Introduction
Diabetic nephropathy (DN) is a serious and common
complication of type 1 and 2 diabetes (1-3).
DN can eventually lead to end-stage renal disease in ≤40% patients
with diabetes (1-3).
It has been previously reported that DN can be considered as the
strongest predictor of mortality (4,5).
Advanced DN is characterized by glomerular sclerosis,
tubulointerstitial degeneration and fibrosis, which may result in a
precipitous decline in the glomerular filtration rate, resulting in
proteinuria (6-8).
Previous studies have shown that podocyte damage can trigger a
number of pathological changes, including glomerular sclerosis and
renal failure (9,10). Furthermore, it has been reported
that podocyte damage is closely associated with the pathogenesis of
several kidney-related diseases, including focal segmental
glomerulosclerosis, minimal change nephropathy and DN (11,12).
Therefore, preventing podocyte injury should be considered to be an
effective strategy for treating DN.
It has been well established that the excessive
production of inflammatory cytokines, reactive oxygen species (ROS)
and lactate dehydrogenase (LDH) release can be induced under high
glucose (HG) conditions, which may lead to podocyte depletion
(13,14). Emerging evidence has suggested that
active components contained within certain Traditional Chinese
Medicine exert suppressive effects on HG-induced inflammation and
oxidative stress (14-16).
Gastrodin is one of the primary components in the rhizome of the
saprophytic perennial herb Gastrodia elata (17). A previous study demonstrated that
gastrodin can exert anti-inflammatory and antioxidative effects on
neurodegenerative diseases including Alzheimer's disease,
Parkinson's disease and cerebral ischemia/reperfusion (18). Another study revealed that gastrodin
treatment conferred neuroprotective effects on patients with type 2
diabetes by inhibiting endoplasmic reticulum stress and activating
the NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3)
inflammasome (19). Additionally,
gastrodin could suppress the apoptosis of human retinal endothelial
cells by blocking the sirtuin 1/Toll-like receptor 4/NF-κB p65
signaling pathway under HG conditions (20). However, to the best of our
knowledge, the role of gastrodin in DN has not been previously
investigated. Based on these aforementioned findings, the present
study hypothesized that gastrodin possesses potential therapeutic
effects against DN.
The AMP-activated protein kinase (AMPK)/nuclear
factor erythroid 2-related factor 2 (Nrf2) signaling pathway has
been reported to regulate several cellular processes, including
inflammation, oxidative stress and apoptosis (21-23).
A previous study has revealed that gastrodin ameliorated oxidative
stress and the inflammatory response whilst improving lipid
metabolism by activating the AMPK/Nrf2 pathway in nonalcoholic
fatty liver disease mouse and rat models (24). Additionally, 4-O-methylhonokiol has
been found to ameliorate type 2 diabetes-induced nephropathy in
mice by activating AMPK-mediated fatty acid oxidation and
Nrf2-mediated oxidative stress reduction (25). Therefore, it was speculated that
gastrodin may inhibit podocyte inflammation, oxidative stress and
apoptosis induced by HG through activation of the AMPK/Nrf2
pathway.
In the present study, immortalized mouse podocytes
were pretreated with different doses of gastrodin under HG
conditions to establish an in vitro model of podocyte
injury. The aim was to reveal evidence to support the potential
protective effects of gastrodin against inflammation and oxidative
stress in podocytes induced by HG.
Materials and methods
Cell culture and treatment
The mouse podocyte MPC5 cell line was obtained from
the American Type Culture Collection (26-28).
Podocytes were maintained in RPMI-1640 medium (Thermo Fisher
Scientific, Inc.) supplemented with 10% inactivated FBS (Thermo
Fisher Scientific, Inc.) and 10 U/ml mouse recombinant interferon-γ
(IFN-γ; Cell Signaling Technology, Inc.) at 33˚C in a humidified
atmosphere of 5% CO2. When they reached 80% confluence,
the podocytes were incubated in RPMI-1640 medium without IFN-γ at
37˚C for 10-14 days to induce podocyte differentiation (29).
MPC5 cells were then divided into the following
groups: i) Normal glucose (NG; 5.5 mM D-glucose); ii) mannitol (MA;
5.5 mM glucose + 24.5 mM D-MA; as osmotic control); iii) HG (30 mM
D-glucose); and iv) HG + gastrodin (Sigma-Aldrich; Merck KGaA).
MPC5 cells were pretreated with different
concentrations of gastrodin (0.1, 1, 10 and 100 µM) for 24 h prior
to HG addition (30). MPC5 cells
were also pretreated with 20 µM Compound C (AMPK inhibitor, also
known as dorsomorphin; Tocris Bioscience) for 1 h before HG
treatment (31,32) at 37˚C.
Cell Counting Kit-8 (CCK-8) assay
A CCK-8 assay kit (Beyotime Institute of
Biotechnology) was used to evaluate cell viability, according to
the manufacturer's protocols. Briefly, MPC5 cells (5x103
cells/well) were seeded into 96-well plates and treated with or
without HG and gastrodin for 24 h at 37˚C. Subsequently, 10 µl/well
of CCK-8 solution was added into each well and the cells were
incubated for an additional 2 h at 37˚C. The absorbance in each
well was measured at a wavelength of 450 nm using a microplate
reader (Thermo Fisher Scientific, Inc.).
ELISA
Following cell treatment, the secretion levels of
TNF-α, IL-1β and IL-6 in the MPC5 cell culture supernatants were
determined using corresponding ELISA kits (cat. nos. RAB0477,
RAB0274 and RAB0308, respectively; Merck KGaA), according to the
manufacturer's protocols.
Western blot analysis
Following treatment, MPC5 cells were lysed with RIPA
buffer (Beyotime Institute of Biotechnology) to extract total
proteins. Protein concentration was determined using the BCA
Protein Assay kit. After denaturation at 100˚C for 10 min, protein
samples (40 µg) from each group were loaded and separated using 10%
SDS-PAGE before they were transferred onto PVDF membranes.
Following blocking with 5% skimmed milk for 2 h at room
temperature, the membranes were incubated at 4˚C overnight with
primary antibodies (all from Abcam except cleaved caspase-6)
against monocyte chemoattractant protein 1 (MCP-1; dilution,
1:1,000; cat. no. ab214819), NLRP3 (dilution, 1:1,000; cat. no.
ab263899), apoptosis-associated speck-like protein (ASC; dilution,
1:1,000; cat. no. ab283684), caspase-1 (dilution, 1:1,000; cat. no.
ab207802), Bcl-2 (dilution, 1:1,000; cat. no. ab32124),
phosphorylated (p)-AMPK (dilution, 1:1,000; cat. no. ab92701), AMPK
(dilution, 1:1,000; cat. no. ab110036), p-Nrf2 (dilution, 1:500;
cat. no. ab76026), Nrf2 (dilution, 1:1,000; cat. no. ab137550), Bax
(dilution, 1:1,000; cat. no. ab32503), cleaved caspase-3 (dilution,
1:500; cat. no. ab32042), caspase-3 (dilution, 1:1,000; cat. no.
ab32351), cleaved caspase-6 (dilution, 1:500; cat. no. 9761; Cell
Signaling Technology, Inc.), caspase-6 (dilution, 1:1,000; cat. no.
ab108335), cleaved caspase-9 (dilution, 1:500; cat. no. ab2324),
caspase-9 (dilution, 1:1,000; cat. no. ab32539) and heme
oxygenase-1 (HO-1; dilution, 1:2,000; cat. no. ab52947). The
membranes were then blotted with species-matched secondary
antibodies (dilution, 1:1,000; cat. nos. ab6728 or ab6721; Abcam)
for 1 h at room temperature. Protein bands were visualized using
the ECL system (Pierce; Thermo Fisher. Scientific, Inc.), and the
band density was analyzed using ImageJ software (version 1.49;
National Institutes of Health).
LDH, malondialdehyde (MDA) and
superoxide dismutase (SOD) level determination
The levels of LDH and MDA and the activity of SOD
were evaluated using the corresponding assay kits according to the
manufacturer's protocols. Briefly, MPC5 cells were seeded into
six-well plates (2x105 cells/well) and treated with or
without HG and gastrodin for 24 h at 37˚C. Following treatment,
cells were harvested and centrifuged at 4˚C, 14,000 x g for 5 min
before the levels of LDH were measured in the cell supernatants
using the LDH Assay kit (cat. no. C0016; Beyotime Institute of
Biotechnology). Accordingly, the levels of MDA and SOD in the cell
supernatants were determined using MDA and SOD Assay kits (cat. no.
S0131S and S0086; both from Beyotime Institute of Biotechnology),
respectively. A microplate reader was used to detect the absorbance
in each well at a wavelength of 595 nm.
TUNEL assay
Following treatment with HG and gastrodin, TUNEL
staining assay was performed to evaluate the apoptosis rate of MPC5
cells. The TUNEL Apoptosis Detection kit (cat. no. 11684817910;
Roche Applied Science) was used according to the manufacturer's
protocols. In brief, the cells were washed twice with PBS after the
experimental procedures, then fixed with 4% paraformaldehyde at
room temperature in the dark for 30 min. Following incubation with
proteinase K for 15 min at 37˚C, cells were placed in 3%
H2O2 for 15 min at room temperature and
stained using the TUNEL detection kit. Subsequently, cells were
incubated with DAB for 10 min, counterstained with hematoxylin for
2 min at room temperature and washed with PBS. Images from
TUNEL-positive cells were captured under a light microscope
(Olympus Corporation)at x200 magnification from five separate
randomly selected fields. The TUNEL apoptosis rate (%)=number of
TUNEL-positive podocytes/total number of podocytes x100%.
Statistical analysis
Data are presented as the mean ± SEM from at least
three independent experiments. All data were analyzed using the
SPSS 19.0 software (IBM Corp.). Multiple group comparisons were
analyzed by one-way ANOVA followed by Tukey's or Bonferroni post
hoc tests. P<0.05 was considered to indicate a statistically
significant difference.
Results
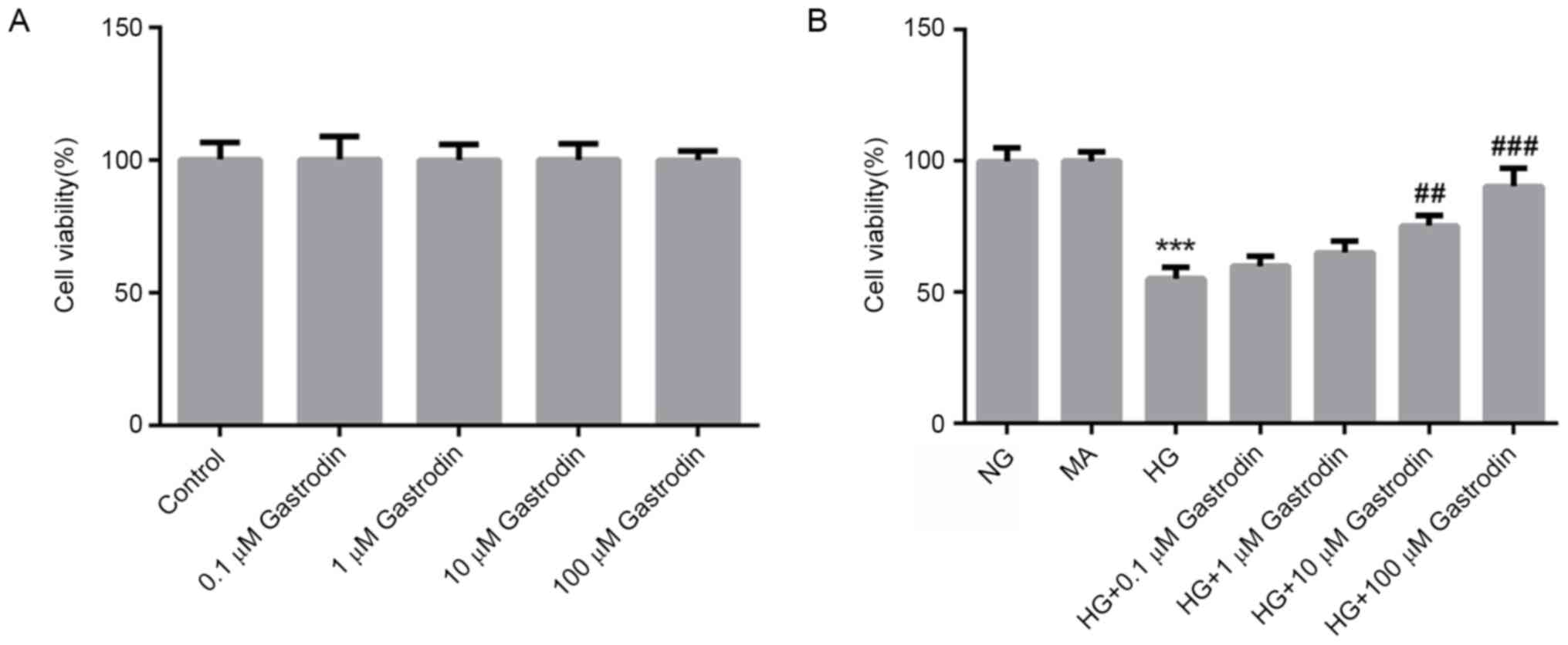
Gastrodin increases the viability of
HG-treated MPC5 cells
To evaluate the cytotoxicity of gastrodin, MPC5
cells were treated with different concentrations of gastrodin for
24 h. CCK-8 assay results showed that treatment with gastrodin had
no effect on cell viability (Fig.
1A). To examine the effects of gastrodin on cell viability
under HG conditions, MPC5 cells were pre-treated with different
doses of gastrodin prior to HG treatment. CCK-8 results showed that
gastrodin treatment increased the viability of HG-induced MPC5
cells in a dose-dependent manner, with the increases becoming
significant at 10 and 100 µM (Fig.
1B). Therefore, the dose of 100 µM gastrodin was selected for
subsequent experiments.
Gastrodin alleviates HG-induced
inflammation and oxidative stress in MPC5 cells
To explore the effect of gastrodin in the
development of DN, the concentration of inflammatory cytokines,
oxidative stress status and the expression levels of NLRP3
signaling-related proteins were next determined. The ELISA results
revealed that HG challenge significantly enhanced the secretion of
TNF-α, IL-1β and IL-6, whilst gastrodin treatment partially but
significantly reversed this effect (Fig. 2A). Furthermore, western blot
analysis results demonstrated that HG challenge significantly
upregulated the protein expression levels of MCP-1, NLRP3, ASC and
caspase-1, which may serve an important role in mediating the
inflammatory responses (Fig. 2B).
Pre-treatment with gastrodin also partially but significantly
reversed the effects of HG on the expression of MCP-1, NLRP3, ASC
and caspase-1. The activities of LDH and SOD, and the levels of MDA
were subsequently detected as biomarkers of oxidative stress. As
shown in Fig. 2C, HG challenge
significantly enhanced MDA levels and LDH activity, but
significantly suppressed SOD activity. These effects were partially
but significantly reversed by gastrodin pre-treatment. These
results suggested that gastrodin can alleviate HG-induced
inflammation and oxidative stress in MPC5 cells.
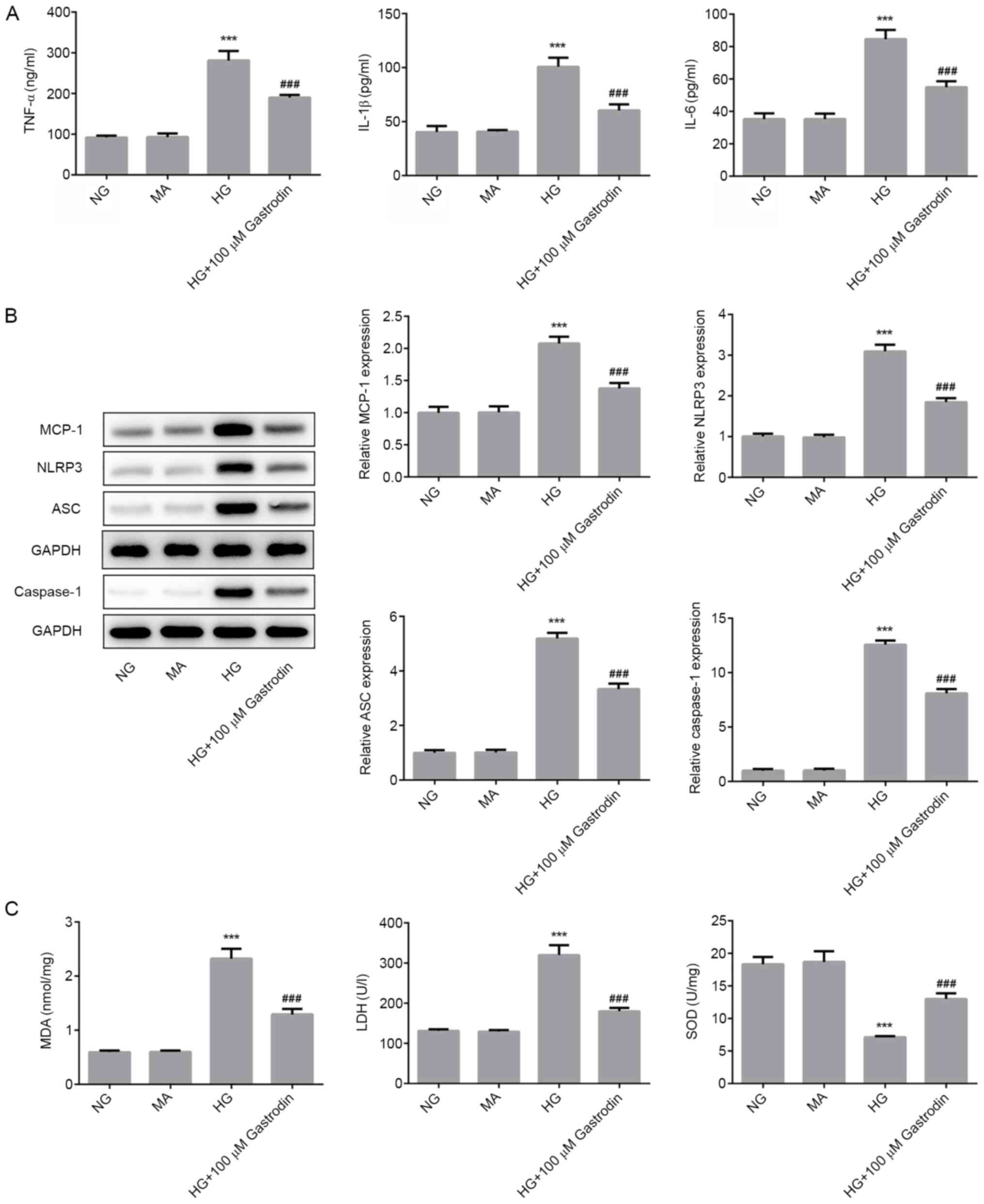 | Figure 2Gastrodin alleviates HG-induced
inflammation and oxidative stress in MPC5 cells. (A) The secretion
of inflammatory cytokines TNF-α, IL-1β and IL-6 were analyzed by
ELISA. (B) The expressions levels of MCP-1, NLRP3, ASC and
Caspase-1 were determined by western blotting and quantified. (C)
MDA levels and the activities of LDH and SOD were quantified using
their respective assay kits. ***P<0.001 vs. NG.
###P<0.001 vs. HG. NG, normal glucose; HG, high
glucose; MA, mannitol; LDH, lactate dehydrogenase; SOD, superoxide
dismutase; MDA, malondialdehyde; MCP-1, monocyte chemoattractant
protein 1; ASC, apoptosis-associated speck-like protein; NLRP3,
NOD-, LRR- and pyrin domain-containing protein 3. |
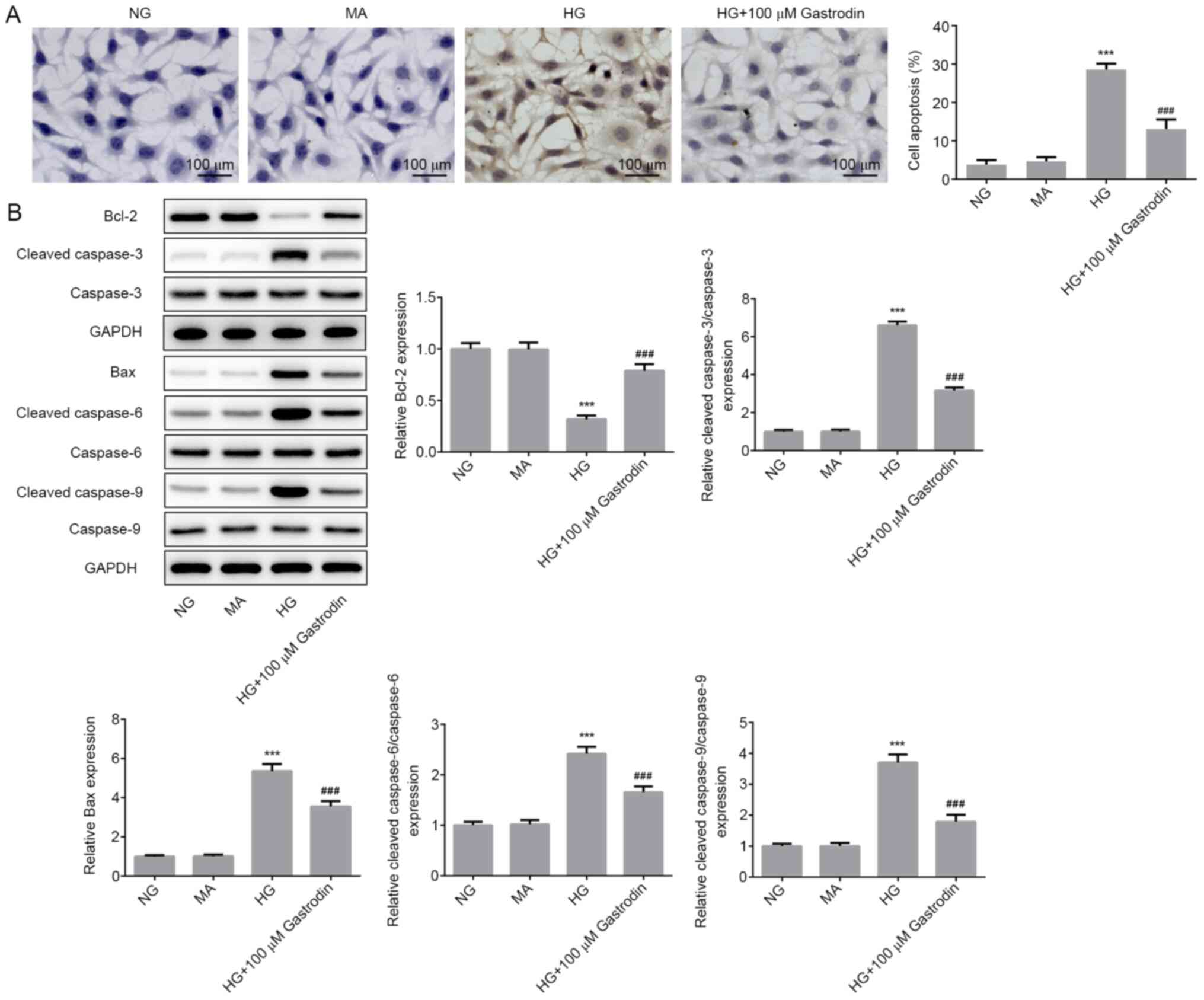
Gastrodin inhibits HG-induced
apoptosis in MPC5 cells
To investigate the effect of gastrodin further, cell
apoptosis and the expression of apoptosis-related proteins were
determined. TUNEL assay results showed that HG stimulation
significantly promoted MPC5 cell apoptosis, whereas cell treatment
with gastrodin significantly attenuated HG-induced apoptosis
(Fig. 3A). Furthermore, the
expression levels of the antiapoptotic protein Bcl-2 were
significantly decreased in the HG group compared with those in the
NG group. Additionally, the expression levels of the proapoptotic
protein Bax and those of cleaved caspase-3, cleaved caspase-6 and
cleaved caspase-9 were all significantly increased in the HG group
compared with those in the NG group. Gastrodin treatment
significantly reversed the HG-induced Bcl-2 downregulation and
upregulation of Bax, cleaved caspase-3, cleaved caspase-6 and
cleaved caspase-9 (Fig. 3B). These
aforementioned findings suggest that gastrodin can inhibit
HG-induced MPC5 cell apoptosis.
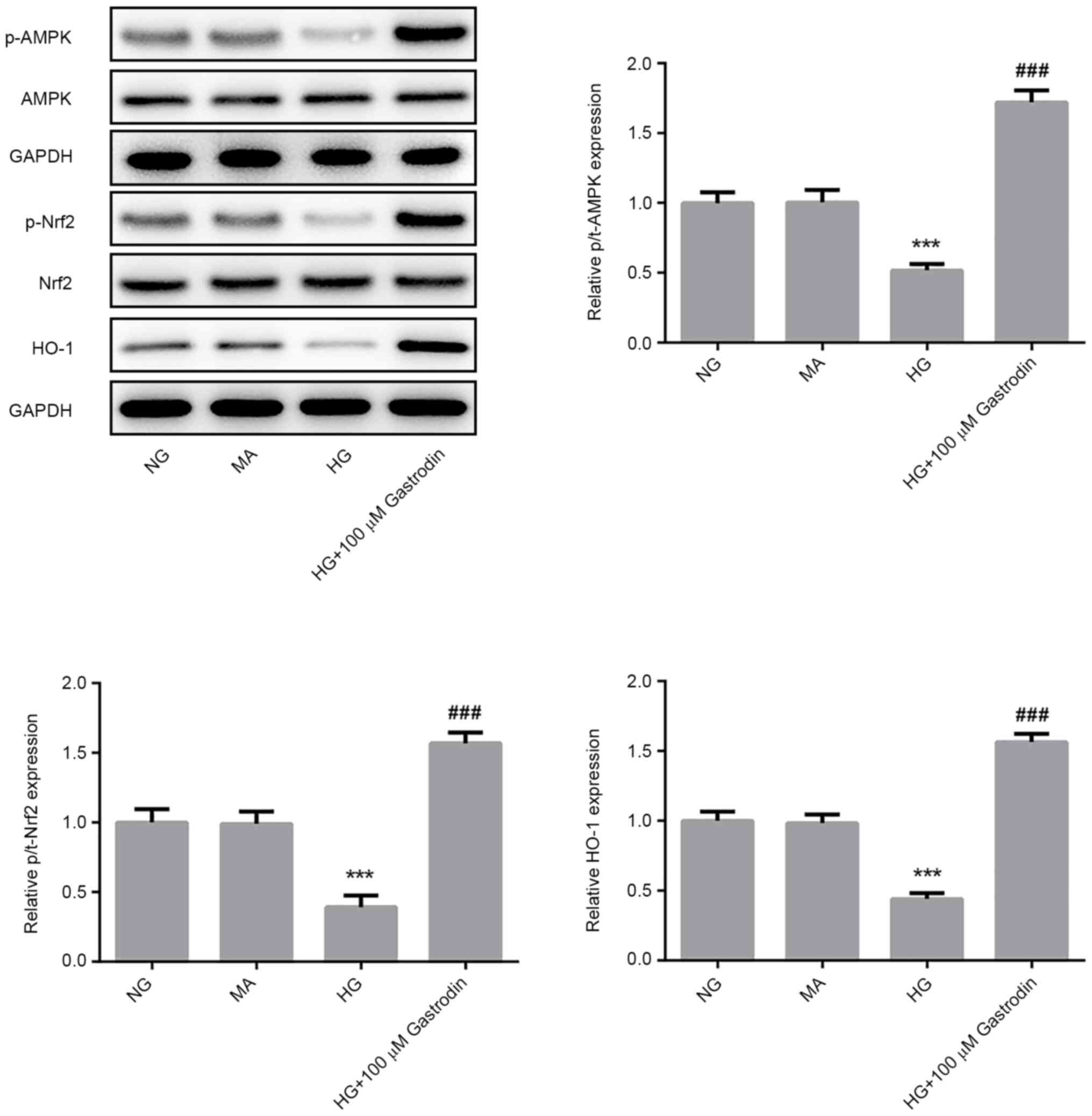
Gastrodin inhibits HG-induced
inflammation, oxidative stress, and apoptosis by activating the
AMPK/Nrf2 signaling pathway in MPC5 cells
To investigate the specific mechanism underlying the
effect of gastrodin on podocyte injury, the protein levels of
p-AMPK/AMPK, p-Nrf2/Nrf2 and HO-1 were detected by western blot
analysis. HG challenge significantly reduced the ratios of
p-AMPK/AMPK and p-Nrf2/Nrf2 and the expression of HO-1 (Fig. 4). The levels of AMPK and Nrf2
phosphorylation and HO-1 expression were found be significantly
upregulated in the HG + 100 µM gastrodin group compared with those
in the HG group (Fig. 4),
suggesting that gastrodin can promote the activation of the
AMPK/Nrf2 signaling pathway.
To verify the biochemical mechanism of gastrodin,
cells were treated with compound C to inhibit AMPK. MPC5 cells were
first pretreated with 20 µM compound C for 1 h prior to HG
treatment before the secretion levels of inflammatory cytokines,
oxidative stress markers, cell apoptosis and the expression of
apoptosis-related proteins were all measured. Pre-treatment with
compound C significantly reversed the inhibitory effects of
gastrodin on the secretion of inflammatory factors TNF-α, IL-1β and
IL-6 under HG (Fig. 5A). The
expression levels of NLRP3, ASC and caspase-1 were significantly
increased in the HG + 100 µM gastrodin + compound C group compared
with those in the HG + 100 µM gastrodin group (Fig. 5B). Additionally, treatment with
compound C significantly reversed the effects of gastrodin on MDA
levels and activities of LDH and SOD (Fig. 5C).
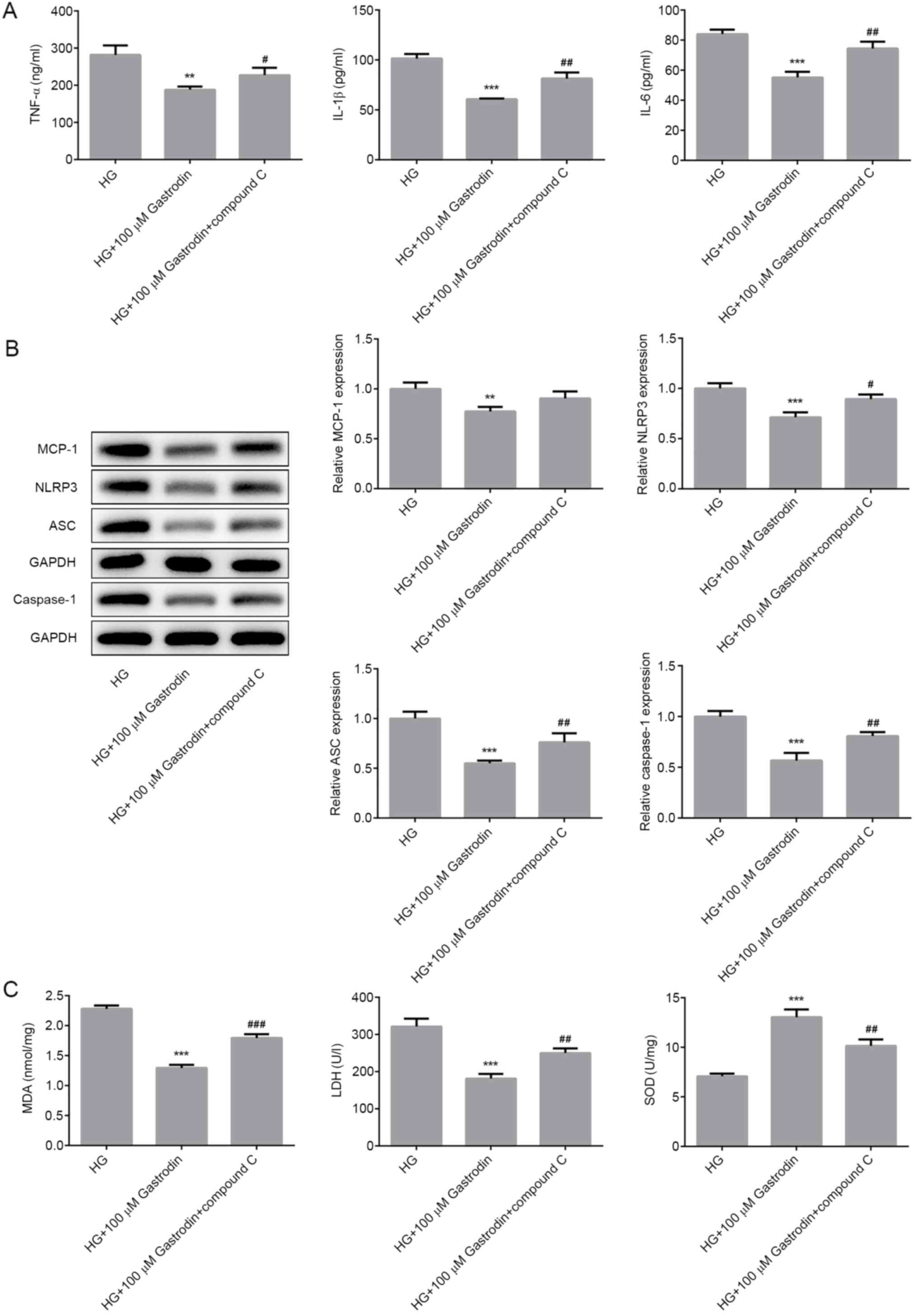 | Figure 5Gastrodin inhibits HG-induced
inflammation and oxidative stress by activating the 5'AMP-activated
protein kinase/nuclear factor erythroid 2-related factor 2
signaling pathway in MPC5 cells. (A) The secretion of inflammatory
cytokines, TNF-α, IL-1β and IL-6 were analyzed by ELISA. (B) The
expression levels of MCP-1, NLRP3, ASC and Caspase-1 were
determined by western blotting and quantified. (C) MDA levels and
the activities of LDH and SOD were quantified using their
respective assay kits. Error bars represent the mean ± SEM from
three independent experiments. **P<0.01 and
***P<0.001 vs. HG. #P<0.05,
##P<0.01 and ###P<0.001 vs. HG + 100 µM
gastrodin. HG, high glucose; LDH, lactate dehydrogenase; SOD,
superoxide dismutase; MDA, malondialdehyde; MCP-1, monocyte
chemoattractant protein 1; ASC, apoptosis-associated speck-like
protein; NLRP3, NOD-, LRR- and pyrin domain-containing protein
3. |
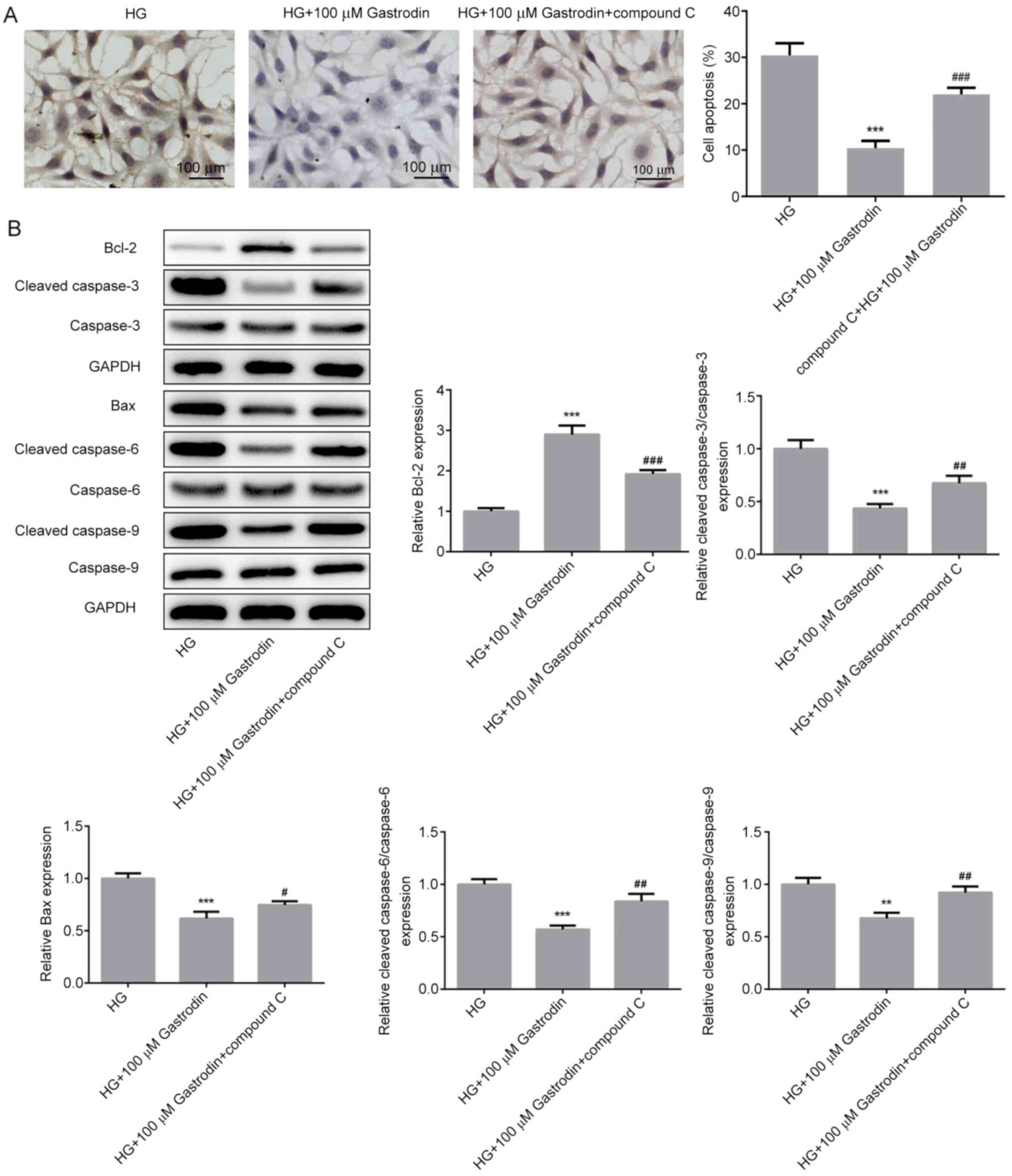
A significantly increased apoptosis rate (Fig. 6A) and expression of proapoptotic
proteins Bax, cleaved caspase-3, cleaved caspase-6 and cleaved
caspase-9 (Fig. 6B) were detected
in cells treated with compound C compared with those in the HG +
100 µM gastrodin group. These findings suggest that gastrodin can
attenuate HG-induced inflammation, oxidative stress and apoptosis
through the activation of the AMPK/Nrf2 signaling pathway in MPC5
cells.
Discussion
DN is characterized by glomerular hypertrophy,
decreased glomerular filtration, proteinuria and renal fibrosis and
is recognized as a chronic complication of diabetes mellitus that
can eventually lead to the loss of renal function (33). The diagnosis of DN is normally made
based on microalbuminuria (34).
Zhang et al (35)
demonstrated that ~20% of patients with diabetes may exhibit
nephropathy complications and the prevalence of DN has been
increasing rapidly in China (36).
However, the pathogenic mechanism of DN remains poorly understood.
Previous studies have shown that podocyte dysfunction serves a
significant role in the development and progression of DN (37,38).
In addition, recent studies have suggested that gastrodin can exert
a protective effect against HG-induced systemic organ injury
(19,20). Therefore, the present study aimed to
explore the effect of gastrodin on podocyte injury and evaluate its
potential mechanism. In the present study, 5.5 and 30 mM glucose
were used to simulate normal and high glucose conditions,
respectively (39). The results
revealed that gastrodin suppressed inflammation, oxidative stress
and apoptosis in HG-stimulated podocytes by activating the
AMPK/Nrf2 pathway, suggesting a protective effect of gastrodin on
DN progression.
Emerging evidence has suggested that HG-induced
oxidative stress is involved the development and progression of
diabetes, where long-term oxidative stress under pathological
conditions can induce excessive inflammatory responses (40). In addition, the multiprotein
inflammasome complex, may serve an important role in initiating the
inflammatory response. NLRP3 induces the recruitment and the
autocatalytic activation of caspase-1, causing the formation of an
inflammasome complex mediated by ASC (41). The formation of NLRP3 inflammasome
and the activation of caspase-1 facilitates the processing of the
cytosolic precursors of IL-1β and IL-18, resulting in the secretion
of these biologically active cytokines (42). In the present study, HG challenge
induced oxidative stress and promoted the secretion of inflammatory
cytokines. In addition, gastrodin alleviated the HG-induced
inflammation and oxidative stress in MPC5 cells, suggesting that
gastrodin can exert therapeutic effects on DN. Inflammation and
oxidative stress contribute to abnormal tubular cell cycle
progression and disrupted cell proliferation, which have been
proposed to be important mechanisms underlying multiple kidney
diseases, including acute kidney injuries, chronic kidney diseases
and polycystic kidney diseases (43). The results of the present study
demonstrated that gastrodin dose-dependently increased the
viability of HG-induced MPC5 cells. Furthermore, gastrodin
inhibited HG-induced MPC5 cell apoptosis, suggesting that gastrodin
can protect against HG-induced podocyte apoptosis.
To investigate the specific mechanism underlying the
protective effect of gastrodin on podocyte injury, the expression
levels of the proteins associated with AMPK/Nrf2 signaling were
determined. Accumulating evidence has suggested that the AMPK/Nrf2
signaling pathway serves a significant role in the pathogenesis of
several inflammation-related diseases, such as atherosclerosis,
diabetes and cancer (44-46).
For example, gastrodin was found to reduce oxidative stress and
inflammation, whilst improving lipid metabolism in a model of
nonalcoholic fatty liver disease by activating AMPK/Nrf2 signaling
(24). In the present study,
gastrodin upregulated the phosphorylation levels of AMPK, Nrf2 and
the expression of HO-1, which was consistent with the previous
study aforementioned. Previous studies also reported that AMPK and
Nrf2 can mediate fatty acid oxidation and reduce oxidative stress,
respectively (47,48). The activation of AMPK/Nrf2 signaling
was previously found to improve lipid metabolism and attenuated
oxidative stress in renal tissues of mice with type 2
diabetes-induced nephropathy (25,49).
Therefore, the present study hypothesized that gastrodin could
inhibit the HG-triggered inflammation, oxidative stress and
apoptosis by activating the AMPK/Nrf2 signaling pathway in MPC5
cells. To verify these hypotheses, MPC5 cells were treated with
compound C, an AMPK inhibitor. The results indicated that the
inhibitory effects of gastrodin on HG-triggered inflammation,
oxidative stress and apoptosis were overturned following cell
exposure to compound C. These findings suggested that gastrodin
could protect against HG-induced podocyte injury, inflammation and
oxidative stress by activating the AMPK/Nrf2 signaling pathway,
indicating this to a promising therapeutic approach for treating
DN.
However, there are several limitations to the
present study. The effects of gastrodin on DN were only observed
in vitro. The role of gastrodin in animal DN models and
human tissues with DN was not explored in the present study. CCK-8
assay was used to detect the effects of different doses of
gastrodin on cell viability, which found that 0-100 µM gastrodin
had no significant effect on the cell viability of untreated MPC5
cells. Due to the substantial difference in the experiments in
vitro and in vivo, the dose of gastrodin used in the
present study can only serve as a reference for future in
vitro experiments. The appropriate dose of gastrodin for
clinical use in humans require further investigation. In addition,
the focus of the present study was to investigate the effects of
gastrodin on inflammation, oxidative stress and apoptosis of
podocytes, but did not explore the effects of HG and gastrodin
treatment on cell cycle progression. Finally, the expression of
proteins in the AMPK/Nrf2 signaling pathway was mainly investigated
in the present study, but it did not explore the effects of
gastrodin on the Sirtuin-1 pathway, which is another signaling
pathway that is associated with AMPK (50).
In summary, gastrodin treatment restored podocyte
viability under HG conditions in a dose-dependent manner. Notably,
gastrodin inhibited HG-induced inflammation, oxidative stress and
podocyte apoptosis. Furthermore, gastrodin promoted the activation
of the AMPK/Nrf2 signaling pathway in HG-stimulated podocytes.
Collectively, these findings suggest that gastrodin can exhibit a
protective effect against podocyte injury by activating AMPK/Nrf2
signaling in DN. Therefore, gastrodin can be considered as an
effective therapeutic agent for treating DN.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LH and YZ designed the experiments. LH and MS
performed the experiments and analyzed the data. YZ and LH confirm
the authenticity of all the raw data. All authors read and approved
the final version of the manuscript.
Ethics aproval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declared that they have no competing
interests.
References
|
1
|
Gnudi L, Coward RJM and Long DA: Diabetic
nephropathy: Perspective on novel molecular mechanisms. Trends
Endocrinol Metab. 27:820–830. 2016.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Yamahara K, Yasuda M, Kume S, Koya D,
Maegawa H and Uzu T: The role of autophagy in the pathogenesis of
diabetic nephropathy. J Diabetes Res. 2013(193757)2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Gheith O, Farouk N, Nampoory N, Halim MA
and Al-Otaibi T: Diabetic kidney disease: World wide difference of
prevalence and risk factors. J Nephropharmacol. 5:49–56.
2016.PubMed/NCBI
|
|
4
|
Feng Y, Weng H, Ling L, Zeng T, Zhang Y,
Chen D and Li H: Modulating the gut microbiota and inflammation is
involved in the effect of Bupleurum polysaccharides against
diabetic nephropathy in mice. Int J Biol Macromol. 132:1001–1011.
2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Morigi M, Perico L, Corna D, Locatelli M,
Cassis P, Carminati CE, Bolognini S, Zoja C, Remuzzi G, Benigni A
and Buelli S: C3a receptor blockade protects podocytes from injury
in diabetic nephropathy. JCI Insight. 5(e131849)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhang P, Fang J, Zhang J, Ding S and Gan
D: Curcumin inhibited podocyte cell apoptosis and accelerated cell
autophagy in diabetic nephropathy via regulating
Beclin1/UVRAG/Bcl2. Diabetes Metab Syndr Obes. 13:641–652.
2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Kim YI, Kim CH, Choi CS, Chung YE, Lee MS,
Lee SI, Park JY, Hong SK and Lee KU: Microalbuminuria is associated
with the insulin resistance syndrome independent of hypertension
and type 2 diabetes in the Korean population. Diabetes Res Clin
Pract. 52:145–152. 2001.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Al-Rubeaan K, Siddiqui K, Alghonaim M,
Youssef AM and AlNaqeb D: The Saudi diabetic kidney disease study
(Saudi-DKD): Clinical characteristics and biochemical parameters.
Ann Saudi Med. 38:46–56. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Lopes TG, de Souza ML, da Silva VD, Dos
Santos M, da Silva WIC, Itaquy TP, Garbin HI and Veronese FV:
Markers of renal fibrosis: How do they correlate with podocyte
damage in glomerular diseases? PLoS One.
14(e0217585)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen Y, Lin L, Tao X, Song Y, Cui J and
Wan J: The role of podocyte damage in the etiology of
ischemia-reperfusion acute kidney injury and post-injury fibrosis.
BMC Nephrol. 20(106)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ilatovskaya DV, Blass G, Palygin O,
Levchenko V, Pavlov TS, Grzybowski MN, Winsor K, Shuyskiy LS,
Geurts AM, Cowley AW Jr, et al: A NOX4/TRPC6 pathway in podocyte
calcium regulation and renal damage in diabetic kidney disease. J
Am Soc Nephrol. 29:1917–1927. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Khalilpourfarshbafi M, Hajiaghaalipour F,
Selvarajan KK and Adam A: Mesenchymal stem cell-based therapies
against podocyte damage in diabetic nephropathy. Tissue Eng Regen
Med. 14:201–210. 2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Chen Y, Liu Q, Shan Z, Zhao Y, Li M, Wang
B, Zheng X and Feng W: The protective effect and mechanism of
catalpol on high glucose-induced podocyte injury. BMC Complement
Altern Med. 19(244)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Zhan X, Yan C, Chen Y, Wei X, Xiao J, Deng
L, Yang Y, Qiu P and Chen Q: Celastrol antagonizes high
glucose-evoked podocyte injury, inflammation and insulin resistance
by restoring the HO-1-mediated autophagy pathway. Mol Immunol.
104:61–68. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yu Q, Zhang M, Qian L, Wen D and Wu G:
Luteolin attenuates high glucose-induced podocyte injury via
suppressing NLRP3 inflammasome pathway. Life Sci. 225:1–7.
2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhong Y, Lee K, Deng Y, Ma Y, Chen Y, Li
X, Wei C, Yang S, Wang T, Wong NJ, et al: Arctigenin attenuates
diabetic kidney disease through the activation of PP2A in
podocytes. Nat Commun. 10(4523)2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Xu CB, Guo QL, Wang YN, Lin S, Zhu CG and
Shi JG: Gastrodin derivatives from gastrodia elata. Nat Prod
Bioprospect. 9:393–404. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liu Y, Gao J, Peng M, Meng H, Ma H, Cai P,
Xu Y, Zhao Q and Si G: A review on central nervous system effects
of gastrodin. Front Pharmacol. 9(24)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ye T, Meng X, Zhai Y, Xie W, Wang R, Sun G
and Sun X: Gastrodin ameliorates cognitive dysfunction in diabetes
rat model via the suppression of endoplasmic reticulum stress and
NLRP3 inflammasome activation. Front Pharmacol.
9(1346)2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhang TH, Huang CM, Gao X, Wang JW, Hao LL
and Ji Q: Gastrodin inhibits high glucose-induced human retinal
endothelial cell apoptosis by regulating the SIRT1/TLR4/NF-κBp65
signaling pathway. Mol Med Rep. 17:7774–7780. 2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Cheng Y, Qi Y, Liu S, Di R, Shi Q, Li J
and Pei C: C1q/TNF-related protein 9 inhibits high glucose-induced
oxidative stress and apoptosis in retinal pigment epithelial cells
through the activation of AMPK/Nrf2 signaling pathway. Cell
Transplant. 29(963689720962052)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Xu W, Zhao T and Xiao H: The implication
of oxidative stress and AMPK-Nrf2 antioxidative signaling in
pneumonia pathogenesis. Front Endocrinol (Lausanne).
11(400)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhou F, Wang M, Ju J, Wang Y, Liu Z, Zhao
X, Yan Y, Yan S, Luo X and Fang Y: Schizandrin A protects against
cerebral ischemia-reperfusion injury by suppressing inflammation
and oxidative stress and regulating the AMPK/Nrf2 pathway
regulation. Am J Transl Res. 11:199–209. 2019.PubMed/NCBI
|
|
24
|
Qu LL, Yu B, Li Z, Jiang WX, Jiang JD and
Kong WJ: Gastrodin ameliorates oxidative stress and proinflammatory
response in nonalcoholic fatty liver disease through the AMPK/Nrf2
pathway. Phytother Res. 30:402–411. 2016.PubMed/NCBI View
Article : Google Scholar
|
|
25
|
Ma T, Zheng Z, Guo H, Lian X, Rane MJ, Cai
L, Kim KS, Kim KT, Zhang Z and Bi L: 4-O-methylhonokiol ameliorates
type 2 diabetes-induced nephropathy in mice likely by activation of
AMPK-mediated fatty acid oxidation and Nrf2-mediated anti-oxidative
stress. Toxicol Appl Pharmacol. 370:93–105. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wang RM, Wang ZB, Wang Y, Liu WY, Li Y,
Tong LC, Zhang S, Su DF, Cao YB, Li L and Zhang LC: Swiprosin-1
promotes mitochondria-dependent apoptosis of glomerular podocytes
via P38 MAPK pathway in early-stage diabetic nephropathy. Cell
Physiol Biochem. 45:899–916. 2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Han X, Li Q, Wang C and Li Y:
MicroRNA-204-3p attenuates high glucose-induced MPC5 podocytes
apoptosis by targeting braykinin B2 receptor. Exp Clin Endocrinol
Diabetes. 127:387–395. 2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Tu Q, Li Y, Jin J, Jiang X, Ren Y and He
Q: Curcumin alleviates diabetic nephropathy via inhibiting podocyte
mesenchymal transdifferentiation and inducing autophagy in rats and
MPC5 cells. Pharm Biol. 57:778–786. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Cao Y, Hao Y, Li H, Liu Q, Gao F, Liu W
and Duan H: Role of endoplasmic reticulum stress in apoptosis of
differentiated mouse podocytes induced by high glucose. Int J Mol
Med. 33:809–816. 2014.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xi Z, Qiao Y, Wang J, Su H, Bao Z, Li H,
Liao X and Zhong X: Gastrodin relieves inflammation injury induced
by lipopolysaccharides in MRC-5 cells by up-regulation of miR-103.
J Cell Mol Med. 24:1451–1459. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kim SH, Hwang JT, Park HS, Kwon DY and Kim
MS: Capsaicin stimulates glucose uptake in C2C12 muscle cells via
the reactive oxygen species (ROS)/AMPK/p38 MAPK pathway. Biochem
Biophys Res Commun. 439:66–70. 2013.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Chuang KC, Chen FW, Tsai MH and Shieh JJ:
EGR-1 plays a protective role in AMPK inhibitor compound C-induced
apoptosis through ROS-induced ERK activation in skin cancer cells.
Oncol Lett. 21(304)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Bhattacharjee N, Barma S, Konwar N,
Dewanjee S and Manna P: Mechanistic insight of diabetic nephropathy
and its pharmacotherapeutic targets: An update. Eur J Pharmacol.
791:8–24. 2016.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Papadopoulou-Marketou N, Chrousos GP and
Kanaka-Gantenbein C: Diabetic nephropathy in type 1 diabetes: A
review of early natural history, pathogenesis, and diagnosis.
Diabetes Metab Res Rev. 33(e2841)2017.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zhang XX, Kong J and Yun K: Prevalence of
diabetic nephropathy among patients with type 2 diabetes mellitus
in China: A meta-analysis of observational studies. J Diabetes Res.
2020(2315607)2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Xiong Y and Zhou L: The signaling of
cellular senescence in diabetic nephropathy. Oxid Med Cell Longev.
2019(7495629)2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Bose M, Almas S and Prabhakar S: Wnt
signaling and podocyte dysfunction in diabetic nephropathy. J
Investig Med. 65:1093–1101. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Tung CW, Hsu YC, Shih YH, Chang PJ and Lin
CL: Glomerular mesangial cell and podocyte injuries in diabetic
nephropathy. Nephrology (Carlton). 23 (Suppl 4):S32–S37.
2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wang S, Zhao X, Yang S, Chen B and Shi J:
Salidroside alleviates high glucose-induced oxidative stress and
extracellular matrix accumulation in rat glomerular mesangial cells
by the TXNIP-NLRP3 inflammasome pathway. Chem Biol Interact.
278:48–53. 2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhao MX, Zhou B, Ling L, Xiong XQ, Zhang
F, Chen Q, Li YH, Kang YM and Zhu GQ: Salusin-β contributes to
oxidative stress and inflammation in diabetic cardiomyopathy. Cell
Death Dis. 8(e2690)2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Sutterwala FS, Ogura Y, Szczepanik M,
Lara-Tejero M, Lichtenberger GS, Grant EP, Bertin J, Coyle AJ,
Galán JE, Askenase PW and Flavell RA: Critical role for
NALP3/CIAS1/Cryopyrin in innate and adaptive immunity through its
regulation of caspase-1. Immunity. 24:317–327. 2006.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Pétrilli V, Dostert C, Muruve DA and
Tschopp J: The inflammasome: A danger sensing complex triggering
innate immunity. Curr Opin Immunol. 19:615–622. 2007.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Lin TA, Wu VC and Wang CY: Autophagy in
chronic kidney diseases. Cells. 8(61)2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Kosuru R, Kandula V, Rai U, Prakash S, Xia
Z and Singh S: Pterostilbene decreases cardiac oxidative stress and
inflammation via activation of AMPK/Nrf2/HO-1 pathway in
fructose-fed diabetic rats. Cardiovasc Drugs Ther. 32:147–163.
2018.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Tanaka M, Kishimoto Y, Sasaki M, Sato A,
Kamiya T, Kondo K and Iida K: Terminalia bellirica (Gaertn.) Roxb.
extract and gallic acid attenuate LPS-induced inflammation and
oxidative stress via MAPK/NF-κB and Akt/AMPK/Nrf2 pathways. Oxid
Med Cell Longev. 2018(9364364)2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Wang Y, Huang Y, Xu Y, Ruan W, Wang H,
Zhang Y, Saavedra JM, Zhang L, Huang Z and Pang T: A Dual AMPK/Nrf2
activator reduces brain inflammation after stroke by enhancing
microglia M2 polarization. Antioxid Redox Signal. 28:141–163.
2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zeng HL, Huang SL, Xie FC, Zeng LM, Hu YH
and Leng Y: Yhhu981, a novel compound, stimulates fatty acid
oxidation via the activation of AMPK and ameliorates lipid
metabolism disorder in ob/ob mice. Acta Pharmacol Sin. 36:343–352.
2015.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhan X, Li J and Zhou T: Targeting
Nrf2-mediated oxidative stress response signaling pathways as new
therapeutic strategy for pituitary adenomas. Front Pharmacol.
12(565748)2021.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Kim Y, Lee H, Kim SY and Lim Y: Effects of
lespedeza bicolor extract on regulation of AMPK associated hepatic
lipid metabolism in type 2 diabetic mice. Antioxidants (Basel).
8(599)2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Huang Q, Wang T, Yang L and Wang HY:
Ginsenoside Rb2 alleviates hepatic lipid accumulation by restoring
autophagy via induction of Sirt1 and activation of AMPK. Int J Mol
Sci. 18(1063)2017.PubMed/NCBI View Article : Google Scholar
|