Introduction
Spinal cord injury (SCI) is a devastating
neurological disorder, leading to the motor, sensory and autonomic
dysfunction below the injured segment, which not only seriously
impacts the quality of life of the patients and represents economic
burdens to the society (1). It is
well documented that the number of deaths, paraplegia and
quadriplegia caused by SCI is gradually increasing. Although
numerous patients with SCI have a response to surgery, cell
transplantation and gene therapy and drug therapy, highly disabling
disease is common, and relapse represents the major cause of
treatment failure (2,3). It has become increasingly clear that
the primary mechanical injury and secondary oxidative stress,
neuronal inflammation and apoptosis are involved in the
pathogenesis of SCI (4,5).
miRNAs are small single-stranded RNAs of 18-25
nucleotides in length that are transcribed from DNA (6). miRNA is partially or completely
complementary to the 3'UTR region of the target gene by
base-pairing interactions to inhibit mRNA translation or induce its
degradation, thereby functioning to control the expression of the
target gene (7). Emerging research
suggest that several miRNAs play pivotal roles in regulating cell
differentiation, proliferation and survival (8,9). A
variety of miRNAs are differentially expressed in the onset and
development of SCI, and are involved in the regulation of multiple
physiological processes such as the proliferation, apoptosis and
autophagy, as well as oxidative damage of neuronal cells (10,11).
There is emerging evidence that the excess production of reactive
oxygen species, such as H2O2, has been
implicated in the secondary damage after SCI (12). Further clarification of the key
miRNA molecules regulating H2O2-caused cell
injury exploits new ideas for the diagnosis and treatment of SCI.
It was recently reported that miR-218-5p was significantly
upregulated in oxygen-glucose deprivation/reoxygenation
(OGD/R)-treated PC12 cells, and downregulation of miR-218-5p
protected against OGD/R-induced cell injury through attenuating
oxidative stress, inflammation and apoptosis (13). A large quantity of evidence has
revealed abnormal expression of miR-218 in cancer (14), cardiovascular diseases (15) and neurological disorders (16). However, little was known about the
roles of miR-218 in SCI to the best of our knowledge.
The present study was designed to investigate the
effect of miR-218 on H2O2-induced apoptosis
and autophagy of rat neuronal PC12 cells, and to further examine
the underlying mechanism.
Materials and methods
Cell culture and treatment
Rat neuronal PC12 cell line, obtained from BioVector
NTCC, were cultured in DMEM medium containing 10% fetal bovine
serum and penicillin/streptomycin (Gibco; Thermo Fisher Scientific,
Inc.) at 37˚C in a humidified atmosphere with 5% CO2.
Culture medium was changed every other day. For the
H2O2 treatment, PC12 cells were seeded into
cell culture multi-well plates and incubation was continued for 24
h. To induce the cell injury model, these cells were then exposed
to fresh medium containing 200 µM H2O2 for 24
h as previously described (17,18).
The cells treated with the same medium without
H2O2 were used as controls.
Cell transfection
The mirVana® miRNA mimic rno-miR-218
(Assay ID: MC20167), mirVana™ miRNA Mimic Negative
Control, mirVana® miRNA inhibitor anti-rno-miR-218
(Assay ID: MH20167), mirVana™ miRNA Inhibitor Negative
Control, predesigned Silencer® Select siRNA spred2
(Assay ID: s156883) and Silencer® Select siRNA negative
control were all were purchased from Thermo Fisher Scientific,
Inc.. The PC12 cells were transfected with the vectors after
confluence reached 70% using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Cells were collected after 48 h
transfection for the following experiments. The sequences of the
transfection agents are shown in Table
I.
 | Table ISequences of transfection
vectors. |
Table I
Sequences of transfection
vectors.
| Vector | Sequence,
5'-3' |
|---|
| miR-218 mimics |
UUGUGCUUGAUCUAACCAUGU |
| miR-NC |
UCGCUUGGUGCAGGUCGGGAA |
|
miR-218-inhibitor |
ACATGGTTAGATCAAGCACAA |
| NC-inhibitor |
UUCUCCGAACGUGUCACGUTT |
| si-spred2 |
GGAAGAUGAUGAAGAGAUAGU |
| si-NC |
CAGUACUUUUGUGUAGUACAA |
Luciferase reporter assay
The coding sequence of spred2 containing the
predicted binding site of miR-218 was amplified, which was inserted
into a pmirGLO Dual-Luciferase miRNA Target Expression Vector
(Promega Corporation) to establish the vectors of spred2-wild-type
(spred2-wt). The spred2-mutated-type (spred2-mut) reporter vector
carrying the mutated binding site of miR-218 in the spred2 was
constructed as negative controls. Luciferase assays were performed
in PC12 cells after co-transfection with the wild/mutated types of
spred2 promoter reporters and miR-218 mimics or miR-NC using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). After 48 h of transfection, the luciferase
activities were tested using the Dual-Luciferase Reporter Assay
system (Promega Corporation).
Cell viability assay
Transfected PC12 cells were re-seeded into 96-well
plates at a density of 2x103 cells per well, and 10 µl
Cell Counting Kit-8 (CCK-8) reagent (Beyotime Institute of
Biotechnology) per well was added at 24 h. After incubation for 2
h, the absorbance was analyzed at 450 nm.
Apoptosis detection
PC12 cells (1x105 cells/well) were
cultured in a 96-well plate and labeled with Annexin V/PI
fluorescent double staining (BD Biosciences) for 30 min as
previously described (19). The
apoptotic cells were analyzed with flow cytometry (FACScan; BD
Biosciences) using FlowJo software (version X; FlowJo LLC).
RNA isolation, reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted using TRIzol®
reagents (Thermo Fisher Scientific, Inc.) and reverse-transcribed
to cDNA using the TaqMan™ MicroRNA Reverse Transcription
kit (Thermo Fisher Scientific, Inc.) and the Prime Script RT
reagent kit (Takara Bio, Inc.) following the manufacturer's
instructions, respectively. The relative expression levels of
miR-218 and spred2 were normalized to U6 and β-actin as an internal
control using the Primer-Script™ RT-PCR kit (Takara),
respectively. The specific primers were as follows: miR-218
forward, 5'-AAGACACCCTGGACGAAGCC-3' and reverse,
5'-ACAACCAGAGTCCACCGGCG-3'; U6 forward,
5'-ATTGGAACGATACAGAGAAGATT-3' and reverse,
5'-GGAACGCTTCACGAATTTG-3'; spred2 forward,
5'-TATATTGTGCGTGTCAAGGCTG-3' and reverse,
5'-GGGGTGCATGACCTTACAGA-3'; β-actin forward,
5'-CATGTACGTTGCTATCCAGGC-3 and reverse,
5'-CGCTCGGTGAGGATCTTCATG-3'.
Western blotting
Protein was extracted through
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology) and protease inhibitors (Roche Diagnostics). Total
protein was measured using a bicinchoninic acid (BCA) assay. The
protein (30 µg per lane) was separated by 10% polyacrylamide gel
electrophoresis and electro-blotted to polyvinylidene fluoride
membranes. After blocking with 5% nonfat milk in TBST for 2 h at
room temperature, the membranes were subsequently incubated
overnight at 4˚C with the indicated antibodies (1:1,000 dilution),
followed by incubation with secondary antibodies (1:2,000 dilution)
for 2 h at room temperature. The primary antibodies included rabbit
polyclonal to spred2 antibody (cat. no. ab153700; Abcam),
monoclonal rabbit p62 antibody (cat. no. 8025; Cell Signaling
Technology, Inc.), monoclonal rabbit LC3-I antibody (cat. no. 4599;
Cell Signaling Technology, Inc.), monoclonal rabbit LC3-II antibody
(cat. no. 3868; Cell Signaling Technology, Inc.), monoclonal rabbit
β-actin antibody (cat. no. 4970; Cell Signaling Technology, Inc.).
The secondary antibodies were HRP-conjugated goat anti-rabbit IgG
antibodies (cat. no. 7074; Cell Signaling Technology, Inc.). The
gray values were measured using ImageJ software (National
Institutes of Health).
Statistical analysis
All data were statistically analyzed by SPSS
software (version 19.0; IBM Corp.) and expressed as mean ± standard
deviation (SD). Unpaired Student's t-test and one-way analysis of
variance (ANOVA) followed by Tukey's post hoc test were used to
assess the difference between groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
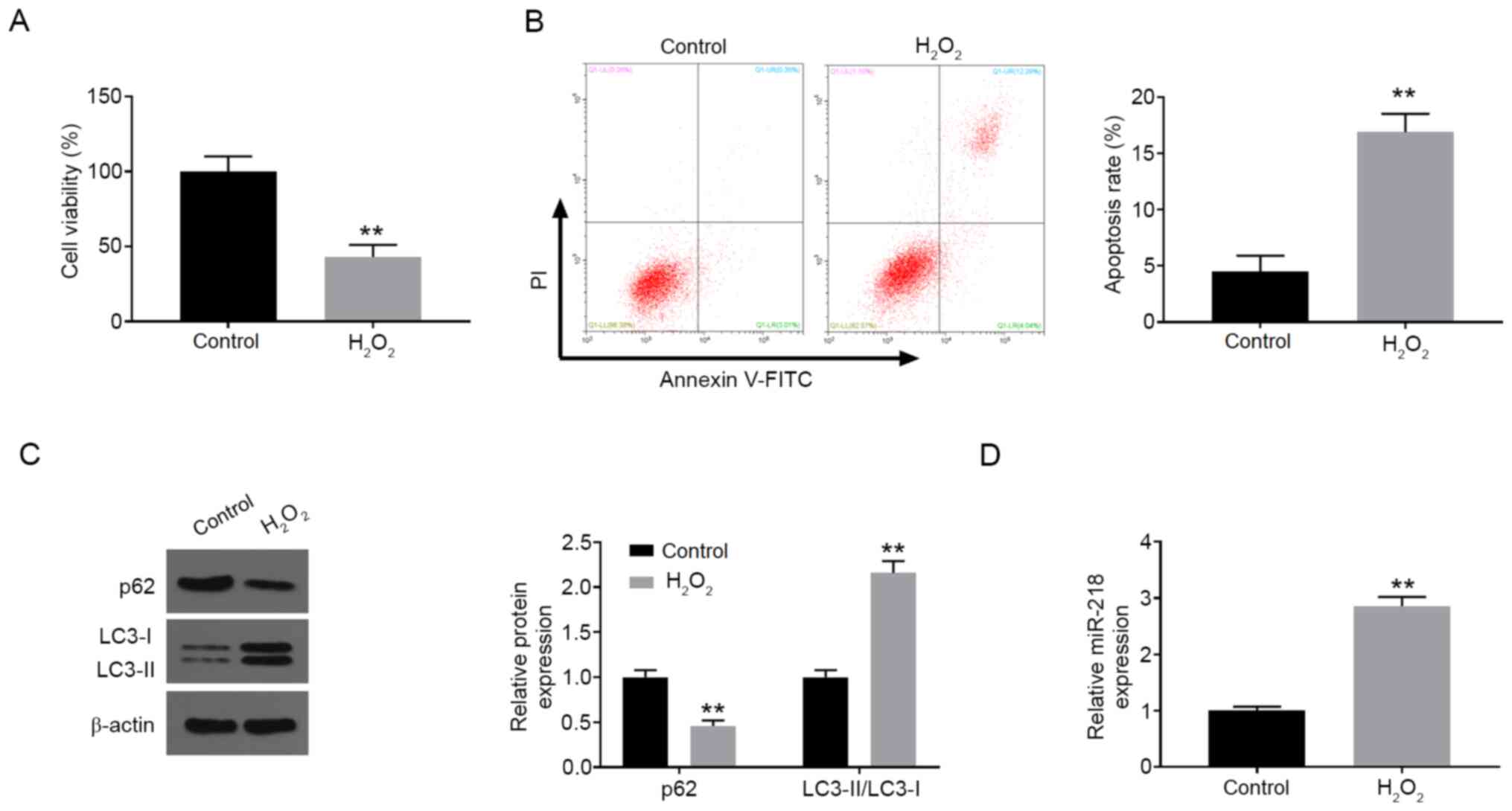
H2O2 induces
PC12 cell injury and miR-218 upregulation
In order to construct the in vitro neuronal
cell injury model, PC12 cells were treated with 200 µM
H2O2 for 24 h. Compared with the control
group, H2O2 treatment markedly suppressed
cell viability detected by CCK-8 assay (Fig. 1A). Furthermore, the percentage of
cell apoptosis was significantly increased after
H2O2 treatment compared with the control
group (Fig. 1B). Moreover, the
expression level of p62 was dramatically downregulated, while the
ratio of LC3-II/I was observably increased after
H2O2 treatment in comparison with the control
group (Fig. 1C). In addition,
H2O2 stimulation also resulted in increased
expression of miR-218 in PC12 cells (Fig. 1D).
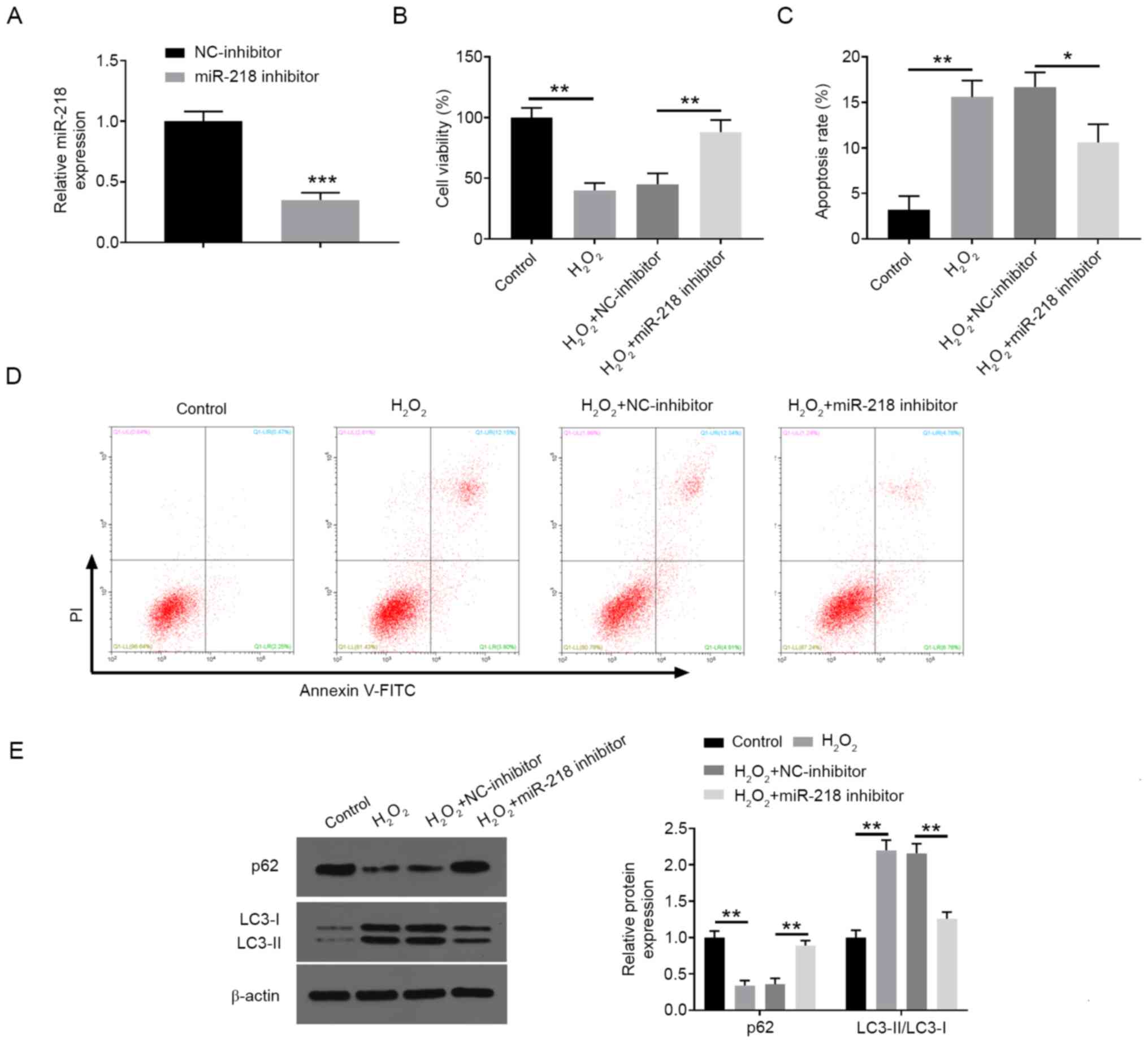
Inhibition of miR-218 increases
proliferation and decreases apoptosis and autophagy in
H2O2-treated PC12 cells
To confirm whether miR-218 is involved in the
regulation of H2O2-disposed neuronal cell
injury, the expression of miR-218 was thus silenced in PC12 cells
by transfection with miR-218 inhibitor (Fig. 2A). In the subsequent experiments,
the knockdown of miR-218 markedly inhibited
H2O2-induced decrease in cell viability
(Fig. 2B) and alleviated
H2O2-induced cell apoptosis (Fig. 2C and D), as well as ameliorating
H2O2-induced cell autophagy by reversing the
changes in the expression levels of autophagy-associated proteins
(Fig. 2E).
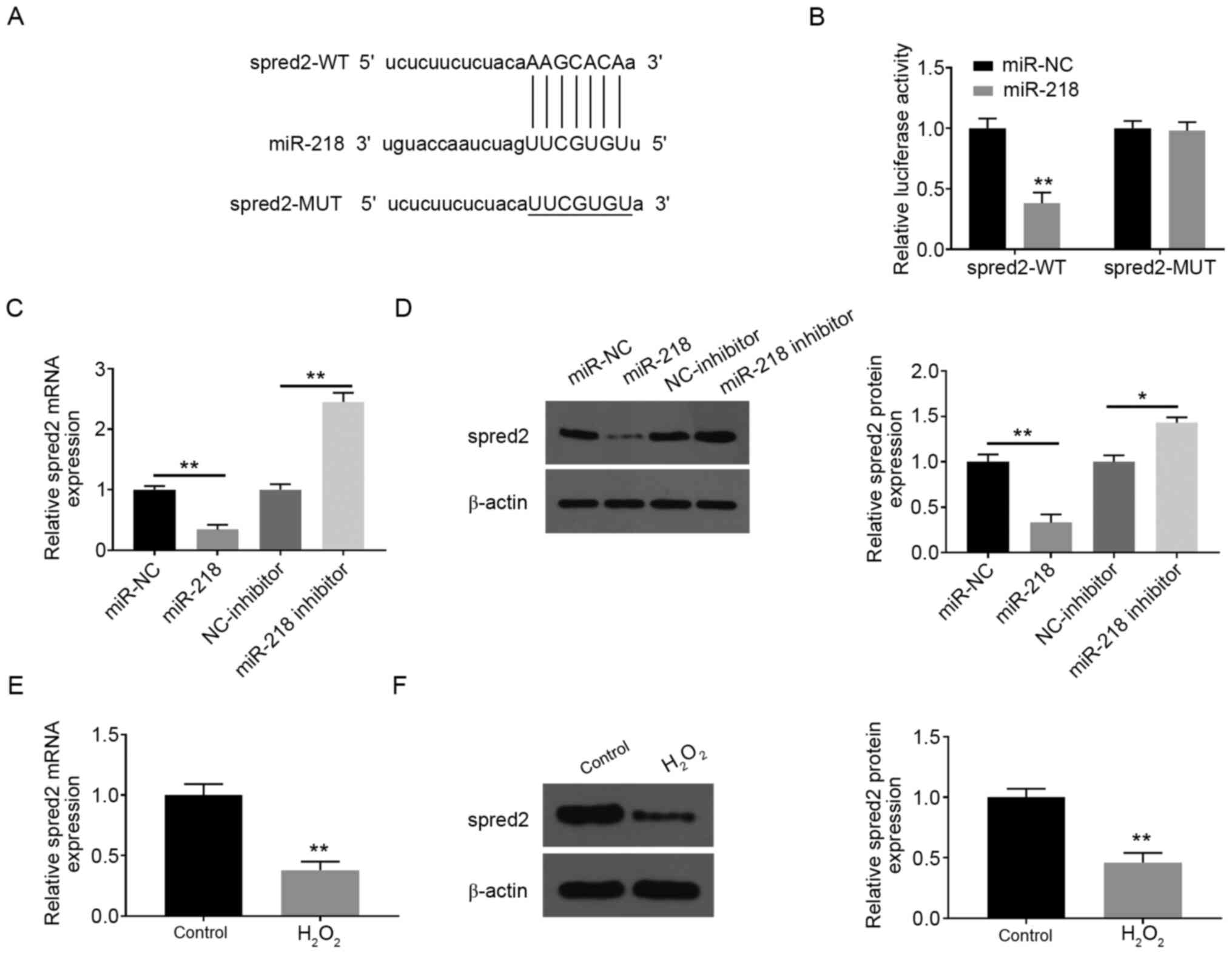
Spred2 is a direct target of miR-218
in PC12 cells
Using the online database starBase, spred2 was
predicted as the potential target of miR-218 (Fig. 3A). The results of luciferase assay
in PC12 cells further verified that miR-218 mimics significantly
suppressed the luciferase report activity of spred2-wt, but not
spred2-mut (Fig. 3B). The
overexpression of miR-218 was achieved in PC12 cells by
transfection with miR-218 mimic, which was further confirmed by
RT-qPCR (Fig. S1). Additionally,
the mRNA and protein levels of spred2 were memorably decreased
after miR-218 overexpression, but were elevated by miR-218
silencing (Fig. 3C and D). The expression levels of spred2 in PC12
cells were detected after H2O2 treatment. The
present study data showed that the expression of spred2 at mRNA and
protein levels was markedly decreased following treatment with
H2O2 compared with the control group
(Fig. 3E and F).
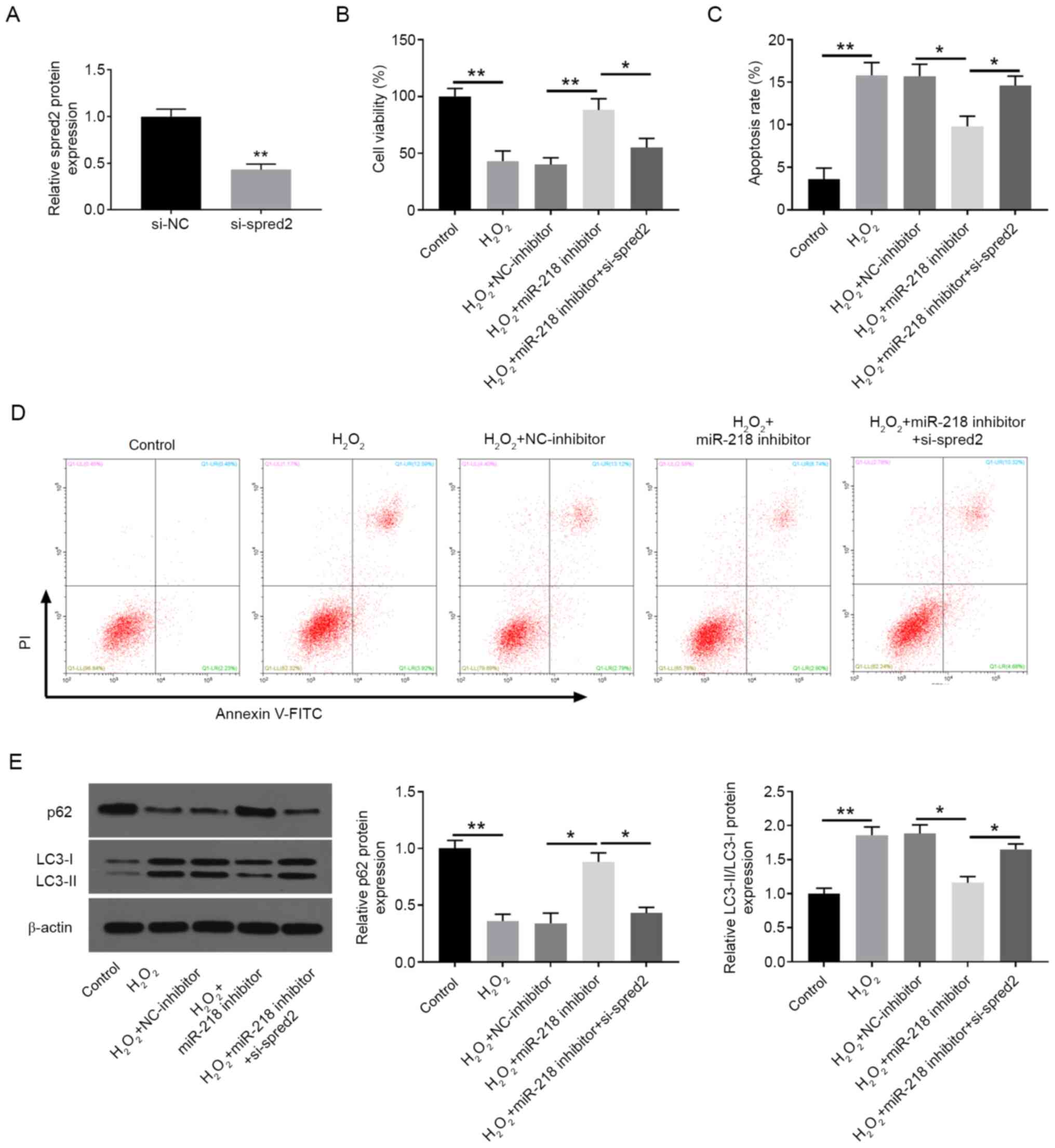
miR-218 enhances
H2O2-induced PC12 cell injury by targeting
spred2
Subsequently, it was investigated whether miR-218
regulated H2O2-induced injury via decreasing
spred2 expression. Spred2 was knocked down in PC12 cells by
transfection with si-spred2 (Fig.
4A). As shown in Fig. 4B,
spred2 inhibition significantly reversed miR-218
downregulation-induced promotion of cell proliferation in
H2O2-exposed PC12 cells (Fig. 4B). Moreover, the inhibitory effects
of miR-218 inhibition on cell apoptosis (Fig. 4C and D) and autophagy (Fig. 4E) were also reversed by spred2
knockdown in H2O2-treated PC12 cells.
Discussion
In the current study, a cell injury model induced by
H2O2 was constructed to investigate the
protective role of miR-218 knockdown in
H2O2-injured PC12 cells. The study initially
indicated that miR-218 expression was significantly increased in
PC12 cells after H2O2 treatment. Moreover,
suppression of miR-218 alleviated
H2O2-induced PC12 cell injury by enhancing
cell proliferation and inhibiting apoptosis and autophagy.
Furthermore, it was identified that spred2 was a direct target of
miR-218 and was negatively regulated by miR-218. Further
investigation demonstrated that miR-218 mediated
H2O2-induced PC12 cell injury through
regulating spred2.
SCI is a severe neurological disease with a high
disability rate (20). To date,
emerging effort has been made to identify critical regulators,
which are involved in the pathogenesis of SCI, and the
pharmacological therapies targeting these regulators have been
developed for clinical trials to prevent neurological dysfunction
(21,22). Recently, there is emerging evidence
that a larger number of miRNAs are aberrantly expressed following
SCI and might play a key role in the development of SCI (23). There are abundant cancer-associated
research regarding the anti-tumor roles of miR-218 (24,25). A
previous study by Zhu et al (13) demonstrated that downregulation of
miR-218 protected against OGD/R-induced injuries of PC12 cells
through lowering inflammation, oxidative stress, apoptosis and
maintenance of endovascular homeostasis; nevertheless, the
involvement of miR-218 in SCI remains unclear.
In recent years, several studies have proposed that
oxidative stress caused by free radicals is implicated in the
pathophysiological processes of secondary injury following SCI. The
suppression of oxidative injury is therefore considered effective
therapeutic strategy to improve SCI. Exposure of neurons to
H2O2 results in oxidative stress and the
activation of a cascade of intracellular toxic events, ultimately
resulting in cell death (26).
Autophagy is a metabolic process degrading long-lived proteins and
organelles, which is not only a highly-conserved defense mechanism,
but also functions as a mechanism of programmed cell death
involving the degradation of damaged organelles and misfolded
proteins in eukaryotes, which is associated with numerous
pathological or physiological processes (27,28),
including the pathogenesis of SCI (29). In the present study,
H2O2 exposure at a concentration of 200 µM
induced proliferation inhibition and increase in apoptosis and
autophagy in PC12 cells, indicating that H2O2
treatment successfully induced cell injury in PC12 cells. It was
demonstrated that miR-218 expression was significantly augmented in
H2O2-stimulated PC12 cells. Employing
H2O2-induced PC12 cells, it was also
discovered that miR-218 inhibition ameliorated
H2O2-induced decrease in cell viability and
increase in apoptosis and autophagy.
Furthermore, the present study findings also
confirmed that spred2 might be a target gene of miR-218, by means
of bioinformatics prediction and luciferase reporter assays. Spred2
is a negative regulator of the ERK-MAPK pathway and is essential
for cell survival, inflammation and angiogenesis (30,31).
Moreover, spred2 is recently identified as a vital regulator of
autophagy and cell death by interacting with LC3 and p62 (32,33).
Studies showed that spred2 interaction with NBR1 downregulated
fibroblast growth factor (FGF) signaling in the regulation of
receptor trafficking (34). Chen
et al (35) demonstrated
that miR-210 directly inhibited spred2 expression, and spred2
decreased oxLDL-mediated migration of vascular cells via the
ERK/c-Fos/MMPs pathway. Spred2 was reported as a target gene of
miR-221-3p, and was involved in the development of LPS-induced lung
inflammation by negatively regulating the ERK1/2 pathway (36,37).
Spred2 was also identified as the downstream target of miR-1246,
wherein downregulated spred2 further reversed the inhibition of the
MAPK pathway (38). The expression
of spred2 in PC12 cells was downregulated after
H2O2 stimulation in the present study. Loss-
and gain-of-function analyses further revealed that spred2
knockdown overturned the anti-apoptotic and anti-autophagy effects
of miR-218 suppression in H2O2-stimulated
PC12 cells. Following results, it was shown that miR-218 deficiency
impaired autophagy and apoptosis by targeting spred2 in PC12 cells
under H2O2 stimulation. According to the
previous studies, differentiated PC12 cells appeared to be
significantly more resistant to H2O2 than
naive PC12 cells (39). Further
research is necessary to investigate the detected mechanisms in
differentiated PC12 cells.
Overall, despite the lack of clinical data and in
vivo experiments, as well as the use of only one cell line, the
present study findings provided a novel insight into the effects
and mechanisms of miR-218 in H2O2-induced
PC12 cell injury. It was preliminarily confirmed that the
downregulation of miR-218 may protect PC12 cells from
H2O2-disposed injury via targeting spred2.
The present study is the first to demonstrate that the
miR-218-spred2 axis may be a promising therapeutic strategy for
SCI.
Supplementary Material
Confirmation of miR-218 overexpression
in PC12 cells. The mRNA expression of miR-218 was determined in
PC12 cells transfected with miR-NC or miR-218 mimic, using reverse
transcription-quantitative PCR analysis. All experiments were
repeated three times. Data are expressed as mean ± SD.
#x002A;**P<0.001. miR, microRNA; NC, negative
control.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed in the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
DC and CL were involved in the conception and
design, acquisition of data, and drafting the article, and
performed the analysis and interpretation of data, and revised the
manuscript critically for important intellectual content. RL
revised the manuscript, analyzed the data and gave approval of the
version to be published and agreed to be accountable for all
aspects of the work. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Schwab JM, Maas AIR, Hsieh JTC and Curt A:
Raising awareness for spinal cord injury research. Lancet Neurol.
17:581–582. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Jendelova P: Therapeutic strategies for
spinal cord injury. Int J Mol Sci. 19(3200)2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Venkatesh K, Ghosh SK, Mullick M,
Manivasagam G and Sen D: Spinal cord injury: Pathophysiology,
treatment strategies, associated challenges, and future
implications. Cell Tissue Res. 377:125–151. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wang JL, Luo X and Liu L: Targeting CARD6
attenuates spinal cord injury (SCI) in mice through inhibiting
apoptosis, inflammation and oxidative stress associated ROS
production. Aging (Albany NY). 11:12213–12235. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Guan B, Chen R, Zhong M, Liu N and Chen Q:
Protective effect of oxymatrine against acute spinal cord injury in
rats via modulating oxidative stress, inflammation and apoptosis.
Metab Brain Dis. 35:149–157. 2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Vishnoi A and Rani S: miRNA biogenesis and
regulation of diseases: An overview. Methods Mol Biol. 1509:1–10.
2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Jiao S, Liu Y, Yao Y and Teng J: miR-124
promotes proliferation and neural differentiation of neural stem
cells through targeting DACT1 and activating Wnt/β-catenin
pathways. Mol Cell Biochem. 449:305–314. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yuan Z, Zhong L, Liu D, Yao J, Liu J,
Zhong P, Yao S, Zhao Y, Li L, Chen M, et al: miR-15b regulates cell
differentiation and survival by targeting CCNE1 in APL cell lines.
Cell Signal. 60:57–64. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Dai J, Xu LJ, Han GD, Sun HL, Zhu GT,
Jiang HT, Yu GY and Tang XM: miR-137 attenuates spinal cord injury
by modulating NEUROD4 through reducing inflammation and oxidative
stress. Eur Rev Med Pharmacol Sci. 22:1884–1890. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Jiao G, Pan B, Zhou Z, Zhou L, Li Z and
Zhang Z: MicroRNA-21 regulates cell proliferation and apoptosis in
H2O2-stimulated rat spinal cord neurons. Mol
Med Rep. 12:7011–7016. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Liu D and Bao F: Hydrogen peroxide
administered into the rat spinal cord at the level elevated by
contusion spinal cord injury oxidizes proteins, DNA and membrane
phospholipids, and induces cell death: Attenuation by a
metalloporphyrin. Neuroscience. 285:81–96. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhu H, Wang X and Chen S: Downregulation
of miR-218-5p protects against oxygen-glucose
deprivation/reperfusion-induced injuries of PC12 cells via
upregulating N-myc downstream regulated gene 4 (NDRG4). Med Sci
Monit. 26(e920101)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhu L, Tu H, Liang Y and Tang D: miR-218
produces anti-tumor effects on cervical cancer cells in vitro.
World J Surg Oncol. 16(204)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Cao Q, Dong P and Wang Y, Zhang J, Shi X
and Wang Y: miR-218 suppresses cardiac myxoma proliferation by
targeting myocyte enhancer factor 2D. Oncol Rep. 33:2606–2612.
2015.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Li L and Zhao G: Downregulation of
microRNA-218 relieves neuropathic pain by regulating suppressor of
cytokine signaling 3. Int J Mol Med. 37:851–858. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yin D, Zheng X, Zhuang J, Wang L, Liu B
and Chang Y: Downregulation of long noncoding RNA Sox2ot protects
PC-12 cells from hydrogen peroxide-induced injury in spinal cord
injury via regulating the miR-211-myeloid cell leukemia-1 isoform2
axis. J Cell Biochem. 119:9675–9684. 2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Guo Z, Li L, Gao Y, Zhang X and Cheng M:
Overexpression of lncRNA ANRIL aggravated hydrogen
peroxide-disposed injury in PC-12 cells via inhibiting
miR-499a/PDCD4 axis-mediated PI3K/Akt/mTOR/p70S6K pathway. Artif
Cells Nanomed Biotechnol. 47:2624–2633. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhang Y, Sun B, Zhao L, Liu Z, Xu Z, Tian
Y and Hao C: Up-regulation of miRNA-148a inhibits proliferation,
invasion, and migration while promoting apoptosis of cervical
cancer cells by down-regulating RRS1. Biosci Rep.
39(BSR20181815)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Eckert MJ and Martin MJ: Trauma: Spinal
cord injury. Surg Clin North Am. 97:1031–1045. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li Z, Wu F, Xu D, Zhi Z and Xu G:
Inhibition of TREM1 reduces inflammation and oxidative stress after
spinal cord injury (SCI) associated with HO-1 expressions. Biomed
Pharmacother. 109:2014–2021. 2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Abou-El-Hassan H, Bsat S, Sukhon F, Assaf
EJ, Mondello S, Kobeissy F, Wang KKW, Weiner HL and Omeis I:
Protein degradome of spinal cord injury: Biomarkers and potential
therapeutic targets. Mol Neurobiol. 57:2702–2726. 2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Pinchi E, Frati A, Cantatore S, D'Errico
S, Russa R, Maiese A, Palmieri M, Pesce A, Viola RV, Frati P and
Fineschi V: Acute spinal cord injury: A systematic review
investigating miRNA FAMILIES INVolved. Int J Mol Sci.
20(1841)2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lu W, Wan X, Tao L and Wan J: Long
non-coding RNA HULC promotes cervical cancer cell proliferation,
migration and invasion via miR-218/TPD52 axis. Onco Targets Ther.
13:1109–1118. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li XC, Hai JJ, Tan YJ, Yue QF and Liu LJ:
miR-218 suppresses metastasis and invasion of endometrial cancer
via negatively regulating ADD2. Eur Rev Med Pharmacol Sci.
23:1408–1417. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wen YD, Wang H, Kho SH, Rinkiko S, Sheng
X, Shen HM and Zhu YZ: Hydrogen sulfide protects HUVECs against
hydrogen peroxide induced mitochondrial dysfunction and oxidative
stress. PLoS One. 8(e53147)2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ravanan P, Srikumar IF and Talwar P:
Autophagy: The spotlight for cellular stress responses. Life Sci.
188:53–67. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Parzych KR and Klionsky DJ: An overview of
autophagy: Morphology, mechanism, and regulation. Antioxid Redox
Signal. 20:460–473. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ray SK: Modulation of autophagy for
neuroprotection and functional recovery in traumatic spinal cord
injury. Neural Regen Res. 15:1601–1612. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Yoshimura A: Regulation of cytokine
signaling by the SOCS and Spred family proteins. Keio J Med.
58:73–83. 2009.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Takahashi S, Yoshimura T, Ohkura T,
Fujisawa M, Fushimi S, Ito T, Itakura J, Hiraoka S, Okada H,
Yamamoto K and Matsukawa A: A novel role of spred2 in the colonic
epithelial cell homeostasis and inflammation. Sci Rep.
6(37531)2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Jiang K, Liu M, Lin G, Mao B, Cheng W, Liu
H, Gal J, Zhu H, Yuan Z, Deng W, et al: Tumor suppressor spred2
interaction with LC3 promotes autophagosome maturation and induces
autophagy-dependent cell death. Oncotarget. 7:25652–25667.
2016.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ullrich M, Aßmus B, Augustin AM, Häbich H,
Abeßer M, Martin Machado J, Werner F, Erkens R, Arias-Loza AP,
Umbenhauer S, et al: Spred2 deficiency elicits cardiac arrhythmias
and premature death via impaired autophagy. J Mol Cell Cardiol.
129:13–26. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Mardakheh FK, Yekezare M, Machesky LM and
Heath JK: Spred2 interaction with the late endosomal protein NBR1
down-regulates fibroblast growth factor receptor signaling. J Cell
Biol. 187:265–277. 2009.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen KC, Liao YC, Wang JY, Lin YC, Chen CH
and Juo SH: Oxidized low-density lipoprotein is a common risk
factor for cardiovascular diseases and gastroenterological cancers
via epigenomical regulation of microRNA-210. Oncotarget.
6:24105–24118. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Xu Y, Ito T, Fushimi S, Takahashi S,
Itakura J, Kimura R, Sato M, Mino M, Yoshimura A and Matsukawa A:
Spred-2 deficiency exacerbates lipopolysaccharide-induced acute
lung inflammation in mice. PLoS One. 9(e108914)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hu L, Ye H and Liao J: LncRNA TUG1
reverses LPS-induced cell apoptosis and inflammation of macrophage
via targeting miR-221-3p/SPRED2 axis. Biosci Biotechnol Biochem.
84:2458–2465. 2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Peng W, Li J, Chen R, Gu Q, Yang P, Qian
W, Ji D, Wang Q, Zhang Z, Tang J and Sun Y: Upregulated METTL3
promotes metastasis of colorectal cancer via miR-1246/SPRED2/MAPK
signaling pathway. J Exp Clin Cancer Res. 38(393)2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Sung YJ, Cheng CL, Chen CS, Huang HB,
Huang FL, Wu PC, Shiao MS and Tsay HJ: Distinct mechanisms account
for beta-amyloid toxicity in PC12 and differentiated PC12 neuronal
cells. J Biomed Sci. 10:379–388. 2003.PubMed/NCBI View Article : Google Scholar
|