Introduction
Acute lung injury (ALI) is caused by epithelial and
capillary endothelial cell damage that is induced by various direct
and indirect injurious factors, such as increased vascular
permeability, overproduction of cytokines, leukocyte recruitment
and dysfunction of surfactant, resulting in diffuse lung
interstitial and alveolar edema and acute hypoxic respiratory
insufficiency (1). ALI is a
disease that can threaten an individual's life. In severe cases, it
can lead to acute respiratory distress syndrome (ARDS) and
respiratory failure, which has a mortality rate of >30%
(2,3). The primary features of ALI and ARDS
are rapid onset of respiratory failure, severe hypoxemia and
reduced static respiratory system compliance; however, ALI and ARDS
are primarily caused by uncontrolled acute inflammation (4,5).
Previous studies have also demonstrated that the inhibition of
inflammation and oxidative stress can prevent ALI (6,7).
Lipopolysaccharide (LPS) is an endotoxin and a
constituent of the membrane of gram-negative bacteria (8). When gram-negative bacteria infect the
lungs, LPS is the primary pathogenic factor that causes ALI
(9). LPS can activate pattern
recognition receptors, thereby activating downstream NF-κB and MAPK
signaling molecules, which in turn promote the synthesis and
release of inflammatory factors, leading to excessive oxidative
stress (8,9). When endothelial and epithelial cells
are damaged, the endothelial barrier is destroyed, which increases
capillary permeability and promotes alveolar edema (10). Thus far, drug treatments for ALI
have not produced favorable results. Therefore, novel methods to
treat ALI using different approaches are required.
At present, an increasing body of evidence has
revealed that microRNAs (miRNAs/miRs) serve an important role in
various diseases (11), including
cancer (12), atherosclerosis
(13) and cardiovascular diseases
(14). miRNAs are a general term
for a class of small-molecule non-coding RNAs that are 20-22
nucleotides in length, as opposed to mRNA-transcribed proteins.
miRNAs do not encode proteins, but instead inhibit target gene
expression (15-17).
miR-122-5p is located at chr18q21.31, and it has been demonstrated
to participate in the development of several diseases, including
cancer (18), skeletal muscle
myogenesis (19), drug-induced
liver injury (20) and
radiation-induced rectal injury (21). In addition, Lu et al
(22) revealed that miR-122-5p was
involved in ALI development; miR-122-5p expression in ALI was
notably upregulated, and it was demonstrated to regulate pulmonary
microvascular endothelial cells by affecting the dual specificity
phosphatase 4 (DUSP4)/ERK signaling pathway to aggravate ALI
(22). However, its specific
mechanism of action is yet to be fully elucidated.
IL-1 receptor antagonist (IL1RN) is a member of the
IL-1 family, which is an immune and pro-inflammatory cytokine that
competitively binds to IL1R1 and prevents it from binding to the
co-receptor IL-1 receptor accessory protein (23). Therefore, IL1RN suppresses IL-1
activity, which regulates several immune and inflammatory responses
associated with IL-1(24). It has
been reported that IL1RN serves a protective role in numerous types
of lung injuries (25,26). Preliminary bioinformatics analysis
for the present study revealed that IL1RN is a target gene of
miR-122-5p. Therefore, it was hypothesized that miR-122-5p may be
involved in ALI through the modulation of IL1RN to regulate
alveolar epithelial cell injury.
Materials and methods
Cell culture and LPS exposure
The A549 human pulmonary epithelial cell line was
acquired from the American Type Culture Collection. Cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% penicillin-streptomycin at 37˚C in a humidified incubator
with 5% CO2. A549 was induced using 0.1, 1, 10 and 100
µg LPS (Sigma-Aldrich; Merck KGaA) for 12 h, or induced using 10 µg
LPS (Sigma) for 4, 8, 12 and 24 h. An ALI cell model was
established through treatment with 10 µg LPS (Sigma-Aldrich; Merck
KGaA) for 12 h (27).
Cell transfection
A549 cells were transfected with 50 nM miR-122-5p
mimic (5'-UGGAGUGUGACAAUGGUGUUUG-3') 50 nM mimic control
(5'-UUCUCCGAACGUGUCACGUTT-3'), 100 nM miR-122-5p inhibitor
(miR-122-5p antagomir; 5'-CAAACACCAUUGUCACACUCCA-3') and 100 nM
inhibitor control (the negative control of miR-122-5p antagomir;
5'-CAGUACUUUUGUGUAGUACAAA-3'), 0.2 µM scrambled control small
interfering (si)RNA (cat no. sc-36869; Santa Cruz Biotechnology,
Inc.), 0.2 µM IL1RN-siRNA (cat no. sc-39617; Santa Cruz
Biotechnology, Inc.), 100 nM miR-122-5p inhibitor + 0.2 µM
control-siRNA, or 100 nM miR-122-5p inhibitor + 0.2 µM IL1RN-siRNA
using Lipofectamine® 2000 (Thermo Fisher Scientific,
Inc.) at 37˚C for 24 h, according to the manufacturer's
instructions. After 24 h, RT-qPCR analysis was conducted to measure
the efficiency of transfection.
Bioinformatics
TargetScan version 7.2 (http://www.targetscan.org/vert_72/) was used to
predict the potential targets of miR-122-5p.
Dual luciferase reporter assay
TargetScan was used to predict the potential targets
of miR-122-5p. The results revealed that IL1RN was identified as a
potential target of miR-122-5p. Therefore, the wild-type (IL1RN-WT;
5'-UCCAAGCUCCAUCUCCACUCCAG-3') and mutant (IL1RN-MUT;
3'UCCAAGCUCCAUCUCGUGAGGUG-5') 3'untranslated regions (UTRs) of
IL1RN, containing miR-122-5p-binding elements, were generated by
reverse transcription (RT). RT was conducted using a HiScript 1st
Strand cDNA Synthesis kit (Vazyme Biotech Co., Ltd.) for 5 min at
25˚C followed by 60 min at 42˚C from total RNA preparations
extracted from A549 cells using TRIzol® reagent (Thermo
Fisher Scientific, Inc.). Samples were then cloned into BamHI and
AscI sites of the pmiRGLO vector (Promega Corporation). The
recombinant plasmids were acquired by using an EndoFree Plasmid
Maxi kit (Vazyme Biotech Co., Ltd.). Subsequently, 293T cells
(American Type Culture Collection) were seeded (5x104
cells/well) in 24-well plates and co-transfected with miR-122-5p
mimics (5'-UGGAGUGUGACAAUGGUGUUUG-3'; Guangzhou RiboBio Co., Ltd.)
or mimic control (5'-UUCUCCGAACGUGUCACGUTT-3'; Guangzhou RiboBio
Co., Ltd.) and the indicated luciferase reporter constructs using
Lipofectamine 2000® (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions at 37˚C for 48 h.
After transfection for 48 h, luciferase activity was assessed using
Dual Luciferase Reporter assay system (Promega Corporation) and
normalized to Renilla luciferase activity.
RT-quantitative (q)PCR assay
Total RNA was extracted from A549 cells using
TRIzol® reagent (Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. RNA concentration was
detected using a NanoDrop™ 2000 spectrophotometer (Thermo Fisher
Scientific, Inc.). Total RNA was reverse transcribed into cDNA
using a HiScript 1st Strand cDNA Synthesis kit (Vazyme Biotech Co.,
Ltd.) according to the manufacturer's protocol. Subsequently, qPCR
was performed using a SYBR® Green PCR kit (Vazyme
Biotech Co., Ltd.) according to the manufacturer's protocol. The
following thermocycling conditions were used for qPCR: Initial
denaturation at 95˚C for 5 min; followed by 38 cycles of 15 sec at
95˚C, 1 min at 60˚C and 30 sec at 72˚C; and a final extension for
10 min at 72˚C. GAPDH (for IL1RN mRNA) or U6 (for miR-122-5p) were
used as the endogenous controls. Gene expression was calculated
using the 2-ΔΔCq method (28). The primer sequences for qPCR were
as follows: GAPDH Forward, 5'-CTTTGGTATCGTGGAAGGACTC-3', and
reverse, 5'-GTAGAGGCAGGGATGATGTTCT-3'; U6 forward,
5'-GCTTCGGCAGCACATATACTAAAAT-3', and reverse,
5'-CGCTTCACGAATTTGCGTGTCAT-3'; miR-122-5p forward
5'-GTGACAATGGTGGAATGTGG-3', and reverse,
3'-CAGAACCGTAGCAAACGAAA-5'; IL1RN forward
5'-AACAGAAAGCAGGACAAGCG-3', and reverse,
5'-CCTTCGTCAGGCATATTGGT-3'.
Western blotting
Total protein was obtained from A549 cells using
RIPA lysis buffer (Beijing Solarbio Science & Technology Co.,
Ltd.). A BCA assay kit (Thermo Fisher Scientific, Inc.) was used to
quantify the total protein concentration. Equal amounts of proteins
(40 µg per lane) were isolated using 12% SDS-PAGE, and then
transferred to PVDF membranes. The membranes were blocked with 5%
non-fat milk to prevent non-specific binding at room temperature
for 1.5 h, and then incubated with primary antibodies against
anti-IL1RN (cat. no. ab124962; 1:1,000; Abcam), cleaved-caspase 3
(cat. no. ab32042; 1:1,000; Abcam), caspase 3 (cat. no. ab32351;
1:1,000; Abcam), and GAPDH (cat. no. ab9485; 1:2,500; Abcam) at 4˚C
overnight. The following day, membranes were incubated with goat
anti-rabbit IgG H&L (HRP)-conjugated secondary antibody
(1:5,000; cat. no. ab7090; Abcam) at room temperature for 2 h.
Protein signals were visualized using an ECL reagent (Cytiva).
ImageJ v.2.0 software (National Institutes of Health) was used to
quantify band intensity.
ELISA
ELISA was performed to examine the levels of TNF-α
(cat. no. PT518), IL-1β (cat. no. PI305) and IL-6 (cat. no. PI330)
in the supernatant of transfected A549 cells. Specific ELISA kits
(Beyotime Institute of Biotechnology) were used to detect the
expression of associated markers according to the manufacturer's
protocol.
Lactate dehydrogenase (LDH) release
assay
LDH release was determined using an LDH Release
Assay kit (cat. no. C0016; Beyotime Institute of Biotechnology)
according to the manufacturer's protocol. After treatment with LPS,
150 µl LDH release reagent was added to DMEM and cells were
incubated at 37˚C for a total of 2 h. Subsequently, samples were
centrifuged at 4˚C and 500 x g for 5 min, and 120 µl supernatant
from each well was used to measure the LDH release according to the
requirements of the kit. Absorbance was measured at a wavelength of
490 nm using a microplate reader.
Flow cytometry
Transfected cells were processed using an
Annexin-V/PI Apoptosis Detection kit (BD Biosciences). After cell
transfection, cells were induced with LPS for 12 h, collected
through centrifugation at 1,000 x g at 4˚C for 5 min and then
resuspended in 100 µl FITC-binding buffer. Subsequently, ~5 µl
ready-to-use Annexin V-FITC and 5 µl PI were added to the buffer
and incubated in the dark for 30 min at room temperature. Annexin
V-FITC and PI fluorescence were assessed using a BD FACSCalibur
flow cytometer (BD Biosciences) and the results were analyzed using
Kaluza Analysis software (v2.1.1.20653; Beckman Coulter, Inc.).
MTT assay
Transfected cells were treated with 10 µg LPS for 12
h and then plated in a 96-well plate and incubated at 37˚C for 24
h. Subsequently, 20 µl MTT (5 mg/ml; Sigma-Aldrich; Merck KGaA) was
added to each well, and the wells were further cultured at 37˚C for
4 h. The absorbance was measured at 570 nm using a multifunctional
plate reader (BioTek Instruments, Inc.).
Statistical analysis
Results were analyzed using GraphPad Prism v6.0
(GraphPad Software, Inc.). Differences between two groups were
compared using an unpaired Student's t-test. The differences among
multiple groups were calculated using one-way ANOVA followed by
Tukey's post hoc test. Data are presented as the mean ± SD of three
independent experiments. P<0.05 was considered to indicate a
statistically significant difference.
Results
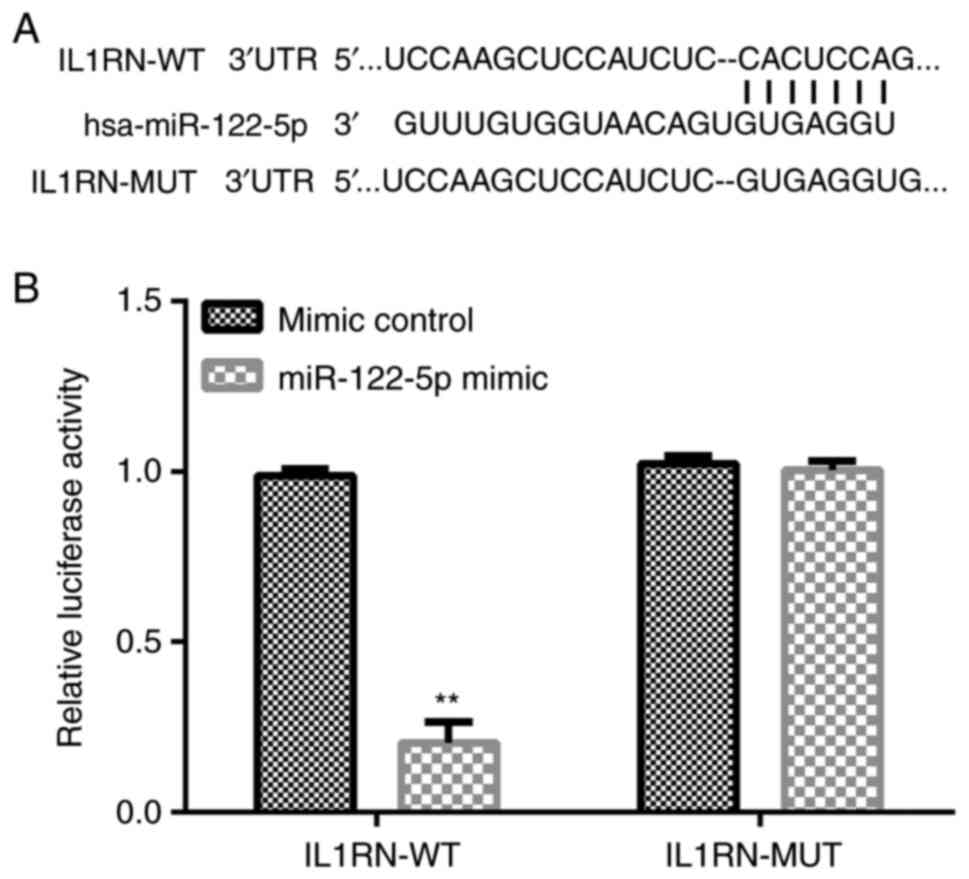
IL1RN is the direct target gene of
miR-122-5p
The target gene downstream of miR-122-5p was
searched using the bioinformatics tool, TargetScan. TargetScan
results revealed that there were thousands of target genes of
miR-122-5p including IL1RN (Fig.
1A). Based on software prediction, miR-122-5p was partially
complementary with the IL1RN 3'-UTR. IL1RN is an antagonist of IL1,
which is a pro-inflammatory cytokine (24). It has been reported that IL1RN
serves a protective role in numerous types of lung injury (25,26).
Thus, the present study hypothesized that miR-122-5p may be
involved in ALI via the modulation of IL1RN expression. Therefore,
IL1RN was selected for further study. Subsequently, 293T cells were
co-transfected with IL1RN-WT or IL1RN-MUT and miR-122-5p mimic or
mimic control for 48 h, after which a dual-luciferase reporter
assay was performed to detect luciferase activity. Dual-luciferase
reporter assays indicated that miR-122-5p mimics could
significantly inhibit the luciferase activity of IL1RN-WT. However,
miR-122-5p mimic did not inhibit the luciferase activity of
IL1RN-MUT (Fig. 1B). Overall,
IL1RN was revealed to be a target gene of miR-122-5p.
Expression of IL1RN and miR-122-5p in
the LPS-induced ALI cell model
To detect the expression of IL1RN and miR-122-5p in
the in vitro model of ALI, A549 cells were treated with LPS.
A549 was induced using 0.1, 1, 10 and 100 µg LPS for 12 h. RT-qPCR
and western blotting were used to detect miR-122-5p and IL1RN
expression. The results revealed that LPS dose-dependently
increased miR-122-5p expression in A549 cells (Fig. 2A), and reduced IL1RN expression at
both the protein and mRNA levels (Fig.
2B and C); these results were
significant at a dosage of ≥1 µg. The effects of treatment with LPS
for different time periods were subsequently assessed. A549 cells
were induced using 10 µg LPS for 4, 8, 12 and 24 h. The results
indicated that LPS significantly increased miR-122-5p expression in
A549 cells (Fig. 2D) and
significantly reduced the protein and mRNA expression levels of
IL1RN in a time-dependent manner (Fig.
2E and F). For subsequent
experiments, A549 cells were induced with 10 µg LPS for 12 h to
establish the ALI cell model (27). The successful establishment of the
LPS-induced ALI cell model was confirmed through the reduction of
A549 cell viability, enhanced LDH activity, increased apoptosis and
enhanced inflammatory cytokine levels.
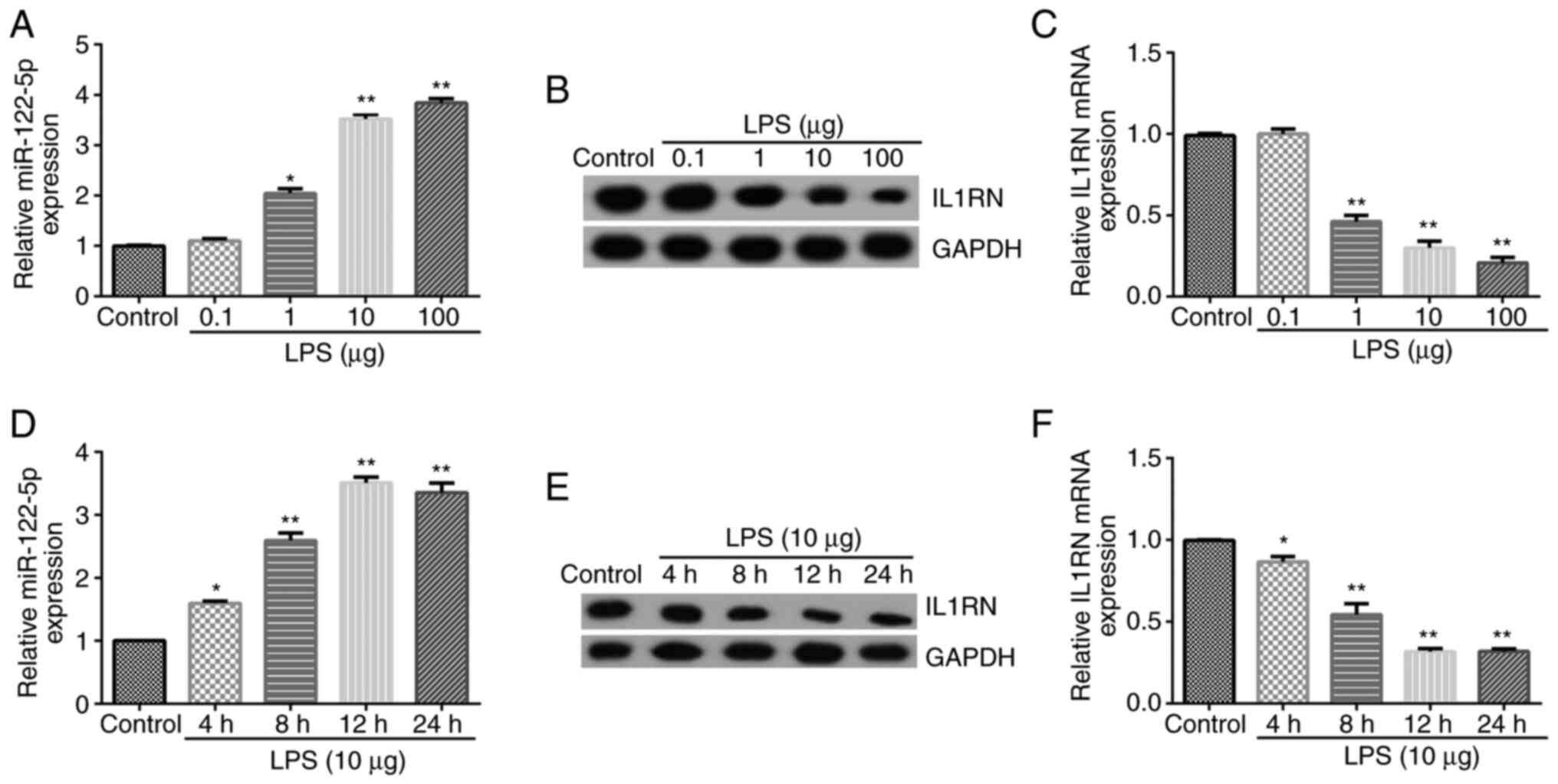 | Figure 2miR-122-5p expression is upregulated
and IL1RN is downregulated in the LPS-treated acute lung injury
cell model. Different concentrations of LPS (0.1, 1, 10 or 100 µg)
were used to induce A549 cells for 12 h. (A) RT-qPCR analysis of
miR-122-5p expression. (B) Western blotting of IL1RN expression.
(C) RT-qPCR analysis of IL1RN expression. (D) RT-qPCR analysis of
miR-122-5p expression when A549 cells were treated with 10 µg LPS
for 4, 8, 12 or 24 h. (E) Protein and (F) mRNA expression analysis
of IL1RN expression in A549 cells treated with 10 µg LPS for 4, 8,
12 or 24 h. *P<0.05 and **P<0.01 vs.
control. miR, microRNA; IL1RN, IL-1 receptor antagonist; LPS,
lipopolysaccharide; RT-qPCR, reverse transcription-quantitative
PCR. |
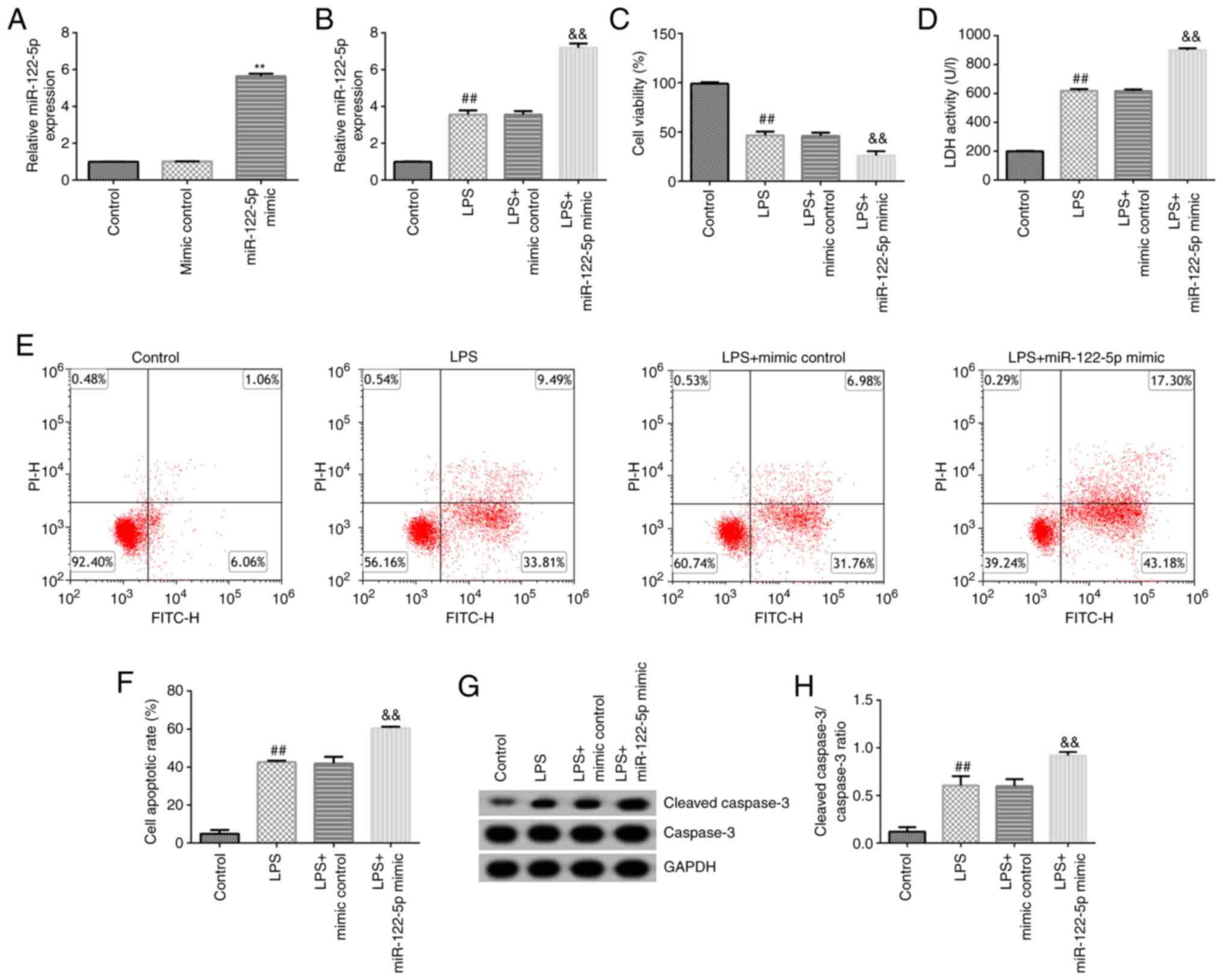
Effects of miR-122-5p mimics on the
LPS-induced ALI cell model
To investigate the effects of miR-122-5p mimics on
LPS-induced ALI cells. A549 cells were transfected with mimic
control or miR-122-5p mimic for 24 h. RT-qPCR was performed to
confirm the transfection efficiency. Compared with the mimic
control group, miR-122-5p mimic significantly increased miR-122-5p
expression in A549 cells (Fig.
3A). After 24 h, transfected cells were induced with 10 µg LPS
for 12 h, and subsequent analysis was performed. RT-qPCR analysis
revealed that miR-122-5p expression levels were significantly
increased in LPS-induced cells compared with their respective
control (Fig. 3B). By contrast,
MTT analysis indicated that LPS significantly decreased cell
viability in the LPS and LPS + miR-122-5p mimic groups compared
with the control and LPS + mimic control, respectively (Fig. 3C). Additionally, LPS significantly
promoted LDH release in these groups (Fig. 3D). Flow cytometry analysis
indicated that LPS significantly increased the rate of apoptosis
(Fig. 3E and F); similarly, western blotting revealed
that cleaved-caspase 3 expression (Fig. 3G) and the cleaved-caspase 3/caspase
3 ratio were significantly increased (Fig. 3H). All these changes were
reinforced by miR-122-5p mimic.
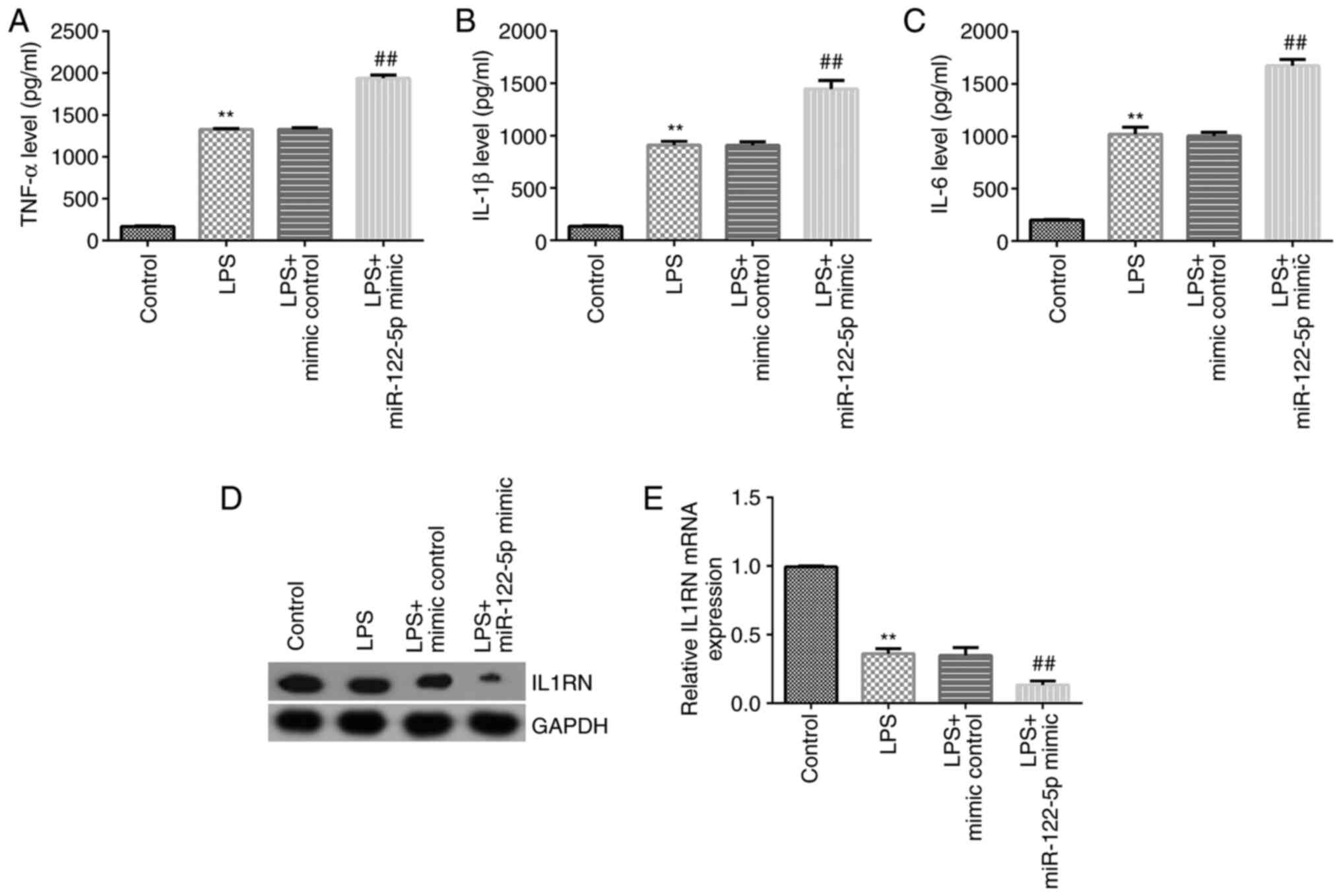
Effect of miR-122-5p mimic on
expression of inflammatory cytokines
ELISA was performed to detect the expression of
inflammatory cytokines. The results revealed that TNF-α, IL-1β and
IL-6 levels were all significantly increased in the cell
supernatant of the LPS-treated group compared with the control. The
miR-122-5p mimic further significantly increased the levels of
TNF-α (Fig. 4A), IL-1β (Fig. 4B) and IL-6 (Fig. 4C) compared with the LPS + mimic
control. Western blotting and RT-qPCR analysis were subsequently
used to detect IL1RN expression. The results revealed that LPS
significantly decreased IL1RN expression compared with the control
group; while miR-122-5p mimics further significantly decreased
IL1RN expression in LPS-induced A549 cells compared with the LPS +
mimic control group (Fig. 4D and
E).
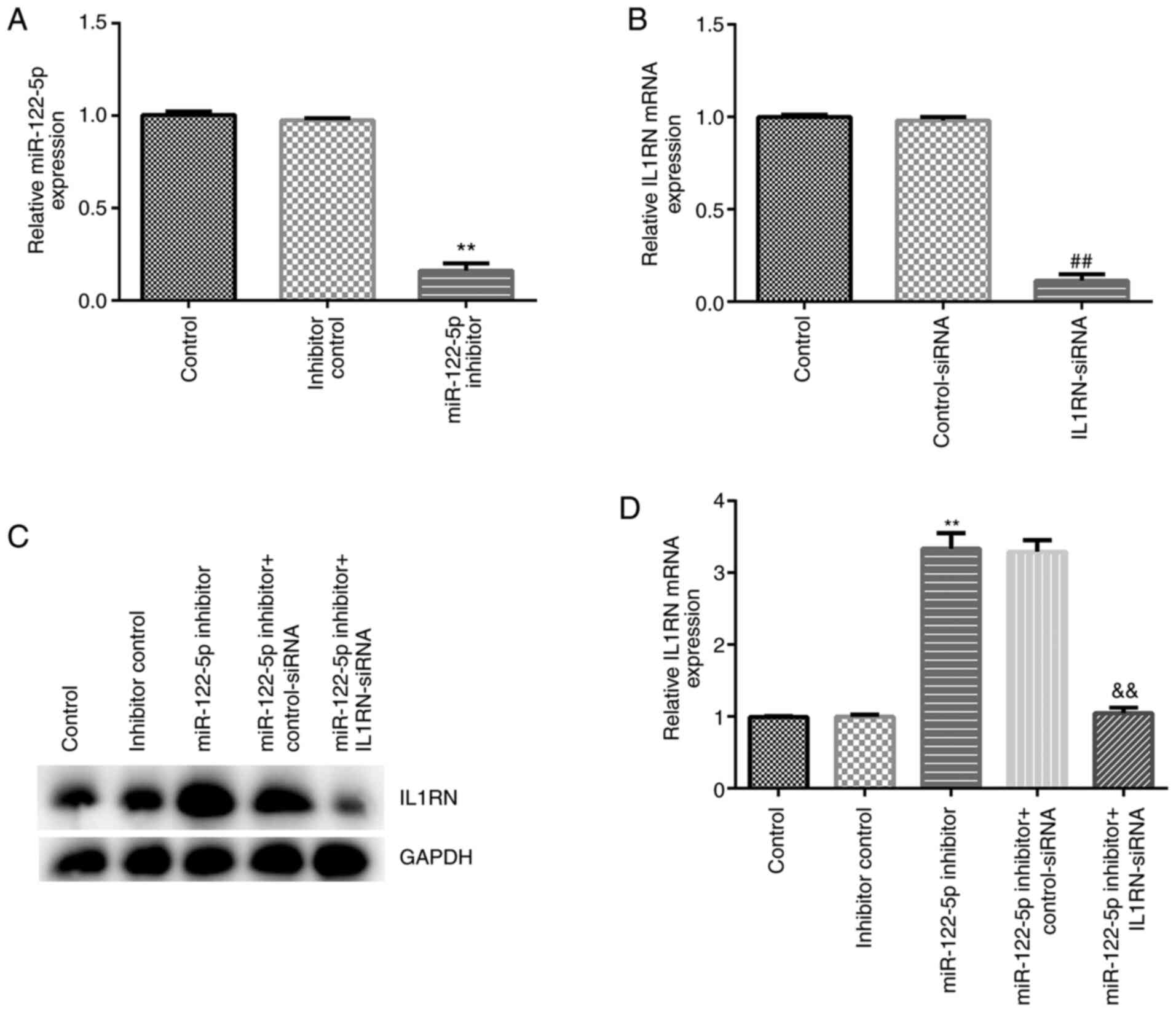
miR-122-5p negatively regulates IL1RN
expression in A549 cells
The effect of miR-122-5p inhibition on the
LPS-induced ALI cell model was assessed. A549 cells were
transfected with inhibitor control, miR-122-5p inhibitor,
control-siRNA, IL1RN-siRNA, miR-122-5p inhibitor + control-siRNA or
miR-122-5p inhibitor + IL1RN-siRNA for 24 h, after which RT-qPCR
was performed to evaluate transfection efficiency. Compared with
the inhibitor control group, the miR-122-5p inhibitor significantly
reduced miR-122-5p expression in A549 cells (Fig. 5A). Compared with the control-siRNA
group, IL1RN-siRNA significantly decreased IL1RN mRNA expression in
A549 cells (Fig. 5B). In addition,
the miR-122-5p inhibitor significantly increased IL1RN expression
compared with the inhibitor control, and this increase was
significantly reversed by IL1RN-siRNA (Fig. 5C and D).
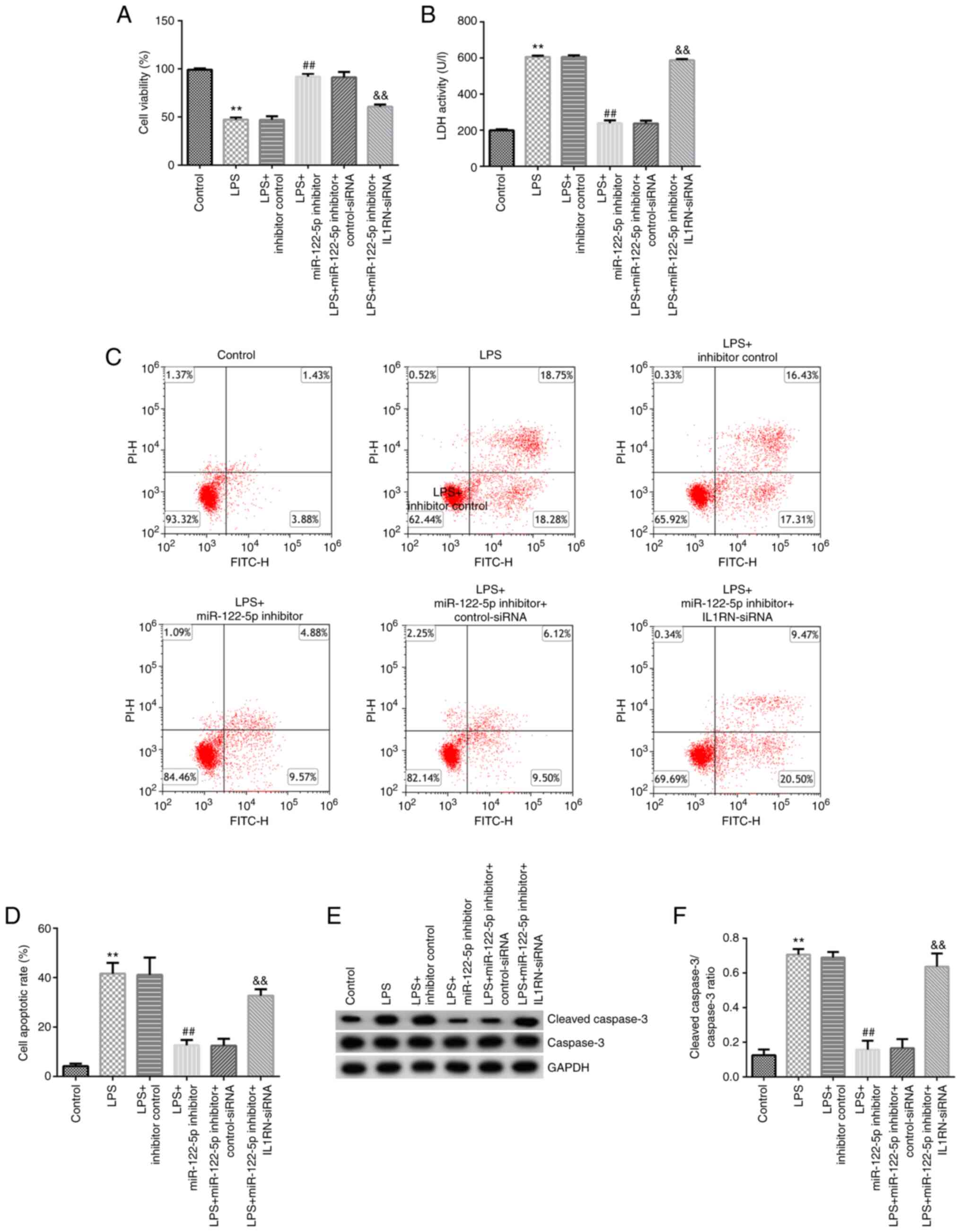
Effect of the miR-122-5p inhibitor on
the LPS-induced ALI cell model
A549 cells were transfected with inhibitor control,
miR-122-5p inhibitor, miR-122-5p inhibitor + control-siRNA or
miR-122-5p inhibitor + IL1RN-siRNA for 24 h, and then subsequently
induced with 10 µg LPS for 12 h. MTT assays revealed that LPS
significantly decreased cell viability compared with the control
(Fig. 6A). The LDH release assay
indicated that LDH release was significantly increased in the LPS
group compared with the control (Fig.
6B). The results from the flow cytometry assays demonstrated
that LPS significantly promoted apoptosis compared with the control
(Fig. 6C and D). Western blotting revealed that LPS
significantly increased cleaved-caspase 3 expression (Fig. 6E) and the cleaved-caspase 3/caspase
3 ratio compared with the control (Fig. 6F). When compared with the LPS +
inhibitor control group, the cell viability of the LPS + miR-122-5p
inhibitor group was significantly increased (Fig. 6A), LDH release was significantly
reduced (Fig. 6B), apoptosis was
significantly decreased (Fig. 6C
and D), the cleaved-caspase 3
protein expression was significantly decreased (Fig. 6E) and the cleaved-caspase 3/caspase
3 ratio was significantly decreased in the LPS + miR-122-5p
inhibitor group (Fig. 6F). All
these changes were significantly reversed by IL1RN-siRNA.
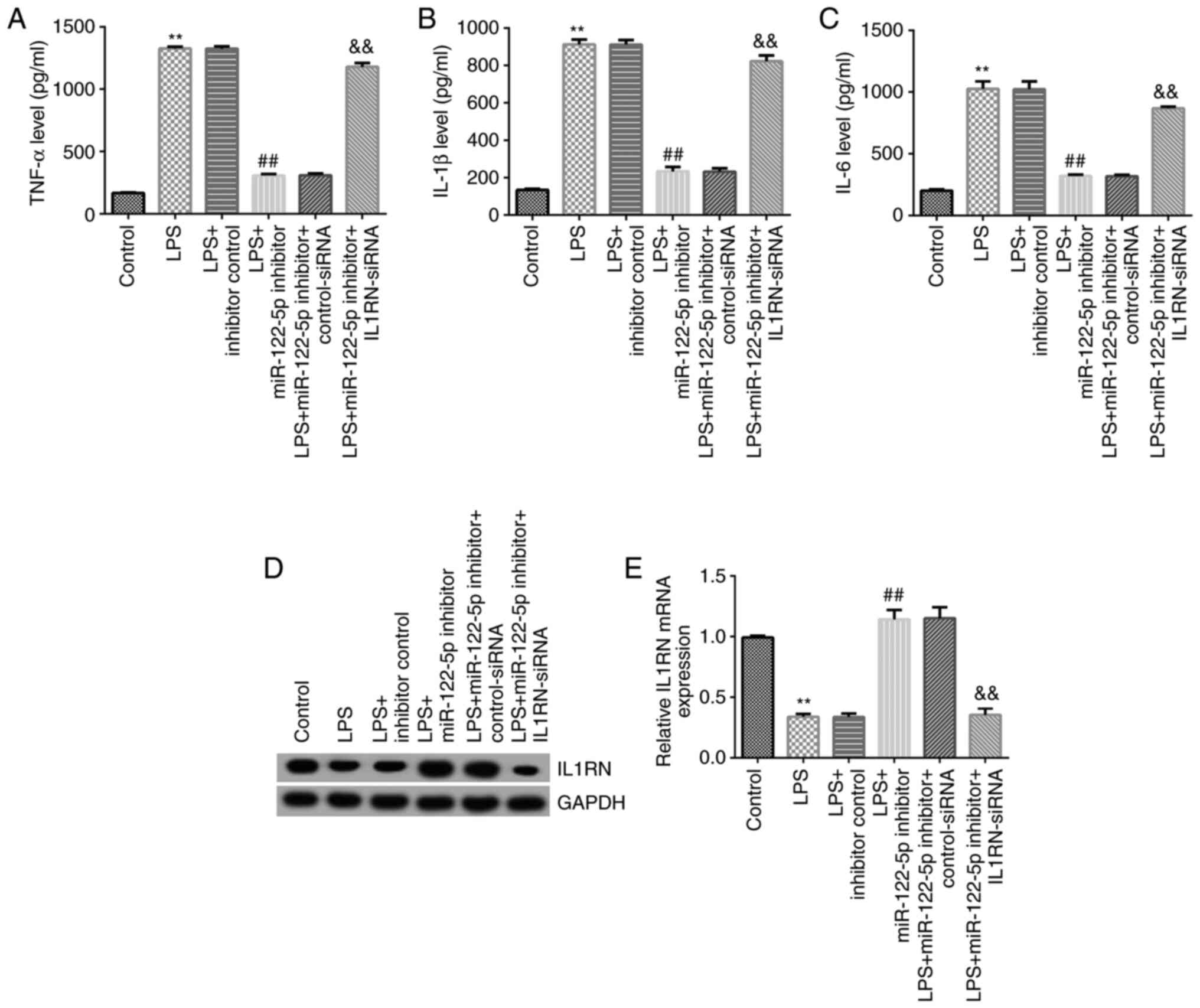
Effects of miR-122-5p inhibitor on the
expression of inflammatory cytokines
The effects of the miR-122-5p inhibitor on the
expression of inflammatory cytokines were assessed. Compared with
the control group, the secretion profiles of TNF-α, IL-1β and IL-6
in the cell supernatant of the LPS group were significantly
increased (Fig. 7A-C), and the
expression of IL1RN was significantly reduced (Fig. 7D and E). Compared with the LPS + inhibitor
control group, the secretion profiles of TNF-α, IL-1β and IL-6 were
significantly reduced in the LPS + miR-122-5p inhibitor group
(Fig. 7A-C), and IL1RN expression
was significantly increased (Fig.
7D and E). All of these
changes were significantly reversed by IL1RN-siRNA.
Discussion
ALI can lead to the development of ARDS, which
severely impacts an individual's life (2,3). The
primary feature of ALI is abnormal respiratory function (4,5).
Although treatment methods for ARDS are constantly improving, the
mortality rate of patients with ARDS remains high at ~30% (29). Recently, miRNAs have been
demonstrated to serve a notable role in the development of several
types of diseases and have attracted significant attention. Yang
and Zhao (30) revealed that
miR-490-3p upregulation suppressed LPS-induced ALI via the IL-1
receptor-associated kinase 1/TNF receptor-associated factor 6
pathway. Furthermore, Li et al (31) demonstrated that miR-150 expression
was downregulated in an LPS-induced ALI cell model, and that
miR-150-overexpression inhibited LPS-induced ALI. Suo et al
(32) demonstrated that miR-1246
inhibited ALI-induced inflammation via the NF-κB and Wnt/β-catenin
signaling pathways. Results of a previous study indicated that
miR-122-5p participates in the development of ALI via the DUSP4/ERK
signaling axis (22). In the
present study, another potential regulatory mechanism involving
miR-122-5p in the development of ALI was assessed.
Several studies have demonstrated that miR-122-5p
participates in the development of various diseases (19,20,33).
Ma et al (33) revealed
that miR-122-5p inhibited osteosarcoma cell proliferation. Ding
et al (19) demonstrated
that miR-122-5p promoted skeletal muscle myogenesis via
transforming growth factor β receptor 2. Yang et al
(20) demonstrated that miR-122-5p
downregulation can protect against acetaminophen-mediated liver
damage by upregulating NDRG family member 3 expression. The results
of the present study indicated that the target gene downstream of
miR-122-5p was IL1RN, and that there was a direct interaction
between IL1RN mRNA and miR-122-5p. However, to validate IL1RN as
the direct target gene of miR-122-5p, further investigations into
the effects of an miR-122-5p inhibitor on the luciferase activity
of IL1RN-WT are required. This was a limitation of the current
study.
The IL-1 family consists of three members, IL-1α,
IL-1β and IL1RN (34). IL1RN is an
anti-inflammatory molecule that exhibits homology with IL-1α and
IL-1β. IL1RN participates in the development of several types of
cancer, such as prostate cancer (34), non-cardia gastric carcinoma
(35), glioma (36) and bladder cancer (37). miRNAs are endogenous, non-coding,
single-stranded small RNAs, which negatively regulate gene
expression by interacting with the 3'-UTRs of target mRNAs at the
posttranscriptional or translational level (38). The present study revealed that
miR-122-5p negatively regulated IL1RN expression in A549 cells.
ALI is characterized by an inflammatory process that
is associated with the upregulation of chemokines and inflammatory
cytokines (39). Niu et al
(40) indicated that when the
lungs are infected, LPS binds to molecules on the surface of
endothelial cells, thereby promoting the expression of various
inflammatory cytokines, including IL-1β, IL-6, IL-8 and TNF-α.
Overproduction of pro-inflammatory cytokines leads to severe lung
damage, and the abnormal apoptosis of pulmonary cells is a
pathophysiological feature of ALI (41,42).
In the present study, miR-122-5p mimics promoted the expression of
inflammatory cytokines. However, the miR-122-5p inhibitor
suppressed TNF-α, IL-1β and IL-6 levels in LPS-treated A549 cells.
Furthermore, miR-122-5p mimic suppressed cell viability and
promoted apoptosis in LPS-treated A549 cells, with the miR-122-5p
inhibitor demonstrating the opposite effect.
Caspase 3, a member of the caspase family (key
effector molecules of apoptosis), is considered to serve an
important role in the cascade of apoptosis and is regarded as an
executor and terminator of multiple apoptotic pathways (43). After cleaved activation
(cleaved-caspase 3), caspase 3 exerts a pro-apoptotic effect, and
the increase of the cleaved-caspase 3/caspase 3 ratio reflects the
activation of caspase 3 (43,44).
The present study also analyzed caspase 3, and the data indicated
that miR-122-5p mimic increased cleaved-caspase 3 protein
expression and the cleaved-caspase 3/caspase 3 ratio in LPS-treated
A549 cells, while the miR-122-5p inhibitor had the opposite
effect.
In conclusion, downregulation of miR-122-5p reduced
LPS-induced ALI by targeting IL1RN. However, the present study was
only a preliminary in vitro study of the effect of
miR-122-5p on an LPS-induced ALI cell model. In order to make the
role of miR-122-5p in ALI more convincing, more in-depth research
is required. For example, the role of miR-122-5p and IL1RN alone in
A549 cells in the absence of LPS should be clarified, and the
effect of miR-122-5p/IL1RN in an animal model of ALI should be
explored. Moreover, the expression of miR-122-5p/IL1RN in patients
with ALI and its association with the clinicopathological
parameters of patients with ALI should be further studied.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JL and XZ contributed to the conception and design
of the study, in addition to data acquisition, analysis and
interpretation. JZ and XZ also drafted and critically revised the
manuscript. WW contributed to data collection, statistical analysis
and manuscript preparation. JL and XZ confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen H, Bai C and Wang X: The value of the
lipopolysaccharide-induced acute lung injury model in respiratory
medicine. Expert Rev Respir Med. 4:773–783. 2010.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Avecillas JF, Freire AX and Arroliga AC:
Clinical epidemiology of acute lung injury and acute respiratory
distress syndrome: Incidence, diagnosis, and outcomes. Clin Chest
Med. 27:549–557. 2006.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bellani G, Laffey JG, Pham T, Fan E,
Brochard L, Esteban A, Gattinoni L, van Haren F, Larsson A, McAuley
DF, et al: Epidemiology, patterns of care, and mortality for
patients with acute respiratory distress syndrome in intensive care
units in 50 countries. JAMA. 315:788–800. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Jongerius I, Porcelijn L, van Beek AE,
Semple JW, van der Schoot CE, Vlaar APJ and Kapur R: The role of
complement in transfusion-related acute lung injury. Transfus Med
Rev. 33:236–242. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Murray DD, Itenov TS, Sivapalan P, Eklöf
JV, Holm FS, Schuetz P and Jensen JU: Biomarkers of acute lung
injury the individualized approach: For phenotyping, risk
stratification and treatment surveillance. J Clin Med.
8(1163)2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Liu H, Yu X, Yu S and Kou J: Molecular
mechanisms in lipopolysaccharide-induced pulmonary endothelial
barrier dysfunction. Int Immunopharmacol. 29:937–946.
2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lei J, Wei Y, Song P, Li Y, Zhang T, Feng
Q and Xu G: Cordycepin inhibits LPS-induced acute lung injury by
inhibiting inflammation and oxidative stress. Eur J Pharmacol.
818:110–114. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Nova Z, Skovierova H and Calkovska A:
Alveolar-capillary membrane-related pulmonary cells as a target in
endotoxin-induced acute lung injury. Int J Mol Sci.
20(831)2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Reiss LK, Uhlig U and Uhlig S: Models and
mechanisms of acute lung injury caused by direct insults. Eur J
Cell Biol. 91:590–601. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Rebetz J, Semple JW and Kapur R: The
pathogenic involvement of neutrophils in acute respiratory distress
syndrome and transfusion-related acute lung injury. Transfus Med
Hemother. 45:290–298. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang Y, Zhang M, Zhong M, Suo Q and Lv K:
Expression profiles of miRNAs in polarized macrophages. Int J Mol
Med. 31:797–802. 2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chen X, Slack FJ and Zhao H: Joint
analysis of expression profiles from multiple cancers improves the
identification of microRNA-gene interactions. Bioinformatics.
29:2137–2145. 2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wei Y, Nazari-Jahantigh M, Chan L, Zhu M,
Heyll K, Corbalán-Campos J, Hartmann P, Thiemann A, Weber C and
Schober A: The microRNA-342-5p fosters inflammatory macrophage
activation through an Akt1- and microRNA-155-dependent pathway
during atherosclerosis. Circulation. 127:1609–1619. 2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ono K, Kuwabara Y and Han J: MicroRNAs and
cardiovascular diseases. FEBS J. 278:1619–1633. 2011.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ro S, Park C, Young D, Sanders KM and Yan
W: Tissue-dependent paired expression of miRNAs. Nucleic Acids Res.
35:5944–5953. 2007.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mallory AC and Vaucheret H: MicroRNAs:
Something important between the genes. Curr Opin Plant Biol.
7:120–125. 2004.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Meng L, Chen Z, Jiang Z, Huang T, Hu J,
Luo P, Zhang H, Huang M, Huang L, Chen Y, et al: MiR-122-5p
suppresses the proliferation, migration, and invasion of gastric
cancer cells by targeting LYN. Acta Biochim Biophys Sin (Shanghai).
521:49–57. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ding Z, Lin J, Sun Y, Cong S, Liu S, Zhang
Y, Chen Q and Chen J: miR-122-5p negatively regulates the
transforming growth factor-β/Smad signaling pathway in skeletal
muscle myogenesis. Cell Biochem Funct. 382:231–238. 2020.PubMed/NCBI View
Article : Google Scholar
|
|
20
|
Yang Z, Wu W, Ou P, Wu M, Zeng F, Zhou B
and Wu S: MiR-122-5p knockdown protects against APAP-mediated liver
injury through up-regulating NDRG3. Mol Cell Biochem.
476:1257–1267. 2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Ge Y, Tu W, Li J, Chen X, Chen Y, Xu Y, Xu
Y, Wang Y and Liu Y: MiR-122-5p increases radiosensitivity and
aggravates radiation-induced rectal injury through CCAR1. Toxicol
Appl Pharmacol. 399(115054)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lu Z, Feng H, Shen X, He R, Meng H, Lin W
and Geng Q: MiR-122-5p protects against acute lung injury via
regulation of DUSP4/ERK signaling in pulmonary microvascular
endothelial cells. Life Sci. 256(117851)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Yazdi AS and Ghoreschi K: The
interleukin-1 family. Adv Exp Med Biol. 941:21–29. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lin SC, Lo YC and Wu H: Helical assembly
in the MyD88-IRAK4-IRAK2 complex in TLR/IL-1R signalling. Nature.
465:885–890. 2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Bui CB, Kolodziej M, Lamanna E, Elgass K,
Sehgal A, Rudloff I, Schwenke DO, Tsuchimochi H, Kroon MAGM, Cho
SX, et al: Interleukin-1 receptor antagonist protects newborn mice
against pulmonary hypertension. Front Immunol.
10(1480)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sun D, Zhao M, Ma D, Liao S and Di C:
Protective effect of interleukin-1 receptor antagonist on oleic
acid-induced lung injury. Chin Med J (Engl). 109:522–526.
1996.PubMed/NCBI
|
|
27
|
Zhou H, Wang X and Zhang B: Depression of
lncRNA NEAT1 antagonizes LPS-evoked acute injury and inflammatory
response in alveolar epithelial cells via HMGB1-RAGE signaling.
Mediators Inflamm. 2020(8019467)2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Gill SE, Yamashita CM and Veldhuizen RA:
Lung remodeling associated with recovery from acute lung injury.
Cell Tissue Res. 367:495–509. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Yang G and Zhao Y: MicroRNA-490-3p
inhibits inflammatory responses in LPS-induced acute lung injury of
neonatal rats by suppressing the IRAK1/TRAF6 pathway. Exp Ther Med.
21(152)2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li P, Yao Y, Ma Y and Chen Y: MiR-150
attenuates LPS-induced acute lung injury via targeting AKT3. Int
Immunopharmacol. 75(105794)2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Suo T, Chen GZ, Huang Y, Zhao KC, Wang T
and Hu K: miRNA-1246 suppresses acute lung injury-induced
inflammation and apoptosis via the NF-κB and Wnt/β-catenin signal
pathways. Biomed Pharmacother. 108:783–791. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ma W, Zhao X, Xue N, Gao Y and Xu Q: The
LINC01410/miR-122-5p/NDRG3 axis is involved in the proliferation
and migration of osteosarcoma cells. IUBMB Life. 73:705–717.
2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Fan YC, Lee KD and Tsai YC: Roles of
interleukin-1 receptor antagonist in prostate cancer progression.
Biomedicines. 8(602)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Rocha GA, Guerra JB, Rocha AM, Saraiva IE,
da Silva DA, de Oliveira CA and Queiroz DM: IL1RN polymorphic gene
and cagA-positive status independently increase the risk of
noncardia gastric carcinoma. Int J Cancer. 115:678–683.
2005.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Wang K, Liu H, Liu J, Wang X, Teng L,
Zhang J, Liu Y, Yao Y, Wang J, Qu Y, et al: IL1RN mediates the
suppressive effect of methionine deprivation on glioma
proliferation. Cancer Lett. 54:146–157. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Worst TS, Reiner V, Gabriel U, Weiß C,
Erben P, Martini T and Bolenz C: IL1RN and KRT13 expression in
bladder cancer: Association with pathologic characteristics and
smoking status. Adv Urol. 2014(184602)2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li P, Zhou Y, Goodwin AJ, Cook JA,
Halushka PV, Zhang XK, Wilson CL, Schnapp LM, Zingarelli B and Fan
H: Fli-1 governs pericyte dysfunction in a murine model of sepsis.
J Infect Dis. 218:1995–2005. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Niu X, Zang L and Li W, Xiao X, Yu J, Yao
Q, Zhao J, Ye Z, Hu Z and Li W: Anti-inflammatory effect of yam
glycoprotein on lipopolysaccharide-induced acute lung injury via
the NLRP3 and NF-κB/TLR4 signaling pathway. Int Immunopharmacol.
81(106024)2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Li S, Zhang Y, Guan Z, Li H, Ye M, Chen X,
Shen J, Zhou Y, Shi ZL, Zhou P and Peng K: SARS-CoV-2 triggers
inflammatory responses and cell death through caspase-8 activation.
Signal Transduct Target Ther. 5(235)2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Yang L, Zhang Z, Zhuo Y, Cui L, Li C, Li
D, Zhang S, Cui N, Wang X and Gao H: Resveratrol alleviates
sepsis-induced acute lung injury by suppressing inflammation and
apoptosis of alveolar macrophage cells. Am J Transl Res.
10:1961–1975. 2018.PubMed/NCBI
|
|
43
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104.
1999.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Fan Y and Bergmann A: The
cleaved-caspase-3 antibody is a marker of caspase-9-like DRONC
activity in Drosophila. Cell Death Differ. 17:534–539.
2010.PubMed/NCBI View Article : Google Scholar
|