Introduction
Colon cancer (CC) is the third most commonly
diagnosed cancer worldwide and the second most common cause of
cancer-related mortalities worldwide (1). The number of new cases and mortalities
of colorectal cancer in China ranks first in the world with ~2.7
million new cases and over 1.3 million deaths, which is much higher
compared with that in other countries and regions (2). At present, although surgical resection
and chemotherapy, such as 5-fluorouracil, cisplatin and
oxaliplatin, have proven effective for patients with colon cancer,
tumor recurrence and chemoresistance remain a major challenge for
the efficacy colon cancer therapy (3). Previous studies have shown that
autophagy is closely associated with the occurrence and development
of numerous cancer types, including leukemia, renal cell cancer,
non-small cell lung cancer, melanoma and advanced solid tumor;
furthermore, it has also been found to be an important mechanism
underlying resistance to chemotherapy (4,5).
Therefore, exploring novel interventions to target autophagy may
contribute to improving the efficacy of chemotherapy for CC
(6).
MicroRNAs (miRNAs or miRs) are a group of
evolutionarily-conserved small RNAs that serve a negative role in
gene regulation at post-transcriptional levels (7). miRNAs were previously found to be
associated with drug resistance by targeting a wide variety of
tumor-related genes in gliomas, including a number of drug
resistance-related genes, such as EGFR and p53(8). Changes in the expression level of a
single miRNA can simultaneously influence a diverse range complex
molecular pathways (9). Therefore,
the possible mechanism underlying miRNA-mediated drug resistance
may be associated with a complex series of pathological,
physiological and biological processes. A number of miRNAs have
been reported to be involved in the occurrence of drug resistance
in CC, as follows: miR-140 Blocks G1 and G2
phase arrest to sensitize colon cancer stem-like cells to 5-FU
(10), miR-20a targets
BCL2-interacting protein 2 to mediate chemotherapeutic resistance
to fluorouracil, oxaliplatin and teniposide (11), miR-137 sensitizes resistant cells to
oxaliplatin by targeting Y box-binding protein 1(12) and miR-195 sensitizes resistant cells
to doxorubicin by targeting BCL2L2(13). As such, elucidating the functions of
miRNAs in mediating drug resistance may lead to improved
therapeutic strategies for CC therapy.
miR-4486 was revealed to be downregulated in glioma
tissues and to facilitate gliomagenesis in vivo. The
overexpression of miR-4486 was found to attenuate the growth of
glioma cells (14). Additionally,
miR-4486 was significantly downregulated in patients with chronic
obstructive pulmonary disease with eosinophilia and was an
indicator of poor prognosis (15).
Furthermore, increased expression of miR-4486 was found to be
associated with febrile seizures (16). However, to the best of our
knowledge, the expression and effect of miR-4486 in CC have not
been previously reported. Autophagy related 7 (ATG7) was reported
to be a target of miR-106a that suppresses tumor cell death in
colorectal cancer, which was associated with inducing autophagy
(17). Notably, researchers have
found that autophagy is one of the major factors leading to drug
resistance in CC (18), which
indicates that autophagy might be a potential target for
DDP-resistance colon cancer. Based on these, the present study was
designed to investigate the function of miR-4486 on apoptosis and
autophagy in DDP-resistant CC cell lines HCT116/DDP and
SW480/DDP.
Materials and methods
Tissue samples
A total of 40 CC tissues resected between Jan 2014
and Dec 2015 were retrieved from the archives of the Affiliated
Hospital of Guangdong Medical University (Zhanjiang, China).
Thereinto, 22 males and 18 females (1.22:1) were enrolled in this
study, and the median age was 57 years (range, 48-67). Inclusion
criteria were: Patients met NCCN colon cancer tumor clinical
practice guidelines (19). CT,
color Doppler ultrasound and MRI were performed to rule out distant
metastasis. According to TNM staging system there were Stage I, II
and III. Patients did not receive previous chemotherapy or
radiotherapy. Patients were diagnosed for the first time, with
detailed clinicopathological data. All the slides of CC tissues
were re-evaluated as I~III graded CC and independently reviewed by
two pathologists according to WHO classifications (20). Exclusion criteria were: Patients
without other malignant tumors, hematological diseases; patients
with severe complications and immune system diseases; patients with
poor treatment compliance caused by severe mental illness; and
patients unwilling to participate in the present study. The CC
tissues were categorized as favorable prognosis group
(DDP-sensitive; survival >5 years; n=20) or unfavorable
prognosis group (DDP-resistant; survival <1 year; n=20). Based
on the time interval between the last dose of DDP and the diagnosis
of recurrence, patients were classified as either DDP-sensitive or
DDP-resistant cases; >6 months of the time interval indicated
DPP sensitivity, while <6 months of the interval indicated DDP
resistance. The tissues were immediately frozen in liquid nitrogen
and stored at -80˚C until use. The present study was approved by
the local Ethics Committee at The Affiliated Hospital of Guangdong
Medical University and all patients provided written informed
consent.
Cell culture
The human CC cell lines, HCT116 (cat. no. MXC469)
and SW480 (cat. no. MXC368) and the corresponding DDP-resistant
cells, HCT116/DDP (cat. no. MXC795) and SW480/DDP (cat. no.
MXBC241), were purchased from Shanghai Meixuan Biological
Technology Co., Ltd. and cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% fetal calf
serum (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
and 100 U/ml streptomycin with 5% CO2 at 37˚C. For the
maintenance of the DDP-resistant phenotype, HCT116/DDP and
SW480/DDP cells were constantly treated with 5 µg/ml cisplatin
(DDP; Sigma-Aldrich; Merck KGaA).
Cell transfection
HCT116, HCT116/DDP, SW480 and SW480/DDP cells were
cultured in the medium supplied with 10% fetal bovine serum in a
moist, wet atmosphere with 5% CO2 at 37˚C to a
confluence of 70-80% for 24 h. miR-4486 mimic
(5'-ACACTCCAGCTGGGGCTGCGCGA-3'), negative control mimic (NC,
5'-UCACAACCUCCUAGAAAGAGUAGA-3'), pcDNA3.1-ATG7 and cDNA3.1 empty
vector (NC vector) were synthesized by Shanghai GenePharma Co.,
Ltd. Subsequently, they were diluted in OptiMEM (Thermo Fisher
Scientific, Inc.) to 10x their final concentrations and were mixed
with equal volumes of Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. A total of 20 ml of the mixture was added per well of the
96-well plate at room temperature for 10 min, and an 80 µl aliquot
of cells (5x103-1x104 cells/well) was plated
at 37˚C for 4 h. Cell viability was assessed at 96 h after
transfection by adding 80 µl CellTiter-Glo® (Promega
Corporation) per well and incubating the plate at room temperature
for 10 min. Cells were then harvested for subsequent
experiments.
Cell Counting Kit-8 (CCK-8) assay
The four cell lines (2x103 cells/well)
were seeded into 96-well plates and cultured overnight. HCT116,
HCT116/DDP, SW480 and SW480/DDP cells were treated with DDP at a
concentration of 1, 3, 9, 27 and 81 µg/ml at 37˚C for 72 h to
determine the half maximal inhibitory concentration
(IC50). The effect of miR-4486 on the cell viability of
HCT116/DDP and SW480/DDP cells treated with an IC50
concentration of DDP was also determined 1 day after transfection.
Next, cells were treated with DDP before 10 µl CCK-8 reagent
(Beijing Solarbio Science & Technology Co., Ltd.) was added to
each well and cultured further at 37˚C for 2 h for the measurement
of cell viability. The absorbance of each well at 450 nm was
measured by using a microplate reader (Bio-Rad Laboratories, Inc.).
Each concentration was analyzed with six replicates. The cell
viability was calculated as: Cell
viability=[A(experimental)-A(blank)]/[A(control)-A(blank)]. Cell
inhibition=1-cell viability.
Cell apoptosis analysis
HCT116/DDP and SW480/DDP cells (2x105
cells/well) were cultured in six-well plates, before the miR-4486
mimic, NC mimic and pcDNA3.1-ATG7 were transfected into HCT116/DDP
and SW480/DDP cells. After 1 day of transfection, a final
concentration of 5 µg/ml DDP was added to the HCT116/DDP and
SW480/DDP cells to maintain their DDP resistance. At 48 h
post-transfection, the cells were washed with PBS and resuspended
in the binding buffer with the final concentration of
5x104 cells/ml, then double-stained with annexin V-FITC
and PI reagents (Abcam) at room temperature for 15 min in the dark.
Next, after being washed three times with PBS to remove excess
antibodies, the cells were analyzed by flow cytometry using BD
FACSCanto II flow cytometer (BD Biosciences). All flow cytometry
data were analyzed using FlowJo v10.1 software for Macintosh
(FlowJo LLC).
Luciferase assay
The potential target of miR-4486 was predicted using
TargetScan v2.0 (http://www.targetscan.org/). The wild-type (Wt) and
mutant (Mut)-type sequences of ATG7 3'UTR were inserted into the
pSi-Check2 reporter vector. The mutant type 3'UTR sequences of ATG7
contained four nucleotide mutations at the miR-4486 targeting site.
Next, 2x105 cells/well 293T cells (Procell Life Science
& Technology Co., Ltd.) were seeded into 24-well plates and
co-transfected with 5 nM miR-4486 mimic or NC mimic and 160 ng
pSI-Check2-ATG7-3'-UTR-Wt or Mut) using Lipofectamine 2000 reagent.
After transfection for 48 h, the luciferase activity was measured
using a Dual-Luciferase Reporter assay system (Promega Corporation)
according to the manufacturer's protocol. The experiment was
repeated ≥ three times. The ratio of Renilla luciferase to
firefly luciferase was calculated for each well.
Reverse transcription-quantitative PCR
(RT-qPCR)
The expression levels of miR-4486 and ATG7 in
tissues and cells were analyzed using RT-qPCR. Briefly, total RNA
was extracted from HCT116, HCT116/DDP, SW480 and SW480/DDP cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
cDNA was synthesized from total RNA (1.5 µg) using a RevertAid
First Strand cDNA Synthesis kit (Takara Bio, Inc.), and the
synthesized cDNA was amplified at 42˚C for 30 min. The sequence of
the primers for miR-4486 were forward,
5'-ACACTCCAGCTGGGGCTGCGCGA-3' and reverse, 5'-TGGTGTCGTGGAGTCG-3'.
The sequence of the primers for ATG7 were forward,
5'-TGGCTGCTACTTCTGCAATGATGT-3' and reverse,
5'-TTAGCACAGGGAACAGCGCTCATGG-3'. PCR reactions were performed using
the thermocycling conditions of pre-denaturation at 95˚C for 5 min,
followed by and 40 cycles of 95˚C for 15 sec and 60˚C for 1 min and
melt curve analysis. To quantify the expression of miR-4486, cells
and tissues were subjected to a RT-qPCR assay with an AceQ qPCR
SYBR® Green Master Mix kit (Vazyme Biotech Co., Ltd.)
according to the manufacturer's instructions. U6 served as a
control. For the detection of ATG7, RT-qPCR was performed using
SYBR Green PCR Master mix (Thermo Fisher Scientific, Inc.) in an
ABI Step One-Plus Detection system (Applied Biosystems; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocols.
β-actin served as a control. The expression levels of miR-4486 and
ATG7 were quantified using the 2-ΔΔCq method (21).
Western blot analysis
To determine the expression levels of ATG7, Beclin
1, LC3-I, LC3-II, Bax, Bcl-2 and cleaved caspase 3, HCT116/DDP and
SW480/DDP cells were lysed with RIPA buffer (Santa Cruz
Biotechnology, Inc.). The protein lysates were centrifuged at
15,000 x g for 15 mins at 4˚C. The protein concentration was
measured by a Pierce BCA protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Protein samples (50 µg/lane) were separated by
10% SDS-PAGE and were transferred onto polyvinylidene difluoride
membranes. Next, the membranes were blocked with 5% non-fat milk
for 1 h at room temperature and then incubated with the
corresponding primary antibodies, including ATG7 (cat. no. ab52472;
1:1,000; Abcam), Beclin 1 (cat. no. ab210498; 1:1,000; Abcam),
microtubule-associated proteins 1A/1B light chain 3B (LC3-I and
LC3-II; cat. no. L8918, 1:1,000; Sigma-Aldrich; Merck KGaA), Bax
(cat. no. 2772T, 1:1,000; Cell Signaling Technology, Inc.), Bcl-2
(cat. no. ab196495, 1:1,000; Abcam), cleaved caspase 3 (cat. no.
ab32042; 1:500; Abcam) and β-actin (cat. no. ab8227; 1:1,000;
Abcam) at 4˚C overnight followed by incubation with a
HRP-conjugated anti-rabbit secondary antibody (cat. no. ab205718,
1:10,000; Abcam) at room temperature for 1 h. Protein expression
was detected with a high-sensitivity ECL detection kit (Vazyme
Biotech Co., Ltd.). The intensity of the bands was expressed as
fold-change by normalizing the data to the values of β-actin using
the ImageJ software (v1.48; National Institutes of Health).
Transmission electron microscopy (TEM)
analysis for autophagosomes
HCT116/DDP and SW480/DDP cells were fixed in 2.5%
glutaraldehyde (Wuhan Goodbio Technology Co., Ltd.) for 4 h at 4˚C,
rinsed with PBS and then fixed in 1% osmium tetroxide (Sinopharm
Chemical Reagent Co., Ltd.) in 0.1 M PBS for 1 h at room
temperature. Following dehydration with graded ethanol solutions
(70~100%) (Sinopharm Chemical Reagent Co., Ltd.) at room
temperature for 10 min each, the cells were embedded in Epon resin
(Epon 812; Nisshin EM Co. Ltd.), and ultrathin sections (50 nm) of
the selected areas were cut using a LKB NOVA ultramicrotome (LKB
Bromma; GE Healthcare) and a diamond blade (Daito Me Holdings Co.,
Ltd.). Cells were then stained with a saturated solution of uranyl
acetate in methanol (50:50; Sinopharm Chemical Reagent Co., Ltd.)
for 12 min at 45˚C, followed by incubation in an aqueous solution
of concentrated bismuth subnitrate (Sinopharm Chemical Reagent Co.,
Ltd.) for 10 min at 25˚C. Subsequently, all the sections were
observed under a Hitachi TEM system (Hitachi, Ltd.) at a
magnification of x8,000.
Statistical analysis
Each experiment was performed at least three times,
and data are presented as the mean ± standard deviation.
Statistical comparisons between a pair of data were carried out
using an Unpaired student's t-test, whilst comparisons among
multiple groups were performed using a one-way or two-way ANOVA
followed by Tukey's post-hoc tests. GraphPad Prism 7 (GraphPad
Software, Inc.) was used to perform statistical analysis and graph
constructions. P<0.05 was considered to indicate a statistically
significant difference.
Results
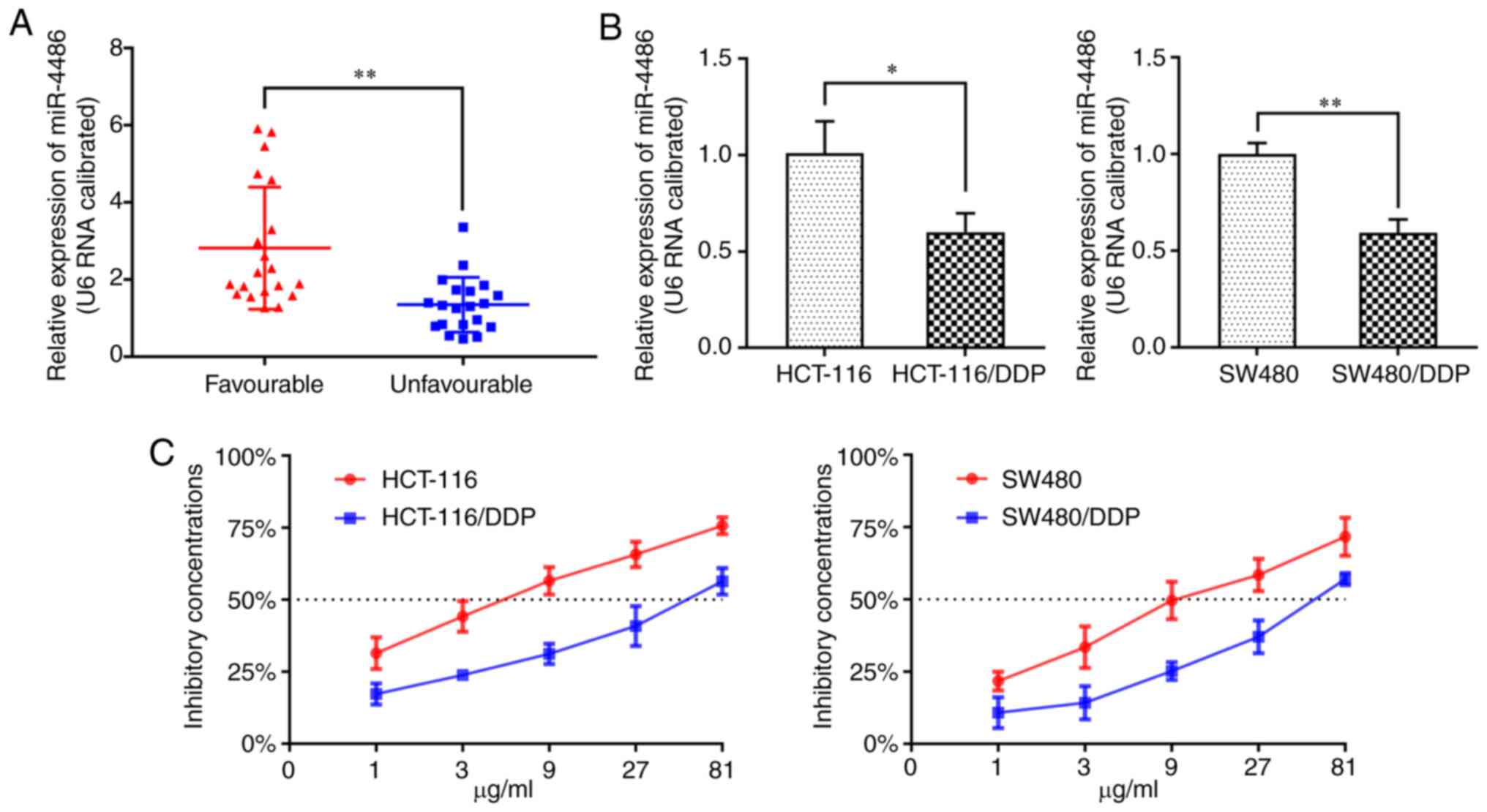
Expression of miR-4486 is decreased in
CC tissues with an unfavorable prognosis and in DDP-resistant CC
cell lines
To investigate the role of miR-4486 in CC, RT-qPCR
was used to quantify the expression of miR-4486 in CC tissues from
patients with an unfavorable prognosis and DDP-resistant CC cell
lines. It was found that miR-4486 was significantly downregulated
in patients with an unfavorable prognosis compared with that in
patients with a favorable prognosis (Fig. 1A). There was also a significant
reduction in miR-4486 expression in HCT116/DDP and SW480/DDP cells
compared with that in their parental HCT116 and SW480 cells
(Fig. 1B). CCK-8 assay revealed
that the IC50 of DDP in HCT116, HCT116/DDP, SW480, and
SW480/DDP cells was 5.5, 52.9, 11.8 and 56.3 µg/ml, respectively
(Fig. 1C). These results suggest
that miR-4486 expression is lower in tissues from patients with CC
with unfavorable outcomes and in DDP-resistant CC cell lines
compared with that in the tissues from patients with CC with
favorable outcomes and parental CC cell lines, respectively.
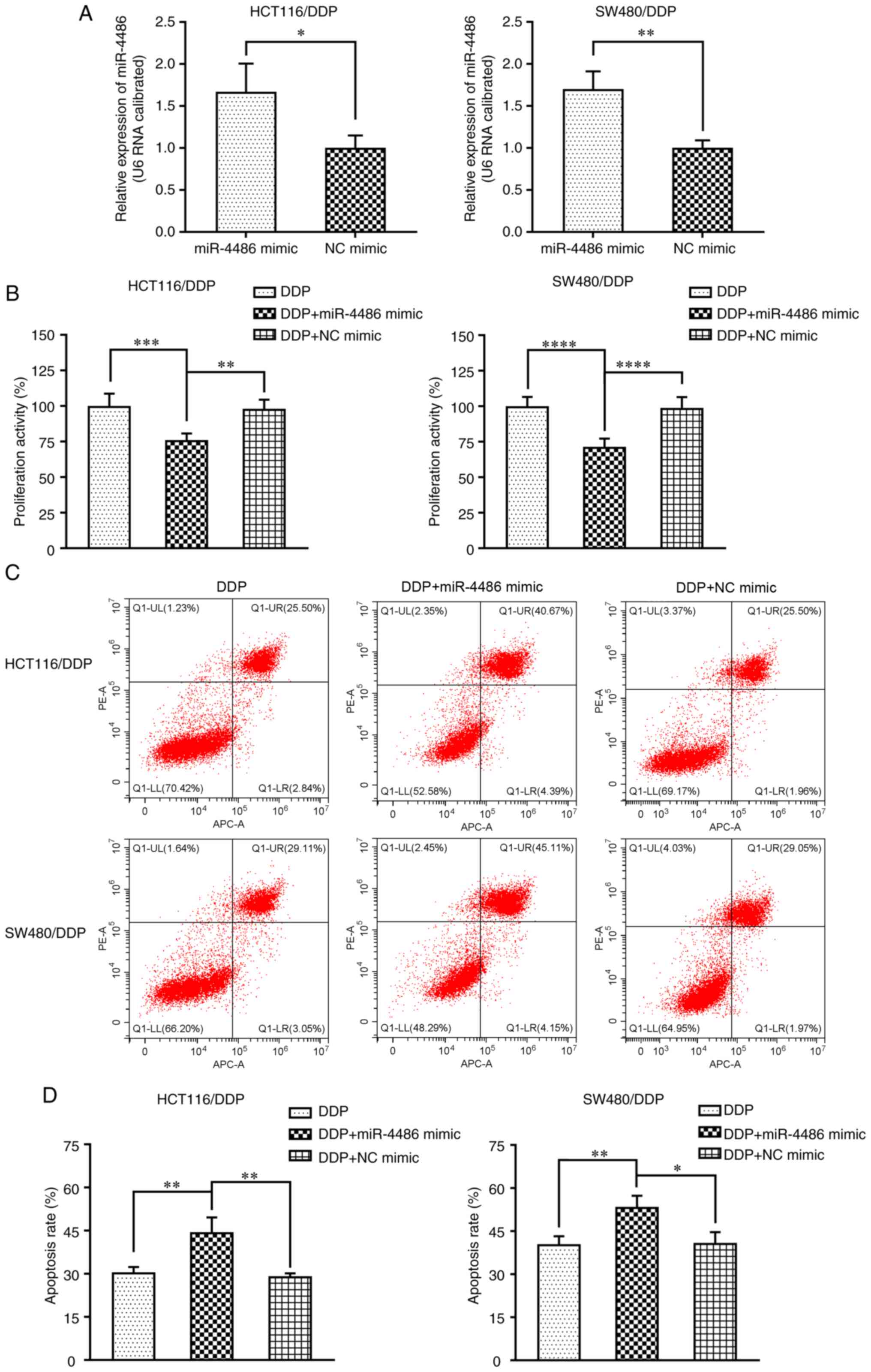
Overexpression of miR-4486 reverse DDP
resistance in HCT116/DDP and SW480/DDP cells
To determine the effect of miR-4486 overexpression
on DDP resistance in HCT116/DDP and SW480/DDP cells, miR-4486 and
NC mimics were synthesized. Firstly, the miR-4486 mimic or NC mimic
were transfected into both cell lines before the expression level
of miR-4486 was determined by RT-qPCR. The results indicated that
miR-4486 expression was significantly promoted by the miR-4486
mimic (Fig. 2A). Subsequently,
HCT116/DDP and SW480/DDP cells were transfected with the miR-4486
mimic or NC mimic and then treated with 5 and 11 µg/ml DDP,
respectively, following which cell viability was subsequently
detected using CCK-8 assay. The results showed that the
overexpression of miR-4486 resulted in a significantly decreased
cell viability compared with that in the DDP-only group (Fig. 2B). Next, the effect of miR-4486 on
HCT116/DDP and SW480/DDP cell apoptosis was analyzed by flow
cytometry (Fig. 2C). The results
showed that the apoptosis rates of HCT116/DDP and SW480/DDP cells
were significantly increased in the miR-4486 mimic group compared
with those in the DDP-only group, suggesting that the
overexpression of miR-4486 can promote the apoptosis of HCT116/DDP
and SW480/DDP cells (Fig. 2D).
These data demonstrated that the upregulation of miR-4486 can
enhance the sensitivity of HCT116/DDP and SW480/DDP cells to
DDP.
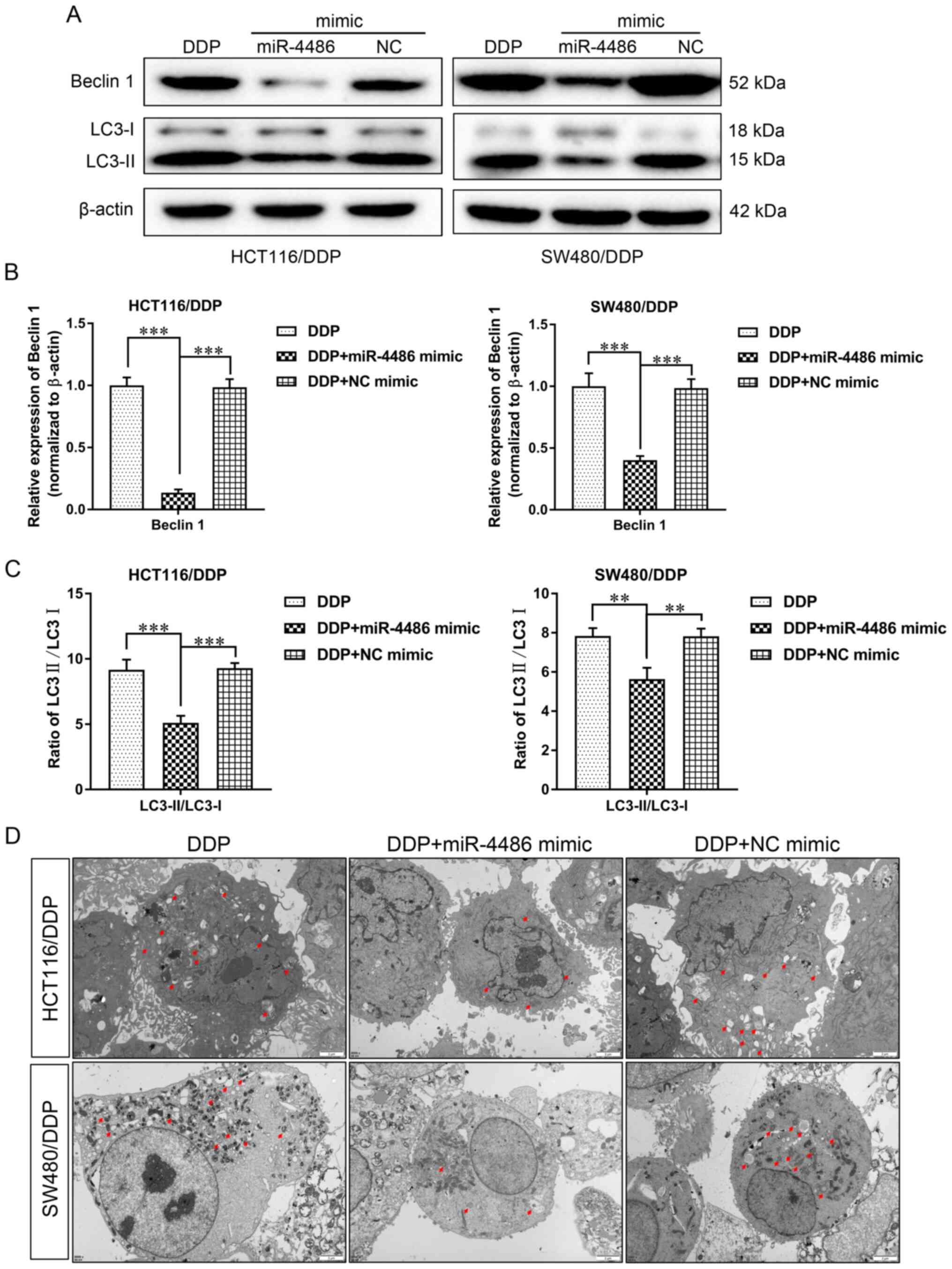
miR-4486 inhibits autophagy in
HCT116/DDP and SW480/DDP cells
Western blotting was subsequently performed to
assess the effect of miR-4486 on autophagy in HCT116/DDP and
SW480/DDP cells. It was found that the expression levels of Beclin
1 and the ratio of LC3-II/LC3-I were significantly decreased in the
miR-4486 mimic group compared with those in the DDP group (Fig. 3A-C), suggesting that the
overexpression of miR-4486 can inhibit the occurrence of autophagy
in HCT116/DDP and SW480/DDP cells. Furthermore, the number of
autophagosomes was also reduced in the miR-4486 mimic group, which
was in accordance with the expression levels of autophagy-related
proteins (Fig. 3D). These
aforementioned results suggest that miR-4486 can increase the DDP
sensitivity of HCT116/DDP and SW480/DDP cells by preventing
autophagy.
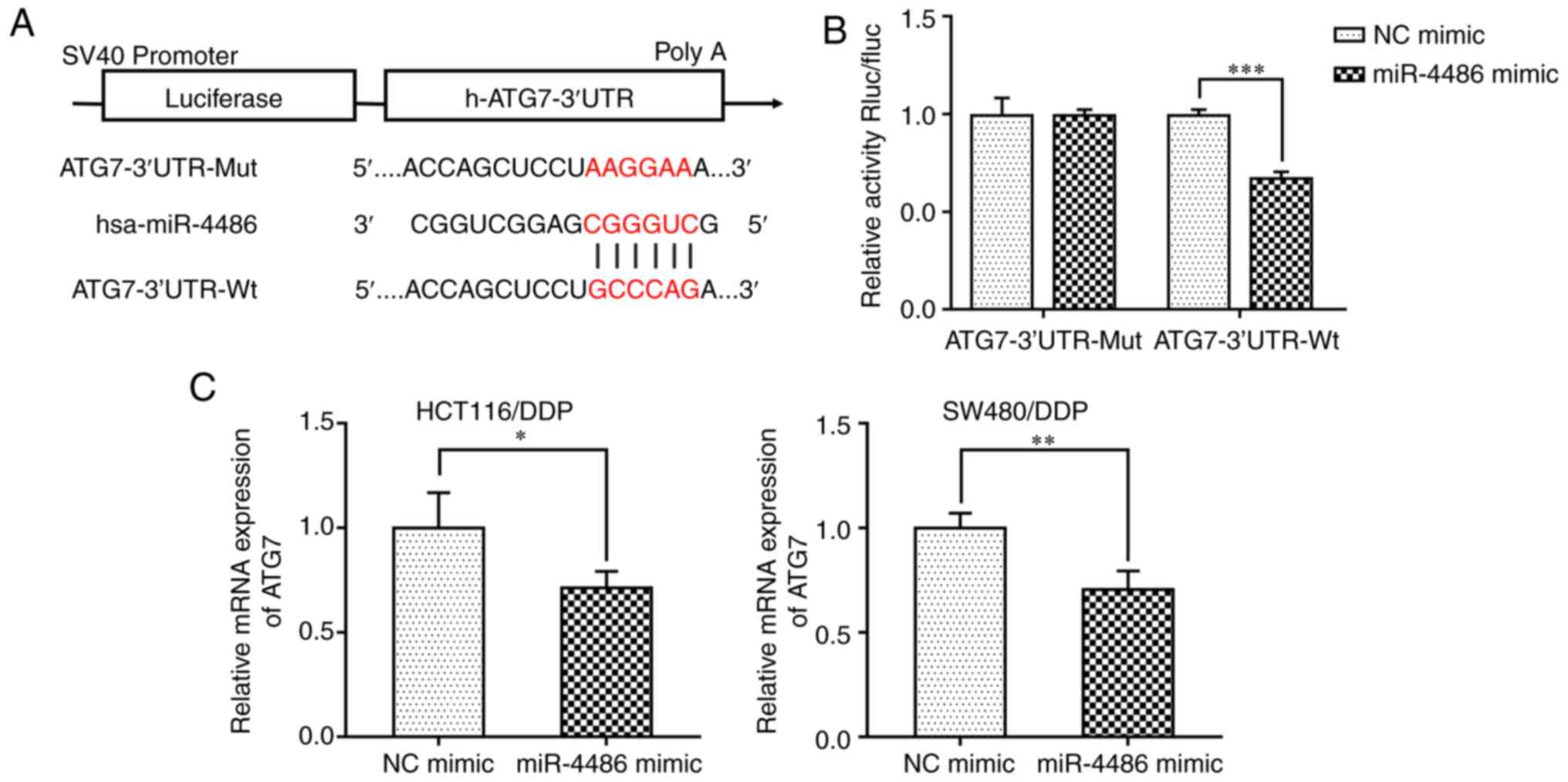
ATG7 is the direct target gene of
miR-4486
Using the online tool TargetScan, the present study
predicted that miR-4486 may directly target the 3'-UTR of ATG7 mRNA
(Fig. 4A). To verify this, Wt or
Mut firefly luciferase reporters containing the 3'-UTR of ATG7 were
constructed and luciferase assay was conducted. The results
revealed that the miR-4486 mimic significantly decreased the
luciferase activity of cells transfected with the ATG7 3'-UTR-Wt
plasmid compared with that in the miR-4486 NC group but not in
cells in the ATG7 3'-UTR-Mut group (Fig. 4B). This suggests that miR-4486 may
directly target ATG7 to reduce the expression of ATG7 by binding to
its 3'-UTR. To verify further if the expression of ATG7 was
regulated by miR-4486, the miR-4486 mimic or NC were transfected
into HCT116/DDP and SW480/DDP cells, where RT-qPCR was conducted to
detect the ATG7 expression levels. The data demonstrated that
transfection with the miR-4486 mimic significantly reduced the ATG7
expression levels in HCT116/DDP and SW480/DDP cells compared with
that in the NC mimic group (Fig.
4C). These results suggest that ATG7 is a target gene of
miR-4486.
Overexpression of ATG7 reverses the
effect of miR-4486 in HCT116/DDP and SW480/DDP cells
To further explore the role of ATG7, ATG7 was cloned
into the pcDNA3.1 vector. HCT116/DDP and SW480/DDP cells were then
transfected with pcDNA3.1-ATG7, where the RT-qPCR and WB results
demonstrated that the mRNA and protein expression levels of ATG7
were both significantly higher in the pcDNA3.1-ATG7 group compared
with those in the NC vector group (Fig.
5A and B).
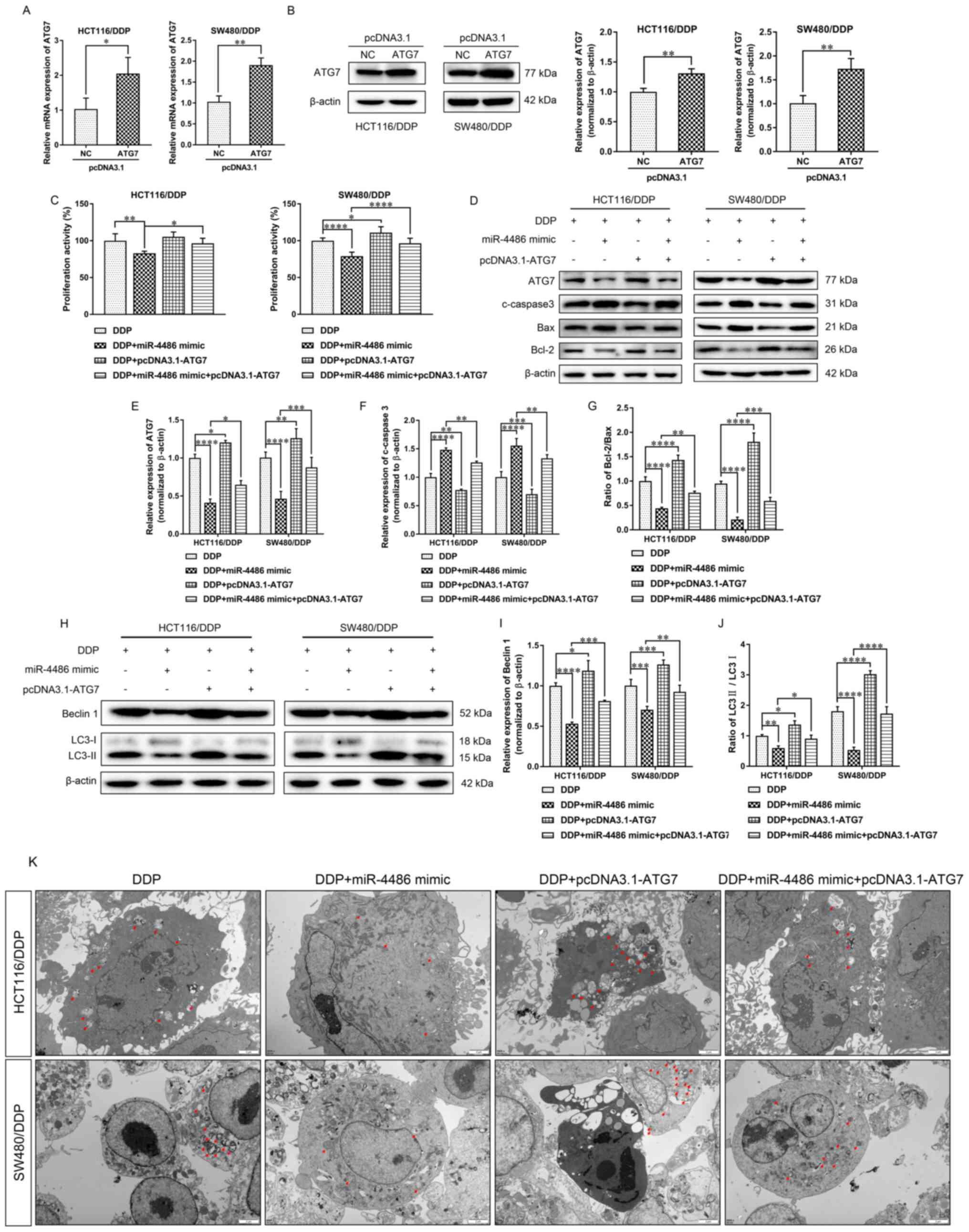 | Figure 5Overexpression of ATG7 reverses the
effect of miR-4486 in HCT116/DDP and SW480/DDP cells. (A) mRNA
expression of ATG7 was analyzed by reverse
transcription-quantitative PCR. (B) The protein expression of ATG7
was measured and quantified by western blotting. (C) The viability
of HCT116/DDP and SW480/DDP cells, which were transfected with
miR-4486, pcDNA3.1-ATG7 or miR-4486 + pcDNA3.1-ATG7, was measured
by Cell Counting Kit-8 assay. (D) The protein expression of ATG7
and the apoptosis-related proteins cleaved caspase 3, Bax and Bcl-2
in each group were measured by western blotting. (E) The levels of
ATG7 protein expression were quantified. (F) The levels of cleaved
caspase 3 protein were quantified. (G) The ratio of Bcl-2/Bax was
calculated. (H) The protein expression of the autophagy-related
proteins Beclin 1, LC3-I and LC3-II were analyzed by western
blotting. (I) The expression of Beclin 1 was quantified. (J) The
ratio of LC3-II/LC3-I was represented. DDP group served as a
control. (K) Autophagosomes in each group were observed by
transmission electron microscopy at x8,000 magnification (scale
bar, 2 µm). Red arrows indicate the autophagosomes. Bars represent
the mean ± standard deviation from ≥3 independent experiments.
*P<0.05, **P<0.01,
***P<0.001 and ****P<0.0001. ATG7,
autophagy-related gene 7; miR, microRNA; DDP, cisplatin;
autophagy-related gene 7; LC3, microtubule-associated proteins
1A/1B light chain 3B. |
Next, the miR-4486 mimic, pcDNA3.1-ATG7 or miR-4486
mimic + pcDNA3.1-ATG7 were transfected into HCT116/DDP and
SW480/DDP cells treated with 5 and 11 µg/ml DDP, respectively,
before cell viability was detected by CCK-8 assay. The results
showed that the overexpression of miR-4486 resulted in
significantly reduced viability compared with that in the DDP group
(Fig. 5C). By contrast, ATG7
overexpression partially but significantly increased the viability
of miR-4486-overexpressing HCT116/DDP and SW480/DDP cells (Fig. 5C). Next, the expression levels of
ATG7 and apoptosis-related proteins were analyzed by western
blotting (Fig. 5D). Compared with
that in the DDP group, the expression level of ATG7 was
significantly decreased in the miR-4486 mimic group but
significantly upregulated in the pcDNA3.1-ATG7 group (Fig. 5D). However, the trend in the
expression of the pro-apoptotic protein cleaved caspase 3 was
opposite to that found in ATG7, where the effects of miR-4486 mimic
were significantly reversed by ATG7 overexpression (Fig. 5E and F). The trend of the Bcl-2/Bax ratio was in
accordance with the trend of ATG7, where the ratio of Bcl-2/Bax was
significantly decreased and increased by the miR-4486 mimic and
pcDNA3.1-ATG7, respectively (Fig.
5G). The inhibitory effects of the miR-4486 mimic on the ratio
of Bcl-2/Bax could be significantly reversed by ATG7 (Fig. 5G).
The expression of the autophagy-related proteins
Beclin 1, LC3-I and LC3-II are shown in Fig. 5H. The levels of Beclin 1 and the
LC3-II/LC3-I ratio in DPP-treated DPP-resistant cells were both
significantly downregulated by the miR-4486 mimic but upregulated
by pcDNA3.1-ATG7 transfection, whilst the overexpression of ATG7
could significantly reverse the inhibitory effects of the miR-4486
mimic on autophagy (Fig. 5I and
J). Additionally, compared with
that in the DDP group, the number of autophagosomes was reduced or
increased after miR-4486 mimic or pcDNA3.1-ATG7 transfection,
respectively. The number of autophagosomes was accordingly
increased after pcDNA3.1-ATG7 transfection in the miR-4486 group
(Fig. 5K). These data suggest that
the overexpression of miR-4486 can reduce the viability of
HCT116/DDP and SW480/DDP cells by reversing their DDP resistance.
The role of miR-4486, which was found to promote apoptosis whilst
inhibiting autophagy, may be associated with the inhibition of ATG7
expression.
Discussion
The present study showed that the expression of
miR-4486 was downregulated in DDP-resistant HCT116 and SW480 cells
(HCT116/DDP and SW480/DDP) compared with that in their parental
cell lines, HCT116 and SW480. In addition, the overexpression of
miR-4486 could enhance the sensitivity of HCT116/DDP and SW480/DDP
cells to DDP by inhibiting autophagy. Since ATG7 was reported to be
an autophagy mediator (17), it was
found that ATG7 was a regulatory target of miR-4486, where the
promotion of apoptosis and inhibition of autophagy mediated by
miR-4486 overexpression could be reversed by ATG7. The present
study may provide novel insights into the role of miR-4486 in the
autophagy-mediated DDP resistance in CC.
A number of miRNAs have been reported to be involved
in regulating DDP-resistant tumor cells, such as osteosarcoma cells
(22) and small-cell lung cancer
cells (23). Some miRNAs reduced
DDP resistance, whilst others can enhance DDP resistance. miR-133b
was found to reduce DDP resistance, where its overexpression
contributed to the suppression of the malignant proliferation and
aggressiveness of DDP-resistant non-small cell lung cancer cells by
targeting glutathione S-transferase π1(24). By contrast, cancer-associated
fibroblasts-derived exosomal miR-196a conferred DDP resistance in
head and neck cancer by targeting cyclin dependent kinase inhibitor
1B and inhibitor of growth family member 5(25). The present study found that miR-4486
could reverse DDP resistance in HCT116/DDP and SW480/DDP cells by
inhibiting cell viability and autophagy. Therefore, the underlying
mechanism was explored further.
The chemotherapeutic agent DDP is a small-molecule
platinum-containing compound that was originally found to inhibit
bacterial growth (26). DDP was
later reported to exert anticancer activity in a variety of tumors,
including tumors of the ovaries and testes, breast, ovarian,
testicular and lung (27,28). It has also been reported that DDP
inhibits tumor growth by inducing apoptosis; for example in the
ovaries, testes and head and neck tumors (29). DDP remained the primary therapeutic
option for several types of solid tumors, such as ovarian cancer,
osteosarcoma and gastric cancer, despite its toxicity even at low
doses, particularly in the kidneys and ear (30). CC is intrinsically resistant to DDP
(31). However, the specific
mechanism remains unclear. It has been reported that DDP causes
ovarian cancer cell death by inducing apoptosis (29), such that a defect in apoptotic
signaling could also confer DDP resistance. DDP-induced genotoxic
stress activates multiple signal transduction pathways including
those involving ATR, p53, p73 and MAPK, which can contribute to
apoptosis or DDP resistance. The present study found that the
expression of miR-4486 in HCT116/DDP and SW480/DDP cells was lower
compared with that in their parental HCT116 and SW480 cells, whilst
the overexpression of miR-4486 could markedly promote the apoptosis
rates of HCT116/DDP and SW480/DDP cells after DDP treatment. These
results suggest that the reduced expression of miR-4486 in
HCT116/DDP and SW480/DDP cells may be involved in DDP
resistance.
Autophagy, as a survival pathway that responds to
metabolic stress, has been reported to mediate the acquired
resistance phenotype of non-small-cell lung and breast cancer
during chemotherapy (5,32). Autophagy can regulate apoptosis,
including mitophagy (33),
microautophagy, chaperone-mediated autophagy (34,35)
and the interaction between Beclin 1 and antiapoptotic Bcl-2 family
members (36,37). When autophagosomes fuse with
lysosomes to form autolysosomes, intra-autophagosomal LC3-II
degrades their contents (38).
Therefore, LC3 could serve as a marker of autophagy. In addition,
LC3 has also been reported to serve a pivotal role in the
interaction between autophagy and apoptosis (39,40).
Beclin 1 regulates every major step in the autophagic pathway, from
autophagosome formation to the maturation of
autophagosomes/endosomes (41). It
was previously found that the antitumor drug, cetuximab, could
induce tumor cell apoptosis through the Beclin 1/LC3 autophagy
pathway in CC (42). However, when
tumor cells continued to divide and proliferate, but the
surrounding environment cannot provide sufficient energy for
maintaining the high levels metabolism, autophagy in tumor cells
would be upregulated (43,44). This is to adapt to any unfavorable
conditions, including nutritional deficiencies, which can result in
chemotherapeutic drug resistance and apoptosis suppression to
increase the survival of tumor cells (43,44).
Protective autophagy is one of the main methods used by tumor cells
to reduce the sensitivity to drugs, where the specific mechanism
has been reported to involve the PI3K/AKT/mTOR and reactive oxygen
species signaling pathways for resistance to erlotinib and
gefitinib in non-small cell lung cancer (45) and to temozolomide in malignant
glioma (46). A previous study has
demonstrated that PI3K/AKT/mTOR signaling-mediated autophagy could
lead to DDP resistance in CC (47).
Therefore, inhibition of autophagy is an important therapeutic
target for relieving DDP resistance in CC.
miRNAs that can regulate autophagy in
multidrug-resistant CC cells have been found. For example,
miR-409-3p was capable of enhancing the chemosensitivity of CC
cells to oxaliplatin by inhibiting Beclin 1-mediated autophagy
(48). By contrast, miR-153-5p
promoted the sensitivity of CC cells to oxaliplatin by targeting
the Bcl-2-mediated autophagy pathway (49). Therefore, the present study explored
the role of miR-4486 on autophagy-mediated DDP resistance in CC
cells. Notably, it was found that miR-4486 could downregulate the
expression of Beclin 1, the ratio of LC3-II/I and the formation of
autophagosomes to prevent autophagy in DPP-treated HCT116/DDP and
SW480/DDP cells. This suggests that suppression of autophagy is
part of the underlying mechanism in which miR-4486 can promote
DDP-induced apoptosis in CC cells.
The present study demonstrated that the
autophagy-related protein ATG7 was a direct inhibitory target of
miR-4486 in autophagy-mediated DDP resistance in CC. A previous
study observed the upregulation of ATG7 in DDP-resistant CC cells
compared with sensitive CC cell lines, which participated in the
autophagic survival and proliferation of CC cells (50). ATG7 was observed to be a key
component of the autophagy machinery, with the main function of
mediating the lipidation of the LC3/GABA type A receptor-associated
protein during autophagosome formation (51). Furthermore, ATG7 was found to
aggravate cell death in CC cell lines HCT116 and SW620, which could
be inhibited by miR-106a (17).
Another previous study showed that ATG7 is essential for autophagy
activation, which significantly promotes dormant breast cancer cell
survival and metastatic burden in vitro and in vivo
(52). Accumulating evidence has
reported the connection between DDP resistance during chemotherapy
with dysregulations in miRNA expression (17,53).
However, the effect and mechanism of miR-4486 on DDP resistance in
CC remain unclear. In the present study, the possible association
between miR-4486 and ATG7 was first identified, which was verified
using dual-luciferase reporter assay. Furthermore, it was found
that ATG7 overexpression could reverse the inhibitory effects of
miR-4486 on DDP resistance in HCT116/DDP and SW480/DDP cells, which
in turn restored cell proliferation and autophagy. These data
suggest further that miR-4486 can reduce DDP resistance in
HCT116/DDP and SW480/DDP cells by targeting ATG7 to inhibit
autophagy.
It should be noted the exact mechanism underlying
these functions remain to be fully elucidated, since miR-4486
inhibitors and autophagy inhibitors were not used in the present
study. The results of this study offered notable insight into the
role of miR-4486, inhibitors of both miR-4486 and autophagy would
need to be used to obtain a comprehensive conclusion.
In conclusion, results from the present study showed
that overexpression of miR-4486 can decrease DDP resistance in
HCT116/DDP and SW480/DDP cells by targeting ATG7 to inhibit
viability and autophagy. Importantly, these findings suggested that
miR-4486 may serve as a new therapeutic target in DDP-resistant
CC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
QX conceived and designed the study. WW, LC and WZ
performed the experiments. XH, LL, ZQ and KS were responsible for
the collection, analysis and interpretation of the data. QX and LC
confirm the authenticity of all the raw data. QX revised the
manuscript critically for important intellectual content. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the local Ethics
Committee at The Affiliated Hospital of Guangdong Medical
University (Zhanjiang, China) and all patients provided written
informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Liu N, Wu C, Jia R, Cai G, Wang Y, Zhou L,
Ji Q, Sui H, Zeng P, Xiao H, et al: Traditional Chinese medicine
combined with chemotherapy and cetuximab or bevacizumab for
metastatic colorectal cancer: A randomized, double-blind,
placebo-controlled clinical trial. Front Pharmacol.
11(478)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Paldino E, Tesori V, Casalbore P,
Gasbarrini A and Puglisi MA: Tumor initiating cells and
chemoresistance: Which is the best strategy to target colon cancer
stem cells? Biomed Res Int. 2014(859871)2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Duffy A, Le J, Sausville E and Emadi A:
Autophagy modulation: A target for cancer treatment development.
Cancer Chemother Pharmacol. 75:439–447. 2015.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Sui X, Chen R, Wang Z, Huang Z, Kong N,
Zhang M, Han W, Lou F, Yang J, Zhang Q, et al: Autophagy and
chemotherapy resistance: A promising therapeutic target for cancer
treatment. Cell Death Dis. 4(e838)2013.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Devenport SN and Shah YM: Functions and
implications of autophagy in colon cancer. Cells.
8(1349)2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Di Leva G and Croce CM: miRNA profiling of
cancer. Curr Opin Genet Dev. 23:3–11. 2013.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Aguda BD: Modeling microRNA-transcription
factor networks in cancer. Adv Exp Med Biol. 774:149–167.
2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Baer C, Claus R and Plass C: Genome-wide
epigenetic regulation of miRNAs in cancer. Cancer Res. 73:473–477.
2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Song B, Wang Y, Xi Y, Kudo K, Bruheim S,
Botchkina GI, Gavin E, Wan Y, Formentini A, Kornmann M, et al:
Mechanism of chemoresistance mediated by miR-140 in human
osteosarcoma and colon cancer cells. Oncogene. 28:4065–4074.
2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chai H, Liu M, Tian R, Li X and Tang H:
miR-20a targets BNIP2 and contributes chemotherapeutic resistance
in colorectal adenocarcinoma SW480 and SW620 cell lines. Acta
Biochim Biophys Sin (Shanghai). 43:217–225. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Guo Y, Pang Y, Gao X, Zhao M, Zhang X,
Zhang H, Xuan B and Wang Y: MicroRNA-137 chemosensitizes colon
cancer cells to the chemotherapeutic drug oxaliplatin (OXA) by
targeting YBX1. Cancer Biomarkers. 18:1–9. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Qu J, Zhao L, Zhang P, Wang J, Xu N, Mi W,
Jiang X, Zhang C and Qu J: MicroRNA-195 chemosensitizes colon
cancer cells to the chemotherapeutic drug doxorubicin by targeting
the first binding site of BCL2L2 mRNA. J Cell Physiol. 230:535–545.
2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu J, Cheng LG and Li HG: LncRNA SNHG20
promoted the proliferation of glioma cells via sponging miR-4486 to
regulate the MDM2-p53 pathway. Eur Rev Med Pharmacol Sci.
23:5323–5331. 2019.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Asensio VJ, Tomás A, Iglesias A, de Llano
LP, Del Pozo V and Cosío BG: CHACOS study group. Eosinophilic COPD
patients display a distinctive serum mirna profile from asthma and
non-eosinophilic COPD. Arch Bronconeumol (Engl ED). 56:234–241.
2020.PubMed/NCBI View Article : Google Scholar : (In English,
Spanish).
|
|
16
|
Kim SH, Yun SW, Kim HR and Chae SA:
Exosomal microRNA expression profiles of cerebrospinal fluid in
febrile seizure patients. Seizure. 81:47–52. 2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hao H, Xia G, Wang C, Zhong F, Liu L and
Zhang D: miR-106a suppresses tumor cells death in colorectal cancer
through targeting ATG7. Med Mol Morphol. 50:76–85. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Xie Q, Liu Y and Li X: The interaction
mechanism between autophagy and apoptosis in colon cancer. Transl
Oncol. 13(100871)2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Benson AB III, Venook AP, Cederquist L,
Chan E, Chen YJ, Cooper HS, Deming D, Engstrom PF, Enzinger PC,
Fichera A, et al: Colon cancer, version 1.2017, NCCN clinical
practice guidelines in oncology. J Natl Compr Canc Netw.
15:370–398. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Bosman FT, Carneiro F, Hruban RH and
Theise ND (eds.): WHO Classification of Tumors of the Digestive
System. Vol 3. 4th edition. IARC Press, Lyon, 2010.
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Song L, Zhou Z, Gan Y, Li P, Xu Y, Zhang
Z, Luo F, Xu J, Zhou Q and Dai F: Long noncoding RNA OIP5-AS1
causes cisplatin resistance in osteosarcoma through inducing the
LPAATbeta/PI3K/AKT/mTOR signaling pathway by sponging the
miR-340-5p. J Cell Biochem. 120:9656–9666. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Pan B, Chen Y, Song H, Xu Y, Wang R and
Chen L: Mir-24-3p downregulation contributes to VP16-DDP resistance
in small-cell lung cancer by targeting ATG4A. Oncotarget.
6:317–331. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lin C, Xie L, Lu Y, Hu Z and Chang J:
miR-133b reverses cisplatin resistance by targeting GSTP1 in
cisplatin-resistant lung cancer cells. Int J Mol Med. 41:2050–2058.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Qin X, Guo H, Wang X, Zhu X, Yan M, Wang
X, Xu Q, Shi J, Lu E, Chen W and Zhang J: Exosomal miR-196a derived
from cancer-associated fibroblasts confers cisplatin resistance in
head and neck cancer through targeting CDKN1B and ING5. Genome
Biol. 20(12)2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Chan H, Pearson CS, Green CM, Li Z, Zhang
J, Belfort G, Shekhtman A, Li H and Belfort M: Exploring intein
inhibition by platinum compounds as an antimicrobial strategy. J
Biol Chem. 291:22661–22670. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Dasari S and Tchounwou PB: Cisplatin in
cancer therapy: Molecular mechanisms of action. Eur J Pharmacol.
740:364–378. 2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Sun CY, Zhang QY, Zheng GJ and Feng B:
Phytochemicals: Current strategy to sensitize cancer cells to
cisplatin. Biomed Pharmacother. 110:518–527. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Siddik ZH: Cisplatin: Mode of cytotoxic
action and molecular basis of resistance. Oncogene. 22:7265–7279.
2003.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Oun R, Moussa YE and Wheate NJ:
Correction: The side effects of platinum-based chemotherapy drugs:
A review for chemists. Dalton Trans. 47(7848)2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Köberle B, Tomicic MT, Usanova S and Kaina
B: Cisplatin resistance: Preclinical findings and clinical
implications. Biochim Biophys Acta. 1806:172–182. 2010.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wu WK, Coffelt SB, Cho CH, Wang XJ, Lee
CW, Chan FK, Yu J and Sung JJ: The autophagic paradox in cancer
therapy. Oncogene. 31:939–953. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang K: Autophagy and apoptosis in liver
injury. Cell Cycle. 14:1631–1642. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Bursch W: The autophagosomal-lysosomal
compartment in programmed cell death. Cell Death Differ. 8:569–581.
2001.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Shimizu S, Kanaseki T, Mizushima N, Mizuta
T, Arakawa-Kobayashi S, Thompson CB and Tsujimoto Y: Role of Bcl-2
family proteins in a non-apoptotic programmed cell death dependent
on autophagy genes. Nat Cell Biol. 6:1221–1228. 2004.PubMed/NCBI View Article : Google Scholar
|
|
36
|
He C and Levine B: The beclin 1
interactome. Curr Opin Cell Biol. 22:140–149. 2010.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Itakura E, Kishi C, Inoue K and Mizushima
N: Beclin 1 forms two distinct phosphatidylinositol 3-kinase
complexes with mammalian Atg14 and UVRAG. Mol Biol Cell.
19:5360–5372. 2008.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Deretic V: Autophagosome and phagosome.
Methods Mol Biol. 445:1–10. 2008.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Fu Ll, Cheng Y and Liu B: Beclin-1:
Autophagic regulator and therapeutic target in cancer. Int J
Biochem Cell Biol. 45:921–924. 2013.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Noguchi M, Hirata N, Tanaka T, Suizu F,
Nakajima H and Chiorini JA: Autophagy as a modulator of cell death
machinery. Cell Death Dis. 11(517)2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Guo GF, Wang YX, Zhang YJ, Chen XX, Lu JB,
Wang HH, Jiang C, Qiu HQ and Xia LP: Predictive and prognostic
implications of 4E-BP1, Beclin-1, and LC3 for cetuximab treatment
combined with chemotherapy in advanced colorectal cancer with
wild-type KRAS: Analysis from real-world data. World J
Gastroenterol. 25:1840–1853. 2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Guo JY, Xia B and White E:
Autophagy-mediated tumor promotion. Cell. 155:1216–1219.
2013.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Bellodi C, Lidonnici MR, Hamilton A,
Helgason GV, Soliera AR, Ronchetti M, Galavotti S, Young KW, Selmi
T, Yacobi R, et al: Targeting autophagy potentiates tyrosine kinase
inhibitor-induced cell death in Philadelphia chromosome-positive
cells, including primary CML stem cells. J Clin Invest.
119:1109–1123. 2009.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Li H, Jin X, Zhang Z, Xing Y and Kong X:
Inhibition of autophagy enhances apoptosis induced by the
PI3K/AKT/mTor inhibitor NVP-BEZ235 in renal cell carcinoma cells.
Cell Biochem Funct. 31:427–433. 2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Lin CJ, Lee CC, Shih YL, Lin TY, Wang SH,
Lin YF and Shih CM: Resveratrol enhances the therapeutic effect of
temozolomide against malignant glioma in vitro and in vivo by
inhibiting autophagy. Free Radic Biol Med. 52:377–391.
2012.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Chen X, Xu H, Yu X, Wang X, Zhu X and Xu
X: Apigenin inhibits in vitro and in vivo tumorigenesis in
cisplatin-resistant colon cancer cells by inducing autophagy,
programmed cell death and targeting m-TOR/PI3K/Akt signalling
pathway. J BUON. 24:488–493. 2019.PubMed/NCBI
|
|
48
|
Tan S, Shi H, Ba M, Lin S, Tang H, Zeng X
and Zhang X: miR-409-3p sensitizes colon cancer cells to
oxaliplatin by inhibiting Beclin-1-mediated autophagy. Int J Mol
Med. 37:1030–1038. 2016.PubMed/NCBI View Article : Google Scholar
|
|
49
|
He Y, Zhang L, Tan F, Wang LF, Liu DH,
Wang RJ and Yin XZ: MiR-153-5p promotes sensibility of colorectal
cancer cells to oxaliplatin via targeting Bcl-2-mediated autophagy
pathway. Biosci Biotechnol Biochem. 84:1645–1651. 2020.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Cernigliaro C, D'Anneo A, Carlisi D,
Giuliano M, Gammazza AM, Barone R, Longhitano L, Cappello F,
Emanuele S and Distefano A: Ethanol-mediated stress promotes
autophagic survival and aggressiveness of colon cancer cells via
activation of Nrf2/HO-1 pathway. Cancers (Basel).
11(505)2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Schaaf MB, Keulers TG, Vooijs MA and
Rouschop KM: LC3/GABARAP family proteins: autophagy-(un)related
functions. FASEB J. 30:3961–3978. 2016.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Vera-Ramirez L, Vodnala SK, Nini R, Hunter
KW and Green JE: Autophagy promotes the survival of dormant breast
cancer cells and metastatic tumour recurrence. Nat Commun.
9(1944)2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Ma J, Dong C and Ji C: MicroRNA and drug
resistance. Cancer Gene Ther. 17:523–531. 2010.PubMed/NCBI View Article : Google Scholar
|