Introduction
Globally, colorectal cancer (CRC) is the third most
frequently detected neoplasm and the second leading cause of
cancer-associated mortality, with >1.8 million new CRC cases and
881,000 deaths being reported globally in 2018, with ~1 death per
10 confirmed cases (1,2). While the incidence of CRC in Middle
Eastern countries is low (3), there
is a higher annual incidence rate of CRC in the Kurdistan region of
Iraq with an estimated ~38-61.7 cases/100,000 people from 2006 to
2014. CRC is the fourth most widespread cancer in males and females
residing in this province, causing ~8.6% mortality of the total
annual fatalities in this region (4). There is strong evidence that CRC is
the most common type of adenocarcinoma in this region (5).
Commonly endogenous gasotransmitters that are used
are NO, CO and H2S (6).
These small molecules of gas, which have limited concentrations and
particular functions, are endogenously and enzymatically generated
(7). NO is produced from L-arginine
by NO synthase (8,9), while H2S is synthesized
from L-cysteine by both cystathionine β-synthase and cystathionine
γ-lyase (CSE), or mercaptopyruvate sulfurtransferase (10,11).
The aforementioned gaseous molecules modulate different biological
pathways and functions such as SMC relaxation, at physiologically
relevant concentrations (6) by
opening a number of membrane ion channels (12). NO is an essential regulator of
angiogenesis (13) and
vasorelaxation via the activation of guanylate cyclase and the
production of cyclic guanidine monophosphate (cGMP) (14). cGMP increases endothelial
Ca+2 concentration and contributes to the opening of
localized Ca+2-dependent K+ channels
(KCa) (15). In
contrast, H2S exerts its effects on vasodilation
(16) and angiogenesis (17) via the direct activation of
KATP channels (18).
NO and H2S also act synergistically or
antagonistically to stimulate their downstream pathways, ranging
from biosynthesis to their signaling cascade within target cells
(19). NO and H2S are
mutually dependent on regulating vasodilation, angiogenesis
(20) and endothelial homeostasis
(21). The role of these gases in
cancer remains unclear, as both exhibit tumor promotion and
anti-tumor properties (22).
Furthermore, NO and H2S have been indicated to modulate
a variety of cancer cell functions, including proliferation,
invasion, metastasis and tumor angiogenesis (23). Additionally, the enzymes responsible
for NO and H2S production are upregulated in CRC cells
and endogenously produce low-to-mid concentrations of
H2S or NO to support cell proliferation, while exogenous
delivery of H2S or NO suppresses the division of colon
cancer cells (24).
Intra-tumor blood vessels are vital for tumor
growth, metastasis, cancer treatment (25) and the acquisition of differential
reactivity by functional mature blood vessels in the tumor
micro-environment, representing an appropriate target for
anti-tumor therapeutic agents (26). Cancerous cells exhibit accelerated
metabolism and, therefore, demand high reactive oxygen species
(ROS) concentrations to maintain high proliferation rates (27). The high level of ROS damages and/or
destroys cells by oxidizing proteins, lipids and nucleic acids
(28). These observations clarify
that oxidative stress and cancer are closely associated (29). Several researchers are investigating
the consequences of ROS and endothelial dysfunction in cancer
(30-32).
Based on literature reviews, to the best of our knowledge, no
studies appear to have presented an association between oxidative
stress, endothelial functions and vascular reactivity of NO and
H2S in patients with CRC. Therefore, the current study
aimed to assess endocan as an endothelial functional marker and
malondialdehyde (MDA) as an oxidative stress marker in patients
with CRC. Additionally, the present study investigated the probable
mechanisms responsible for NO- and H2S-induced
vasodilation in the human mesenteric artery of patients with CRC.
To elucidate these mechanisms, the possible roles of different
K+ channels in the vasodilation response produced by NO
and H2S were explored.
Materials and methods
Patients
The current study was a case-control study. For the
first experiment, patients were recruited at two hospitals in
Erbil, Iraq: The Oncology Department of Rizgary and Nanakaly
Hospitals. Venous blood samples were taken from 44 patients (male,
24; female, 20) with different stages of CRC. Additionally, 40
healthy volunteers (male, 22; female, 18) of similar ages were
recruited randomly in Erbil city as control samples. For the second
experiment, colorectal tumour specimens were obtained from patients
with CRC undergoing partial colectomy at Consultancy Medical City
and Welfare private hospitals in Erbil. In both experiments,
patients were recruited between August to November 2016 and
patient's median age was 55 years old (ranged between 35-70).
Patients with underlying immunodeficiency disorder or
immunodeficiency state and individuals who had other co-morbid
health problems which could introduce heterogeneity to the sample,
such as additional acquired brain injury, arthritis, chronic
obstructive pulmonary disease, asthma, diabetes mellitus,
ankylosing spondylitis, connective tissue diseases and other
inflammatory diseases were excluded.
The blood samples were obtained by phlebotomy under
an aseptic technique. Blood was placed into a clot activator tube
for serum separation. The sera were then separated under
centrifugation at 448 x g for 5 min at 37˚C. Patients with an
underlying immunodeficiency disorder or state of immunodeficiency,
and individuals who presented with other co-morbid health problems
which could introduce heterogeneity to the sample (including
arthritis, asthma, diabetes mellitus, hypertension and other
inflammatory diseases) were excluded.
Determination of endocan
The concentration of endocan was determined using
the Human ESM1 ELISA kit (cat. no. E-EL-H1557; Elabscience, Inc.)
by the Sandwich-ELISA method (33).
The micro ELISA plate was pre-coated with antibodies specific to
endocan. Standards or samples were added to the suitable micro
ELISA plate wells and combined with the endocan-specific antibodies
(Elabscience, Inc.). A biotinylated detection antibody, part of the
aforementioned kit, specific for endocan and avidin-horseradish
peroxidase (HRP)-conjugate and substrate were then added to each
well. Only the wells that contained endocan, biotinylated detection
antibodies and avidin-HRP conjugate appeared blue in color and the
reaction was stopped with a 1N H2SO4 solution
and the color turned yellow. The absorbance was measured
spectrophotometrically (ELISA reader; Biotek) at a wavelength of
450 nm.
Determination of serum MDA
MDA was determined according to the Ohkawa method
(34). The procedure started by
thiobarbituric acid (TBA) preparation, in which 0.66 g TBA was
dissolved in 100 ml of 0.05 M of NaOH with simple heating at 45˚C.
Trichloroacetic acetic acid (TCA) was then prepared by dissolving
17.5 g TCA in 100 ml of distilled water. TCA2 was prepared by
dissolving 70 g of TCA in 100 ml of distilled water. Finally, 150
ml of either control or CRC patient's serum was added, 1 ml of TCA1
was mixed for 2 min and placed in a boiling water bath for 15 min
and 1 ml of TCA2 was added and incubated for 20 min at 37˚C. The
solution was centrifuged for 5 min at 448 x g. The supernatant was
read (Unico SpectroQuest SQ2800 UV-Visible Spectrophotometer;
Unico) at 532 nm.
Myographical recording Vessel
collection and preparation
Human mesenteric arteries were collected during
surgery from patients with CRC undergoing partial colectomy. The
arteries supplying blood to the tumors were dissected surgically
and placed into beakers containing cold modified Krebs solution
(Sigma-Aldrich; Merck KGaA) (Fig.
1) and aerated with 95% O2 and 5% CO2.
The excess tissue and fat were removed in the laboratory and the
arteries were cut into rings (length, ~3-4 mm).
Recording of isometric tension
The procedure by Furchgott and Zawadzki (35) with certain modifications in the
Krebs solution's concentration was followed to study the
vasodilator activity of the isolated mesenteric arteries. The
arterial rings were propped by two stainless steel clamps. The
first clamp was attached to a hook at the underside of the organ
bath and the second was connected to the force transducer through a
thread to record the isometric tension of the mesenteric arteries.
Data were recorded using LabChart 7.1 data acquisition software
(ADInstruments, Inc.). The propped arterial rings were immersed in
modified Krebs solution (NaCl 5.10 gm/l, NaHCO3 1.94
gm/l, MgSO4 0.686 gm/l, KCl 2.24 gm/l,
KH2PO4 0.15 gm/l, CaCl2 0.277 gm/l
and C6H12O6 2 gm/l) and contained
in a 10 ml organ chamber. Krebs solution was maintained at a pH 7.4
and was constantly aerated with 95% O2 5% CO2
at 37˚C (panLab; Harvard Apparatus).
The mesenteric arterial rings were tensed to a
stable basal strain of 4 gm prior to being left to be equilibrated
for 2 h. The Krebs solution was replaced at 15-20 min intervals in
the bath chamber until it reached stability. Experimental
substances were added to the bath chambers, according to protocol.
The arteries were incubated at 37˚C with channel blockers for 20
min prior to pre-contraction with norepinephrine (NE; 1 µM;
Sigma-Aldrich; Merck KGaA). Following this, relaxation occurred by
bolus dose application of sodium nitroprusside (SNP; Sigma-Aldrich;
Merck KGaA) or sodium sulphide (Na2S; Hangzhou J&H
Chemical Co.).
Experimental protocol
The arterial rings, which were pre-contracted with
NE, were first relaxed by the cumulative addition of either SNP (30
nM-30 µM) or Na2S (1-6 mM). Based on these initial
experiments, the relative half-inhibitory concentration
(IC50) of SNP (2.3 µM) or Na2S (2.4 mM) was
used to retest the ability to relax pre-contracted rings in three
separate sets of experiments. In the first experiment, when the
NE-induced contraction reached the uppermost value, SNP (2.3 µM) or
Na2S (2.4 mM) was added and left for 60 min, and the
maximal relaxation rate (%) was calculated four times at each 15
min interval (n=8). Following this, the role of K+
channels in the progress of SNP and Na2S mediated
relaxation were tested via incubation at 37˚C of the arterial rings
for 20 min using tetraethylammonium (TEA; 1 mM), glibenclamide
(GLIB; 0.1 µM), barium chloride (BaCl2; 1 mM) and 4-aminopyridine
(4-AP; 1 mM) (all supplied from Hangzhou J&H Chemical Co.). In
the second experiment, the role of endogenous NO and H2S
were tested by the pre-incubation of arterial rings with
endothelial NO and CSE antagonists, L-nitro-arginine methyl ester
(L-NAME; 3x10-4 M) or D,L-propargylglycine (PAG; 10 mM)
(Sigma-Aldrich; Merck KGaA), respectively for 20 min prior to
applying SNP (n=8). Finally, to examine whether the combination of
H2S and NO potentiates or inhibits vasorelaxation when
the NE-induced contraction reached the highest value, SNP and
Na2S were added simultaneously and left for 60 min
(n=8).
Statistical analysis
Comparisons between patients with CRC and healthy
individuals were performed using a Mann-Whitney test, and values
were presented as median and quartiles. Statistical analysis of
myographical data was performed using a two-way ANOVA followed by
Dunnett post-hoc test. Maximum relaxation responses were calculated
as a percentage of the contraction produced by NE and expressed as
the mean ± standard error of the mean. The tension created by NE
was defined as 0% relaxation, and the baseline tension prior to the
addition of NE was determined as 100% relaxation.
The graphs, calculations and statistical analyses
were performed using GraphPad Prism software (version 6.0; GraphPad
Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Serum endocan and MDA
concentrations
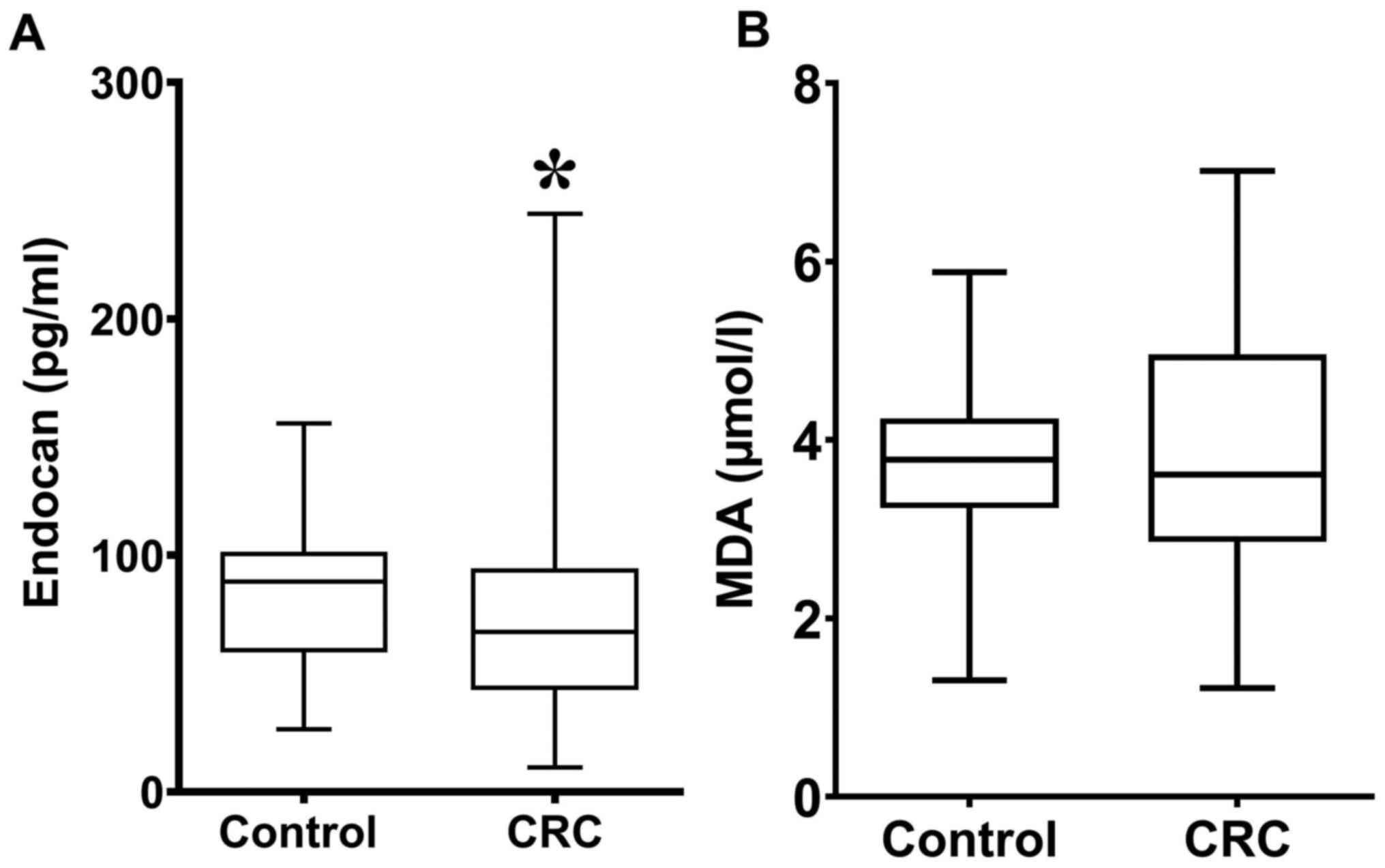
Serum endocan concentration was significantly lower
in patients with CRC (67.56; 43.04-94.28) compared with healthy
individuals (88.68; 59-101.3; Fig.
2A). There were no significant differences in MDA concentration
between patients with CRC (3.62; 2.86-4.96) and healthy individuals
(3.78; 3.23-4.24; Fig. 2B).
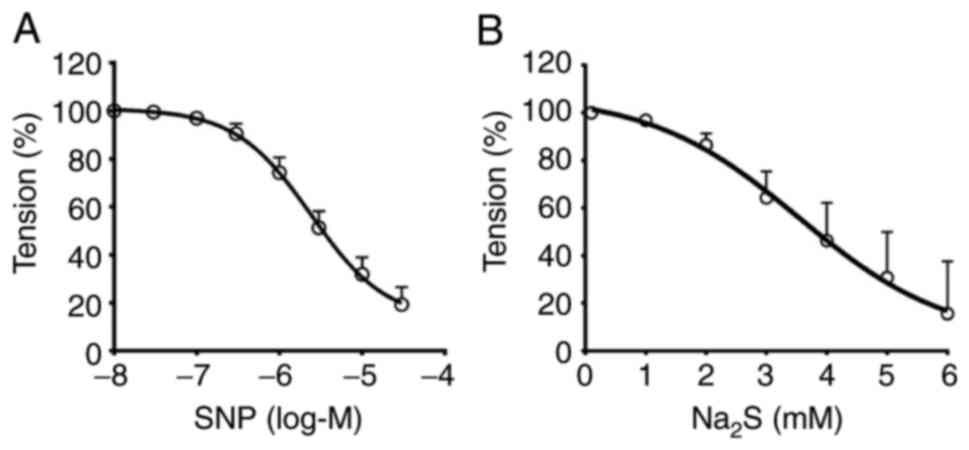
Measurement of IC50
SNP concentrations ranging from 30 nM-30 µM induced
a relaxant effect on CRC mesenteric arteries following
pre-contraction with NE (1 µM) with an IC50 value of
2.42±0.16 µM (CI 95%, 1.18-4.95 µM). The percentage of relaxation
was 80.74±7.256%. Na2S at concentrations from 1-6 mM had
a relaxant effect on mesenteric arteries of CRC pre-contracted with
NE. The calculated IC50 value was 3.54±1.07 mM (CI 95%,
1.4-5.68 mM) and the percentage of relaxation was 84.43±22.05%. The
concentration-response curve for the effect of SNP and
Na2S against NE-mediated contractions are presented in
Fig. 3A and B, respectively.
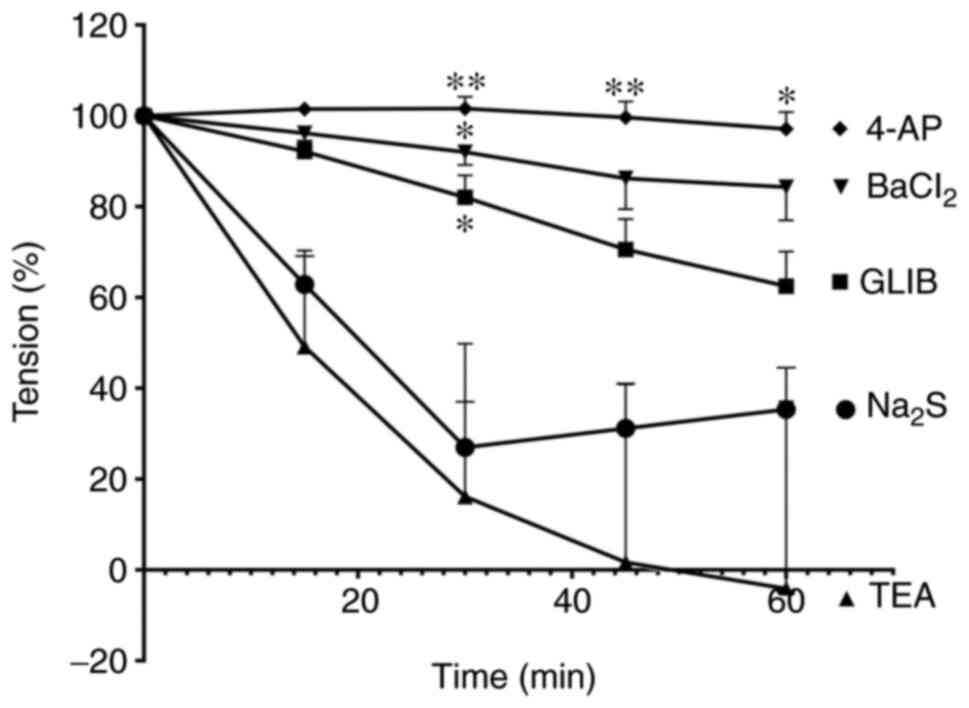
The role of K+ channels in
the NO-induced relaxation
Pre-incubation of mesenteric arteries with either
GLIB (0.1 µM; n=6) or TEA (1 mM; n=6) exhibited a significant
reduction of net SNP-induced vasorelaxation in mesenteric arteries
at all-time points (Fig. 4). In
contrast, vasorelaxation reduction by BaCl2 (1 mM; n=6)
and 4-AP (1 mM; n=6) was significant for 15-45 min; however,
reduction was not significant at 60 min compared with the SNP
treatment group.
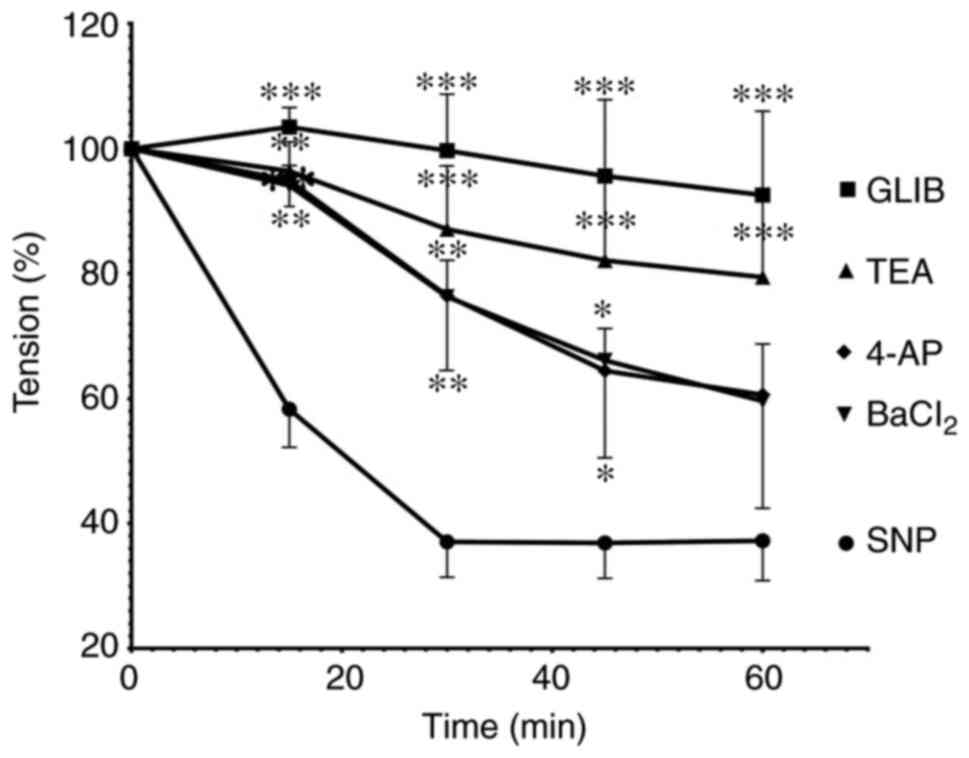 | Figure 4Time-dependent change of relaxation
responses to SNP in mesenteric arteries preincubated with GLIB (10
µM), TEA (1 mM), 4-AP (1 mM) and BaCl2 (1 mM).
SNP-induced vasorelaxation was significantly inhibited by GLIB,
TEA, 4-AP and BaCl2 pretreatment. *P<0.05,
**P<0.01 and ***P<0.001 vs. the SNP
treatment group. SNP, sodium nitroprusside; GLIB, glibenclamide;
TEA, tetraethylammonium; 4-AP, 4-aminopyridine; BaCl2,
barium chloride. |
The role of K+ channels in
the H2S-induced relaxation
The impairment of Na2S-inducing
relaxation in mesenteric arteries was significantly sustained at
time points 30, 45 and 60 mins following 4-AP incubation (n=6;
Fig. 5). GLIB (n=6) and
BaCl2 (n=6) reduced vasorelaxation responses produced by
Na2S only at time point 30 min. In contrast, TEA failed
to ameliorate the vasorelaxation response of Na2S.
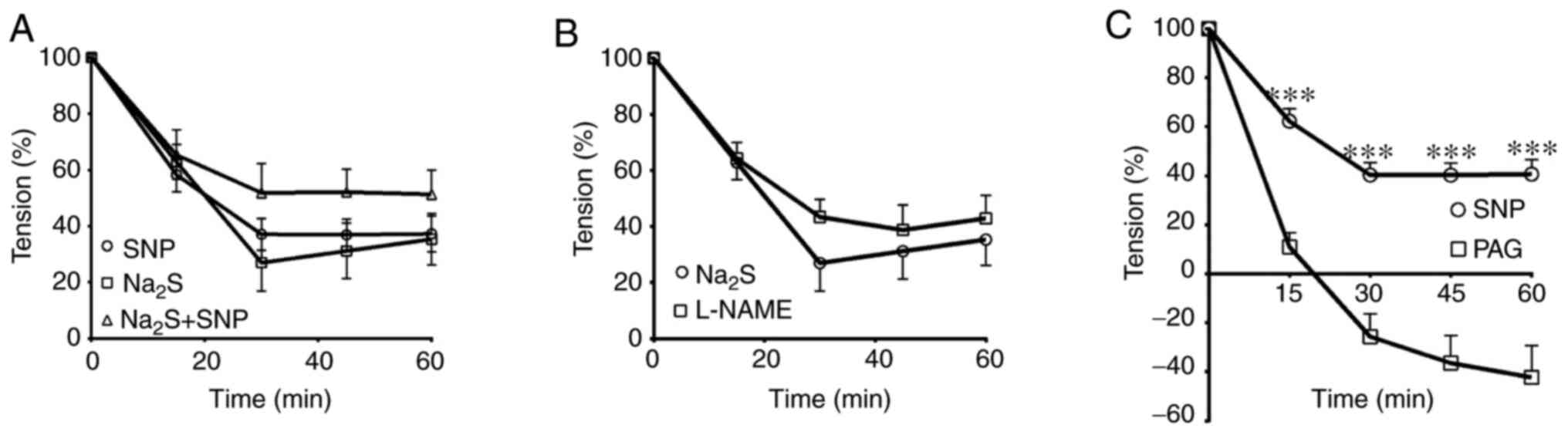
Interaction effects of SNP and
Na2S
The combination of SNP and Na2S did not
significantly alter the relaxation responses at any time point
compared with the relaxation induced by the application of SNP or
Na2S alone (Fig. 6A).
Additionally, pre-incubation of the arterial rings with L-NAME
(n=6) did not significantly change the extent of
Na2S-induced relaxation at any time point compared with
the NasS treatment group (Fig. 6B).
However, treating the mesenteric arterial rings with PAG (n=6)
significantly increased vasorelaxation induced by the SNP at
all-time points (Fig. 6C).
Discussion
The current study revealed that the endothelium
cells of patients with CRC were functioning normally since the
levels of serum endocan, which is an endothelial cell marker, were
significantly decreased compared with the controls (36). In this regard (37-39),
the reduction of endocan may be associated with chemotherapy, VEGF
receptor-2 kinase inhibitor treatment or the downregulation of
endocan expression, indicating that the expression of endocan is
associated with the development and differentiation of CRC
(36). Furthermore, it has been
demonstrated that endocan is associated with colon tumor size,
depth of invasion, lymph node metastasis, distant metastasis and
Dukes' staging (40). In addition
to endocan, there has been a growing interest in MDA as a marker of
oxidative stress in the progression of cancer (41). The present investigation
demonstrated that there were no changes in the serum MDA levels in
patients with CRC compared with controls. This is in contrast to a
previous study that reported a considerable elevation in serum MDA
levels in patients with CRC (42).
Subramanyam et al (43)
reported that chemotherapeutics normalized oxidative stress in
patients with CRC. Collectively, these results indicated that the
arteries in patients with CRC were intact and that their endothelia
were functioning properly.
Furthermore, the current study demonstrated that SNP
markedly relaxed mesenteric arteries in patients with CRC. A
previous study by (44) recorded
69% relaxation in the mesenteric arteries in experimental rats
compared with the control group, while a relaxation of 103% was
recorded in a study investigating human arm veins (45). To investigate the mechanism of
SNP-induced relaxation, the role of K+ channels was
investigated in the mesenteric arteries of patients with CRC.
The results of the present study revealed that
K+ channels exhibited a significant role of SNP-induced
relaxation in mesenteric arteries following pre-treatment with TEA,
GLIB, BaCl2 or 4-AP. All of these significantly
inhibited vasodilation. A previous study demonstrated that NO
activated several K+ channels of the small muscle cells
(SMCs) of mesenteric and cerebral arteries in rats and rabbits,
including ATP-sensitive K+ channels, and induced
membrane hyperpolarization by lowering
[Ca2+]i levels via the inhibition of
Ca2+ influx or Ca2+ release from
intracellular storage (46).
Furthermore, NO hyperpolarized arterial SMCs via the activation of
both KV and KCa channels on vascular SMCs
(VSMCs) in the rat superior mesenteric, coronary, cerebral and
large arteries (47-49)
through a cGMP-dependent mechanism, subsequently inhibiting the
depolarization of the evoked membrane and upsurge in
[Ca2+]i (50). The data obtained are compatible with
earlier findings, where it was observed that NO regulated VSMC
KIR currents (51). In
contrast, Hempelmann et al (52) reported that neither 4-AP nor
BaCl2 modulated NO-induced relaxation in the rat basilar
artery. This conclusion indicated that NO may exert vasodilation,
possibly by opening different K+ channels. Consequently,
it can be concluded that K+ channels serves a crucial
role in the vasodilation mechanism of NO.
To the best of our knowledge, in terms of the
influence of Na2S on arterial relaxation, the current
study was the first to observe the potency of Na2S in
relaxing the mesenteric arteries of patients with CRC. The present
study demonstrated that this relaxation was dependent on the
activation of KATP and KV channels. The
importance of KATP channel activation has been observed
in the mesenteric arteries of the human colon (53), rat arterial smooth muscle (54) and human mammary arteries (55). The latter mechanism occurred either
through the hyperpolarization of SMC membranes, which may close
voltage-gated Ca2+ channels (56), or through channel protein
sulfhydration (16). On the other
hand (53), concluded that
H2S relaxed pre-contracted human mesenteric arterial
rings in a concentration-dependent assay. Similarly, H2S
induced vasorelaxation in rat aortas, which was diminished by
KCNQ-type KV channel blockage (57).
In contrast, it has previously been shown that
KATP channels do not mediate H2S-induced
relaxation in the guinea-pig ileum or the trout urinary bladder
(58). Previous studies have
reported that KIR channels weakly participate in the
relaxation of mesenteric arteries in patients CRC and that the
mechanism of relaxation in rat aortas was mainly mediated by the
stimulation of KIR channels and subsequent
KIR-dependent hyperpolarization from endothelium to the
SMCs (59-61).
However, H2S was demonstrated to activate
BKCa (62),
IKCa and SKCa channels in endothelial cells
(16) and BKCa channels
in SMCs of mesenteric arteries (62) and cerebral arterioles (63). The results of the present study
reported that TEA did not alter the vasodilation of mesenteric
arteries induced by Na2S, indicating that KCa
may not be a considerable factor for H2S-induced
vasorelaxation. Similar results were observed by Tang et al
(64), who noted that different
KCa channel blockers were ineffective in the vascular
impact of H2S. Contrary to the current results, the
maximum relaxation of VSMC in rat (16,54,59)
and human mammary (55) arteries
induced by sodium hydrogen sulfide was significantly attenuated by
KCa channel blockers. Whereas both H2S and NO
are vasorelaxant factors with dissimilar mechanisms of action when
applied in combination (54), the
results of the current study reported that the combination of SNP
and Na2S donors did not significantly alter maximum
relaxation compared with the administration of SNP or
Na2S alone. The generation of a novel molecule (perhaps
nitrosothiol) via the combination of H2S and NO does not
relax blood vessels in vitro or in vivo (65). Consequently, the formation of this
unique molecule most likely signifies an approach to biological
inactivation or possibly sequestration of released NO (66). In contrast, rat aortic relaxation
was prolonged when the gas donors were combined (67). This synergistic action may be due to
the production of S-nitrosothiol (HSNO) and nitroxyl (HNO) as the
result of a chemical reaction between H2S and nitrite
(68), which releases NO and
polysulfides, and relaxes VSMCs through soluble guanylyl cyclase
activation (69).
Simultaneously, pre-incubation of L-NAME did not
alter Na2S-induced relaxation. Similar results have been
reported by (70,71). These authors demonstrated that
L-NAME did not modify the Na2S-induced relaxation in
isolated porcine irides. This conclusion indicated that endogenous
NO does not have an impact on the vasoactivity of the
H2S donor. In contrast, pre-incubation of the arterial
rings of patients with CRC with PAG did increase the relaxation
activity induced by the NO donor. The justification for this
reaction is associated with the activity of endogenous
H2S in inhibiting the action of NO. In a similar manner,
SNP-induced vasorelaxation in rat aortas and human internal mammary
arteries was diminished by a low concentration of H2S
through the suppression of NO action or inhibition of NO synthase
(56,64).
In 2015, Kashfi et al (72) investigated the effect of a NO- and
H2S-releasing hybrid on the growth properties of various
(HCT116 and NCM356) CRC cell lines. Additionally, in 2017, Oláh
et al (24) explored the
expression of NO- and H2S-generating enzymes in primary
CRC tissues and the HCT116 CRC cell line. A limitation of the
current study is that only the vasodilatory activity of NO and
H2S donors in the human mesenteric artery of patients
with CRC was investigated. Therefore, future studies should focus
on assessing the molecular signaling pathways in CRC tissues and
cell lines to provide a more comprehensive model of the expression
patterns of NO and H2S enzymes.
In conclusion, low endocan and normal MDA levels in
patients with CRC revealed that endothelial dysfunction and
oxidative stress were not involved in the pathogenesis of CRC.
Furthermore, the mechanism of NO and H2S-induced
mesenteric artery vasodilation was time- and K+
channel-dependent, as NO dilates mesenteric arteries via the
activation of KATP, KCa, KIR and
KV channels, while the vasodilation activity of
H2S is due to the modulation of KATP and
KV channels. Additionally, NO and H2S
interacted at the enzyme level and the activity of the CSE enzyme
inhibited the ability of exogenous NO in the vasodilatation
process.
Acknowledgements
The authors would like to thank Dr Saeb Gailany and
Dr Imad at the at Consultancy Medical City and Welfare hospitals
(Kurdistan region of Iraq, Iraq) for providing the human CRC
specimens. Finally, the authors would like to thank Dr Karim
Khoshnaw (Salahaddin University-Erbil), Mrs. Lynne Colley (Soran
University), Mrs. Marley Tinnock (UNDP) and Mr. Kumar Tiku (UNDP)
for their diligent proofreading of this article.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
AYH performed the experiments and co-wrote the
manuscript. IMM designed the experiments and co-wrote the paper.
ASS designed the experiments, analyzed data and co-wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was authorized and approved by the
Human Ethics Committee of Salahaddin University-Erbil. Patients
provided written informed consent.
Patient consent for publication
All patients provided written informed consent for
the publication of data in the current study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Arafa MA and Farhat KH: Recent diagnostic
procedures for colorectal cancer screening: Are they
cost-effective? Arab J Gastroenterol. 18:136–139. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Alhurry AM, Rezaianzadeh A,
Rahimikazerooni S, Abd-Zaid Akool M, Bahrami F, Saeedeh Shahidinia
S and Pourahmad M: A review of the incidence of colorectal cancer
in the Middle East. Ann Colorectal Res. 5(e46292)2017.
|
|
4
|
Khoshnaw N, Mohammed HA and Abdullah DA:
Patterns of cancer in Kurdistan-results of eight years cancer
registration in sulaymaniyah Province-Kurdistan-Iraq. Asian Pac J
Cancer Prev. 16:8525–8531. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hamza H and Rasul K: The epidemiology of
colorectal cancer in Erbil. Cancer Sci Res. 4:1–7. 2018.
|
|
6
|
Wang R: Two's company, three's a crowd:
Can H2S be the third endogenous gaseous transmitter? FASEB J.
16:1792–1798. 2002.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Yang G, Sener A, Ji Y, Pei Y and Pluth MD:
Gasotransmitters in biology and medicine: Molecular mechanisms and
drug targets. Oxid Med Cell Longev. 2016(4627308)2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Moncada S, Palmer RM and Higgs EA:
Biosynthesis of nitric oxide from L-arginine. A pathway for the
regulation of cell function and communication. Biochem Pharmacol.
38:1709–1715. 1989.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Palmer RM, Rees DD, Ashton DS and Moncada
S: L-arginine is the physiological precursor for the formation of
nitric oxide in endothelium-dependent relaxation. Biochem Biophys
Res Commun. 153:1251–1256. 1988.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Kabil O and Banerjee R: Enzymology of H2S
biogenesis, decay and signaling. Antioxid Redox Signal. 20:770–782.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Filipovic MR: Persulfidation
(S-sulfhydration) and H2S. Handb Exp Pharmacol. 230:29–59.
2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wang R, Cheng Y and Wu L: The role of
hydrogen sulfide as an endogenous vasorelaxant factor. In: Signal
transduction and the gasotransmitters. Wang R (ed). Humana Press,
pp323-332, 2004.
|
|
13
|
Nematollahi S, Nematbakhsh M,
Haghjooyjavanmard S, Khazaei M and Salehi M: Inducible nitric oxide
synthase modulates angiogenesis in ischemic hindlimb of rat. Biomed
Pap Med Fac Univ Palacky Olomouc Czech Repub. 153:125–129.
2009.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lüscher TF, Boulanger CM, Dohi Y and Yang
ZH: Endothelium-derived contracting factors. Hypertension.
19:117–130. 1992.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chadha PS, Liu L, Rikard-Bell M,
Senadheera S, Howitt L, Bertrand RL, Grayson TH, Murphy TV and
Sandow SL: Endothelium-dependent vasodilation in human mesenteric
artery is primarily mediated by myoendothelial gap junctions
intermediate conductance calcium-activated K+ channel and nitric
oxide. J Pharmacol Exp Ther. 336:701–708. 2011.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Mustafa AK, Sikka G, Gazi SK, Steppan J,
Jung SM, Bhunia AK, Barodka VM, Gazi FK, Barrow RK, Wang R, et al:
Hydrogen sulfide as endothelium-derived hyperpolarizing factor
sulfhydrates potassium channels. Circ Res. 109:1259–1268.
2011.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Szabó C and Papapetropoulos A: Hydrogen
sulphide and angiogenesis: Mechanisms and applications. Br J
Pharmacol. 164:853–865. 2011.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Tang G, Wu L, Liang W and Wang R: Direct
stimulation of K(ATP) channels by exogenous and endogenous hydrogen
sulfide in vascular smooth muscle cells. Mol Pharmacol.
68:1757–1764. 2005.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Coletta C, Papapetropoulos A, Erdelyi K,
Olah G, Módis K, Panopoulos P, Asimakopoulou A, Gerö D, Sharina I,
Martin E and Szabo C: Hydrogen sulfide and nitric oxide are
mutually dependent in the regulation of angiogenesis and
endothelium-dependent vasorelaxation. Proc Natl Acad Sci USA.
109:9161–9166. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Mistry RK and Brewer AC: Redox regulation
of gasotransmission in the vascular system: A focus on
angiogenesis. Free Radic Biol Med. 108:500–516. 2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Hu Q, Wu D, Ma F, Yang S, Tan B, Xin H, Gu
X, Chen X, Chen S, Mao Y and Zhu YZ: Novel angiogenic activity and
molecular mechanisms of ZYZ-803, a slow-releasing hydrogen
sulfide-nitric oxide hybrid molecule. Antioxid Redox Signal.
25:498–514. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kashfi K: The dichotomous role of
H2S in cancer cell biology? Deja vu all over again.
Biochem Pharmacol. 149:205–223. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Szabo C: Gasotransmitters in cancer: From
pathophysiology to experimental therapy. Nat Rev Drug Discov.
15:185–203. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Oláh G, Módis K, Törö G, Hellmich MR,
Szczesny B and Szabo C: Role of endogenous and exogenous nitric
oxide, carbon monoxide and hydrogen sulfide in HCT116 colon cancer
cell proliferation. Biochem Pharmacol. 149:186–204. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kather JN, Zöllner FG, Schad LR, Melchers
SM, Sinn HP, Marx A, Gaiser T and Weis CA: Identification of a
characteristic vascular belt zone in human colorectal cancer. PLoS
One. 12(e0171378)2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Sonveaux P: Provascular strategy:
Targeting functional adaptations of mature blood vessels in tumors
to selectively influence the tumor vascular reactivity and improve
cancer treatment. Radiother Oncol. 86:300–313. 2008.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Sosa V, Moline T, Somoza R, Paciucci R,
Kondoh H and ME LL: Oxidative stress and cancer: An overview.
Ageing Res Rev. 12:376–390. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Gill JG, Piskounova E and Morrison SJ:
Cancer, oxidative stress, and metastasis. Cold Spring Harb Symp
Quant Biol. 81:163–175. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Reuter S, Gupta SC, Chaturvedi MM and
Aggarwal BB: Oxidative stress, inflammation, and cancer: How are
they linked? Free Radic Biol Med. 49:1603–1616. 2010.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Perše M: Oxidative stress in the
pathogenesis of colorectal cancer: Cause or consequence? Biomed Res
Int. 2013(725710)2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Perillo B, Di Donato M, Pezone A, Di Zazzo
E, Giovannelli P, Galasso G, Castoria G and Migliaccio A: ROS in
cancer therapy: The bright side of the moon. Exp Mol Med.
52:192–203. 2020.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Hsu T, Nguyen-Tran HH and Trojanowska M:
Active roles of dysfunctional vascular endothelium in fibrosis and
cancer. J Biomed Sci. 26(86)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Orbegozo D, Rahmania L, Irazabal M,
Mendoza M, Annoni F, De Backer D, Creteur J and Vincent JL: Endocan
as an early biomarker of severity in patients with acute
respiratory distress syndrome. Ann Intensive Care.
7(93)2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ohkawa H, Ohishi N and Yagi K: Assay for
lipid peroxides in animal tissues by thiobarbituric acid reaction.
Anal Biochem. 95:351–358. 1979.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Furchgott RF and Zawadzki JV: The
obligatory role of endothelial cells in the relaxation of arterial
smooth muscle by acetylcholine. Nature. 288:373–376.
1980.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zuo L, Zhang SM, Hu RL, Zhu HQ, Zhou Q,
Gui SY, Wu Q and Wang Y: Correlation between expression and
differentiation of endocan in colorectal cancer. World J
Gastroenterol. 14:4562–4568. 2008.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hatfield KJ, Lassalle P, Leiva RA, Lindås
R, Wendelboe Ø and Bruserud Ø: Serum levels of endothelium-derived
endocan are increased in patients with untreated acute myeloid
leukemia. Hematology. 16:351–356. 2011.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Yang J, Yang Q, Yu S and Zhang X: Endocan:
A new marker for cancer and a target for cancer therapy. Biomed
Rep. 3:279–283. 2015.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Delehedde M, Devenyns L, Maurage CA and
Vives RR: Endocan in cancers: A lesson from a circulating dermatan
sulfate proteoglycan. Int J Cell Biol. 2013(705027)2013.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Pardo LA and Stuhmer W: The roles of K(+)
channels in cancer. Nat Rev Cancer. 14:39–48. 2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Gönenç A, Erten D, Aslan S, Akinci M,
Simşek B and Torun M: Lipid peroxidation and antioxidant status in
blood and tissue of malignant breast tumor and benign breast
disease. Cell Biol Int. 30:376–380. 2006.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Skrzydlewska E, Sulkowski S, Koda M,
Zalewski B, Kanczuga-Koda L and Sulkowska M: Lipid peroxidation and
antioxidant status in colorectal cancer. World J Gastroenterol.
11:403–406. 2005.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Subramanyam D, Subbaiah KV, Rajendra W and
Lokanatha V: Serum selenium concentration and antioxidant activity
in cervical cancer patients before and after treatment. Exp Oncol.
35:97–100. 2013.PubMed/NCBI
|
|
44
|
Plane F, Sampson LJ, Smith JJ and Garland
CJ: Relaxation to authentic nitric oxide and SIN-1 in rat isolated
mesenteric arteries: Variable role for smooth muscle
hyperpolarization. Br J Pharmacol. 133:665–672. 2001.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chalon S, Tejura B, Moreno H, Urae A,
Blaschke TF and Hoffman BB: Role of nitric oxide in isoprenaline
and sodium nitroprusside-induced relaxation in human hand veins. Br
J Clin Pharmacol. 47:91–98. 1999.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Brayden JE: Functional roles of
KATP channels in vascular smooth muscle. Clin Exp
Pharmacol Physiol. 29:312–316. 2002.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Félétou M: Calcium-activated potassium
channels and endothelial dysfunction: Therapeutic options? Br J
Pharmacol. 156:545–562. 2009.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Ko EA, Han J, Jung ID and Park WS:
Physiological roles of K+ channels in vascular smooth muscle cells.
J Smooth Muscle Res. 44:65–81. 2008.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Stankevicius E, Dalsgaard T, Kroigaard C,
Beck L, Boedtkjer E, Misfeldt MW, Nielsen G, Schjorring O, Hughes A
and Simonsen U: Opening of small and intermediate calcium-activated
potassium channels induces relaxation mainly mediated by
nitric-oxide release in large arteries and endothelium-derived
hyperpolarizing factor in small arteries from rat. J Pharmacol Exp
Ther. 339:842–850. 2011.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Jiang J, Thoren P, Caligiuri G, Hansson GK
and Pernow J: Enhanced phenylephrine-induced rhythmic activity in
the atherosclerotic mouse aorta via an increase in opening of
KCa channels: Relation to Kv channels and nitric oxide.
Br J Pharmacol. 128:637–646. 1999.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Schubert R, Krien U, Wulfsen I, Schiemann
D, Lehmann G, Ulfig N, Veh RW, Schwarz JR and Gago H: Nitric oxide
donor sodium nitroprusside dilates rat small arteries by activation
of inward rectifier potassium channels. Hypertension. 43:891–896.
2004.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Hempelmann RG, Seebeck J, Kruse ML,
Ziegler A and Mehdorn HM: Role of potassium channels in the
relaxation induced by the nitric oxide (NO) donor DEA/NO in the
isolated rat basilar artery. Neurosci Lett. 313:21–24.
2001.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Materazzi S, Zagli G, Nassini R, Bartolini
I, Romagnoli S, Chelazzi C, Benemei S, Coratti A, De Gaudio AR and
Patacchini R: Vasodilator activity of hydrogen sulfide (H2S) in
human mesenteric arteries. Microvasc Res. 109:38–44.
2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Zhao W, Zhang J, Lu Y and Wang R: The
vasorelaxant effect of H(2)S as a novel endogenous gaseous K(ATP)
channel opener. EMBO J. 20:6008–6016. 2001.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Webb GD, Lim LH, Oh VM, Yeo SB, Cheong YP,
Ali MY, El Oakley R, Lee CN, Wong PS, Caleb MG, et al: Contractile
and vasorelaxant effects of hydrogen sulfide and its biosynthesis
in the human internal mammary artery. J Pharmacol Exp Ther.
324:876–882. 2008.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Zhao W and Wang R: H(2)S-induced
vasorelaxation and underlying cellular and molecular mechanisms. Am
J Physiol Heart Circ Physiol. 283:H474–H480. 2002.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Köhn C, Schleifenbaum J, Szijártó IA,
Markó L, Dubrovska G, Huang Y and Gollasch M: Differential effects
of cystathionine-γ-lyase-dependent vasodilatory H2S in
periadventitial vasoregulation of rat and mouse aortas. PLoS One.
7(e41951)2012.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Teague B, Asiedu S and Moore PK: The
smooth muscle relaxant effect of hydrogen sulphide in vitro:
Evidence for a physiological role to control intestinal
contractility. Br J Pharmacol. 137:139–145. 2002.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Al-Magableh MR and Hart JL: Mechanism of
vasorelaxation and role of endogenous hydrogen sulfide production
in mouse aorta. Naunyn Schmiedebergs Arch Pharmacol. 383:403–413.
2011.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Doughty JM, Boyle JP and Langton PD:
Blockade of chloride channels reveals relaxations of rat small
mesenteric arteries to raised potassium. Br J Pharmacol.
132:293–301. 2001.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Salihi A: Activation of inward rectifier
potassium channels in high salt impairment of hydrogen
sulfide-induced aortic relaxation in rats. Physiol Pharmacol.
19:263–273. 2016.
|
|
62
|
Jackson-Weaver O, Osmond JM, Riddle MA,
Naik JS, Gonzalez Bosc LV, Walker BR and Kanagy NL: Hydrogen
sulfide dilates rat mesenteric arteries by activating endothelial
large-conductance Ca2+-activated K+ channels
and smooth muscle Ca2+ sparks. Am J Physiol Heart Circ
Physiol. 304:H1446–H1454. 2013.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Liang GH, Xi Q, Leffler CW and Jaggar JH:
Hydrogen sulfide activates Ca2+ sparks to induce
cerebral arteriole dilatation. J Physiol. 590:2709–2720.
2012.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Tang G, Wu L and Wang R: Interaction of
hydrogen sulfide with ion channels. Clin Exp Pharmacol Physiol.
37:753–763. 2010.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Whiteman M, Li L, Kostetski I, Chu SH,
Siau JL, Bhatia M and Moore PK: Evidence for the formation of a
novel nitrosothiol from the gaseous mediators nitric oxide and
hydrogen sulphide. Biochem Biophys Res Commun. 343:303–310.
2006.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Ali MY, Ping CY, Mok YY, Ling L, Whiteman
M, Bhatia M and Moore PK: Regulation of vascular nitric oxide in
vitro and in vivo; a new role for endogenous hydrogen sulphide? Br
J Pharmacol. 149:625–634. 2006.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Salihi AB, Shekha MS and Al-Habib OA:
Vasodilatory effects of nitric oxide, hydrogen sulfide and sulfur
dioxide in rats: Time-dependent interaction study. Prog Biol Sci.
6:19–30. 2016.
|
|
68
|
Filipovic MR, Miljkovic JL, Nauser T,
Royzen M, Klos K, Shubina T, Koppenol WH, Lippard SJ and
Ivanović-Burmazović I: Chemical characterization of the smallest
S-Nitrosothiol, HSNO; cellular cross-talk of H2S and
S-nitrosothiols. J Am Chem Soc. 134:12016–12027. 2012.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Cortese-Krott MM, Fernandez BO, Santos JL,
Mergia E, Grman M, Nagy P, Kelm M, Butler A and Feelisch M:
Nitrosopersulfide (SSNO(-)) accounts for sustained NO bioactivity
of S-nitrosothiols following reaction with sulfide. Redox Biol.
2:234–244. 2014.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Ohia SE, Opere CA, Monjok EM, Kouamou G,
Leday AM and Njie-Mbye YF: Role of hydrogen sulfide production in
inhibitory action of L-cysteine on isolated porcine irides. Curr
Eye Res. 35:402–407. 2010.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Monjok EM, Kulkarni KH, Kouamou G, McKoy
M, Opere CA, Bongmba ON, Njie YF and Ohia SE: Inhibitory action of
hydrogen sulfide on muscarinic receptor-induced contraction of
isolated porcine irides. Exp Eye Res. 87:612–616. 2008.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Kashfi K, Chattopadhyay M and Kodela R:
NOSH-sulindac (AVT-18A) is a novel nitric oxide- and hydrogen
sulfide-releasing hybrid that is gastrointestinal safe and has
potent anti-inflammatory, analgesic, antipyretic, anti-platelet,
and anti-cancer properties. Redox Biol. 6:287–296. 2015.PubMed/NCBI View Article : Google Scholar
|