Introduction
In mammalian cells, post-transcriptional gene
regulation involves multistep processes, including precursor mRNA
(pre-mRNA) splicing and mRNA export, storage, stability and
translation. Post-transcriptional gene regulation is substantially
governed by two types of factor: RNA-binding proteins (RBPs) and
microRNAs (miRNAs/miRs) (1).
AU-rich element RNA binding protein 1 (AUF1), also known as
heterogeneous nuclear ribonucleoprotein D, is one of the most
extensively characterized RBPs and primarily functions as an mRNA
destabilizing factor (2).
A total of four isoforms of AUF1 arising from
differential exon splicing of a single pre-mRNA have been
identified and named according to their apparent molecular masses
(p37, p40, p42 and p45) (2-4).
AUF1 degrades diverse groups of transcripts, including regulators
of the cell cycle and apoptosis, inflammatory mediators and
cytokines and proto-oncogenes; thus, AUF1 influences oncogenic
processes and inflammatory diseases (5,6). For
example, AUF1 in esophageal squamous cell carcinoma (ESCC) tumor
tissues exhibits high expression, while knockdown of AUF1 causes
markedly reduced proliferation and enhanced cell apoptosis in
Eca-109 cells (7). IL-10, an
important anti-inflammatory cytokine, exerts its action by
inhibiting proinflammatory cytokine production by mononuclear
phagocytes (8). The AUF1 isoform
p40 plays a positive role in regulating lipopolysaccharide
exposure-induced expression of IL-10 mRNA and protein in human
promonocytic leukemia cells (8).
AUF1 may also be involved in thyroid carcinoma
progression. A previous study has revealed that thyroid tissues
have revealed increased cytoplasmic expression of AUF1 in malignant
tissues (9). In hepatocellular
carcinoma (HCC), AUF1 suppresses the expression of liver-specific
miR-122, an established tumor suppressor miRNA, while knockdown of
AUF1 promotes HCC cell death (10). AUF1 delays the senescence of
vascular endothelial cells, indicating its influences on delaying
aging and prolonging life (11).
AUF1 was also revealed to involve the antioxidant system through
regulation of superoxide dismutase 1 expression in human cancer
cells (12).
Skin is the largest organ of the human body and acts
as a protective barrier against pathogens and injuries (13). Mice lacking AUF1 have been
demonstrated to develop chronic dermatitis, underscoring the role
of AUF1 in the complex inflammatory response in skin (14). However, to the best of our
knowledge, the role of AUF1 in human skin homeostasis is still
unclear. The present study explored the function of AUF1 in human
skin cells and characterized its downstream mRNAs by RNA-sequencing
(RNA-Seq).
Materials and methods
Cell culture
Human immortalized keratinocyte HaCaT cells and
human embryonic dermal fibroblast WS1 cells were purchased from the
American Type Culture Collection. They were both cultured in DMEM
(cat. no. D0822; Sigma-Aldrich; Merck KGaA) supplemented with 10%
fetal bovine serum (FBS; cat. no. 04-001-1A; Biological
Industries), 100 U/ml streptomycin and 100 U/ml penicillin (cat.
no. 15140122; Gibco; Thermo Fisher Scientific, Inc.) at 37˚C in a
5% CO2 atmosphere.
Cell infection
To explore the function of AUF1 in skin cells,
overexpression and RNA interference experiments were carried out.
AUF1 complementary (c)DNA was obtained from OriGene Technologies,
Inc. (cat. no. SC107836). AUF1 cDNA was subcloned into the
pHBAD-EF1-mcs-CMV vector (Hanbio Biotechnology Co., Ltd.) for
adenovirus construction. Packaging and purification of adenovirus
overexpressing AUF1 (Ad-AUF1) and negative control adenovirus
(Ad-NC) were carried out by Hanbio Biotechnology Co., Ltd. AUF1
cDNA was also subcloned into the pHBLV-U6-ZsGreen-Puro vector for
lentivirus construction. Lentiviruses targeting AUF1 [named short
hairpin (sh)AUF1-1 and shAUF1-2, respectively] and a scrambled
negative control lentivirus (sh-NC) were purchased from Hanbio
Biotechnology Co., Ltd. The target sequences against AUF1 are
listed in Table SI. A
mock-infected control group (without vector) was also constructed
to demonstrate the background expression value.
HaCaT and WS1 cells were cultured in a six-well
culture plate. After reaching 70% confluence, the cells were both
infected with adenoviruses and lentiviruses at a multiplicity of
infection of 20 with DMEM free-serum for 6 h at 37˚C in 5%
CO2. After 48 h, the cells were harvested for further
experiments, and the efficiency of infection was determined by
reverse transcription-quantitative PCR (RT-qPCR) and western
blotting.
RNA preparation and RT-qPCR
Total RNA of HaCaT and WS1 cells was extracted using
TRIzol® (cat. no. 15596018; Thermo Fisher Scientific,
Inc.), and the absorbance values at 230, 260 and 280 nm were
measured to determine the purity and density of the RNA.
Subsequently, 1 µg of total RNA from each sample was used to
synthesize single-stranded cDNA with a PrimeScript™ RT reagent kit
(cat. no. RR047A; Takara Bio, Inc.). The reaction was performed at
37°C for 15 min followed by 85˚C for 5 sec. cDNA was amplified via
qPCR with TB Green® dye (cat. no. RR420S Takara Bio,
Inc.) on a Prism 7500 real-time PCR machine (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The thermocycling conditions
included an initial denaturation step of 95˚C for 60 sec, followed
by 40 cycles of amplification at 95˚C for 15 sec and then annealing
at 60˚C for 30 sec. The β-actin gene was used to normalize the
relative fold changes of the target genes (15). The details of the primer sequences
are listed in Table SII.
Western blotting
HaCaT and WS1 cells were harvested and lysed in RIPA
lysis buffer (containing 50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1%
Triton X-100, 1% sodium deoxycholate and 0.1% SDS) containing
protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA) for 30 min
at 4˚C. Protein concentration was subsequently measured using a BCA
Protein Assay kit (cat. no. P0012; Beyotime Institute of
Biotechnology). Protein (50 µg/lane) from each lysate was
fractionated by 10% SDS-PAGE and transferred to polyvinylidene
difluoride membranes (MilliporeSigma). After blocking with 5%
non-fat milk in PBS and 0.1% Tween-20 for 1 h at room temperature,
the membranes were blotted with AUF1 (cat. no. ab282018; 1:1,000)
and GAPDH (cat. no. ab181602; 1:50,000) antibodies overnight at
4˚C. After washing four times with PBST, the membranes were
incubated with horseradish peroxidase-conjugated goat anti-rabbit
IgG (cat. no. A21020; 1:10,000; Abbkine Scientific, Co., Ltd.)
secondary antibodies for 2 h. Following washing in PBST, the
protein-bound antibodies were detected using an enhanced
chemiluminescence stable peroxide solution (cat. no. DC10100;
PointBio). All protein bands were visualized by a FluroChem MI
imaging system (Alpha Innotech Corporation) at room temperature.
Densitometry was performed using Image J Java 1.8.0-172 software
(National Institutes of Health).
Cell viability and proliferation
assay
After infection, cell viability was assayed using a
Cell Counting Kit-8 assay (CCK-8; cat. no. CK04; Dojindo
Laboratories, Inc.). Cells were seeded in a 96-well plate
(1x104 cells/well) and cultured for 24 and 48 h at 37˚C
in a CO2 incubator. After 10 µl of CCK-8 solution was
added to each well, the cells were incubated for another 2 h.
Absorbance values were determined at 450 nm using a microplate
reader (Bio-Rad Laboratories, Inc.).
To measure the proliferation of infected skin cells,
an EdU incorporation assay was performed by an EdU-based cell
proliferation kit (cat. no. C0071; Beyotime Institute of
Biotechnology) according to the manufacturer's instructions after
48 h of infection. In brief, HaCaT and WS1 cells at 50-70%
confluence were incubated with 50 µM EdU for 2 h at 37˚C.
Subsequently, the cells were fixed with 4% paraformaldehyde for 15
min at room temperature, permeabilized with 0.3% Triton X-100 and
washed with phosphate-buffered saline (PBS). The cell nuclei were
stained with 5 µg/ml Hoechst 33342 for 30 min at room temperature.
A florescence microscope (Olympus Corporation; magnification, x20)
was used to examine the cells.
Colony formation assay
HaCaT and WS1 cells infected with the indicated
virus were seeded in 60-mm petri dishes at low density (1,000
cells/dish). After 10 days, the cells were fixed with 3.7% methanol
for 30 min at room temperature and stained with 0.1% crystal violet
for 30 min at room temperature. Colonies consisting of >50 cells
were counted manually under a light microscope (Olympus
Corporation; magnification, x40).
Scratch-wound assay
Infected HaCaT and WS1 cells were seeded onto 6-well
plates and cultured for 24 h at 37˚C to form a 50% confluent
monolayer. A straight scratch was gently and slowly created using a
p200 pipette tip in the cell monolayer. Images of the wound were
captured using a light microscope (Olympus Corporation;
magnification, x10). Following two gentle washes with PBS, the
plates were replenished with fresh DMEM containing 2% FBS for
regular culture. Images of the width of the scratched regions after
24 and 48 h were captured and wound closure was assessed using
Image J Java 1.8.0-172 software (National Institute of Health). The
relative wound width rate was calculated by dividing the changed
distance in the scratched region by the initial distance.
Apoptosis assay
Early and late stage of apoptosis was evaluated
using an Annexin V-FITC apoptosis detection kit (cat. no. C1062;
Beyotime Institute of Biotechnology). HaCaT and WS1 cells were
harvested 48 h post-infection and then stained with Annexin V/PI
for 30 min at room temperature. To compute the percentage of
apoptotic cells, a flow cytometer (FACSCalibur; BD Biosciences)
with ModFit's LT v.3.0 software (BD Diagnostics) was used for data
analysis.
Cell senescence assay
HaCaT and WS1 cells were fixed in 2%
formaldehyde/0.2% glutaraldehyde for 5 min at room temperature.
β-Galactosidase staining solution containing X-gal (cat. no. C0602;
Beyotime Institute of Biotechnology) was added after rinsing with
PBS. The cells were then incubated for 6-10 h in a 37˚C incubator
without CO2. Senescent cells (stained blue) were
observed and images were captured using light microscopy (Olympus
Corporation; magnification, x10), and positive staining areas were
calculated by determining the percentage of SA-β-gal+
cells in five random fields in each of the three wells.
Cell cycle assay
HaCaT and WS1 cells were seeded in 100-mm petri
dishes. At 24 h, the cells were collected, washed with PBS and
fixed with 70% precooled ethanol overnight at 4˚C. After the cells
were stained with propidium iodide staining solution, including
RNase A (50X; cat. no. C1052; Beyotime Institute of Biotechnology)
in the dark for 30 min at 37˚C to detect S phase, G1
phase and G2/M phase arrest. The cell cycle distribution
was analyzed using a FACSCalibur system with ModFit's LT
software.
RNA-Seq of gene expression and
pathway-based analysis
Total RNA was extracted from infected WS1 cells with
TRIzol® reagent (cat. no. 15596018; Invitrogen; Thermo
Fisher Scientific, Inc.) and quantified by a NanoDrop ND-2000C
spectrophotometer (Thermo Fisher Scientific, Inc.). RNA integrity
was assessed using the Agilent 2100 Bioanalyzer (Agilent
Technologies, Santa Clara, CA, USA). The libraries were
subsequently constructed using a TruSeq Stranded mRNA LT Sample
Prep kit (cat. no. RS-122-2302, Illumina, Inc.) according to the
manufacturer's protocol. Transcriptome sequencing and analysis were
conducted by Shanghai OE Biotech Co., Ltd. (Shanghai, China)
http://oebiotech.bioon.com.cn/. The
libraries were sequenced on an Illumina Novaseq platform (Illumina,
Inc.) where 150 bp paired-end reads were generated. Raw data (raw
reads) of fastq format were firstly processed using in-house perl
scripts and the low-quality reads were removed to obtain the clean
reads. The clean reads were subsequently mapped to the human genome
(GRCh38) using Hierarchical Indexing for Spliced Alignment of
Transcripts (Hisat2 v2.0.5.). Fragments per kilobase per million
reads sequenced of each gene was calculated using StringTie, after
which the read counts of each gene were obtained using
featureCounts v1.5.0-p3. Differential expression analysis was
performed using the DESeq2 R package (1.16.1). P<0.05 and a
fold-change >2 or <0.5 were set as the thresholds for
significant differential expression. Hierarchical cluster analysis
of differentially expressed genes (DEGs) was performed to
demonstrate the expression pattern of genes in different groups and
samples. Kyoto Encyclopedia of Genes and Genomes pathway enrichment
analysis of DEGs were performed respectively using the
clusterProfiler R package sed on the hypergeometric distribution.
The Gene Expression Omnibus (GEO) number was GSE138621.
Statistical analysis
All the data are presented as the mean ± SEM of at
least three independent experiments and were analyzed using SPSS
19.0 software (SPSS Inc.). Differences between two groups were
determined using a paired Student's t-test, and differences among
>2 groups were analyzed by one-way analysis of variance and
Tukey post hoc tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
AUF1 promotes the viability and
proliferation of skin cells
To explore the role of AUF1 in human skin cells,
AUF1 was overexpressed or silenced using an adenovirus or a
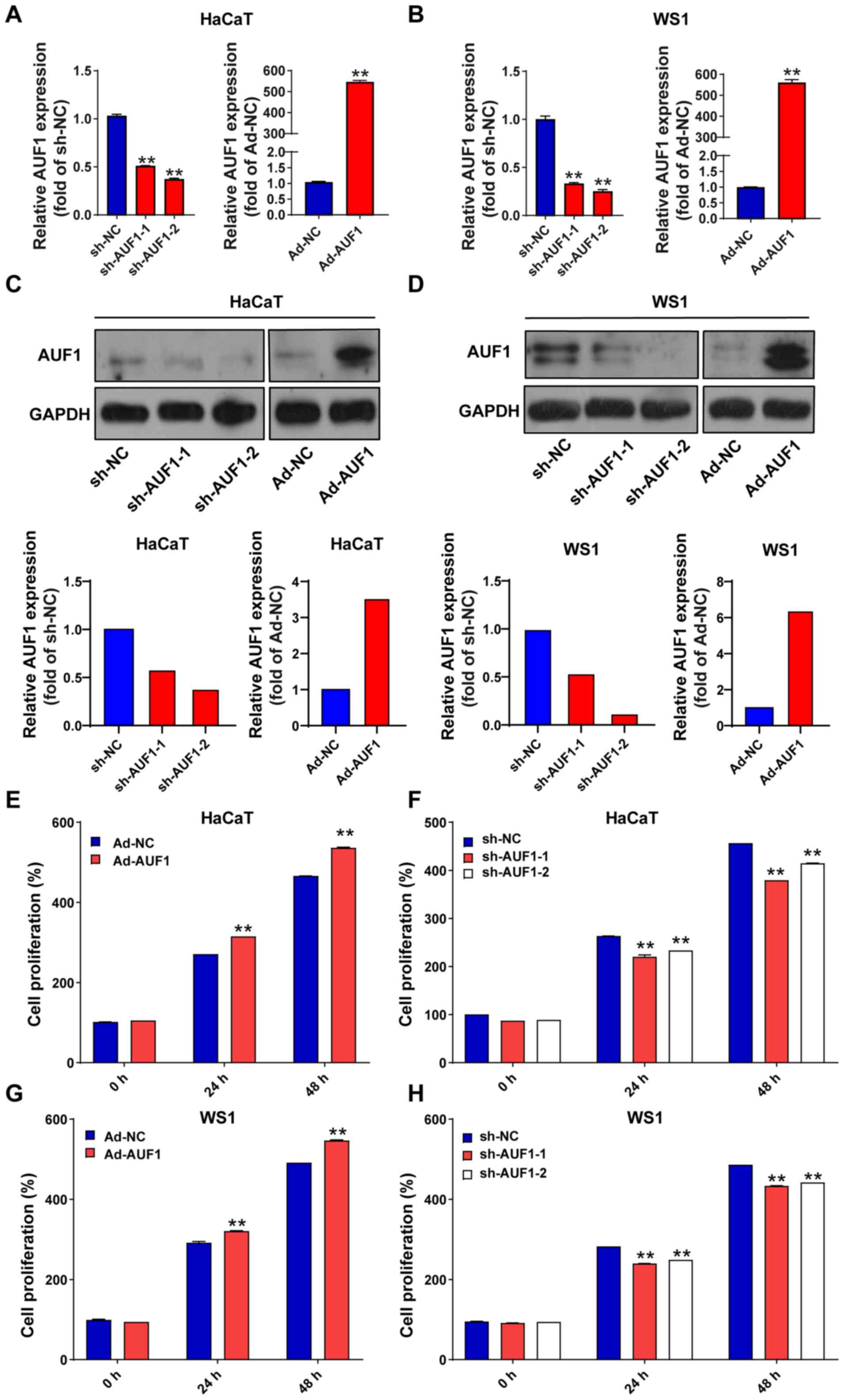
lentivirus, respectively. The efficiency of infection in HaCaT and
WS1 cells was verified by RT-qPCR (Fig. 1A and B) and western blotting analysis (Fig. 1C and D). Compared with the sh-NC group,
sh-AUF1-1- and sh-AUF1-2-infected cells demonstrated reduced levels
of AUF1 protein. As expected, AUF1 protein expression in the
Ad-AUF1-infected group was increased markedly As presented in
Fig. 1A-D, AUF1 expression was
successfully modulated by virus infection. Subsequently, cell
viability was assayed using CCK-8-based analysis. As presented in
Fig. 1E-H, AUF1 overexpression
significantly increased cell viability compared with Ad-NC at 24
and 48 h; whereas AUF1 silencing (sh-AUF1-1 and sh-AUF1-2)
significantly suppressed cell viability in HaCaT and WS1 cells
compared with sh-NC at 24 and 48 h.
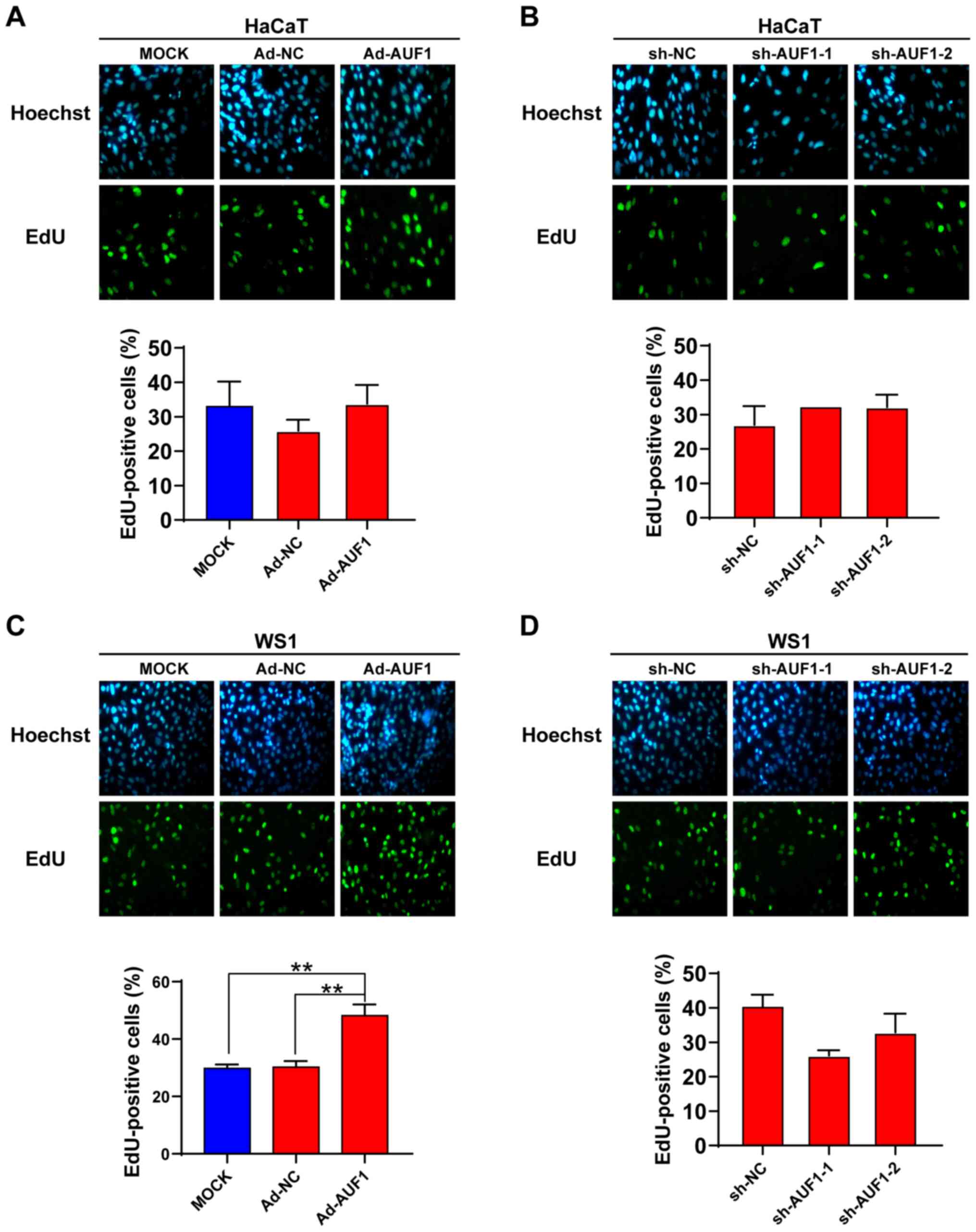
To explore the role of AUF1 in cell proliferation of
skin cells, EdU staining and colony formation assays were
performed. As presented in Fig. 2A
and C, WS1 cells in the AUF1
overexpression group revealed a significantly increased EdU
staining percentage compared with that in the control groups, while
no significant difference was observed in HaCaT cells. Knockdown of
AUF1 did not show a significant effect on the EdU-positive cell
percentage in the two types of skin cells (Fig. 2B and D).
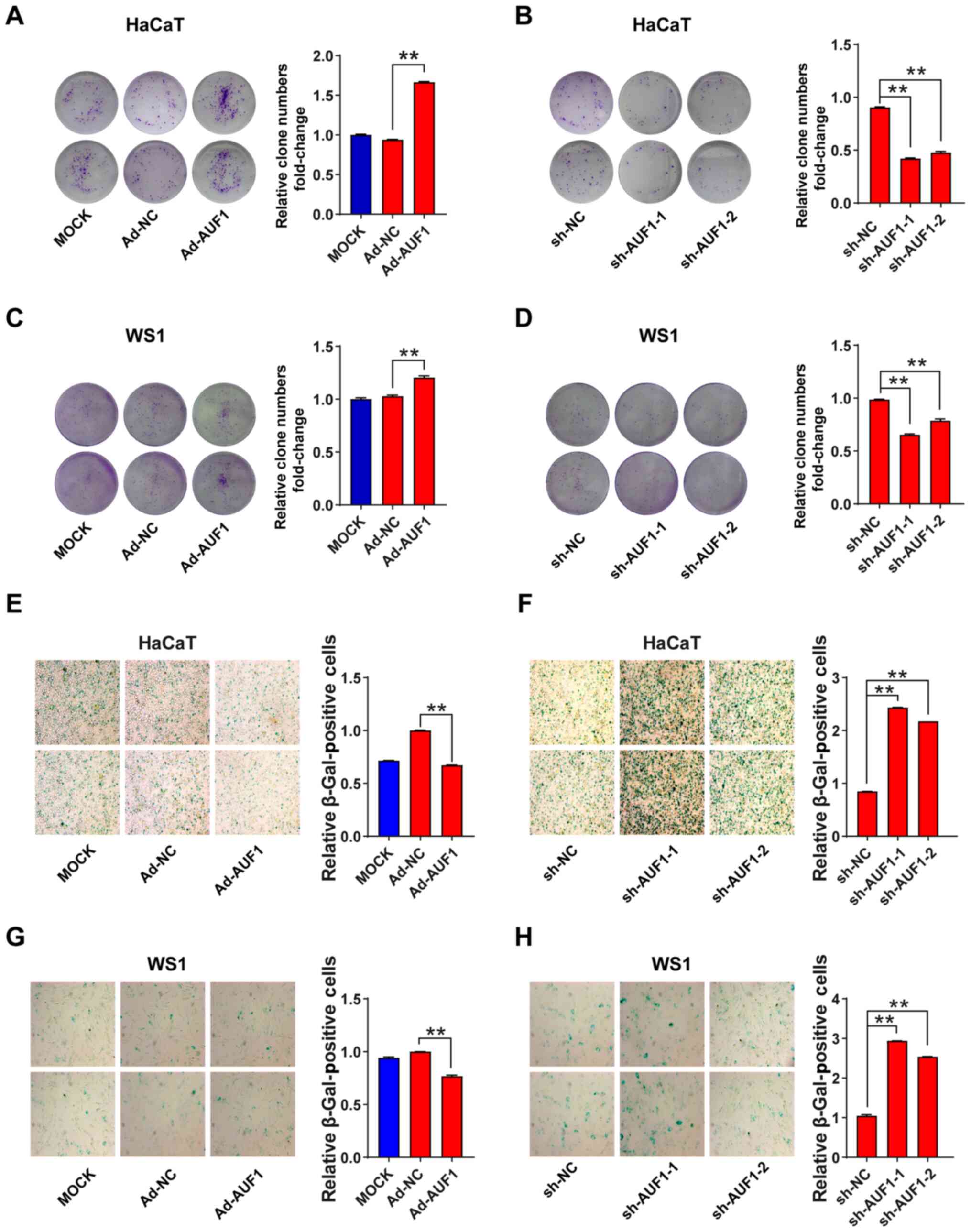
Next, the role of AUF1 in skin cell colony formation
was investigated. Overexpression of AUF1 resulted in a significant
increase of HaCaT and WS1 colony foci compared with Ad-NC (Fig. 3A and C). Conversely, infection with the two
AUF1-silencing lentiviruses caused a significant decrease in the
number of colonies in both HaCaT cells and WS1 cells compared with
sh-NC (Fig. 3B and D). Colony formation analysis consistently
demonstrated that AUF1 expression promoted cell proliferation.
AUF1 enhances the migration of skin
cells
A scratch-wound assay was used to evaluate cell
migration, which is important for wound healing. As presented in
Fig. 4A and C, when compared with mock or Ad-NC group,
the migration of HaCaT and WS1 cells was significantly increased
with AUF1 overexpression at 24 and 48 h. By contrast, when compared
with the sh-NC group, fewer AUF1-silenced HaCaT and WS1 cells
migrated toward the center of the gap of the cells, leading to a
decreased rate of wound healing at 24 and 48 h (Fig. 4B and D). Migration inhibition induced by AUF1
depletion was observed even after incubation for 48 h. Overall,
these results demonstrated that AUF1 expression can affect the
migration of skin cells.
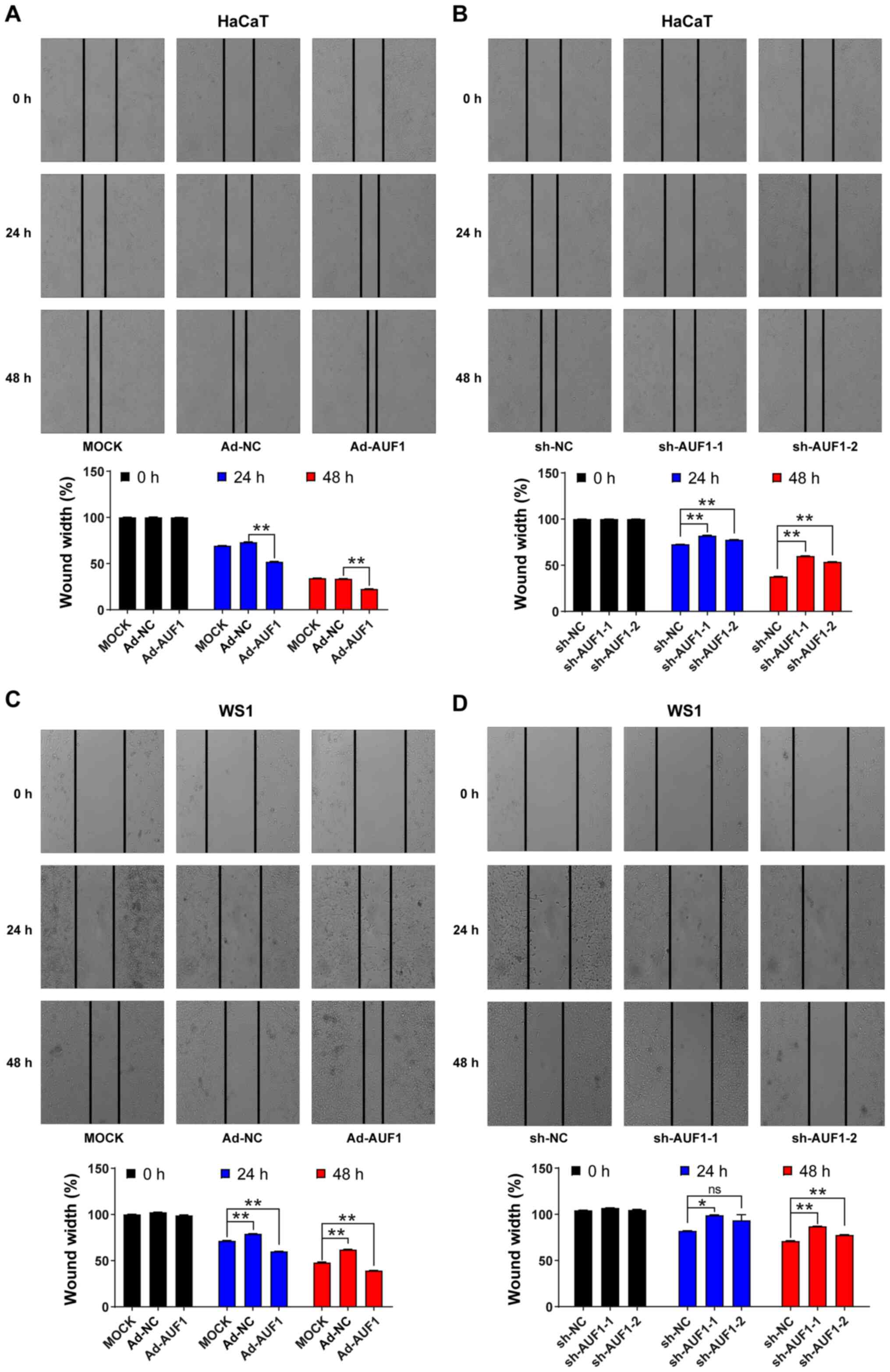 | Figure 4Effect of AUF1 on skin cell
migration. HaCaT or WS1 cell migration after infection with the
indicated viruses. Images of the scratched region were captured at
0, 24 and 48 h after the wounds were created, and the relative
wound width rate was calculated by dividing the distance of the
scratched region by the initial distance. HaCaT cells infected with
(A) Ad-AUF1, Ad-NC or the mock, or (B) sh-NC or AUF1-targeted
lentiviruses (shRNA-1 and shRNA-2). WS1 cells infected with (C)
Ad-AUF1, Ad-NC or the mock, or (D) with sh-NC or AUF1-targeted
lentiviruses (shRNA-1 and shRNA-2). *P<0.05,
**P<0.01. AUF1, AU-rich element RNA-binding factor 1;
sh, short hairpin; NC, negative control; ns, not significant. |
AUF1 attenuates cell senescence in
skin cells
Cell senescence was analyzed using β-galactosidase
staining, and the relative numbers of β-galactosidase-positive skin
cells were identified. In the AUF1 overexpression group of skin
cells, there was a significant decrease in the number of
β-galactosidase-positive cells following infection with
AUF1-overexpressing adenovirus compared with the Ad-NC groups
(Fig. 3E and G). The positive areas were significantly
reduced by Ad-AUF1 infection in HaCaT and WS1 cells (Fig. 3E and G). Conversely, as presented in Fig. 3F and H, knocking down AUF1 nearly tripled the
β-galactosidase staining-positive areas in the two shRNA
lentivirus-infected HaCaT cells and WS1 cells compared with the
sh-NC. These results reflected the positive function of AUF1 in
cell senescence.
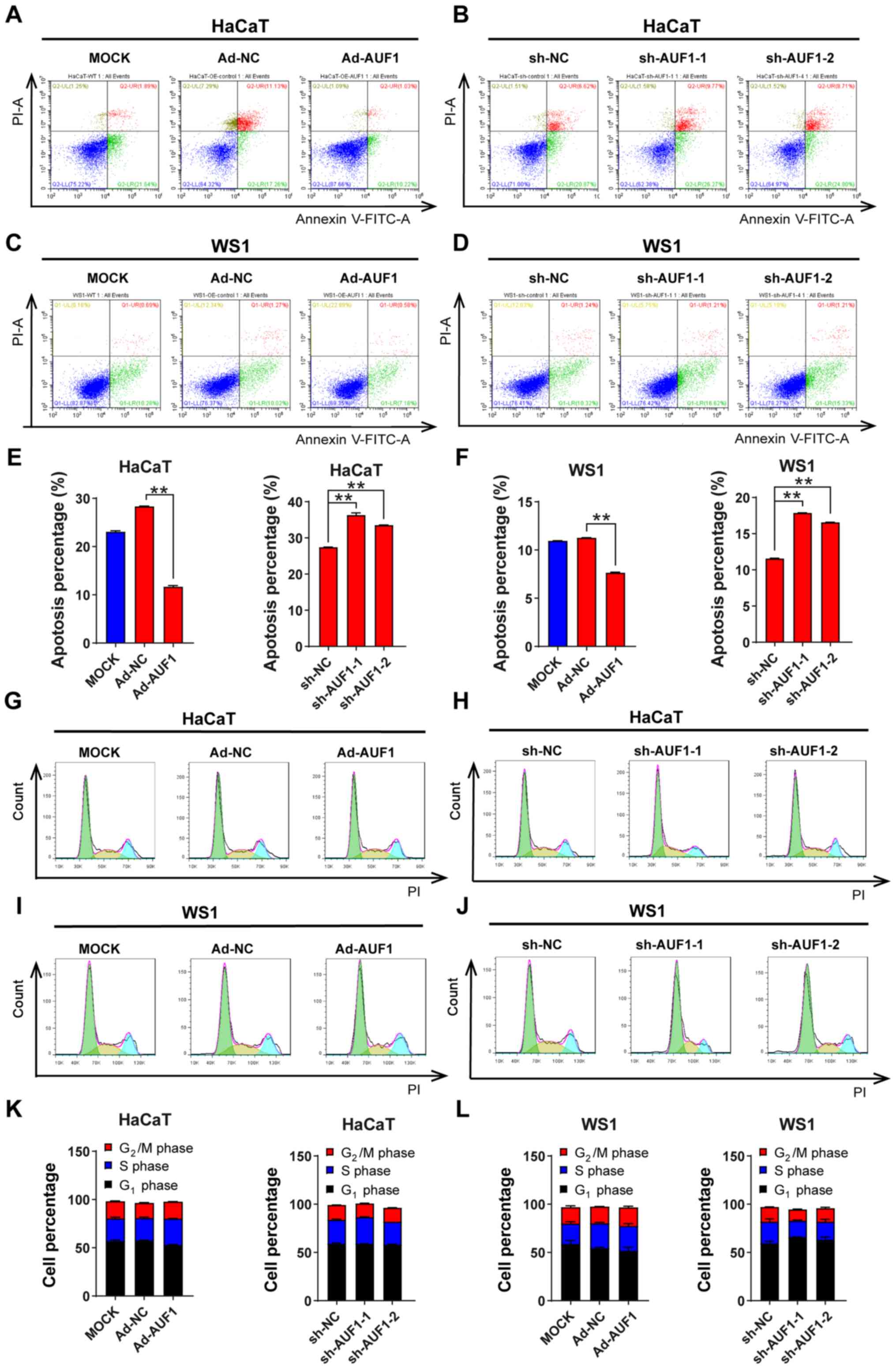
AUF1 decreases apoptosis in skin
cells
As cell proliferation was affected by AUF1
expression, the function of AUF1 expression on apoptosis was
investigated using an Annexin V/PI double-staining assay. Due to
the pretreatment time before cytometry analysis, there were higher
basal levels of apoptosis in HaCaT cells. The Ad-AUF1 groups in
both HaCaT and WS1 cells demonstrated significantly reduced
apoptosis rates compared with the control groups; by contrast, the
apoptosis rates of AUF1-downregulated skin cells were significantly
increased compared with the controls (Fig. 5A-F). These results indicated the
antiapoptotic role of AUF1 in skin cells.
AUF1 has no significant effect on cell
cycle progression
AUF1 may play a role in cell proliferation by
disturbing the cell cycle; therefore, a cell cycle analysis was
conducted using flow cytometry. However, no statistical
significance for the percentage of cells in different phases was
observed between any infected group (either the AUF1 overexpression
group or AUF1 knockdown groups) and the corresponding control group
in the two types of cells (Fig.
5G-J). As presented in Fig.
5K-L, AUF1 did not modulate cell cycle progression in HaCaT and
WS1 cells.
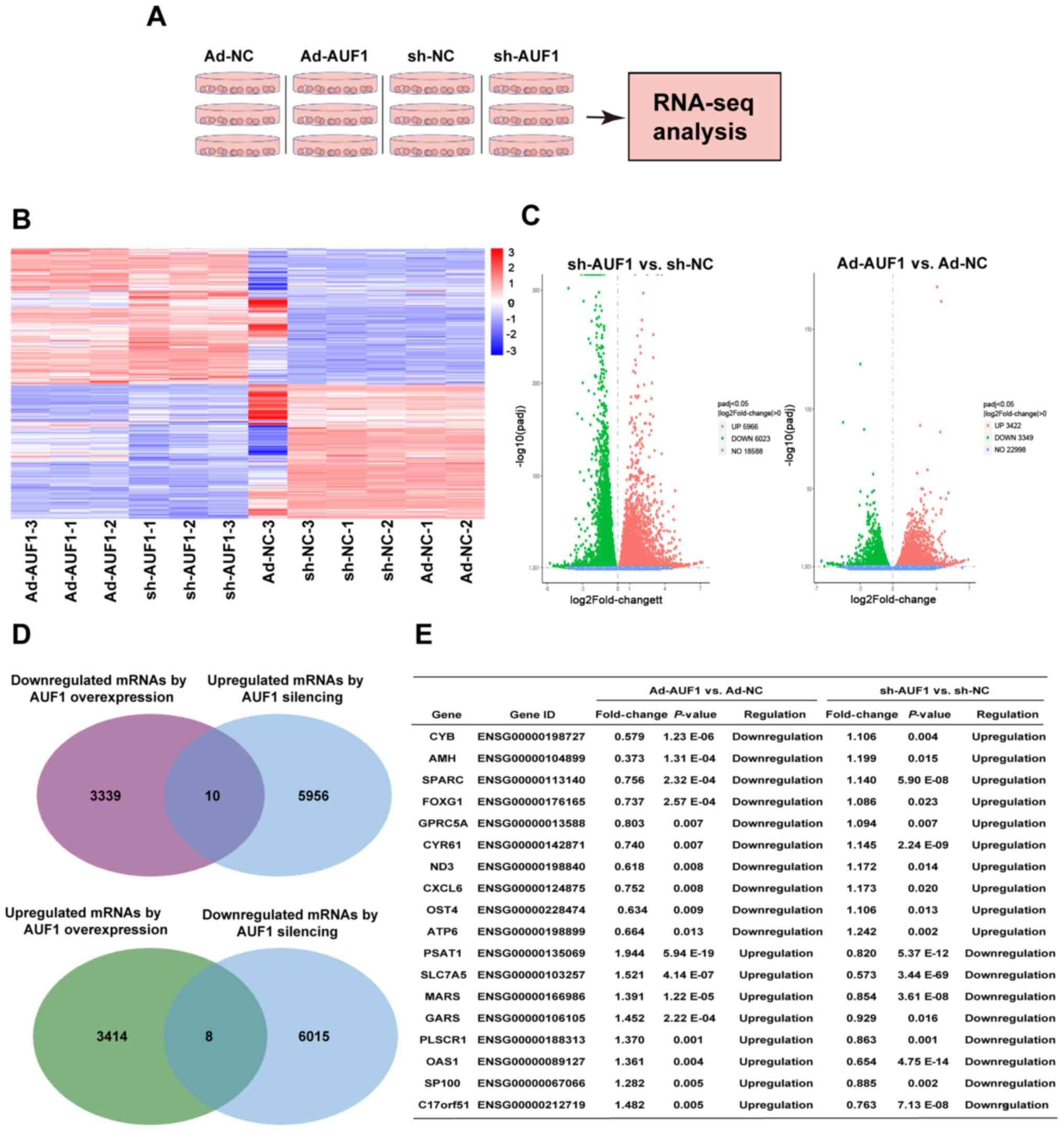
Transcripts and pathways that are
affected by AUF1
As AUF1 is an RBP, multiple transcripts and pathways
may be affected by the action of AUF1. To explore AUF1-affected
mRNAs, WS1 cells with AUF1 overexpression and silencing were used
to perform RNA-Seq and KEGG pathway analysis (Fig. 6A). The raw data are accessible at
GSE138621. The results indicated that knockdown of AUF1 influenced
>10,000 mRNAs, while overexpression of AUF1 caused a significant
expression change in 6,771 mRNAs (Fig.
6B-D). A total of 6,771 mRNAs differentially expressed in
AUF1-overexpressing cells included 3,422 up- and 3,349
downregulated mRNAs. AUF1 silencing involved 5,966 up- and 6,023
downregulated transcripts (Fig.
6D). A total of 18 mRNAs (eight mRNAs with positive
associations and 10 mRNAs with negative associations) demonstrated
consistent associations with both AUF1 overexpression and silencing
(Fig. 6D): Mitochondrially encoded
cytochrome B, anti-muellerian hormone (AMH), secreted protein
acidic and cysteine rich (SPARC), forkhead box G1 (FOXG1), G
protein-coupled receptor class c group 5 member A, cysteine rich
angiogenic inducer 61 (CYR61), NADH-ubiquinone oxidoreductase chain
3, C-X-C motif chemokine ligand 6, oligosaccharyltransferase
complex subunit 4, non-catalytic, mitochondrially encoded ATP
synthase membrane subunit 6, phosphoserine aminotransferase 1,
solute carrier family 7 member 5, methionyl-TRNA synthetase 1
(MARS), glycyl-TRNA synthetase 1, phospholipid scramblase 1
(PLSCR1), 2'-5'-oligoadenylate synthetase 1, nuclear autoantigen
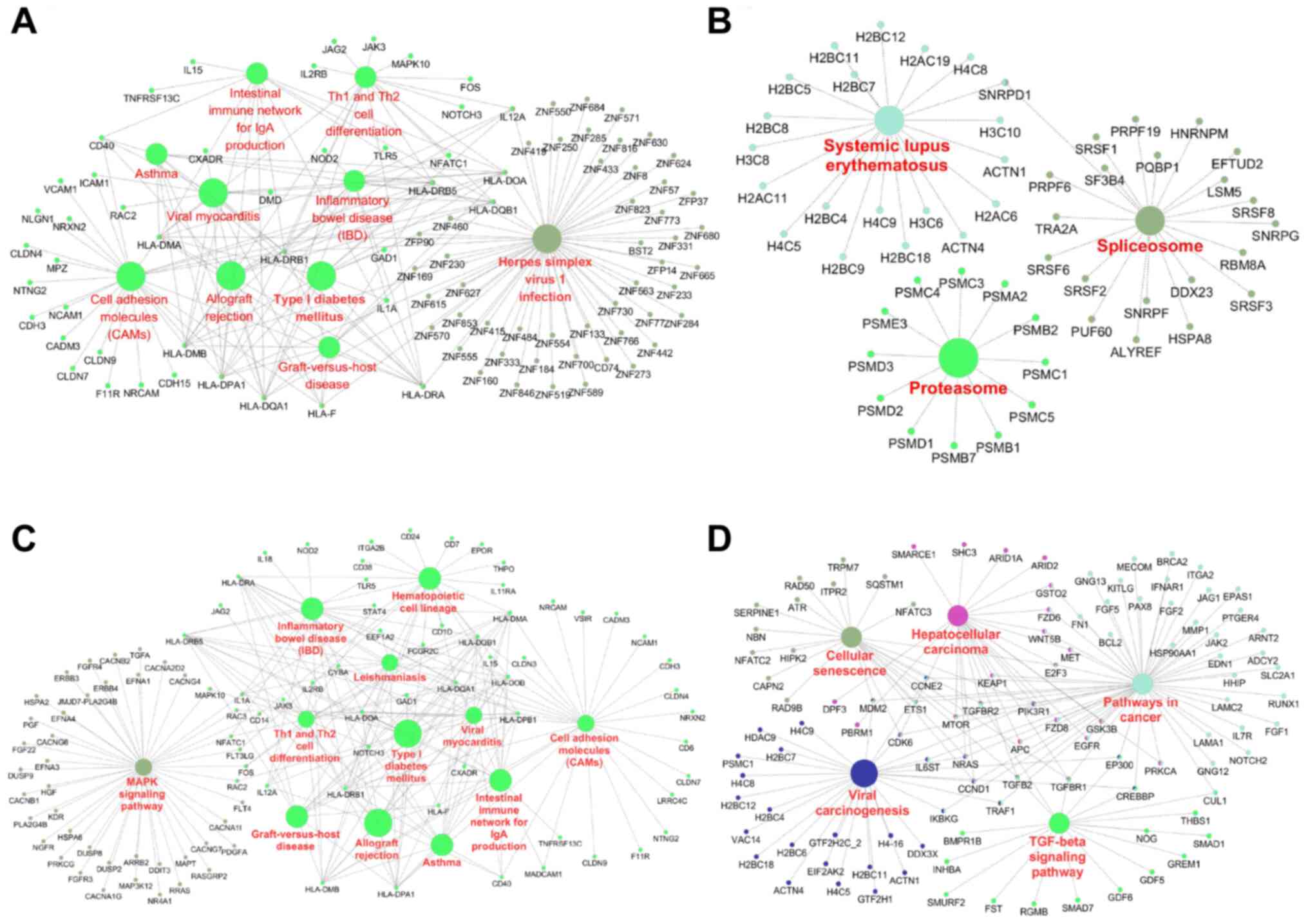
Sp-100 (SP100), chromosome 17 open reading frame 51 (Fig. 6E). KEGG pathway analysis revealed
AUF1-affected networks involved multiple biological processes,
including cell adhesion molecules, the proteasome and the
spliceosome (Fig. 7A and B). mRNAs downregulated by AUF1 were
associated with cell adhesion molecules, pathways in cancer,
cellular senescence and the TGF-β signaling pathway, among others
(Fig. 7C and D). These results suggested a complex
regulatory network of AUF1 in skin cells.
Discussion
AUF1 regulates various cell processes, such as cell
proliferation, apoptosis, cell cycle progression and/or senescence,
by interacting with adenylate-uridylate (AU)-rich element-bearing
mRNAs (7,11,16,17).
The functional significance of AUF1 has been explored in various
types of cells, including tumor cells and noncancerous cells
(7-11,16-18).
However, to the best of our knowledge, the role of AUF1 in skin
cells has not been reported, even in skin diseases.
The present study aimed to explore whether AUF1
played a notable role in the homeostasis of skin cells in
vitro. It was revealed that, when WS1 cells were infected with
different types of viruses, there were expected changes in AUF1
expression. These included AUF1 downregulation when exposed to AUF1
silencing viruses and AUF1 overexpression following Ad-AUF1
infection. Generally, overexpression of AUF1 promoted the
proliferation and migration of HaCaT and WS1 cells, while knockdown
of AUF1 demonstrated the opposite effects. Although HaCaT and WS1
cells are non-cancerous cells, they are immortalized by the
American Type Culture Collection. A previous report indicated that
HaCaT cells have various capacities for cloning (19). The present study used colony
formation assays to confirm the pro-proliferative role of AUF1, as
AUF1 has been reported to stimulate cell proliferation in human
chronic myeloid leukemia K562 cells (20). Consistently, downregulation of AUF1
expression has been reported to reduce cell proliferation in ESCC
cells and human colorectal carcinoma HCT116 cells (7,16).
However, a limitation of the present study was that an in
vivo study based on animal models was not performed. In
vitro results may be different to those of cells in vivo
and primary skin cells. When AUF1 is continuously overexpressed,
the cells may be regulated by other mechanisms to control cell
proliferation as AUF1 is constitutively expressed in normal skin
tissues (21). However, the
difference between in vitro and in vivo conditions
warrants further investigation. In vivo experiments will be
considered for improved evaluation of the role of AUF1 in skin
cells in follow-up studies. Regarding the effect on cell migration,
AUF1 served as a promoter in the present study, which was
consistent with a previous study which demonstrated that AUF1
expression and wound healing were associated (18). Similarly, AUF1 overexpression
contributes to cell migration (22). However, AUF1 knockdown in B16 mouse
melanoma cells is beneficial to cell migration (23).
The present study them examined whether AUF1 was
associated with apoptosis, senescence and cell cycle alteration.
Apoptosis was significantly decreased with AUF1 overexpression, but
was enhanced by AUF1 knockdown in HaCaT and WS1 cells. Cell
senescence was affected in a similar pattern as apoptosis, whereas
cell cycle changes were not significant after the cells were
treated with the different viruses. Apoptosis and senescence
reflected the cell proliferation ability and cell motility
modulated by AUF1, which is consistent with observations in a
report involving ESCC cells (7).
This indicates that silencing AUF1 inhibits the proliferation and
enhances the apoptosis of ESCC cells (7). AUF1 has been suggested to have an
inhibitory effect on apoptosis, and upregulated AUF1 contributes to
uncontrolled proliferation of liver cells (10). Previous studies have demonstrated
that AUF1 suppresses cellular senescence, and plays roles in
anti-aging and maintaining cell proliferation (11,24).
However, the cell cycle was not affected by AUF1 in the present
study, although AUF1 has been considered to control the mRNA decay
of cell cycle-regulatory proteins (6,25).
It was hypothesized that the pro-proliferative role of AUF1 was
likely attributed to decreased apoptosis and senescence. In
addition, a number of examples demonstrate that cell proliferation
is associated with cell death, meaning that changes of the cell
cycle are not essential (26-30).
Thus, a more sophisticated mechanism other compared with the cell
cycle was implicated in AUF1 modulation of skin cells.
Diverse targets of AUF1 have been explored to
elucidate the mechanism of AUF1 in different cell types, such as
GTP cyclohydrolase 1 for cell proliferation in ESCC cells, neural
precursor cell expressed developmentally downregulated 4-like in
HK2 cells and myocyte-specific enhancer factor 2C for the
myogenesis program in mouse myoblasts (1,18,26,31).
To explore the underlying mechanism of AUF1 in skin cells, the
present study performed RNA-Seq, which revealed the landscape of
the transcriptome in skin cells with up/downregulated AUF1
expression. Thousands of genes demonstrated differential expression
following AUF1 upregulation or downregulation, and hundreds of
pathways were subsequently annotated with KEGG pathway functions.
By integrating RNA-Seq data from skin cells with AUF1 upregulation
and downregulation, 18 mRNAs (eight mRNAs with positive
associations and 10 mRNAs with negative associations) revealed
consistent associations with AUF1 expression levels. Among these 18
potentially regulated AUF1 molecules, several have been
demonstrated to be associated with cell function. For example, PAST
homolog 1 contributes to cell proliferation and cell cycle
progression (32,33). Reduced expression of 2-5A synthase
1 by a non-coding RNA named TINCR facilitates proliferation in
breast cancer cells (34). MARS
has an important role in initiating translation and protection
against cellular damage (35).
MARS has been revealed to promote the proliferation of cancer cells
(36). Taken together, AUF1
exerted its influence through wide-spectrum targets, which was
consistent with its behavior in other types of cell, such as ESCC
cells, human colorectal carcinoma HCT116 cells and human chronic
myeloid leukemia K562 cells (7,16,21).
However, the targets of AUF1 are likely to be cell-type specific.
Querying published papers revealed a number of genes that could be
the exact targets for AUF1 that lead to cell proliferation,
including AMH, SPARC, FOXG1, CYR61, PLSCR1 and SP100 (37-42).
These genes were explored by RNA-Seq in the present study. Among
them, CYR61 (CCN1) and SP100 are associated with apoptosis, and
FOXG1 is connected with cell senescence (43-45).
However, which one could be the exact target for AUF1 that leads to
cell proliferation needs to be further studied.
In conclusion, AUF1 played a positive role in the
proliferation and migration of immortalized skin cells. AUF1
inhibited cell senescence and apoptosis in human skin cells and
modulated downstream mRNAs implicated in multiple pathways in skin
cells.
Supplementary Material
Targeting sequences in AU-rich element
RNA binding protein 1.
Primer sequences for reverse
transcription quantitative PCR analysis.
Acknowledgements
Not applicable.
Funding
Funding: This work is supported by the National Natural Science
Foundation of China (grant nos. 31770911, 81803166, 32071238 and
82073477), Military Logistics Research Program (grant no.
BKJ18J003), the Young Talent Project of China National Nuclear
Corporation and the Fundamental Research Funds for the Central
Universities.
Availability of data and materials
The datasets generated and/or analyzed during the
current study are available in the Gene Expression Omnibus
repository, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE138621.
Authors' contributions
DY and XL conceived the experiments and designed the
research. DY, XL and ZW performed the molecular biology
experiments. SJ, TY and ZJ analyzed and interpreted the data. SZ,
YS and KF performed statistical analysis. SZ drafted the
manuscript. KF performed language editing of the manuscript. YS
provided general supervision. ZJ and KF confirm the authenticity of
all the raw data. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stavast CJ and Erkeland SJ: The
Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation.
Cells. 8(1465)2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Loflin P, Chen CY and Shyu AB: Unraveling
a cytoplasmic role for hnRNP D in the in vivo mRNA destabilization
directed by the AU-rich element. Genes Dev. 13:1884–1897.
1999.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhang W, Wagner BJ, Ehrenman K, Schaefer
AW, DeMaria CT, Crater D, DeHaven K, Long L and Brewer G:
Purification, characterization, and cDNA cloning of an AU-rich
element RNA-binding protein, AUF1. Mol Cell Biol. 13:7652–7665.
1993.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Wagner BJ, DeMaria CT, Sun Y, Wilson GM
and Brewer G: Structure and genomic organization of the human AUF1
gene: Alternative pre-mRNA splicing generates four protein
isoforms. Genomics. 48:195–202. 1998.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Zucconi BE and Wilson GM: Modulation of
neoplastic gene regulatory pathways by the RNA-binding factor AUF1.
Front Biosci (Landmark Ed). 16:2307–2325. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Lozano-Rosas MG, Chávez E, Velasco-Loyden
G, Domínguez-López M, Martínez-Pérez L and Chagoya De Sánchez V:
Diminished S-adenosylmethionine biosynthesis and its metabolism in
a model of hepatocellular carcinoma is recuperated by an adenosine
derivative. Cancer Biol Ther. 21:81–94. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Gao Y, Wang W, Cao J, Wang F, Geng Y, Cao
J, Xu X, Zhou J, Liu P and Zhang S: Upregulation of AUF1 is
involved in the proliferation of esophageal squamous cell carcinoma
through GCH1. Int J Oncol. 49:2001–2010. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Sarkar S, Sinsimer KS, Foster RL, Brewer G
and Pestka S: AUF1 isoform-specific regulation of anti-inflammatory
IL10 expression in monocytes. J Interferon Cytokine Res.
28:679–691. 2008.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Trojanowicz B, Sekulla C, Dralle H and
Hoang-Vu C: Expression of ARE-binding proteins AUF1 and HuR in
follicular adenoma and carcinoma of thyroid gland. Neoplasma.
63:371–377. 2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wu X, Yang Y, Huang Y, Chen Y, Wang T, Wu
S, Tong L, Wang Y, Lin L, Hao M, et al: RNA-binding protein AUF1
suppresses miR-122 biogenesis by down-regulating Dicer1 in
hepatocellular carcinoma. Oncotarget. 9:14815–14827.
2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
He J, Jiang YF, Liang L, Wang DJ, Wei WX,
Ji PP, Huang YC, Song H, Lu XL and Zhao YX: Targeting of AUF1 to
vascular endothelial cells as a novel anti-aging therapy. J Geriatr
Cardiol. 14:515–523. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang S, Xue J, Zheng J, Wang S, Zhou J,
Jiao Y, Geng Y, Wu J, Hannafon BN and Ding WQ: The superoxide
dismutase 1 3'UTR maintains high expression of the SOD1 gene in
cancer cells: The involvement of the RNA-binding protein AUF-1.
Free Radic Biol Med. 85:33–44. 2015.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gund R, Zirmire R, J H, Kansagara G and
Jamora C: Histological and Immunohistochemical Examination of Stem
Cell Proliferation and Reepithelialization in the Wounded Skin. Bio
Protoc. 11(e3894)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Sadri N and Schneider RJ:
Auf1/Hnrnpd-deficient mice develop pruritic inflammatory skin
disease. J Invest Dermatol. 129:657–670. 2009.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ma W, Qiao J, Zhou J, Gu L and Deng D:
Characterization of novel LncRNA P14AS as a protector of ANRIL
through AUF1 binding in human cells. Mol Cancer.
19(42)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yan J, Du F, Li SD, Yuan Y, Jiang JY, Li
S, Li XY and Du ZX: AUF1 modulates TGF-β signal in renal tubular
epithelial cells via post-transcriptional regulation of Nedd4L
expression. Biochim Biophys Acta Mol Cell Res. 1865:48–56.
2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Al-Khalaf HH and Aboussekhra A: AUF1
positively controls angiogenesis through mRNA
stabilization-dependent up-regulation of HIF-1α and VEGF-A in human
osteosarcoma. Oncotarget. 10:4868–4879. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Tate S, Imai M, Matsushita N, Nishimura
EK, Higashiyama S and Nanba D: Rotation is the primary motion of
paired human epidermal keratinocytes. J Dermatol Sci. 79:194–202.
2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Lu M, Pan C, Zhang L, Ding C, Chen F, Wang
Q, Wang K and Zhang X: ING4 inhibits the translation of
proto-oncogene MYC by interacting with AUF1. FEBS Lett.
587:1597–1604. 2013.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Moore AE, Chenette DM, Larkin LC and
Schneider RJ: Physiological networks and disease functions of
RNA-binding protein AUF1. Wiley Interdiscip Rev RNA. 5:549–564.
2014.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Qian W, Cai X, Qian Q, Wang D and Zhang L:
Angelica sinensis polysaccharide suppresses
epithelial-mesenchymal transition and pulmonary fibrosis via a
DANCR/AUF-1/FOXO3 regulatory axis. Aging Dis. 11:17–30.
2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Sun S, Zhang X, Lyu L, Li X, Yao S and
Zhang J: Autotaxin expression is regulated at the
post-transcriptional level by the RNA-binding proteins HuR and
AUF1. J Biol Chem. 291:25823–25836. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Lee JW, Chun YL, Kim AY, Lloyd LT, Ko S,
Yoon JH and Min KW: Accumulation of mitochondrial RPPH1 RNA
is associated with cellular senescence. Int J Mol Sci Jan.
22(782)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Panda AC, Abdelmohsen K, Yoon JH,
Martindale JL, Yang X, Curtis J, Mercken EM, Chenette DM, Zhang Y,
Schneider RJ, et al: RNA-binding protein AUF1 promotes myogenesis
by regulating MEF2C expression levels. Mol Cell Biol. 34:3106–3119.
2014.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Xu Y, Jiang Y, Wang Y, Ren Y, Zhao Z, Wang
T and Li T: LINC00473 regulated apoptosis, proliferation and
migration but could not reverse cell cycle arrest of human bone
marrow mesenchymal stem cells induced by a high-dosage of
dexamethasone. Stem Cell Res (Amst). 48(101954)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Alimirah F, Pulido T, Valdovinos A,
Alptekin S, Chang E, Jones E, Diaz DA, Flores J, Velarde MC,
Demaria M, et al: Cellular senescence promotes skin carcinogenesis
through p38MAPK and p44/42MAPK signaling. Cancer Res. 80:3606–3619.
2020.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Walczak K, Langner E, Makuch-Kocka A,
Szelest M, Szalast K, Marciniak S and Plech T: Effect of
Tryptophan-Derived AhR Ligands, Kynurenine, Kynurenic Acid and
FICZ, on Proliferation, Cell Cycle Regulation and Cell Death of
Melanoma Cells-In Vitro Studies. Int J Mol Sci.
21(7946)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Lin Q, Jin HJ, Zhang D and Gao L: DDX46
silencing inhibits cell proliferation by activating apoptosis and
autophagy in cutaneous squamous cell carcinoma. Mol Med Rep.
22:4236–4242. 2020.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu Q, Dong J, Li J, Duan Y, Wang K, Kong
Q and Zhang H: LINC01255 combined with BMI1 to regulate human
mesenchymal stromal senescence and acute myeloid leukemia cell
proliferation through repressing transcription of MCP-1. Clin
Transl Oncol. 23:1105–1116. 2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
White EJ, Brewer G and Wilson GM:
Post-transcriptional control of gene expression by AUF1:
Mechanisms, physiological targets, and regulation. Biochim Biophys
Acta. 1829:680–688. 2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yang Y, Wu J, Cai J, He Z, Yuan J, Zhu X,
Li Y, Li M and Guan H: PSAT1 regulates cyclin D1 degradation and
sustains proliferation of non-small cell lung cancer cells. Int J
Cancer. 136:E39–E50. 2015.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Duan W and Liu X: PSAT1 upregulation
contributes to cell growth and cisplatin resistance in cervical
cancer cells via regulating PI3K/AKT signaling pathway. Ann Clin
Lab Sci. 50:512–518. 2020.PubMed/NCBI
|
|
34
|
Lu D, Di S, Zhuo S, Zhou L, Bai R, Ma T,
Zou Z, Chen C, Sun M, Tang J, et al: The long noncoding RNA TINCR
promotes breast cancer cell proliferation and migration by
regulating OAS1. Cell Death Discov. 7(41)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Suh YS, Yeom E, Nam JW, Min KJ, Lee J and
Yu K: Methionyl-tRNA synthetase regulates lifespan in
Drosophila. Mol Cells. 43:304–311. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Jin Q, Liu G, Wang B, Li S, Ni K, Wang C,
Ren J, Zhang S and Dai Y: High methionyl-tRNA synthetase expression
predicts poor prognosis in patients with breast cancer. J Clin
Pathol. 73:803–812. 2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Wu MF, Stachon T, Langenbucher A, Seitz B
and Szentmáry N: Effect of amniotic membrane suspension (AMS) and
amniotic membrane homogenate (AMH) on human corneal epithelial cell
viability, migration and proliferation in vitro. Curr Eye Res.
42:351–357. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
PLOS ONE Editors. Retraction: SPARC
overexpression inhibits cell proliferation in neuroblastoma and is
partly mediated by tumor suppressor protein PTEN and AKT. PLoS One.
15(e0228246)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhen J, Zhang H, Dong H and Tong X:
miR-9-3p inhibits glioma cell proliferation and apoptosis by
directly targeting FOXG1. Oncol Lett. 20:2007–2015. 2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Cheng Z, Zhang Y, Tian Y, Chen Y, Ding F,
Wu H, Ji Y and Shen M: Cyr61 promotes Schwann cell proliferation
and migration via αvβ3 integrin. BMC Mol Cell Biol.
22(21)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Gui L, Zhu YW, Xu Q, Huang JJ, Hua P, Wu
GJ, Lu J, Ni JB, Tang H and Zhang LL: RNA interference-mediated
downregulation of phospholipid scramblase 1 expression in primary
liver cancer in vitro. Oncol Lett. 20(361)2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Held-Feindt J, Hattermann K,
Knerlich-Lukoschus F, Mehdorn HM and Mentlein R: SP100 reduces
malignancy of human glioma cells. Int J Oncol. 38:1023–1030.
2011.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Dang T, Modak C, Meng X, Wu J, Narvaez R
and Chai J: CCN1 induces apoptosis in esophageal adenocarcinoma
through p53-dependent downregulation of survivin. J Cell Biochem.
120:2070–2077. 2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Wang R, Li KM, Zhou CH, Xue JL, Ji CN and
Chen JZ: Cdc20 mediates D-box-dependent degradation of Sp100.
Biochem Biophys Res Commun. 415:702–706. 2011.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Verginelli F, Perin A, Dali R, Fung KH, Lo
R, Longatti P, Guiot MC, Del Maestro RF, Rossi S, di Porzio U, et
al: Transcription factors FOXG1 and Groucho/TLE promote
glioblastoma growth. Nat Commun. 4(2956)2013.PubMed/NCBI View Article : Google Scholar
|