Introduction
Knee osteoarthritis (KOA) is a common degenerative
joint disease caused by several factors, such as age, obesity,
inflammation and trauma, and is characterized by the degeneration
of articular cartilage (1). KOA
results in the destruction of articular cartilage, and triggers
pathological changes in the surrounding synovial membrane,
ligaments and soft tissue (2).
These pathological changes result in the damage of articular
cartilage and the potential destruction of the entire articular
surface (3). At present, there are
no available strategies for the prevention and treatment of KOA.
Thus, further investigations into the mechanisms underlying KOA may
provide novel treatment strategies, and research into the diagnosis
of KOA and further treatment options is ongoing in a number of
countries (4).
Dual specificity protein phosphatase 4 (DUSP4), also
known as MAPK phosphatase 2 (MKP2), is a member of the DUSP family
(5). DUSP4 serves an important
role in the regulation of cell proliferation and differentiation
(6). As a negative regulator of
the MAPK signaling pathway, DUSP4 also plays a tumor-suppressive
role in the majority of human cancer types, including colorectal,
breast and pancreatic cancer (7).
The results of a previous study demonstrated that DUSP4
overexpression in vascular endothelial cells reduced oxidative
stress-induced cell damage and apoptosis to protect the heart
following cardiac ischemia or reperfusion (8). In lipopolysaccharide (LPS)-induced
acute lung injury, interference with DUSP4 expression promoted the
expression of inflammatory cytokines, such as TNF α, IL-1 and
IL-6(9). These results suggested
that DUSP4 may serve an important role in the regulation of cell
inflammation, injury and apoptosis in numerous diseases, including
cancer, and inflammatory and vascular diseases. Osteoarthritis (OA)
is characterized by the progressive degeneration and loss of
articular cartilage caused by chondrocyte apoptosis and excessive
degradation of the cartilage matrix. Cellular inflammatory factors
are the main factors that promote excessive degradation of the
cartilage matrix (10).
Furthermore, the results of a previous study demonstrated that
antisense long-non coding RNA in the INK4 locus regulated the
proliferation and apoptosis of OA-related cells via the
microRNA-122-5p/DUSP4 axis, indirectly suggesting that DUSP4 may be
involved in the occurrence of OA (11). However, the specific mechanism
underlying DUSP4 in KOA is yet to be fully elucidated.
In the present study, chondrocytes were induced with
LPS to establish an OA injury model. Furthermore, the expression of
DUSP4 was determined in the OA injury model to uncover the
regulatory mechanism underlying DUSP4, thus providing novel
insights into the occurrence and development of KOA.
Materials and methods
Cell culture and model induction
Murine cartilage ATDC5 cells were purchased from the
Institute of Biochemistry and Cell Biology at the Chinese Academy
of Sciences. Cells were maintained in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37˚C in a humidified incubator containing 5%
CO2. Cells were pretreated with 100 ng/ml phorbol
12-myristate 13-acetate (PMA; cat. no. HY-18739; MedChemExpress)
for 2 h and then treated with 5 µg/ml LPS for 24 h at 37˚C, while
untreated cells served as the control group.
GSE1919 microarray analysis
The Gene Expression Omnibus (GEO) database
(https://www.ncbi-nlm-nih-gov.elib.tcd.ie/geo/) was
used to search for the key word ‘Osteoarthritis’, and the GSE1919
chip data were selected for further analysis (12). The healthy samples (n=5) and OA
samples (n=5) were selected. The data were analyzed using GEO2R
online tools, and GraphPad Prism version 8 (GraphPad Software,
Inc.) was used to aggregate the data into a histogram.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from ATDC5 cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), according to the manufacturer's protocols. Total RNA was
reverse transcribed into cDNA using the PrimeScript RT Reagent kit
(Takara Bio, Inc.). cDNA was synthesized from total RNA (0.5 µg) at
48˚C for 30 min, followed by 95˚C for 10 min. The relative Mrna
expression levels were determined using the One Step
SYBR® PrimeScript® PLUS RT-RNA PCR Kit
(Takara Biotechnology Co., Ltd.). The synthesized cDNA (0.7 µg) was
subjected to qPCR under the following thermocycling conditions:
Initial denaturation at 95˚C for 15 min, followed by 45 cycles of
95˚C for 10 sec, 66˚C for 20 sec, and 72˚C for 10 sec, according to
the manufacturer's instructions. GAPDH was used to normalize the
expression levels of the target genes and data were calculated by
the 2-ΔΔCq method (13). The primer sequences used for qPCR
were as follows: DUSP4 forward, 5'-TGGTTCATGGAAGCCATAGAG-3' and
reverse, 5'-CCCGTTTCTTCATCATCAGG-3'; and GAPDH forward,
5'-AACTTTGGCATTGTGGAAGG-3' and reverse,
5'-GGAGACAACCTGGTCCTCAG-3'.
Western blot analysis
Total proteins were extracted from ATDC5 cells using
RIPA buffer (Beijing Solarbio Science & Technology Co., Ltd.)
and the protein concentration was determined using a BCA protein
assay kit (Beijing Solarbio Science & Technology Co., Ltd.).
Subsequently, 30 µg/lane of total protein extracts were separated
via SDS-PAGE on 10% gel and transferred onto nitrocellulose
membranes. Following blocking with 5% non-fat milk for 1 h at room
temperature, the membranes were incubated with the following
primary antibodies: anti-DUSP4 (1:1,000; cat. no. ab216576; Abcam),
anti-p38 (1:1,000; cat. no. ab170099; Abcam), anti-phosphorylated
(p)-p38 (1:1,000; cat. no. ab178867; Abcam), anti-ERK1/2 (1:1,000;
cat. no. ab184699; Abcam), anti-p-ERK1/2 (1:1,000, cat. no.
ab278538; Abcam), anti-JNK (1:1,000; cat. no. ab110724; Abcam),
anti-p-JNK (1:1,000; cat. no. ab215208; Abcam), anti-IL-6 (1:1,000;
cat. no. ab233706; Abcam), anti-TNF-α (1:1,000; cat. no. ab183218;
Abcam), anti-IL-1β (1:1,000; cat. no. ab216995; Abcam), anti-Bcl-2
(1:1,000; cat. no. ab32124; Abcam), anti-Bax (1:1,000; cat. no.
ab182733; Abcam), anti-cleaved caspase 3 (1:1,000; cat. no.
ab32042; Abcam), anti-caspase 3 (1:1,000; cat. no. ab32351; Abcam)
and GAPDH (1:1,000; cat. no. ab8245; Abcam) at 4˚C overnight.
Following washing with PBS containing 0.05% Tween-20 (1,000 ml PBS
and 5 ml Tween-20) three times, the membranes were incubated with
the corresponding HRP-conjugated goat anti-rabbit (1:5,000; cat.
no. LK2001; Sungene Biotech Co., Ltd.) or anti-mouse (1:5,000, cat.
no. LK2003; Sungene Biotech Co., Ltd.) secondary antibodies for 1 h
at room temperature. The immunoblots were detected with an ECL kit
(Advansta, Inc.) and the intensity of the bands was analyzed using
ImageJ software (version 146; National Institutes of Health). GAPDH
served as the loading control for normalization.
Cell transfection
The DUSP4 overexpression plasmid (ov-DUSP4) was
designed and synthesized by Shanghai GeneChem Co., Ltd., using the
pBluescript vector. A total of 1 µg ov-DUSP4 plasmid or an empty
vector (NC) were transfected into ATDC5 cells using
Lipofectamine® 2000 at 37˚C for 48 h (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
Following incubation for 48 h, cells were used for subsequent
experiments.
Cell Counting Kit-8 (CCK-8)
ATDC5 cells were seeded into 96-well microplates at
a density of 1x104 cells/well at 37˚C for 12 h.
Following cell transfection and treatment with LPS and PMA, 10 µl
CCK-8 reagent (Thermo Fisher Scientific, Inc.) was added to each
well and cells were cultured at 37˚C for 4 h. The Infinite™ M200
microplate reader (Tecan Group, Ltd.) was utilized to measure the
optical density of each well at a wavelength of 460 nm.
Reactive oxygen species (ROS), lactate
dehydrogenase (LDH), superoxide dismutase (SOD) and malondialdehyde
(MDA) assays
The levels of ROS, LDH, SOD and MDA were determined
using the ROS/Superoxide Detection Assay kit (cat. no. ab113851;
Abcam), LDH cytotoxicity assay kit (cat. no. C0017, Beyotime
Institute of Biotechnology), total SOD assay kit (cat. no. BC0175,
Beijing Solarbio Science & Technology Co., Ltd.) and MDA kit
(cat. no. E0597Ge, Wuhan EIAab Science Co., Ltd.) respectively,
according to the corresponding manufacturers' protocols.
ELISA
The levels of IL-1β (cat. no. MLB00C), IL-6 (cat.
no. D6050) and TNF-α (cat. no. MTA00B) in the cell supernatant were
measured using the IL-1β, IL-6 and TNF-α ELISA kits (R&D
Systems, Inc.), according to the manufacturer's protocol.
TUNEL assay
Cell apoptosis was evaluated using a TUNEL assay kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. The cells were fixed with Immunol Staining
Fix Solution (Beyotime Institute of Biotechnology) for 30 min at
37˚C, followed by washing with PBS (Thermo Fisher Scientific,
Inc.). Subsequently, cells were incubated with PBS containing 0.3%
Triton X-100 (Sigma-Aldrich; Merck KGaA) at room temperature for 5
min and were washed again with PBS followed by TUNEL solution for 1
h at 37˚C. Apoptotic cell nuclei were stained with
fluorescein-labeled dUTP solution (Invitrogen; Thermo Fisher
Scientific, Inc.) at 37˚C for 60 min, while all cell nuclei were
stained with 5 mg/ml DAPI for 3 min at 37˚C. A total of 10 fields
were randomly selected and observed by fluorescence microscope
(magnification, x200; CKX53; Olympus Corporation). TUNEL-positive
staining indicated apoptotic cells.
Statistical analysis
All data are expressed as the mean ± standard
deviation. One-way ANOVA followed by Tukey's post hoc test was used
for comparisons among multiple groups. P<0.05 was considered to
indicate a statistically significant difference. All analyses were
carried out using SPSS 22.0 statistical software (IBM Corp.). Each
experiment was repeated three times.
Results
DUSP4 is downregulated in OA
chondrocytes and DUSP4 overexpression inhibits the MAPK signaling
pathway
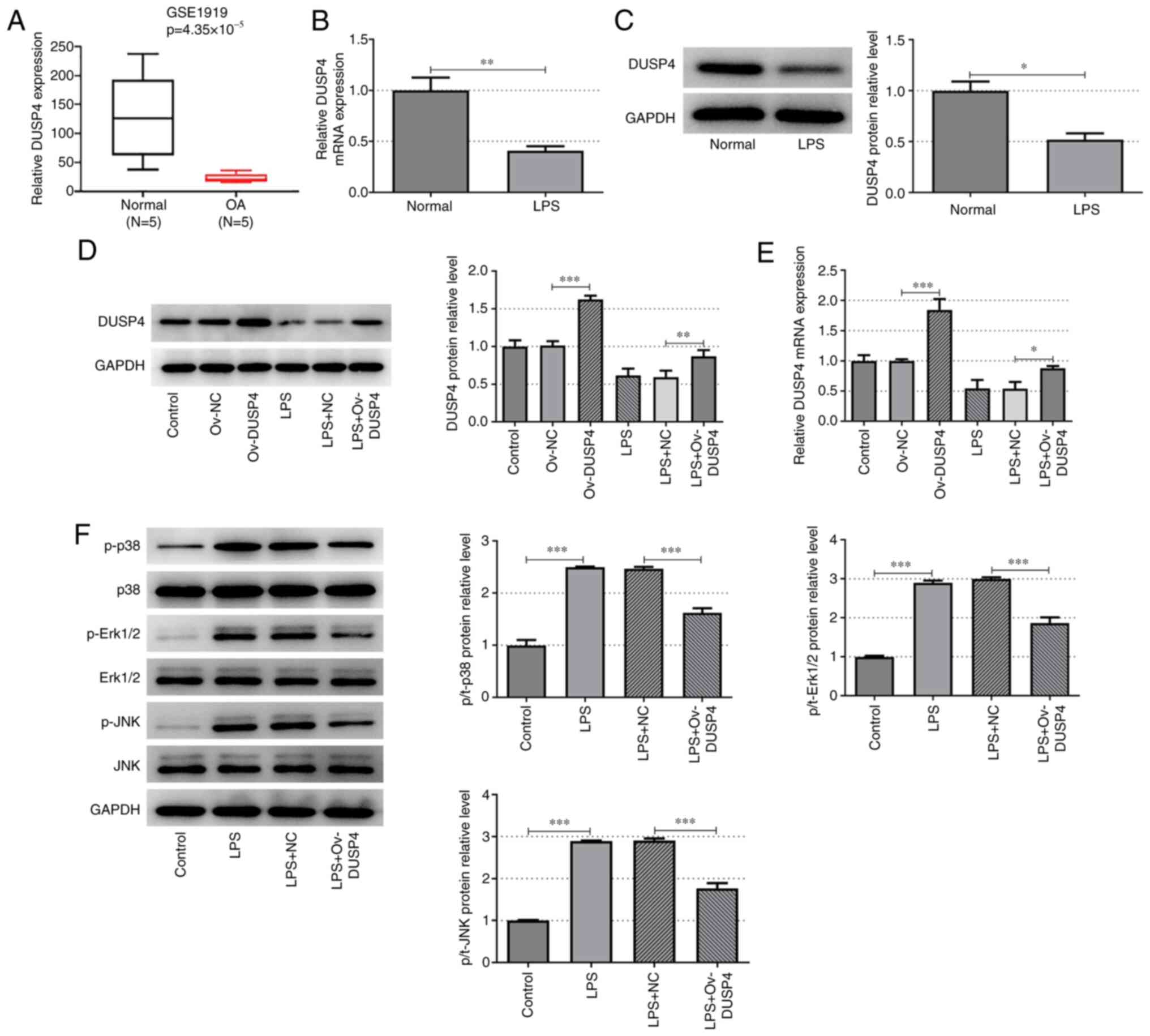
GSE1919 microarray analysis revealed that DUSP4 was
significantly downregulated in synovial tissues of patients with OA
(Fig. 1A). Additionally, in
vitro experiments demonstrated that the expression of DUSP4 was
significantly decreased in LPS-induced ATDC5 cells compared with
the control group at both the protein and mRNA level (Fig. 1B and C). These results indicated that the
expression of DUSP4 was decreased in OA chondrocytes. The ov-DUSP4
plasmid was constructed and the subsequent transfection efficiency
was verified by RT-qPCR and western blot analyses in ATDC5 cells.
DUSP4 was significantly overexpressed in ATDC5 cells and
LPS-induced ATDC5 cells compared with the corresponding control
group at both the protein and mRNA level (Fig. 1D and E). Subsequently, cells were divided into
four groups: i) Control; ii) LPS; iii) LPS + NC; and iv) LPS +
ov-DUSP4. The expression of MAPK pathway-related proteins was
assessed by western blot analysis. The expression levels of p-p38,
p-ERK1/2 and p-JNK were significantly increased in the LPS group
compared with the control group. Furthermore, the expression of
p-p38, p-ERK1/2 and p-JNK was significantly downregulated following
DUSP4 overexpression, compared with the LPS + NC group (Fig. 1F). The results of the present study
suggested that DUSP4 may serve an important role in promoting
chondrocyte injury in OA by inhibition of the MAPK signaling
pathway.
DUSP4 enhances cell viability and
attenuates oxidative stress and inflammatory responses in OA
chondrocytes via the MAPK signaling pathway
To further investigate whether DUSP4 exerts its
regulatory effects on OA via the MAPK signaling pathway, cells were
treated with PMA, an agonist of the MAPK signaling pathway. Cells
were divided into five groups: i) Control; ii) LPS; iii) LPS + NC;
iv) LPS + ov-DUSP4; and v) LPS + ov-DUSP4 + PMA. A CCK-8 assay was
performed to assess cell viability and the results demonstrated
that cell viability was significantly reduced in the LPS group
compared with the control group. Furthermore, cell viability in the
LPS + ov-DUSP4 group was significantly increased compared with the
LPS + NC group, and was significantly attenuated following cell
treatment with PMA compared with the LPS + ov-DUSP4 group (Fig. 2A).
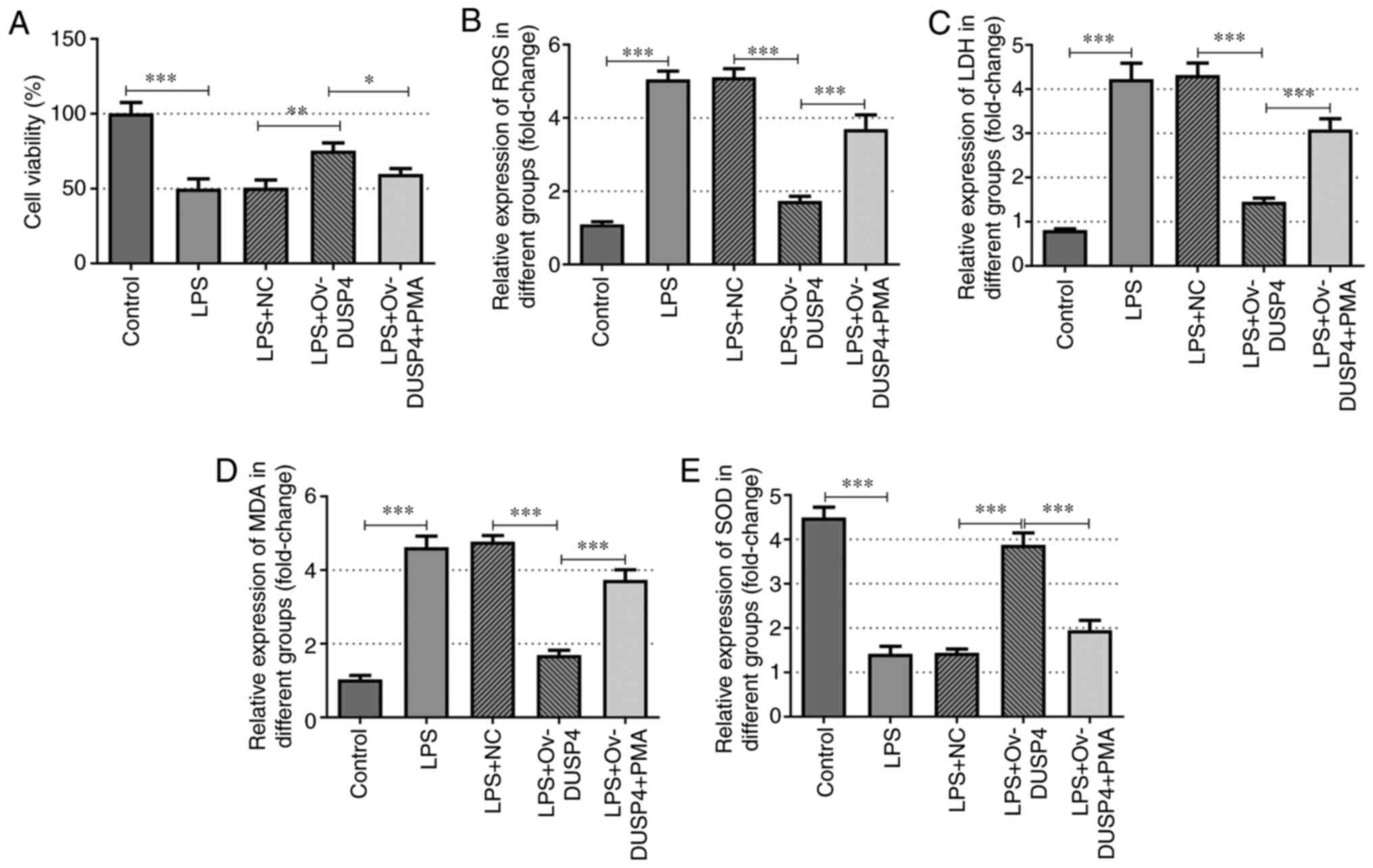 | Figure 2DUSP4 increases the activity and
reduces the oxidative stress level of osteoarthritis chondrocytes
via the MAPK signaling pathway. (A) MTT assays were used to detect
the cell viability. ELISA assays detected the expression of (B)
ROS, (C) LDH, (D) MDA and (E) SOD. *P<0.05;
**P<0.01; ***P<0.001. LPS,
lipopolysaccharide; NC, negative control; Ov, overexpression;
DUSP4, dual specificity protein phosphatase 4; PMA, phorbol
12-myristate 13-acetate; ROS, reactive oxygen species; LDH, lactate
dehydrogenase; MDA, malondialdehyde; SOD, superoxide dismutase. |
The generation of ROS, the levels of LDH and MDA and
the activity of SOD in cells were determined by ELISA. The results
demonstrated that the levels of ROS, LDH and MDA was significantly
enhanced in the LPS group, while the activity of SOD was markedly
decreased compared with the control group. Additionally, the
production of ROS, LDH and MDA was significantly diminished in the
LPS + ov-DUSP4 group, whereas the production of SOD was
significantly enhanced, compared with the LPS + NC group. In
addition, the production of ROS, LDH and MDA was notably decreased
in the LPS + ov-DUSP4 group compared with the LPS + ov-DUSP4 + PMA
group, while the opposite trend was observed in the activity of SOD
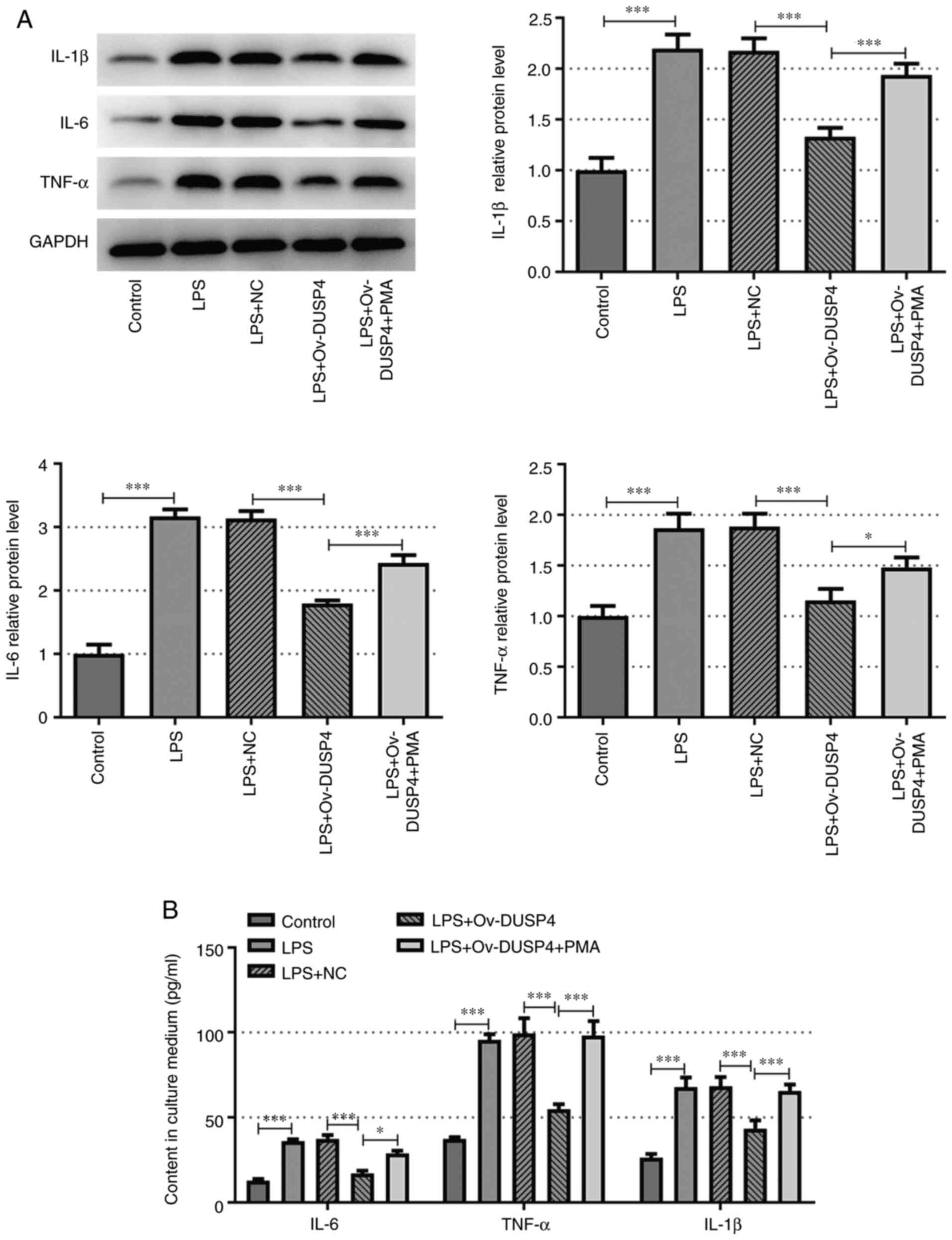
(Fig. 2B-E). Furthermore, the
secretion levels of the inflammation-related factors, IL-6, TNF-α
and IL-1β were measured in each group using ELISA. The results
demonstrated that PMA reversed the inhibitory effect of DUSP4
overexpression on the protein expression levels of IL-6, TNF-α and
IL-1β compared with the LPS + ov-DUSP4 group (Fig. 3A and B).
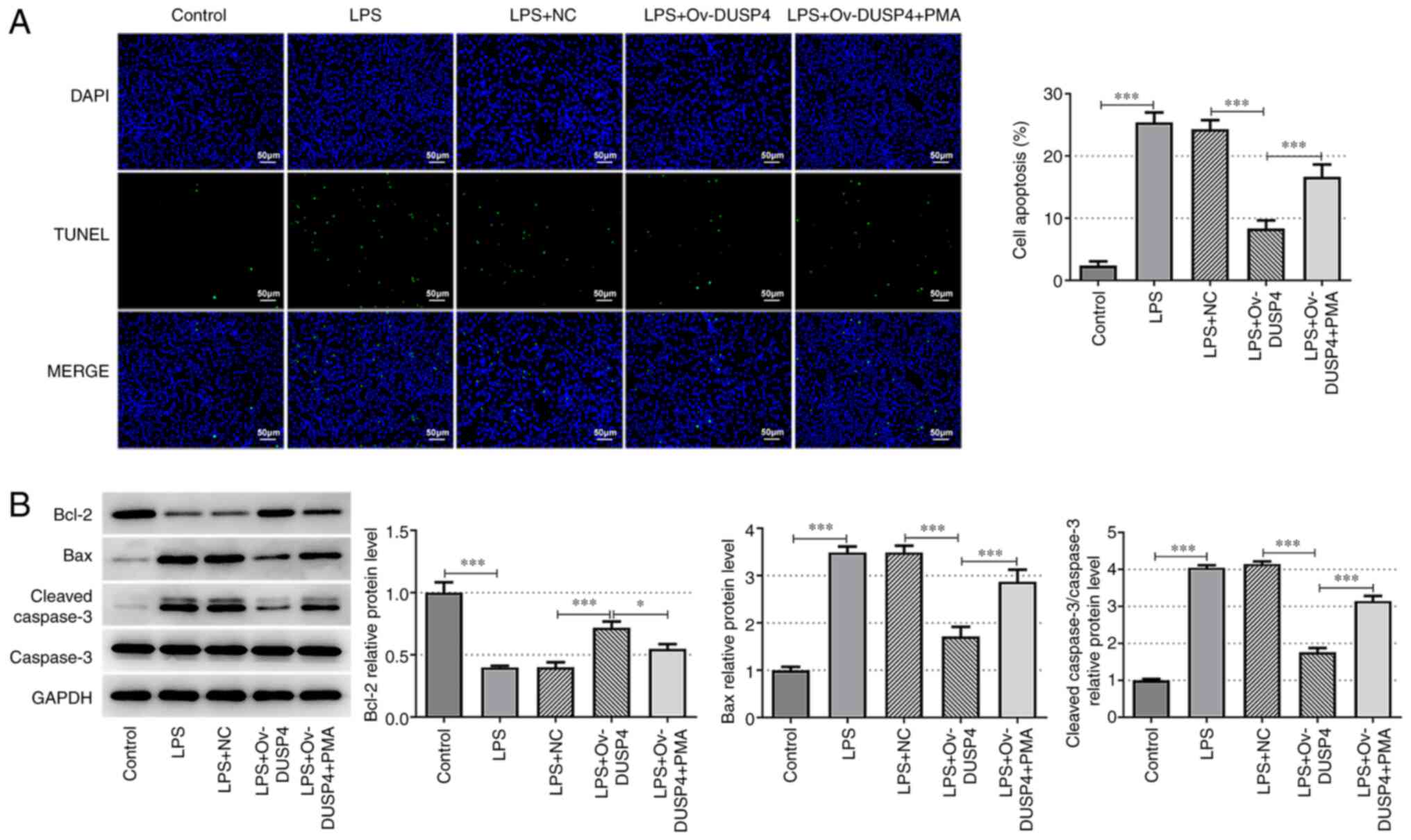
DUSP4 alleviates OA chondrocyte
apoptosis via the MAPK signaling pathway
TUNEL and western blotting assays were carried out
to evaluate cell apoptosis. Compared with the control group, cell
apoptosis was significantly increased in the LPS group, accompanied
by upregulation of Bax and cleaved caspase 3, and the significant
downregulation of Bcl-2. Additionally, apoptosis was reduced in the
LPS + ov-DUSP4 group compared with the LPS + NC group. Consistent
with the aforementioned results, the expression of Bax and cleaved
caspase 3 was decreased, while that of Bcl-2 was increased in the
LPS + ov-DUSP4 group compared with the LPS + NC group. The number
of apoptotic cells and the expression of Bax and cleaved caspase 3
were increased in the LPS + ov-DUSP4 + PMA group compared with the
LPS + ov-DUSP4 group. However, Bcl-2 was downregulated in the LPS +
ov-DUSP4 + PMA group compared with the LPS + ov-DUSP4 group
(Fig. 4A and B). These results indicated that DUSP4
alleviated apoptosis of OA chondrocyte by inhibition of the MAPK
signaling pathway.
Discussion
OA involves a number of complex mechanisms,
including cartilage degradation, synovitis, and destruction and
loss of articular cartilage (14).
Chondrocytes, the only specialized cell type found in cartilage
tissue, serve a critical role in maintaining joint stability
(15). As prochondrocytes, ATDC5
cells differentiate in a similar way to cells that undergo
chondrogenesis (16). ATDC5 cells
are used as in vitro phytopharmacological models to reflect
the complete process of chondrocyte proliferation, differentiation
and mineralization during chondrogenesis (17). At present, the application of ATDC5
cells in OA research has been widely accepted (18-20).
In the present study, ATDC5 cells were induced with LPS to
establish an OA cell model, as previously described (20,21).
In the present study, the mouse OA chondrocyte injury model was
utilized to explore the regulatory role and mechanism underlying
DUSP4 in chondrocyte injury. However, the limitations of the
present study include limited cell line selection, leading to a
lack of analysis in human chondrocytes. Human chondrocytes will be
used for verification in future experiments.
Emerging evidence has demonstrated that DUSP4 serves
a regulatory role in cell inflammation, injury and apoptosis in a
variety of diseases (9,22,23).
However, the exact mechanism underlying DUSP4 in OA is yet to be
elucidated. In the present study, results of the GSE1919 dataset
analysis demonstrated that the expression of DUSP4 was notably
downregulated in OA. In addition, the expression of DUSP4 was also
significantly decreased in LPS-induced ATDC5 cells. DUSP4, also
known as MKP2, serves a key role as a negative regulator of the
MAPK signaling pathway in a number of biological processes
(24). Rapid phosphorylation of
ERK1/2 following LPS induction in microglia induces the expression
of DUSP1, DUSP4, DUSP2 and DUSP5(25). The result demonstrated that DUSPs
exhibit different substrate preferences for specific MAPKs. In the
present study, phosphorylation of p38, ERK1/2 and JNK in the MAPK
signaling pathway resulted in decreased expression of DUSP4
following LPS induction. These results indicated that LPS inhibited
the expression of DUSP4 by promoting p38, ERK1/2 and JNK
phosphorylation. Li et al (11) demonstrated that regulation of DUSP4
expression enhanced the proliferation and apoptosis of OA cells,
thus suggesting that DUSP4 may be involved in the occurrence of OA.
In the present study, DUSP4 overexpression markedly enhanced
LPS-induced ATDC5 cell viability and attenuated oxidative stress,
cell apoptosis and inflammatory response.
DUSP4 dephosphorylates several key signaling
molecules, including those involved in the MAPK signaling pathway
(26). The results of a previous
study indicated that DUSP4 silencing in diabetes enhanced the
activity of the p38/JNK signaling pathway (27). Additionally, simultaneously
activating the MAPK signaling pathway aggravated LPS-induced
chondrocyte injury (28). Ham
et al (25) revealed an
association between LPS, DUSPs and ERK1/2 in microglia cells, and
highlighted that LPS induced ERK1/2 phosphorylation, thereby
regulating the expression of DUSP4 and 6. Inhibition of ERK1/2
phosphorylation also inhibited LPS-induced DUSP4 expression. In
addition, inhibition of DUSP6 increased ERK1/2 phosphorylation
(25). However, the role of DUSP4
in ERK1/2 phosphorylation and regulation of the MAPK signaling
pathway are yet to be fully elucidated. Thus, we hypothesized that
DUSP4 attenuated LPS-induced chondrocyte injury in the KOA model
via inhibition of the MAPK signaling pathway. To further verify
this hypothesis, cells were co-treated with the MAPK signaling
pathway agonist, PMA. The results demonstrated that PMA reversed
the inhibitory effect of DUSP4 overexpression on oxidative stress,
inflammatory response and LPS-induced ATDC5 cell apoptosis. The
aforementioned findings suggested that DUSP4 alleviated LPS-induced
chondrocyte injury in the KOA model via inhibition of the MAPK
signaling pathway.
Further limitations of the present study include the
investigation of DUSP4 expression at a cellular level, as the
mechanisms underlying the effects of DUSP4 on OA were not evaluated
in vivo. Therefore, the effects of DUSP4 on OA must be
further verified using in vivo experiments. In addition, it
is unclear whether DUSP4 plays a role in OA by targeting the MAPK
signaling pathway, or by targeting alternate genes that regulate
the MAPK pathway.
The detection of inflammatory-related factors and
apoptosis-related indicators were the main focus of the present
study. In following experiments, the roles of these factors will be
further investigated.
In conclusion, the present study demonstrated that
DUSP4 alleviated LPS-induced chondrocyte injury in KOA via
inhibition of the MAPK signaling pathway. The present study
provided a theoretical basis for further investigation into the
mechanisms underlying the development of KOA.
Acknowledgements
The authors would like to acknowledge Ganzhou Key
Laboratory of Bone and Joint (Jiangxi, China), which offered help
in the procurement of experimental materials and data collection,
and was used to perform experiments in the present study.
Funding
The present study was supported by the Science and Technology
Plan of Jiangxi Provincial Health Commission (grant no.
20204611).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL wrote the manuscript, analyzed the data,
performed the experiments and supervised the study. BC searched the
literature, revised the manuscript for important intellectual
content, made substantial contributions to the conception and
design of the study, and contributed to analysis and interpretation
of data. ZL and BC confirm the authenticity of all the raw data.
All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kan HS, Chan PK, Chiu KY, Yan CH, Yeung
SS, Ng YL, Shiu KW and Ho T: Non-surgical treatment of knee
osteoarthritis. Hong Kong Med J. 25:127–133. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Mathiessen A and Conaghan PG: Synovitis in
osteoarthritis: Current understanding with therapeutic
implications. Arthritis Res Ther. 19(18)2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Jeffries MA: Osteoarthritis year in review
2018: Genetics and epigenetics. Osteoarthritis Cartilage.
27:371–377. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Barnett R: Osteoarthritis. Lancet.
391(1985)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Low HB and Zhang Y: Regulatory roles of
MAPK phosphatases in cancer. Immune Netw. 16:85–98. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ratsada P, Hijiya N, Hidano S, Tsukamoto
Y, Nakada C, Uchida T, Kobayashi T and Moriyama M: DUSP4 is
involved in the enhanced proliferation and survival of
DUSP4-overexpressing cancer cells. Biochem Biophys Res Commun.
528:586–593. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kang X, Li M, Zhu H, Lu X, Miao J, Du S,
Xia X and Guan W: DUSP4 promotes doxorubicin resistance in gastric
cancer through epithelial-mesenchymal transition. Oncotarget.
8:94028–94039. 2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Barajas-Espinosa A, Basye A, Angelos MG
and Chen CA: Modulation of p38 kinase by DUSP4 is important in
regulating cardiovascular function under oxidative stress. Free
Radic Biol Med. 89:170–181. 2015.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lu Z, Feng H, Shen X, He R, Meng H, Lin W
and Geng Q: MiR-122-5p protects against acute lung injury via
regulation of DUSP4/ERK signaling in pulmonary microvascular
endothelial cells. Life Sci. 256(117851)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Varela-Eirin M, Loureiro J, Fonseca E,
Corrochano S, Caeiro JR, Collado M and Mayan MD: Cartilage
regeneration and ageing: Targeting cellular plasticity in
osteoarthritis. Ageing Res Rev. 42:56–71. 2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Li X, Huang TL, Zhang GD, Jiang JT and Guo
PY: LncRNA ANRIL impacts the progress of osteoarthritis via
regulating proliferation and apoptosis of osteoarthritis
synoviocytes. Eur Rev Med Pharmacol Sci. 23:9729–9737.
2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Giovannini-Chami L, Marcet B, Moreilhon C,
Chevalier B, Illie MI, Lebrigand K, Robbe-Sermesant K, Bourrier T,
Michiels JF, Mari B, et al: Distinct epithelial gene expression
phenotypes in childhood respiratory allergy. Eur Respir J.
39:1197–1205. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Martel-Pelletier J, Barr AJ, Cicuttini FM,
Conaghan PG, Cooper C, Goldring MB, Goldring SR, Jones G, Teichtahl
AJ and Pelletier JP: Osteoarthritis. Nat Rev Dis Primers.
2(16072)2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Charlier E, Deroyer C, Ciregia F, Malaise
O, Neuville S, Plener Z, Malaise M and de Seny D: Chondrocyte
dedifferentiation and osteoarthritis (OA). Biochem Pharmacol.
165:49–65. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ogawa Y, Takahashi N, Takemoto T, Nishiume
T, Suzuki M, Ishiguro N and Kojima T: Hyaluronan promotes
TRPV4-induced chondrogenesis in ATDC5 cells. PLoS One.
14(e0219492)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wilhelm D, Kempf H, Bianchi A and Vincourt
JB: ATDC5 cells as a model of cartilage extracellular matrix
neosynthesis, maturation and assembly. J Proteomics.
219(103718)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhang H, Lin C, Zeng C, Wang Z, Wang H, Lu
J, Liu X, Shao Y, Zhao C, Pan J, et al: Synovial macrophage M1
polarisation exacerbates experimental osteoarthritis partially
through R-spondin-2. Ann Rheum Dis. 77:1524–1534. 2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Fan L, Li M, Cao FY, Zeng ZW, Li XB, Ma C,
Ru JT and Wu XJ: Astragalus polysaccharide ameliorates
lipopolysaccharide-induced cell injury in ATDC5 cells via
miR-92a/KLF4 mediation. Biomed Pharmacother.
118(109180)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Liu M, Zhang J, Liu W and Wang W:
Salidroside protects ATDC5 cells against lipopolysaccharide-induced
injury through up-regulation of microRNA-145 in osteoarthritis. Int
Immunopharmacol. 67:441–448. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Shu Y, Long J, Guo W and Ye W:
MicroRNA-195-5p inhibitor prevents the development of
osteoarthritis by targeting REGγ. Mol Med Rep. 19:4561–4568.
2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Westphal S, Stoppe C, Gruenewald M, Bein
B, Renner J, Cremer J, Coburn M, Schaelte G, Boening A, Niemann B,
et al: Genome-wide association study of myocardial infarction,
atrial fibrillation, acute stroke, acute kidney injury and delirium
after cardiac surgery-a sub-analysis of the RIPHeart-Study. BMC
Cardiovasc Disord. 19(26)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Dougherty JA, Kilbane Myers J, Khan M,
Angelos MG and Chen CA: Dual-specificity phosphatase 4
overexpression in cells prevents hypoxia/reoxygenation-induced
apoptosis via the upregulation of eNOS. Front Cardiovasc Med.
4(22)2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Kirchner A, Bagla S, Dachet F and Loeb JA:
DUSP4 appears to be a highly localized endogenous inhibitor of
epileptic signaling in human neocortex. Neurobiol Dis.
145(105073)2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ham JE, Oh EK, Kim DH and Choi SH:
Differential expression profiles and roles of inducible DUSPs and
ERK1/2-specific constitutive DUSP6 and DUSP7 in microglia. Biochem
Biophys Res Commun. 467:254–260. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chen HF, Chuang HC and Tan TH: Regulation
of dual-specificity phosphatase (DUSP) ubiquitination and protein
stability. Int J Mol Sci. 20(2668)2019.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Denhez B, Rousseau M, Dancosst DA, Lizotte
F, Guay A, Auger-Messier M, Côté AM and Geraldes P:
Diabetes-induced DUSP4 reduction promotes podocyte dysfunction and
progression of diabetic nephropathy. Diabetes. 68:1026–1039.
2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lei J, Fu Y, Zhuang Y and Zhang K: Sema4D
aggravated LPS-induced injury via activation of the MAPK signaling
pathway in ATDC5 chondrocytes. Biomed Res Int.
2020(8691534)2020.PubMed/NCBI View Article : Google Scholar
|