Introduction
Sevoflurane is a general inhalation anesthetic
commonly used in the clinic, which exhibits rapid induction and
recovery, and is widely used in cesarean sections and pediatric
surgery (1,2). However, the use of sevoflurane may
result in the decline of cognitive function during the recovery
period, which may be associated with the apoptosis of nerve cells
during anesthesia (3). The
hippocampus is an important area of consciousness and memory in
mammals (4), and sevoflurane may
damage neurons in this particular area, causing cognitive
impairment in anesthetized individuals (5). In the present study, the cause of
cognitive impairment induced by sevoflurane was investigated.
It has been reported that sevoflurane inhibits the
activity of neurons and even causes neuronal apoptosis (6-8).
In the present study, the effect of sevoflurane on neurons and its
association with Bax/BCL-2 was investigated. Bax is a well-known
apoptosis-promoting factor, which activates the caspase cascade
reaction to induce apoptosis (9).
BCL-2 is an anti-apoptotic molecule and BCL-2-like protein 11
(BCL2L11) is a member of the BCL-2 family (10). The increase of BCL-2 levels and the
decrease of Bax levels indicate that the resistance of cells to
apoptosis is enhanced (11). It has
been revealed that inhibition of BCL-2 or BCL2L11 gene expression
promotes apoptosis (12).
MicroRNA (miRNA/miR)-367-3p is a type of non-coding
endogenous miRNA consisting of 21 nucleotides, which widely exists
in eukaryotic cells (13).
miR-367-3p specifically acts on target mRNAs, and regulates gene
expression via incomplete complementation, inhibition of
translation and complete complementation (14). It has been confirmed that miR-367-3p
is involved in the process of hippocampal neuronal damage, although
the specific underlying mechanism remains unclear, and miR-367-3p
target gene regulation is still unknown (15). In the present study, TargetScan
software (16) was used to screen
targets for miR-367-3p and a luciferase reporter assay was employed
to explore the interaction between miR-367-3p and the BCL2L11 gene.
The potential mechanisms underlying the apoptosis of astrocytes in
the hippocampus induced by sevoflurane were also studied.
Materials and methods
Animal treatment and primary
astrocytes culture
All animal experiments were approved by the
Committee on Animal Experimentation of The Affiliated Hospital of
Putian University (Putian, China; approval no. 2018-0023) and
performed in compliance with the Guidelines for the Care and Use of
Laboratory Animals (17). A total
of 36 4-month-old adult male Sprague-Dawley rats (weight, 180-250
g) purchased from Beijing Vital River Laboratory Animal Technology
Co., Ltd. were divided into six groups (n=3). The rats were kept in
an animal breeding room for 2 weeks under the same conditions of
temperature (22±2˚C), humidity (50±10%) and controlled habitat with
a 12-h light/dark cycle (lights were on between 8:00 a.m. and 8:00
p.m.). Rats were administered food and water ab libitum.
Primary astrocytes were used in the present study,
which were isolated as follows: Rats were anesthetized with
different concentrations of sevoflurane (cat. no. 1612540;
Millipore Sigma) as follows: 0, 1, 2, 4, 8 or 16%, from 9:00 a.m.
to 3:00 p.m. Specifically, rats were housed in a chamber (length,
50 cm; width, 30 cm; height, 30 cm) containing 30% O2,
68% N2 and the corresponding concentrations of
sevoflurane for 6 h. Immediately after anesthesia, the animals were
euthanized via cervical dislocation and the heads were removed. The
isolated brains of rats were sterilized with PBS containing 10%
penicillin and streptomycin (cat. no. 10378016; Gibco; Thermo
Fisher Scientific, Inc.) and placed in precooled sterile calcium-
and magnesium-free (CMF) Hank's Balanced Salt Solution (HBSS) (cat.
no. H9394; Sigma-Aldrich; Merck KGaA). All the cerebral hemispheres
were collected in a petri dish on ice containing 5 ml CMF-HBSS, and
the collected cortical tissue was minced as finely as possible. The
tissue was transferred into a 15 ml tube with 12 ml CMF-HBSS. A
total of 1.5 ml 2.5% trypsin and 10 mg/ml DNase solution were
added, and the mixture was incubated at 37˚C for 15 min. The tissue
was triturated 10-15 times using a glass Pasteur pipette and the
mixture was further incubated at 37˚C for 5 min. The supernatant
was filtered using a 100-µm cell strainer. The suspension was
centrifuged at 130 x g for 6 min at 4˚C and the supernatant was
carefully discarded. It was ensured that the pelleted cells at the
bottom were not disturbed. Every group of samples was mixed
together with up to 20 ml of astrocyte medium (cat. no. A1261301;
Thermo Fisher Scientific, Inc.) to resuspend the combined
dissociated glial cells, which were subsequently cultured in a
flask in a humidified incubator at 37˚C with 5% CO2. The
astrocyte medium was changed on the next day. A total of 4 days
after seeding the cells, the flask was vigorously shaken 5-10 times
to remove the unwanted cell types, and the supernatant was
discarded. A total of 10 ml fresh CMF-HBSS was added in each flask,
which was vigorously shaken 5-10 times and the supernatant was
again discarded. A total of 20 ml fresh astrocyte medium was added
into each flask. The confluent cells were the desired astrocytes.
The astrocytes were maintained in high glucose DMEM (cat. no.
11965084) containing 10% FBS (cat. no. 16141061), and 1% penicillin
and streptomycin (cat. no. 10378016; all from Gibco; Thermo Fisher
Scientific, Inc.) in a humidified incubator at 37˚C with 5%
CO2. The astrocytes were then used for the following
experiments.
Transfection
Astrocytes were grouped into four groups:
Anti-miR-negative control (NC) [infected with lentivirus
(LV)-miR-367-3p inhibitor negative control], anti-miR-367-3p
(infected with LV-miR-367-3p inhibitor), miR-NC (infected with
LV-miR-367-3p mimic negative control), miR-367-3p (infected with
LV-miR-367-3p mimic). The lentiviral miRNA inhibitor vector was
pCLenti-U6-shRNA-CMV-EGFP-WPRE, and the lentiviral miRNA
overexpression vector was pCLenti-U6-miR30
(miRNA)-CMV-EGFP-F2A-Puro-WPRE, which were purchased from OBiO
Technology (Shanghai) Corp., Ltd. miR-367-3p inhibitor
(5'-AGTAATGGCCATCACCATTGCT-3'), inhibitor-NC
(5'-CCATTACTAGCAATGGTGATGG-3'), miR-367-3p mimic
(5'-CCATTACATGCAATGGTGATGGAU-3') and mimic-NC
(5'-AUCCATCACCATTGCTAGTAATGG-3') were constructed by Sangon Biotech
Co., Ltd. Lentiviral particles were packaged by co-transfection
with lentivirus and pHelper 1.0 and 2.0 vectors carrying the
miR-367-3p inhibitor, inhibitor-NC, miR-367-3p mimic or mimic-NC
into 293T cells (Procell Life Science & Technology Co., Ltd.).
In brief, a total of 20 µg lentiviral plasmids, including pHelper
1.0: pHelper 2.0: pLVX (LV-miR-367-3p inhibitor or NC;
LV-miR-367-3p mimic or NC)=7.5 µg: 2.5 µg: 10 µg, were added into
500 µl serum free DMEM (Solution A). A total of 60 µl
Lipofectamine® 3000 (cat. no. L3000015; Invitrogen;
Thermo Fisher Scientific, Inc.) was added into 500 µl serum free
DMEM (Solution B). Solution B was added drop by drop into solution
A, mixed gently and incubated for 20 min at room temperature to
prepare solution C. Solution C was added into 293T cells drop by
drop and mixed gently, and 293T cells were cultured in a 37˚C
incubator for 16 h. Subsequently, serum free DMEM was changed into
complete DMEM medium with 10% FBS. A total of 48 h after
transfection at 37˚C, lentivirus particles were harvested from the
cell culture supernatant and concentrated to 108
transduction units/ml.
A total of 24 h before lentivirus transduction,
astrocytes were seeded in 6-well plates with ~1x105
cells to obtain ~30% confluency on the 2nd day. On day 2, 6.8 µl
lentiviral particles (MOI=80) were added to each well. A total of
16 h later, the medium was replaced with lentivirus-free complete
DMEM medium, and the astrocytes were cultured for another 48 h for
transduction. Then, the lentiviral transduction efficiency was
evaluated under a fluorescence microscope (Eclipse Ts2; Nikon
Corporation).
For overexpression of BCL2L11, BCL2L11 open reading
frame was inserted into pGV358-GFP vector (TsingKe Biological
Technology) to construct pGV358-GFP-BCL2L11. pGV358-GFP was used as
NC. The following primers (forward, 5'-TCAGTGCCTTCTCCAGACCAGACG-3';
and reverse, 5'-CATCAGAAGGTTGCTTGGCCAT-3') were designed to amplify
the cDNA of BCL2L11. The lentiviral particles of pGV358-GFP-BCL2L11
were packaged following the procedures aforementioned. In brief, a
total of 20 µg lentiviral plasmids including pHelper 1.0 and 2.0
vectors and pGV358-GFP-BCL2L11 were incubated with 60 µl
Lipofectamine 3000 in 500 µl serum free DMEM for 20 min at room
temperature and then added to 293T cells for co-transfection for 48
h in a 37˚C incubator. Subsequently, lentiviral particles were
obtained from the cell supernatant and concentrated to 108
transduction units/ml. Astrocytes were then infected with the
packaged pGV358-GFP-BCL2L11 lentiviral particles for 48 h, followed
by adding G418 to the astrocytes for 4 weeks to obtain stable
BCL2L11-overexpressing astrocytes.
For co-transfection with pGV358-GFP-BCL2L11 and
LV-miR-367-3p mimic, stable BCL2L11-overexpressing astrocytes were
used, and 6.8 µl LV-miR-367-3p mimic lentiviral particles (MOI=80)
were added to each well of 6-well plate. A total of 16 h later, the
medium was replaced with lentivirus-free complete DMEM, and the
astrocytes were cultured for another 48 h.
Cell Counting Kit-8 (CCK-8) assay for
cell viability
As aforementioned, the hippocampal regions of the
brain were minced and digested into cell suspensions. These cell
suspensions were then added to a 96-well plate at 3x103
cells per well in 100 µl complete DMEM medium, and cultured at 37˚C
in a 5% CO2 incubator for 6 h. A total of 10 µl CCK-8
reagent (cat. no. 96992; Sigma-Aldrich; Merck KGaA) was added to
each well and cultured for 4 h. The absorbance was measured at 450
nm. Each sample was assayed in triplicate (18). The absorbance of the astrocytes
sample from rats anesthetized with 0% sevoflurane was used as
control group and calculated as 100% cell viability. The cell
viability of the other groups was calculated as the absorbance of
1, 2, 4, 8 or 16% sevoflurane group/the absorbance of 0%
sevoflurane group x100%.
Western blotting
Total protein was extracted from the astrocytes that
had been previously digested from hippocampal tissue using trypsin.
RIPA buffer (cat. no. 89900; Thermo Fisher Scientific, Inc.) with
PMSF (ratio, 1 ml:10 µl) was used to lyse the cells. Following
homogenization, the supernatant was prepared by centrifugation at
1,000 x g at 4˚C for 20 min, and the transparent colloidal mucus
was discarded. The protein was quantified using the BCA assay. A
total of 20 µg of the total protein per lane was mixed with SDS
loading buffer and proteins were denatured at 95-100˚C for 5 min.
The proteins were separated by 10% SDS-PAGE, and the separated
proteins were transferred to a PVDF membrane. The membrane was
blocked using 5% skimmed milk containing 0.1% Tween-20 (TBST) at
room temperature for 1 h, prior to incubation with the
corresponding primary antibody at 4˚C for 12 h. The membranes were
subsequently washed in TBST and incubated with the goat anti-rabbit
IgG H&L (HRP) secondary antibody (1:10,000; cat. no. ab6721;
Abcam) at room temperature for 2 h. The primary antibodies were as
follows: i) Bax (1:5,000; anti-rabbit; cat. no. ab32503; Abcam);
ii) BCL-2 (1:1,000; anti-rabbit; cat. no. ab32124; Abcam); iii)
BCL2L11 (1:2,000; anti-rabbit; cat. no. ab32158; Abcam); and iv)
GAPDH as a loading control (1:10,000; anti rabbit; cat. no.
ab181602; Abcam). Bands were developed using Immobilon ECL Ultra
Western HRP Substrate (cat. no. WBULS0100; MilliporeSigma) and the
image were analyzed using ImageJ v1.8.0 (National Institutes of
Health) (19).
TUNEL staining
TUNEL staining was carried out using Click-iT™ TUNEL
Alexa Fluor™ 488 Imaging Assay kit (cat. no. C10245; Invitrogen;
Thermo Fisher Scientific, Inc.). The experiment was conducted
following the manufacturer's protocol. In brief, a total of
2.5x105 astrocytes were seeded on coverslips and
cultured in 6-well plates (Corning, Inc.) for 24 h before TUNEL
staining. The astrocytes were rinsed with PBS three times, and then
fixed with 4% paraformaldehyde for 15 min at room temperature. A
total of 100 µl 0.25% Triton X-100 was added to completely cover
the coverslips and incubated for 20 min at room temperature. A
total of 100 µl terminal deoxynucleotidyl transferase (TdT)
reaction buffer was added on each coverslip and the solution was
allowed to spread completely over the surface. The coverslips were
incubated for 10 min at room temperature. Another 100 µl of the TdT
reaction cocktail containing TdT reaction buffer, EdUTP (dUTP
modified with an alkyne) and TdT, was added to each coverslip and
incubated them for 60 min at 37˚C. The coverslips were washed twice
with 3% BSA (cat. no. B2064; MilliporeSigma) in PBS. A total of 100
µl Click-iT reaction cocktail containing Click-iT reaction buffer
and Click-iT reaction buffer additive was added to each coverslip
and incubated them for 30 min at room temperature in the dark. The
coverslips were washed twice with 3% BSA in PBS. The primary
antibody solution was added as supplemented by the manufacturer and
incubated for 6 h at 4˚C, protected from light. The coverslips were
washed with 3% BSA in PBS twice, and subsequently incubated with
the secondary antibody for 0.5 h at room temperature, protected
from light, followed by washing with 3% BSA in PBS twice. A total
of 100 µl of 1X Hoechst 33342 solution per coverslip was added for
15 min at room temperature in the dark. The coverslips were washed
twice with PBS, and subsequently imaged coverslips under a
fluorescence microscope (Nikon Corporation) with fluorescence
excitation at 495 nm and emission at 519 nm. At least 5 fields were
captured per coverslip and 3 coverslips per group were analyzed to
calculate the TUNEL-positive cells (%).
Dual luciferase reporter assay
TargetScan v7.2 software (http://www.targetscan.org/vert_72/) was used to
predict the potential miRNAs interacting with the BCL2L11
3'-untranslated region (3'-UTR). Dual luciferase reporter assay was
carried out using Pierce™ Renilla-Firefly Luciferase Dual
Assay Kit (cat. no. 16185; Thermo Fisher Scientific, Inc.). In
brief, the reporter plasmids (80 ng) containing wild type (WT) or
mutant (Mut) 3'-UTR of BCL2L11were co-transfected with miR-367-3p
mimic (50 nM) or miR-NC (50 nM) into 293T cells using Lipofectamine
3000. miR-367-3p mimic (5'-CCATTACATGCAATGGTGATGGAU-3') and
mimic-NC (5'-AUCCATCACCATTGCTAGTAATGG-3') were constructed by
Sangon Biotech Co., Ltd.
A total of 48 h after transfection, the luciferase
assay reagent was added and luminescence signals were read at 640
nm for firefly luciferase and at 525 nm for green Renilla
luciferase. Firefly luciferase was an experimental reporter and
Renilla luciferase was used as a normalization control.
Reverse transcription-quantitative
(RT-q)PCR
Hippocampal tissue that had been previously digested
into individual cells using trypsin was used for PCR. RNA was
extracted from the cells using 1 ml TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and 200 µl chloroform
according to the manufacturer's protocol, incubated on a shaker,
and centrifuged at 10,000 x g at 4˚C for 15 min. The upper solution
was transferred to a clean RNase-free centrifuge tube and mixed
with an equal volume of isopropanol, incubated for 15 min on ice,
and centrifuged at 10,000 x g at 4˚C for 15 min. The RNA quality
was evaluated as the absorbance at 260/280 nm ratio being 1.8-2.1.
The samples were reverse transcribed to cDNA with PrimeScript RT
reagent kit (cat. no. RR036Q; Takara Bio, Inc.). The RT reaction
was performed at 37˚C for 15 min followed by 85˚C for 5 sec and 4˚C
on hold. The TB Green® Premix Ex Taq™ II kit (cat. no.
RR820A; Takara Bio, Inc. Takara) with an ABI 7500 real-time qPCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) was
used to conduct qPCR. The thermocycling conditions used were as
follows: 95˚C for 30 sec; followed by 40 cycles of 95˚C for 5 sec
and 60˚C for 10 sec. The 2-IICq method was applied for
calculating the relative quantification (20). U6 was used as reference control for
miR-367-3p.
The following primers were used: miR-367-3p forward,
5'-TTCTCCGAACTTTGCACGTTT-3' and reverse,
5'-ACGTGACACGTTCGGAGAATT-3'; U6 forward,
5'-GCTTCGGCAGCACATATACTAA-3' and reverse,
5'-AACGCTTCACGAATTTGCGT-3'.
Statistical analysis
All experimental data are presented as the mean ±
standard deviation. The data were analyzed using GraphPad Prism 6
statistical software (GraphPad Software, Inc.). Unpaired Student's
t-test was used for comparisons between two groups (21) and one-way ANOVA followed by
Bonferroni's correction was used for comparisons between multiple
groups. All experiments were repeated three times (n=3). P<0.05
was considered to indicate a statistically significant
difference.
Results
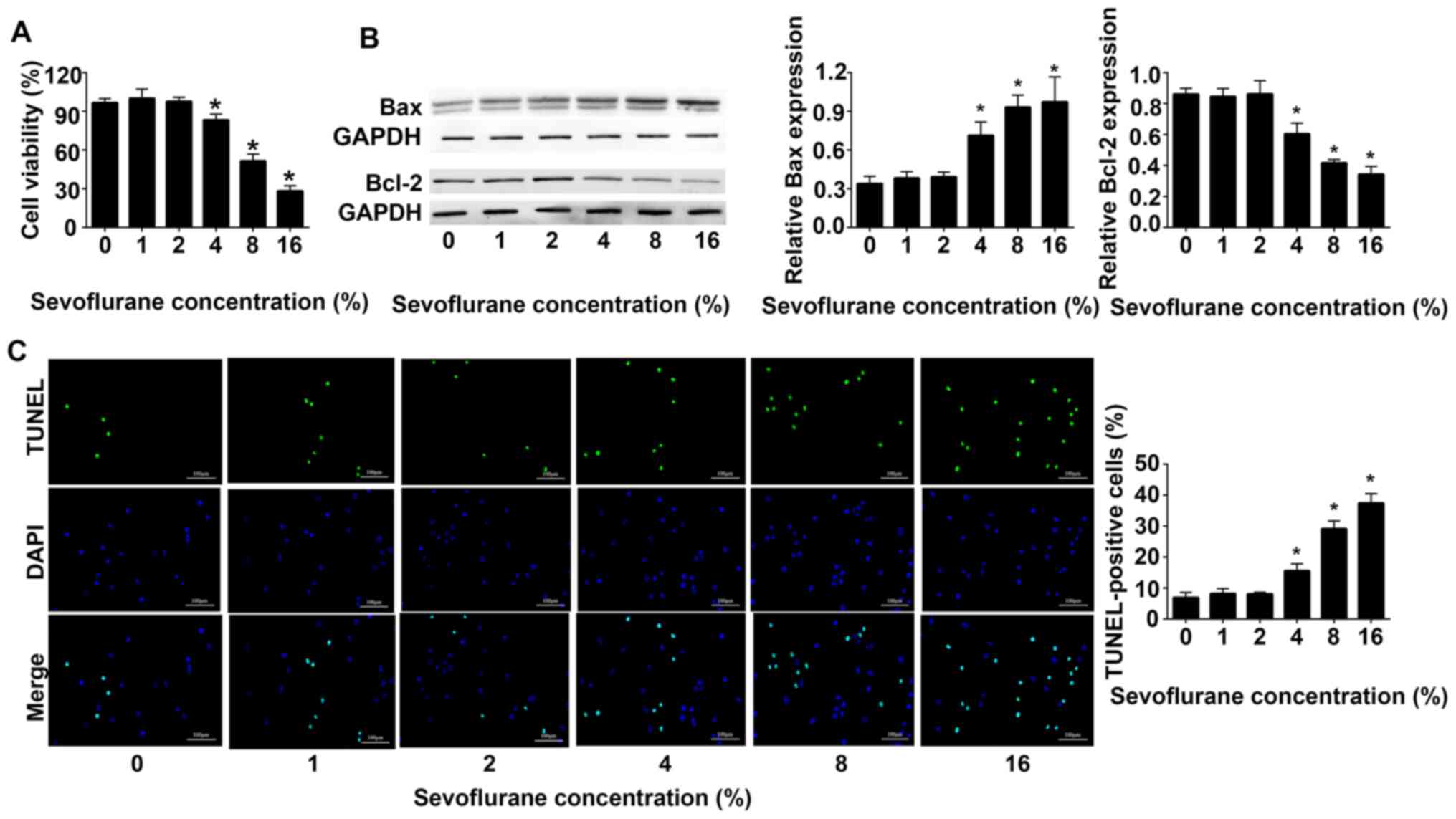
Sevoflurane inhibits the viability of
astrocytes in the hippocampus of adult rats and promotes astrocytes
apoptosis
Sevoflurane inhibited the viability of astrocytes in
a dose-dependent manner. The results of the CCK-8 cell viability
assay revealed that at ≥4% sevoflurane the cell viability decreased
in a dose-dependent manner compared with 0% sevoflurane (Fig. 1A). Western blot analysis indicated
that compared with 0% sevoflurane, the expression levels of Bax
were gradually increased, whereas the expression levels of Bcl-2
were gradually decreased. When the concentration of sevoflurane was
16%, the expression levels of Bax reached a maximum, and those of
Bcl-2 were the lowest (Fig. 1B).
The results of TUNEL assays demonstrated that with the increase of
sevoflurane concentration, the apoptosis of astrocytes increased.
When the concentration of sevoflurane reached 16%, the apoptosis of
astrocytes was the highest (Fig.
1C).
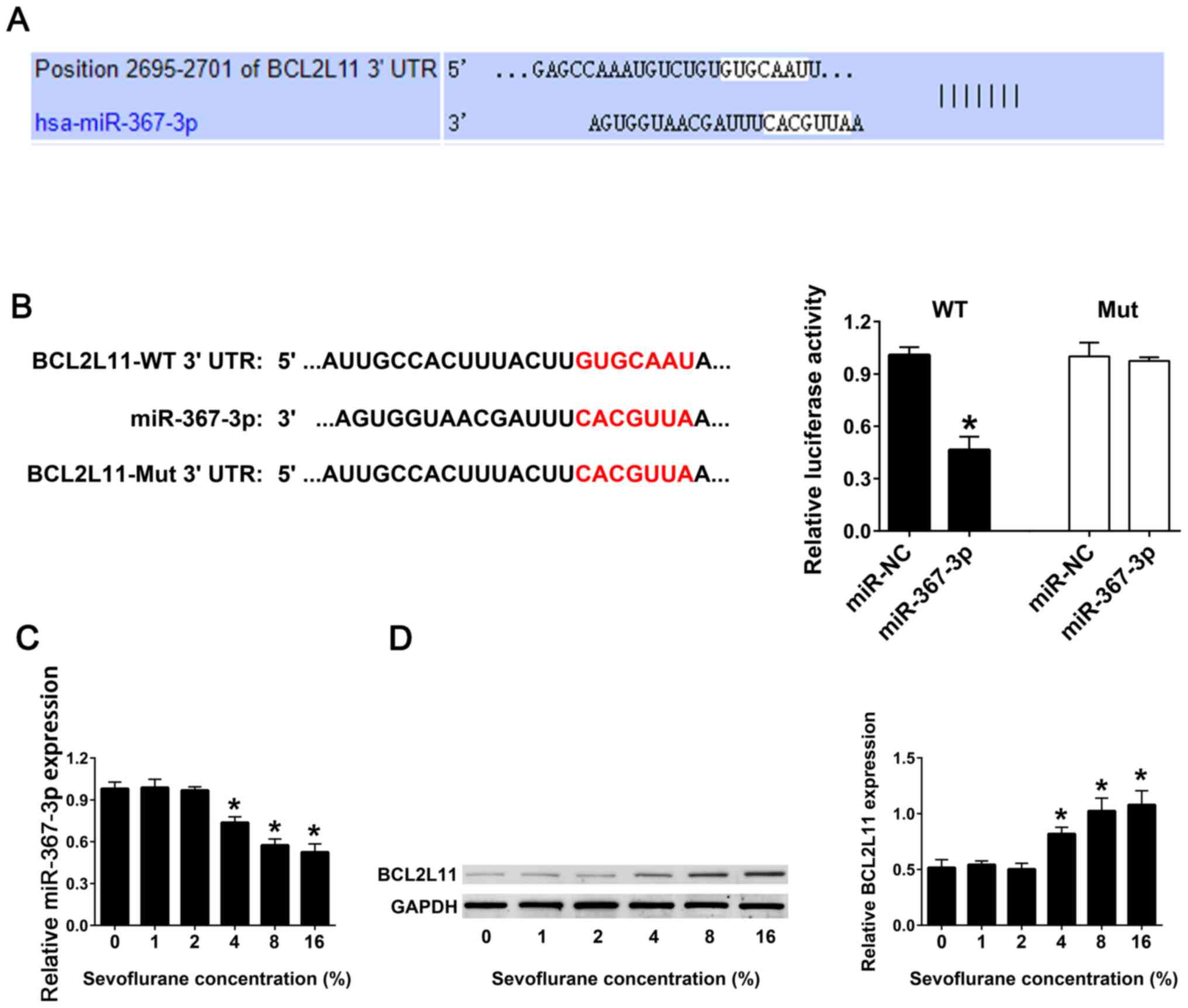
Sevoflurane inhibits the expression of
miR-367-3p in the hippocampal astrocytes
TargetScan software was used to predict the
potential miRNAs interacting with the BCL2L11 3'-UTR, and
miR-367-3p was identified (Fig.
2A). Luciferase reporter assay was used to analyze the binding
ability between the WT or Mut 3'-UTRs of BCL2L11 and miR-367-3p.
The results demonstrated that co-transfection of miR-367-3p and
BCL2L11-WT 3'-UTR exhibited a lower luciferase activity compared
with co-transfection of miR-NC and BCL2L11-WT 3'-UTR.
Co-transfection of the miR-367-3p or miR-NC and BCL2L11-Mut 3'-UTR
demonstrated no difference in the luciferase activity (Fig. 2B). RT-qPCR results revealed that
with an increase in the concentration of sevoflurane, the
expression levels of miR-367-3p in astrocytes gradually decreased,
compared with 0% sevoflurane. When the concentration of sevoflurane
reached 16%, the expression levels of miR-367-3p were the lowest,
compared with 0% sevoflurane (Fig.
2C). Western blot analysis was used to detect the protein
expression levels of BCL2L11 in astrocytes. The results
demonstrated that with the increase of sevoflurane concentration,
the expression levels of the BCL2L11 protein in hippocampal
astrocytes were significantly increased. When the concentration of
sevoflurane reached 16%, the expression of BCL2L11 protein was the
highest, compared with 0% sevoflurane (Fig. 2D).
miR-367-3p inhibits the expression of
BCL2L11 in astrocytes
In order to detect the interaction between
miR-367-3p and BCL2L11, lentiviral infection with anti-miR-367-3p
or miR-367-3p into the astrocytes from rats not treated with
sevoflurane was performed. RT-qPCR results indicated that
transfection with anti-miR-367-3p significantly decreased the
expression levels of miR-367-3p compared with the anti-miR-NC
group, whereas transfection with miR-367-3p significantly increased
the expression levels of miR-367-3p compared with miR-NC group
(Fig. 3A). Western blot analysis
revealed that the protein expression levels of BCL2L11 were
significantly increased following transfection with
anti-miR-367-3p, and were significantly decreased after
transfection with miR-367-3p, compared with anti-miR-NC or miR-NC,
respectively (Fig. 3B).
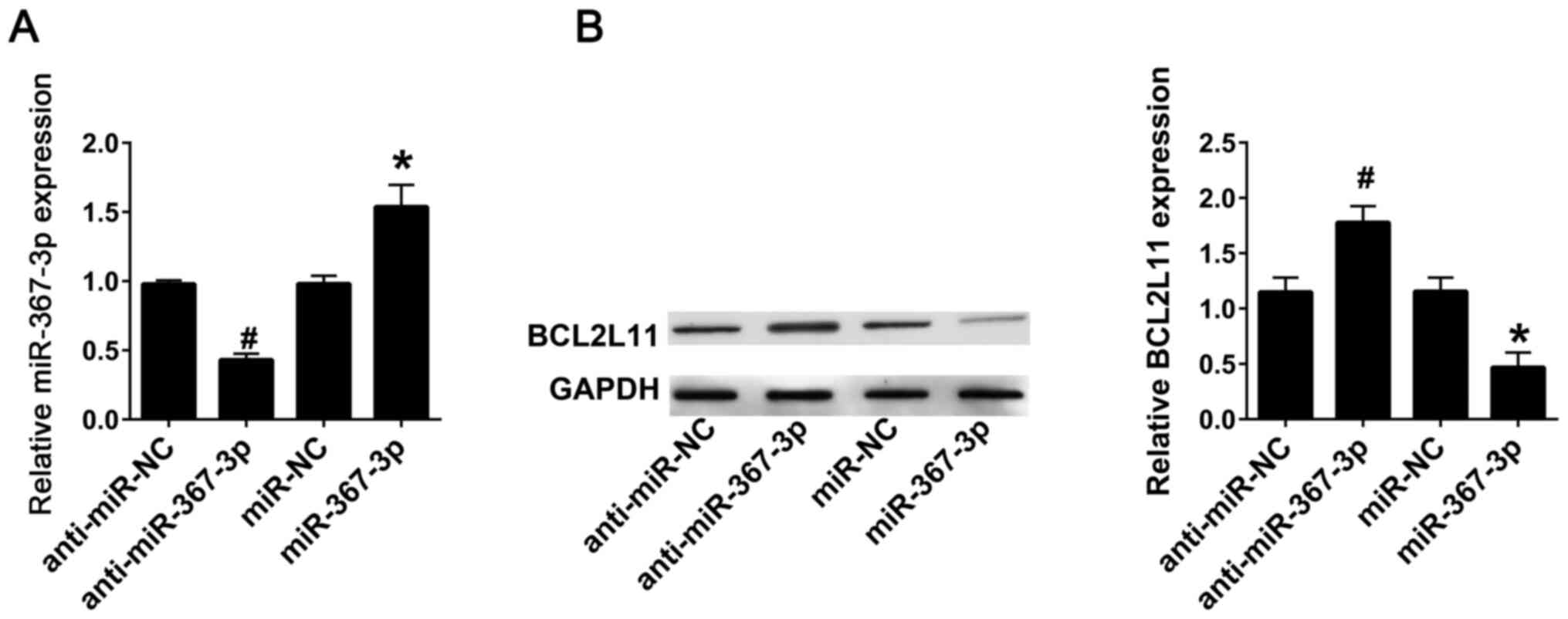 | Figure 3miR-367-3p inhibits BCL2L11 in
astrocytes. (A) The expression of miR-367-3p was detected using
reverse transcription-quantitative PCR following transfection with
anti-miR-NC, anti-miR-367-3p, miR-NC or miR-367-3p. (B) The
expression of BCL2L11 was detected using western blotting following
transfection with anti-miR-NC, anti-miR-367-3p, miR-NC or
miR-367-3p. (n=3). #P<0.05 vs. anti-miR-NC;
*P<0.05 vs. miR-NC. miR, microRNA; BCL2L11,
BCL-2-like protein 11; NC, negative control; WT, wild-type; Mut,
mutant; UTR, untranslated region. |
Transfection with miR-367-3p inhibits
the expression of BCL2L11 and prevents the sevoflurane-induced
apoptosis of rat hippocampal astrocytes
Western blot analysis revealed that under 8%
sevoflurane treatment, the protein expression of BCL2L11 and Bax in
the astrocytes transfected with anti-miR-367-3p was significantly
increased, while the protein expression of Bcl-2 was decreased,
compared with astrocytes transfected with anti-miR-NC. The protein
expression of BCL2L11 and Bax in astrocytes transfected with
miR-367-3p were significantly decreased, and the protein expression
of BcL-2 were significantly increased, compared with astrocytes
transfected with miR-NC (Fig. 4A).
The results of TUNEL staining revealed that apoptosis of the
anti-miR-367-3p-transfected astrocytes were significantly
increased, and that of the miR-367-3p-transfected astrocytes were
significantly decreased (Fig. 4B),
compared with anti-miR-NC or miR-NC, respectively.
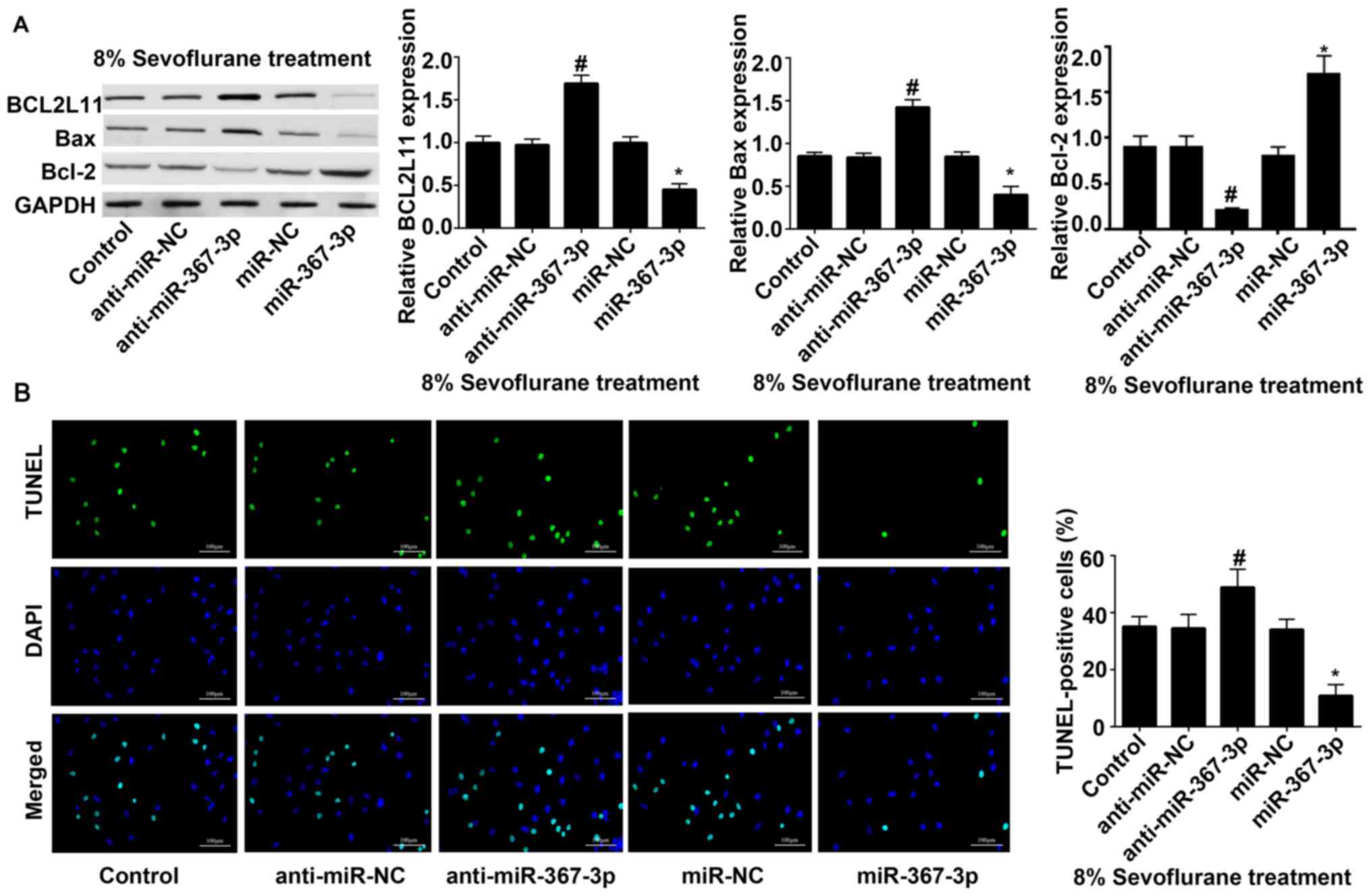 | Figure 4Transfection with miR-367-3p inhibits
the expression of BCL2L11 and prevents sevoflurane-induced
apoptosis of rat hippocampal astrocytes. (A) The expression of
BCL2L11, Bax and Bcl-2 was detected using western blot assay
following transfection with anti-miR-NC, anti-miR-367-3p, miR-NC,
miR-367-3p under the condition of 8% sevoflurane treatment. (B)
TUNEL staining was conducted following transfection with
anti-miR-NC, anti-miR-367-3p, miR-NC, miR-367-3p under the
condition of 8% sevoflurane treatment. Scale bars, 100 µm. (n=3).
#P<0.05 vs. anti-miR-NC; *P<0.05 vs.
miR-NC. miR, microRNA; BCL2L11, BCL-2-like protein 11; NC, negative
control. |
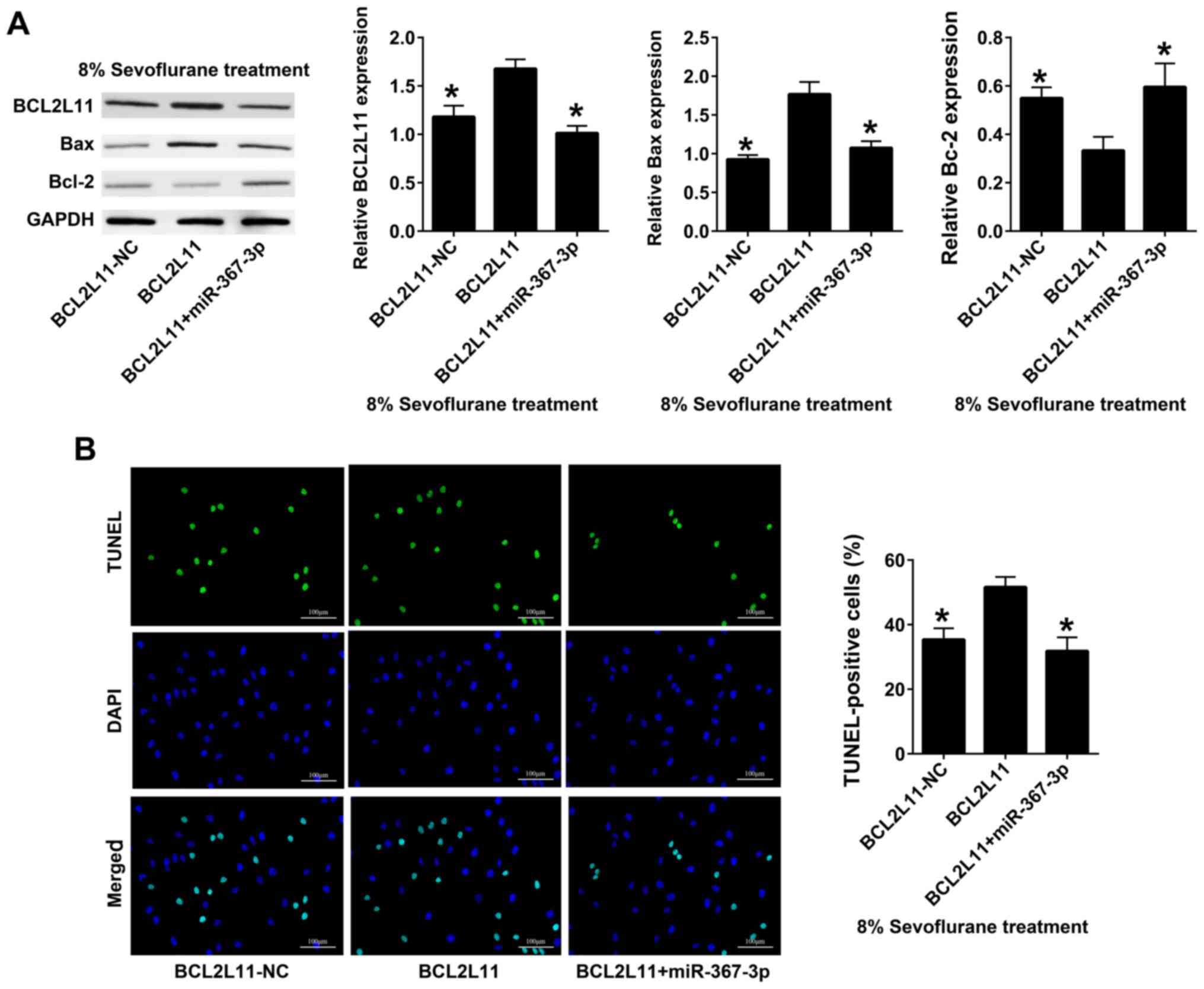
miR-367-3p decreases the effect of
recombinant BCL2L11 on astrocytes apoptosis in sevoflurane-treated
rats
BCL2L11 and miR-367-3p were co-transfected into rat
hippocampal astrocytes; BCL2L11-NC was used as the control group.
Western blot analysis revealed that the expression levels of
BCL2L11 and Bax in co-transfected BCL2L11 + miR-367-3p astrocytes
were significantly lower compared with those transfected with
BCL2L11 alone, whereas the expression levels of BcL-2 in
co-transfected BCL2L11 + miR-367-3p astrocytes were significantly
higher than those transfected with BCL2L11 alone (Fig. 5A). The results of TUNEL staining
demonstrated that the apoptosis of co-transfected BCL2L11 +
miR-367-3p astrocytes was lower than that of cells transfected with
BCL2L11 alone (Fig. 5B).
Discussion
Sevoflurane was initially discovered by Terrell
(22) and was approved by the
Japanese Drug Administration as a clinically inhaled anesthetic in
1990(23). Sevoflurane is a common
drug in inhalation anesthesia (24). However, in recent years, studies
have demonstrated that sevoflurane is neurotoxic. For example,
Beekoo et al (25)
demonstrated that sevoflurane may cause behavioral and
developmental cognitive disorders in children <3-years-old. Zhou
et al (26) treated
7-day-old rats with 2.3% sevoflurane for 6 h, in order to
demonstrate that exposure of the developing brain to sevoflurane
may lead to hippocampal synaptic plasticity damage, hippocampal
area-dependent learning and memory impairment. Yang et al
(27) used the Morris water maze
test to detect the cognitive function of rats (P30) who had inhaled
3% sevoflurane for 4 h. The results revealed that sevoflurane
caused cognitive impairment in rats, which may have been due to the
promotion of apoptosis of neurons in the hippocampus. The results
of the present study confirmed the aforementioned hypothesis.
Sevoflurane caused apoptosis of nerve cells in the hippocampus,
promoted the expression of the apoptotic protein Bax and BCL2L11,
and inhibited the expression of the anti-apoptotic proteins BCL-2.
BCL-2 has been indicated to serve an important role in
sevoflurane-mediated cell death; sevoflurane was previously shown
to facilitate cell apoptosis in glioma progression by inhibiting
BCL-2(28). miR-34c has previously
been shown to participate in apoptosis mediated by sevoflurane in
rat brains via the mitochondrial pathway (29). Zhu et al (30) exposed 14-day-old mice to 3%
sevoflurane for 2 h for 3 consecutive days. The results revealed
that sevoflurane increased the expression of Bax,
caspase-3 and other apoptosis-related genes in the
hippocampus, and inhibited the expression of BCL-2.
miR-367-3p is an endogenous small molecule
non-coding RNA. In the present study, miR-367-3p reversed the
induction of apoptosis by sevoflurane, potentially rendering
neurons in the hippocampus resistant to sevoflurane. A study by
Kaid et al (31)
demonstrated that miR-367-3p exhibited certain stem cell-enhancing
characteristics, and improved the pluripotency of embryonic stem
cells and the invasiveness of cancer-like stem cells. Wang et
al (32) revealed that
treatment with adriamycin induced apoptosis of osteosarcoma cells
with upregulation of miR-367-3p. Additionally, miR-367-3p has been
demonstrated to be downregulated in brain homogenates with
sustained ischemia (15). However,
the association between miR-367-3p and neuronal cell apoptosis
remains unclear. The results of the present study revealed that
miR-367-3p inhibited the expression of Bax and BCL2L11, and
promoted the expression of BCL-2. The target gene of miR-367-3p was
also verified as BCL2L11 using TargetScan, a target gene database
of mammalian miRNA. Therefore, these findings indicated that
miR-367-3p inhibited apoptosis and protected neuronal cells.
In conclusion, miR-367-3p reversed the
apoptosis-promoting effect of sevoflurane on neurons. miR-367-3p
suppressed sevoflurane-induced neuronal apoptosis by targeting
BCL2L11. Sevoflurane promoted the expression of Bax and BCL2L11,
and inhibited the expression of BCL-2, whereas miR-367-3p reversed
these expression changes. miR-367-3p may therefore be a target in
the field of anesthesia recovery and also an important target in
protection from anesthesia toxicity/adverse effects.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DMX and CBZ conceived and supervised the study; JYL
designed the experiments; WHC and WL performed the experiments and
analyzed the data; DMX and CBZ confirm the authenticity of all the
raw data; DMX wrote the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the
Committee on Animal Experimentation of The Affiliated Hospital of
Putian University (Putian, China). All applicable international,
national and/or institutional guidelines for the care and use of
animals were followed.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Edgington TL, Muco E and Maani CV (eds):
Sevoflurane. In: StatPearls [Internet]. StatPearls Publishing,
Treasure Island, FL, 2021.
|
|
2
|
Brioni JD, Varughese S, Ahmed R and Bein
B: A clinical review of inhalation anesthesia with sevoflurane:
From early research to emerging topics. J Anesth. 31:764–778.
2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Yang X, Zhang W, Wu H, Fu S, Yang J, Liu
S, Zhao Y, Zhang X and Liu J: Downregulation of CDK5 restores
sevoflurane-induced cognitive dysfunction by promoting
SIRT1-mediated autophagy. Cell Mol Neurobiol. 40:955–965.
2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Guo S, Liu L, Wang C, Jiang Q, Dong Y and
Tian Y: Repeated exposure to sevoflurane impairs the learning and
memory of older male rats. Life Sci. 192:75–83. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Satomoto M, Sun Z, Adachi YU, Kinoshita H
and Makita K: Sevoflurane preconditioning ameliorates
lipopolysaccharide-induced cognitive impairment in mice. Exp Anim.
67:193–200. 2018.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Andropoulos DB: Effect of anesthesia on
the developing brain: Infant and fetus. Fetal Diagn Ther. 43:1–11.
2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Istaphanous GK and Loepke AW: General
anesthetics and the developing brain. Curr Opin Anaesthesiol.
22:368–373. 2009.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Zuo Y, Chang Y, Thirupathi A, Zhou C and
Shi Z: Prenatal sevoflurane exposure: Effects of iron metabolic
dysfunction on offspring cognition and potential mechanism. Int J
Dev Neurosci. 81:1–9. 2021.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Qi Z, Li S, Su Y, Zhang J, Kang Y, Huang
Y, Jin F and Xing Q: Role of microRNA-145 in protection against
myocardial ischemia/reperfusion injury in mice by regulating
expression of GZMK with the treatment of sevoflurane. J Cell
Physiol: Mar 14, 2019 (Epub ahead of print). doi:
10.1002/jcp.28323.
|
|
10
|
Chen X, Zhou X, Lu D, Yang X, Zhou Z, Chen
X, Chen Y, He W and Feng X: Aberrantly expressed long noncoding
RNAs are involved in sevoflurane-induced developing hippocampal
neuronal apoptosis: A microarray related study. Metab Brain Dis.
31:1031–1040. 2016.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bedirli N, Bagriacik EU, Yilmaz G, Ozkose
Z, Kavutçu M, Cavunt Bayraktar A and Bedirli A: Sevoflurane exerts
brain-protective effects against sepsis-associated encephalopathy
and memory impairment through caspase 3/9 and Bax/Bcl signaling
pathway in a rat model of sepsis. J Int Med Res. 46:2828–2842.
2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang LM, Zhao XC, Sun WB, Li R and Jiang
XJ: Sevoflurane post-conditioning protects primary rat cortical
neurons against oxygen-glucose deprivation/resuscitation via
down-regulation in mitochondrial apoptosis axis of Bid, Bim,
Puma-Bax and Bak mediated by Erk1/2. J Neurol Sci. 357:80–87.
2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Subramanyam D, Lamouille S, Judson RL, Liu
JY, Bucay N, Derynck R and Blelloch R: Multiple targets of miR-302
and miR-372 promote reprogramming of human fibroblasts to induced
pluripotent stem cells. Nat Biotechnol. 29:443–448. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Kaid C, Silva PB, Cortez BA, Rodini CO,
Semedo-Kuriki P and Okamoto OK: miR-367-3p promotes proliferation
and stem-like traits in medulloblastoma cells. Cancer Sci.
106:1188–1195. 2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Tabet F, Lee S, Zhu W, Levin MG, Toth CL,
Cuesta Torres LF, Vinh A, Kim HA, Chu HX, Evans MA, et al:
MicroRNA-367-3p regulation of GPRC5A is suppressed in ischemic
stroke. J Cereb Blood Flow Metab. 40:1300–1315. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhou Z, Dong Y, Zhou H, Liu J and Zhao W:
miR-143-3p directly targets GLUT9 to reduce uric acid reabsorption
and inflammatory response of renal tubular epithelial cells.
Biochem Biophys Res Commun. 517:413–420. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Jones-Bolin S: Guidelines for the care and
use of laboratory animals in biomedical research. Curr Protoc
Pharmacol. 59:A.4B.1–A.4B.9. 2012.PubMed/NCBIdoi: 10.1002/0471141755.pha04bs59.
|
|
18
|
Xu Y, Wang H, Li F, Heindl LM, He X, Yu J,
Yang J, Ge S, Ruan J, Jia R and Fan X: Long non-coding RNA
LINC-PINT suppresses cell proliferation and migration of melanoma
via recruiting EZH2. Front Cell Dev Biol. 7(350)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Abd-Elrahim Batran S: Toxicity of
environmental ozone exposure on mice olfactory bulbs, using western
blot technique. Toxicol Rep. 7:453–459. 2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Geng C, Qiao Y, Guo Y, Han W, Wu B, Wang
C, Zhang J, Chen D, Yang M and Jiang P: Integrated metabolomics and
lipidomics profiling of hippocampus reveal metabolite biomarkers in
a rat model of chronic unpredictable mild stress-induced
depression. Ann Transl Med. 7(781)2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Terrell RC: The invention and development
of enflurane, isoflurane, sevoflurane, and desflurane.
Anesthesiology. 108:531–533. 2008.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kasahara M, Ichinohe T, Okamoto S, Okada
R, Kanbe H and Matsuura N: Concomitant administration of nitrous
oxide and remifentanil reduces oral tissue blood flow without
decreasing blood pressure during sevoflurane anesthesia in rabbits.
J Anesth. 29:421–425. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Olutoye OA, Baker BW, Belfort MA and
Olutoye OO: Food and drug administration warning on anesthesia and
brain development: Implications for obstetric and fetal surgery. Am
J Obstet Gynecol. 218:98–102. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Beekoo D, Yuan K, Dai S, Chen L, Di M,
Wang S, Liu H and ShangGuan W: Analyzing electroencephalography
(EEG) waves provides a reliable tool to assess the depth of
sevoflurane anesthesia in pediatric patients. Med Sci Monit.
25:4035–4040. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhou X, Li W, Chen X, Yang X, Zhou Z, Lu D
and Feng X: Dose-dependent effects of sevoflurane exposure during
early lifetime on apoptosis in hippocampus and neurocognitive
outcomes in Sprague-Dawley rats. Int J Physiol Pathophysiol
Pharmacol. 8:111–119. 2016.PubMed/NCBI
|
|
27
|
Yang F, Shan Y, Tang Z, Wu X, Bi C, Zhang
Y, Gao Y and Liu H: The neuroprotective effect of hemin and the
related mechanism in sevoflurane exposed neonatal rats. Front
Neurosci. 13(537)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li H, Xia T, Guan Y and Yu Y: Sevoflurane
regulates glioma progression by Circ_0002755/miR-628-5p/MAGT1 axis.
Cancer Manag Res. 12:5085–5098. 2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhou X, Xian D, Xia J, Tang Y, Li W, Chen
X, Zhou Z, Lu D and Feng X: MicroRNA-34c is regulated by p53 and is
involved in sevoflurane-induced apoptosis in the developing rat
brain potentially via the mitochondrial pathway. Mol Med Rep.
15:2204–2212. 2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhu J, Zhang Z, Jia J, Wang L, Yang Q,
Wang Y and Chen C: Sevoflurane induces learning and memory
impairment in young mice through a reduction in neuronal glucose
transporter 3. Cell Mol Neurobiol. 40:879–895. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kaid C, Jordan D, Bueno HMS, Araujo BHS,
Assoni A and Okamoto OK: miR-367 as a therapeutic target in
stem-like cells from embryonal central nervous system tumors. Mol
Oncol. 13:2574–2587. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang GC, He QY, Tong DK, Wang CF, Liu K,
Ding C, Ji F and Zhang H: miR-367 negatively regulates apoptosis
induced by adriamycin in osteosarcoma cells by targeting KLF4. J
Bone Oncol. 5:51–56. 2016.PubMed/NCBI View Article : Google Scholar
|