Sents W, Ivanova E
and Janssens V: Structure, regulation, and pharmacological
modulation of PP2A phosphatases. Methods Mol Biol. 1053:283–305.
2013.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Ou YC, Li JR, Wang JD, Chen WY, Kuan YH,
Yang CP, Liao SL, Lu HC and Chen CJ: Aspirin restores
ABT-737-mediated apoptosis in human renal carcinoma cells. Biochem
Biophys Res Commun. 502:187–193. 2018.PubMed/NCBI View Article : Google Scholar
|
|
91
|
de Fatima A, Zambuzzi WF, Modolo LV,
Tarsitano CA, Gadelha FR, Hyslop S, de Carvalho JE, Salgado I,
Ferreira CV and Pilli RA: Cytotoxicity of goniothalamin enantiomers
in renal cancer cells: Involvement of nitric oxide, apoptosis and
autophagy. Chem Biol Interact. 176:143–150. 2008.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Seo SU, Woo SM, Min KJ and Kwon TK:
Z-FL-COCHO, a cathepsin S inhibitor, enhances oxaliplatin-induced
apoptosis through upregulation of Bim expression. Biochem Biophys
Res Commun. 498:849–854. 2018.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Tsai YT, Chuang MJ, Tang SH, Wu ST, Chen
YC, Sun GH, Hsiao PW, Huang SM, Lee HJ, Yu CP, et al: Novel cancer
therapeutics with allosteric modulation of the mitochondrial
C-Raf-DAPK complex by raf inhibitor combination therapy. Cancer
Res. 75:3568–3582. 2015.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Ou YC, Kuan YH, Li JR, Raung SL, Wang CC,
Hung YY and Chen CJ: Induction of apoptosis by luteolin involving
akt inactivation in human 786-o renal cell carcinoma cells. Evid
Based Complement Alternat Med. 2013(109105)2013.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Liu Y: Renal fibrosis: New insights into
the pathogenesis and therapeutics. Kidney Int. 69:213–217.
2006.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Daehn I and Bottinger EP: Microvascular
endothelial cells poised to take center stage in experimental renal
fibrosis. J Am Soc Nephrol. 26:767–769. 2015.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Bohle A, Mackensen-Haen S and Wehrmann M:
Significance of postglomerular capillaries in the pathogenesis of
chronic renal failure. Kidney Blood Press Res. 19:191–195.
1996.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Fine LG, Orphanides C and Norman JT:
Progressive renal disease: The chronic hypoxia hypothesis. Kidney
Int Suppl. 65 (Suppl):S74–S78. 1998.PubMed/NCBI
|
|
99
|
Wu F and Wilson JX:
Peroxynitrite-dependent activation of protein phosphatase type 2A
mediates microvascular endothelial barrier dysfunction. Cardiovasc
Res. 81:38–45. 2009.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Kása A, Czikora I, Verin AD, Gergely P and
Csortos C: Protein phosphatase 2A activity is required for
functional adherent junctions in endothelial cells. Microvasc Res.
89:86–94. 2013.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Kriz W, Kaissling B and Le Hir M:
Epithelial-mesenchymal transition (EMT) in kidney fibrosis: Fact or
fantasy? J Clin Invest. 121:468–474. 2011.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Lipphardt M, Dihazi H, Jeon NL, Dadafarin
S, Ratliff BB, Rowe DW, Müller GA and Goligorsky MS: Dickkopf-3 in
aberrant endothelial secretome triggers renal fibroblast activation
and endothelial-mesenchymal transition. Nephrol Dial Transplant.
34:49–62. 2019.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Zeisberg EM, Potenta SE, Sugimoto H,
Zeisberg M and Kalluri R: Fibroblasts in kidney fibrosis emerge via
endothelial-to-mesenchymal transition. J Am Soc Nephrol.
19:2282–2287. 2008.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Matsumoto K, Xavier S, Chen J, Kida Y,
Lipphardt M, Ikeda R, Gevertz A, Caviris M, Hatzopoulos AK,
Kalajzic I, et al: Instructive role of the microenvironment in
preventing renal fibrosis. Stem Cells Transl Med. 6:992–1005.
2017.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Chen CL, Chou KJ, Fang HC, Hsu CY, Huang
WC, Huang CW, Huang CK, Chen HY and Lee PT: Progenitor-like cells
derived from mouse kidney protect against renal fibrosis in a
remnant kidney model via decreased endothelial mesenchymal
transition. Stem Cell Res Ther. 6(239)2015.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Xavier S, Vasko R, Matsumoto K, Zullo JA,
Chen R, Maizel J, Chander PN and Goligorsky MS: Curtailing
endothelial TGF-β signaling is sufficient to reduce
endothelial-mesenchymal transition and fibrosis in CKD. J Am Soc
Nephrol. 26:817–829. 2015.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Li J, Qu X and Bertram JF:
Endothelial-myofibroblast transition contributes to the early
development of diabetic renal interstitial fibrosis in
streptozotocin-induced diabetic mice. Am J Pathol. 175:1380–1388.
2009.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Xie T, Chen C, Peng Z, Brown BC, Reisz JA,
Xu P, Zhou Z, Song A, Zhang Y, Bogdanov MV, et al: Erythrocyte
metabolic reprogramming by sphingosine 1-phosphate in chronic
kidney disease and therapies. Circ Res. 127:360–375.
2020.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Hou T, Xiao Z, Li Y, You YH, Li H, Liu YP,
Xi YY, Li J, Duan SB, Liu H, et al: Norcantharidin inhibits renal
interstitial fibrosis by downregulating PP2Ac expression. Am J
Transl Res. 7:2199–2211. 2015.PubMed/NCBI
|
|
110
|
Deng Y, Cai Y, Liu L, Lin X, Lu P, Guo Y,
Han M and Xu G: Blocking Tyr265 nitration of protein phosphatase 2A
attenuates nitrosative stress-induced endothelial dysfunction in
renal microvessels. FASEB J. 33:3718–3730. 2019.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Wright RS, Reeder GS, Herzog CA, Albright
RC, Williams BA, Dvorak DL, Miller WL, Murphy JG, Kopecky SL and
Jaffe AS: Acute myocardial infarction and renal dysfunction: A
high-risk combination. Ann Intern Med. 137:563–570. 2002.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Rodrigues FB, Bruetto RG, Torres US,
Otaviano AP, Zanetta DM and Burdmann EA: Effect of kidney disease
on acute coronary syndrome. Clin J Am Soc Nephrol. 5:1530–1536.
2010.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Barnes JL and Glass Ii WF: Renal
interstitial fibrosis: A critical evaluation of the origin of
myofibroblasts. Contrib Nephrol. 169:73–93. 2011.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Tobisawa T, Yano T, Tanno M, Miki T, Kuno
A, Kimura Y, Ishikawa S, Kouzu H, Nishizawa K, Yoshida H and Miura
T: Insufficient activation of Akt upon reperfusion because of its
novel modification by reduced PP2A-B55α contributes to enlargement
of infarct size by chronic kidney disease. Basic Res Cardiol.
112(31)2017.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Sato Y and Yanagita M: Immune cells and
inflammation in AKI to CKD progression. Am J Physiol Renal Physiol.
315:F1501–F1512. 2018.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Rahman MM, Rumzhum NN, Morris JC, Clark
AR, Verrills NM and Ammit AJ: Basal protein phosphatase 2A activity
restrains cytokine expression: Role for MAPKs and tristetraprolin.
Sci Rep. 5(10063)2015.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Crispin JC, Apostolidis SA, Rosetti F,
Keszei M, Wang N, Terhorst C, Mayadas TN and Tsokos GC: Cutting
edge: Protein phosphatase 2A confers susceptibility to autoimmune
disease through an IL-17-dependent mechanism. J Immunol.
188:3567–3571. 2012.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Hsieh CY, Hsiao G, Hsu MJ, Wang YH and
Sheu JR: PMC, a potent hydrophilic α-tocopherol derivative,
inhibits NF-κB activation via PP2A but not IKBα-dependent signals
in vascular smooth muscle cells. J Cell Mol Med. 18:1278–1289.
2014.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Yang J, Fan GH, Wadzinski BE, Sakurai H
and Richmond A: Protein phosphatase 2A interacts with and directly
dephosphorylates RelA. J Biol Chem. 276:47828–47833.
2001.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Zhang Y, Cuevas S, Asico LD, Escano C,
Yang Y, Pascua AM, Wang X, Jones JE, Grandy D, Eisner G, et al:
Deficient dopamine D2 receptor function causes renal inflammation
independently of high blood pressure. PLoS One.
7(e38745)2012.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Asghar M, Chugh G and Lokhandwala MF:
Inflammation compromises renal dopamine D1 receptor function in
rats. Am J Physiol Renal Physiol. 297:F1543–F1549. 2009.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Yang S, Yao B, Zhou Y, Yin H, Zhang MZ and
Harris RC: Intrarenal dopamine modulates progressive angiotensin
II-mediated renal injury. Am J Physiol Renal Physiol.
302:F742–F749. 2012.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Chugh G, Lokhandwala MF and Asghar M:
Oxidative stress alters renal D1 and AT1 receptor functions and
increases blood pressure in old rats. Am J Physiol Renal Physiol.
300:F133–F138. 2011.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Yang Y, Zhang Y, Cuevas S, Villar VA,
Escano C, D Asico L, Yu P, Grandy DK, Felder RA, Armando I and Jose
PA: Paraoxonase 2 decreases renal reactive oxygen species
production, lowers blood pressure, and mediates dopamine D2
receptor-induced inhibition of NADPH oxidase. Free Radic Biol Med.
53:437–446. 2012.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Armando I, Wang X, Villar VA, Jones JE,
Asico LD, Escano C and Jose PA: Reactive oxygen species-dependent
hypertension in dopamine D2 receptor-deficient mice. Hypertension.
49:672–678. 2007.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Zhang Y, Jiang X, Qin C, Cuevas S, Jose PA
and Armando I: Dopamine D2 receptors' effects on renal inflammation
are mediated by regulation of PP2A function. Am J Physiol Renal
Physiol. 310:F128–F134. 2016.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Marasa BS, Xiao L, Rao JN, Zou T, Liu L,
Wang J, Bellavance E, Turner DJ and Wang JY: Induced TRPC1
expression increases protein phosphatase 2A sensitizing intestinal
epithelial cells to apoptosis through inhibition of NF-kappaB
activation. Am J Physiol Cell Physiol. 294:C1277–C1287.
2008.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Li S, Wang L, Berman MA, Zhang Y and Dorf
ME: RNAi screen in mouse astrocytes identifies phosphatases that
regulate NF-kappaB signaling. Mol Cell. 24:497–509. 2006.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Kim SI, Kwak JH, Wang L and Choi ME:
Protein phosphatase 2A is a negative regulator of transforming
growth factor-beta1-induced TAK1 activation in mesangial cells. J
Biol Chem. 283:10753–10763. 2008.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Jung KJ, Lee EK, Kim SJ, Song CW, Maruyama
N, Ishigami A, Kim ND, Im DS, Yu BP and Chung HY: Anti-inflammatory
activity of SMP30 modulates NF-κB through protein tyrosine
kinase/phosphatase balance. J Mol Med (Berl). 93:343–356.
2015.PubMed/NCBI View Article : Google Scholar
|