Introduction
Retinoblastoma (RB) is an aggressive eye cancer that
occurs during retinal development in childhood, and it accounts for
2-4% of all childhood malignancies (1,2).
Moreover, the occurrence of RB is a very complicated process and it
can be present in a single eye or both eyes (3,4).
Although the treatment methods have improved over the past few
years, the mortality remains at ~70% in low- and middle-income
countries (5). Therefore, it is
important to improve the understanding of the pathogenesis of RB
and develop more effective treatment strategies.
Long non-coding RNAs (lncRNAs) are a class of
non-coding RNAs (ncRNAs) with >200 nucleotides (nts), which have
no protein-encoding ability (6).
lncRNAs have been reported to serve crucial roles in the occurrence
and progression of various cancer types (7,8).
Furthermore, several lncRNAs have been revealed to participate in
the regulation of RB. For example, knockdown of lncRNA Antisense
ncRNA in the INK4 locus inhibits RB cell invasion and induces
apoptosis via the ataxia-telangiectasia mutated/E2F transcription
factor 1 pathway (9). Colon cancer
associated transcript 1 has been demonstrated to promote RB cell
proliferation via sponging microRNA (miRNA/miR)-218-5p (10). It has also been revealed that H19
imprinted maternally expressed transcript suppresses RB progression
by targeting miR-17-92(11).
Previous studies have reported that Nuclear paraspeckle assembly
transcript 1 (NEAT1) plays an important role in different human
cancer type progression (12). For
example, NEAT1 promotes cell proliferation and invasion by
regulating the miR-211/High mobility group AT-hook 2 axis in breast
cancer (13). Moreover, NEAT1
promotes cell proliferation and invasion by targeting AKT/PI3K in
cervical carcinoma (14). However,
understanding of the role of NEAT1 in RB remains limited, and the
potential effects and mechanisms of NEAT1 in RB require further
investigation.
miRNAs are a group of small ncRNAs with 22-25 nts
that exert their functions by binding to the 3'untranslated region
(UTR) of their target gene (15).
Previous studies have reported that miRNAs are aberrantly expressed
in RB (16). Furthermore, miR-24 is
downregulated in RB tissues compared with healthy retinas tissues
(17). A recent study by Yu et
al (18) reported that miR-24
suppresses cell proliferation, migration and invasion in RB.
However, the mechanism underlying the role of miR-24-3p in the
development of RB is not fully understood.
Leucine-rich-α-2-glycoprotein1 (LRG1) is a
glycoprotein containing 312 amino acid residues (19). LRG1 has been reported to influence a
variety of biological processes, including proliferation, apoptosis
and invasion (20,21). In addition, previous studies have
revealed that the dysregulation of LRG1 is associated with the
progression of human cancer types, such as non-small cell lung
cancer (22), bladder cancer
(23) and ovarian cancer (24). Furthermore, Amer et al
(25) demonstrated that LRG1 is
elevated in RB, suggesting that LRG1 may be an important regulator
of RB progression.
Therefore, the aim of the present study was to
explore the function and underlying molecular mechanism of NEAT1 in
RB through functional and mechanistic analysis.
Materials and methods
Patients and tissue samples
RB tissue samples and adjacent healthy retina tissue
samples were collected from 20 patients with RB who underwent
surgery at Renmin Hospital, Hubei University of Medicine (Shiyan,
China) between October 2015 and March 2018. The patients were
within the ages of 8 months to 9 years, including 9 girls and 11
boys. The patients were diagnosed as having RB by two experienced
pathologists and none of the patients received any treatment before
surgery. All tissue samples were stored in liquid nitrogen
immediately after resection and kept at -80˚C until use. Written
informed consent was obtained from every patient and the study was
approved by the Ethics Committee of Renmin Hospital, Hubei
University of Medicine.
Cell culture and transfection
Human RB cell lines (Y79 and WERI-RB1) and normal
human retinal pigment epithelial cell line (ARPE-19) were obtained
from the American Type Culture Collection. RB cell lines SO-Rb50
and HXO-Rb44 were obtained from the Cell Bank of Type Culture of
Chinese Academy of Sciences. Y79, WERI-RB1, SO-Rb50 and HXO-Rb44
cells were cultured in RPMI 1640 medium (Gibco; Thermo Fisher
Scientific, Inc.). ARPE-19 cells were cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.). All mediums were supplemented with
10% FBS (HyClone; GE Healthcare Life Sciences), 100 U/ml penicillin
and 100 mg/ml streptomycin (HyClone; GE Healthcare Life Sciences).
Cells were cultured at 37˚C in a humidified incubator containing 5%
CO2.
Short hairpin RNA (shRNA) targeting NEAT1
(sh-NEAT1), shRNA targeting LRG1 (sh-LRG1) and their negative
control (sh-NC), pcDNA3.1-NEAT1 overexpression vector
(pcDNA-NEAT1), pcDNA3.1-LRG1 overexpression vector (pcDNA-LRG1) and
their control pcDNA3.1 empty vector (pcDNA-NC) were purchased from
Shanghai GenePharma Co., Ltd. miR-24-3p mimic (miR-24-3p;
5'-UGGCUCAGUUCAGCAGGAACAG-3' and 5'-GUUCCUGCUGAACUGAGCCAUU-3') and
control mimic (miR-NC; 5'-UUCUCCGAACGUGUCACGUTT-3' and
5'-ACGUGACACGUUCGGAGAATT-3'), miR-24-3p inhibitor (anti-miR-24-3p;
5'-CUGUUCCUGCUGAACUGAGCCA-3') and its control (anti-miR-NC;
5'-CAGUACUUUUGUGUAGUACAA-3') were also purchased from Shanghai
GenePharma Co., Ltd. Y79 and WERI-RB1 cells were seeded into
24-well plates at a density of 2.0x104 cells/well,
following which 50 nM synthetic oligonucleotides or 2 µg vectors
were transfected into the cells using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. Cells were collected after 48 h for
further experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from tissues and cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocols. To analyze the
expression of lncRNA and mRNA, the RNAs were reverse transcribed
into cDNAs using PrimeScript™ RT reagent kit (Takara, Biotechnology
Co., Ltd.) using the temperature protocol consisting of 50˚C for 15
min followed by 85˚C for 5 sec. To analyze the expression of miRNA,
cDNAs were synthesized by All-in-One miRNA First strand cDNA
Synthesis kit (GeneCopoeia, Inc.) using the temperature protocol
consisting of 42˚C for 50 min followed by 70˚C for 15 min. qPCR was
then performed using SYBR® Green qPCR Master Mix
(Bio-Rad Laboratories, Inc.) with GAPDH (for lncRNA and mRNA) or U6
(for miRNA) as an endogenous control. The thermocycling conditions
of the qPCR reaction were: Initial denaturation at 95˚C for 5 min,
followed by 40 cycles of 95˚C for 30 sec and 60˚C for 45 sec. The
relative expression levels of NEAT1, miR-24-3p and LRG1 were
calculated via the 2-ΔΔCq method (26). The primers sequences were listed as
follows: NEAT forward, 5'-GTGGCTGTTGGAGTCGGTAT-3' and reverse,
5'-TAACAAACCACGGTCCATGA-3'; miR-24-3p forward,
5'-GCCGAGTGGCTCAGTTCAGC-3' and reverse, 5'-CTCAACTGGTGTCGTGGA-3';
LRG1 forward, 5'-CCATGTCAGTGTGCAGATTC-3' and reverse,
5'-AAGAGTGAGAGGTGGAAGAG-3'; U6 forward, 5'-CTCGCTTCGGCAGCACA-3' and
reverse, 5'-AACGCTTCACGAATTTGCGT-3'; and GAPDH forward,
5'-TCGGAGTCAACGGATTTGGT-3' and reverse,
5'-TTCCCGTTCTCAGCCTTGAC-3'.
Transwell assay
Cell migration and invasion abilities were evaluated
using Transwell chambers (8-µm pore size; EMD Millipore). Cell
suspension (2.0x105 cells/ml) was prepared in DMEM
(Gibco; Thermo Fisher Scientific, Inc.) without serum. For the
Transwell migration assay, ~500 µl cell suspension was seeded into
the upper chamber and 300 µl DMEM (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS (HyClone; GE Healthcare Life
Science) was added into the lower chamber. After culturing for 24 h
at 37˚C, cells that did not pass through the membrane were scraped
using a cotton swab. Migrated cells that passed through the
membrane were fixed in 90% ethyl alcohol for 15 min at room
temperature, stained with 0.1% crystal violet (Sigma-Aldrich; Merck
KGaA) for 20 min at room temperature, observed and counted under a
light microscope (Olympus Corporation) at x100 magnification.
For the Transwell invasion assay, the upper chamber
was pre-coated with Matrigel (BD Biosciences) for 30 min at 37˚C,
and the subsequent steps were the same as those in the cell
migration assay.
Dual-luciferase reporter assay
The potential binding sites between NEAT1 and
miR-24-3p, as well as miR-24-3p and LRG1 3'UTR were predicted using
the online software starBase2.0 (http://starbase.sysu.edu.cn/index.php) and MiRcode 11
(http://mircode.org/), respectively. To assess the
prediction, the sequences of NEAT1 and LRG1 3'UTR containing the
potential wild-type (WT) or mutant (MUT) binding sites of miR-24-3p
were amplified via PCR and cloned into pGL3-control luciferase
reporter vectors (Promega Corporation), named as WT-NEAT1,
MUT-NEAT1, LRG1 3'UTR-WT and LRG1 3' UTR-MUT. These procedures were
conducted by Shanghai Sangon Co., Ltd. Y79 and WERI-RB1 cells
(5.0x104 cells) were then seeded into 24-well plates
before 50 nM miR-24-3p mimic or miR-NC and 100 ng WT-NEAT1,
MUT-NEAT1, LRG1 3'UTR-WT or LRG1 3'UTR-MUT were co-transfected into
Y79 and WERI-RB1 cells using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to
manufacturer's instructions. After incubation for 48 h, firefly
luciferase activity and Renilla luciferase activity were
detected using a Dual Luciferase Reporter assay kit (Promega
Corporation). Renilla luciferase activity was normalized to
firefly luciferase activity.
Western blotting
Total protein was extracted from tissues and cells
using RIPA lysis buffer (Beijing Solarbio Science & Technology
Co., Ltd.) and protein concentrations were measured using the
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology) according to the manufacturer's instructions. In
total, 20 µg proteins were separated via 10% SDS-PAGE and
transferred to PVDF membranes (EMD Millipore). The membranes were
blocked in 5% non-fat milk for 1 h at room temperature.
Subsequently, the membranes were immunoblotted with primary
antibodies: E-cadherin (cat. no. ab231303; 1:2,000; Abcam),
N-cadherin (cat. no. ab76011; 1:5,000; Abcam), matrix
metalloproteinase (MMP)9 (cat. no. ab38898; 1:1,000; Abcam), LRG1
(cat. no. ab170953; 1:5,000; Abcam) or GAPDH (cat. no. ab9485;
1:2,500; Abcam) at 4˚C overnight. After the membranes were washed
three times with TBS-1% Tween-20, the membranes were incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG secondary
antibody (cat. no. ab6721; 1:5,000; Abcam) for 1 h at room
temperature. All antibodies used in the current study were
purchased from Cell Signaling Technology, Inc.. The immunoreactive
bands were detected using enhanced chemiluminescence (Pierce;
Thermo Fisher Scientific, Inc.) and analyzed using ImageJ v1.8.0
software (National Institutes of Health). GAPDH was used as an
internal reference.
Statistical analysis
All data in the present study were obtained from
three independent experiments, analyzed by GraphPad Prism 7
software (GraphPad Software, Inc.) and are presented as the mean ±
SD. Paired Student's t-test was used to analyze the difference
between two groups, while one-way ANOVA followed by Sidak's test or
Dunnett's test were used to analyze the differences between
multiple groups. The correlations between NEAT1 and miR-24-3p,
miR-24-3p and LRG1 mRNA expression levels were analyzed by
Pearson's correlation analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
NEAT1 is upregulated and miR-24-3p is
downregulated in RB tissues and cell lines
To investigate the potential roles of NEAT1 and
miR-24-3p in RB progression, the expression levels of NEAT1 and
miR-24-3p were detected in RB tissues via RT-qPCR. It was revealed
that NEAT1 expression was significantly increased (P<0.001) and
miR-24-3p expression was significantly decreased (P<0.001) in RB
tissues compared with adjacent healthy tissues (Fig. 1A and B). Furthermore, Pearson's correlation
analysis results suggested that the expression of miR-24-3p
exhibited a moderate negatively correlation with the expression of
NEAT1 in RB tissues (P<0.05; r=-0.455; Fig. 1C).
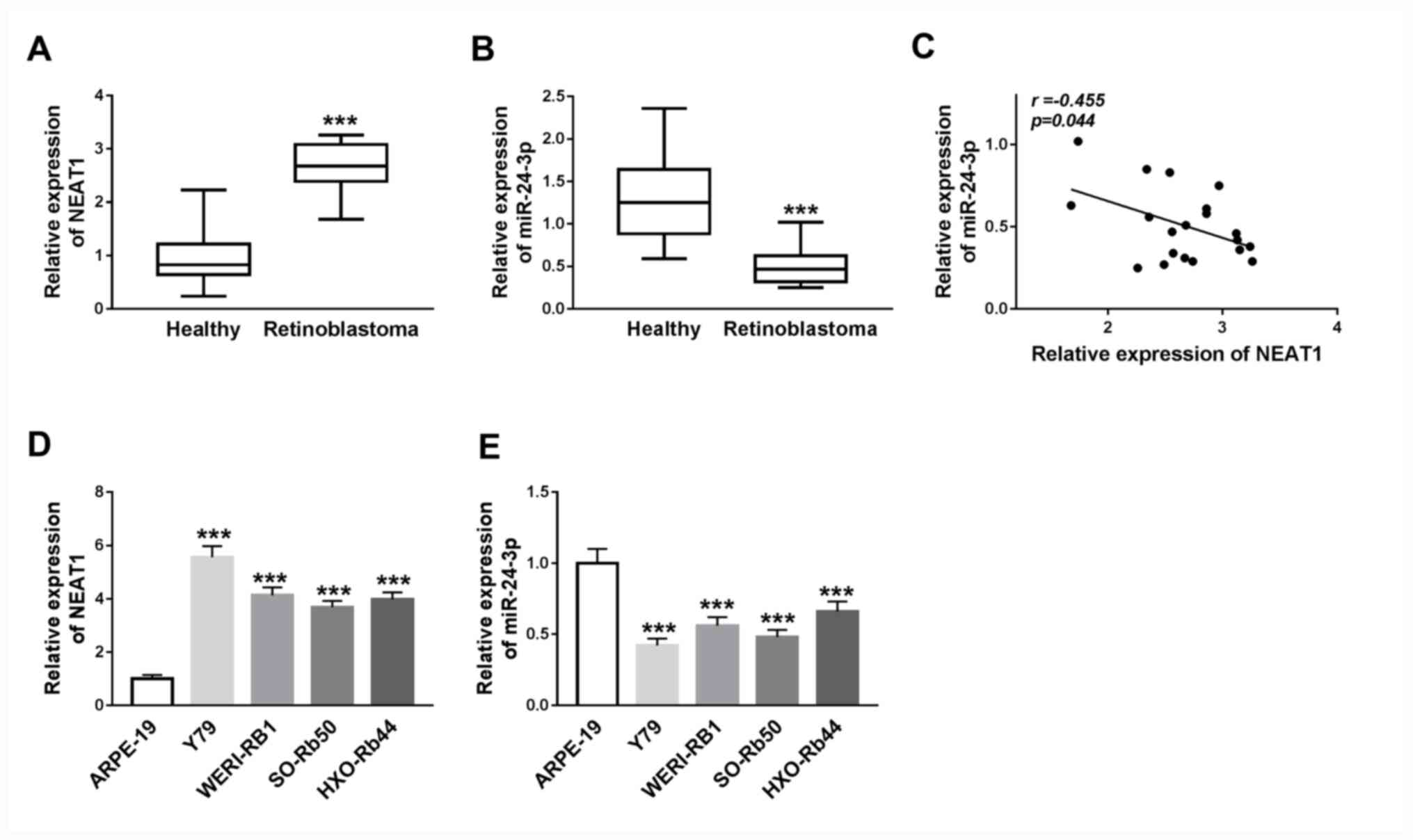 | Figure 1NEAT1 expression is upregulated and
miR-24-3p expression is downregulated in RB tissues and cells. (A)
Expression levels of NEAT1 in RB tissues and adjacent healthy
tissues were determined via RT-qPCR. ***P<0.001 vs.
Healthy. (B) Expression levels of miR-24-3p in RB tissues and
adjacent healthy tissues were determined via RT-qPCR.
***P<0.001 vs. Healthy. (C) Correlation between NEAT1
and miR-24-3p expression was analyzed using Pearson's correlation
analysis. (D) NEAT1 expression in Y79, WERI-RB1, SO-Rb50, HXO-Rb44
and ARPE-19 cell lines was detected via RT-qPCR. (E) miR-24-3p
expression in Y79, WERI-RB1, SO-Rb50, HXO-Rb44 and ARPE-19 cell
lines was detected via RT-qPCR. ***P<0.001 vs.
ARPE-19. RT-qPCR, reverse transcription-quantitative PCR; miR,
microRNA; NEAT1, nuclear paraspeckle assembly transcript 1; RB,
retinoblastoma. |
The expression levels of NEAT1 and miR-24-3p were
subsequently detected in RB cell lines (Y79, WERI-RB1, SO-Rb50 and
HXO-Rb44) and the normal human retinal pigment epithelial cell line
(ARPE-19) via RT-qPCR. NEAT1 expression was significantly
upregulated, whilst miR-24-3p was significantly downregulated, in
Y79, WERI-RB1, SO-Rb50 and HXO-Rb44 cells compared with that in
ARPE-19 cells (all P<0.001; Fig.
1D and E). Thus, it was
indicated that the dysregulation of NEAT1 and miR-24-3p may serve
vital roles in RB progression.
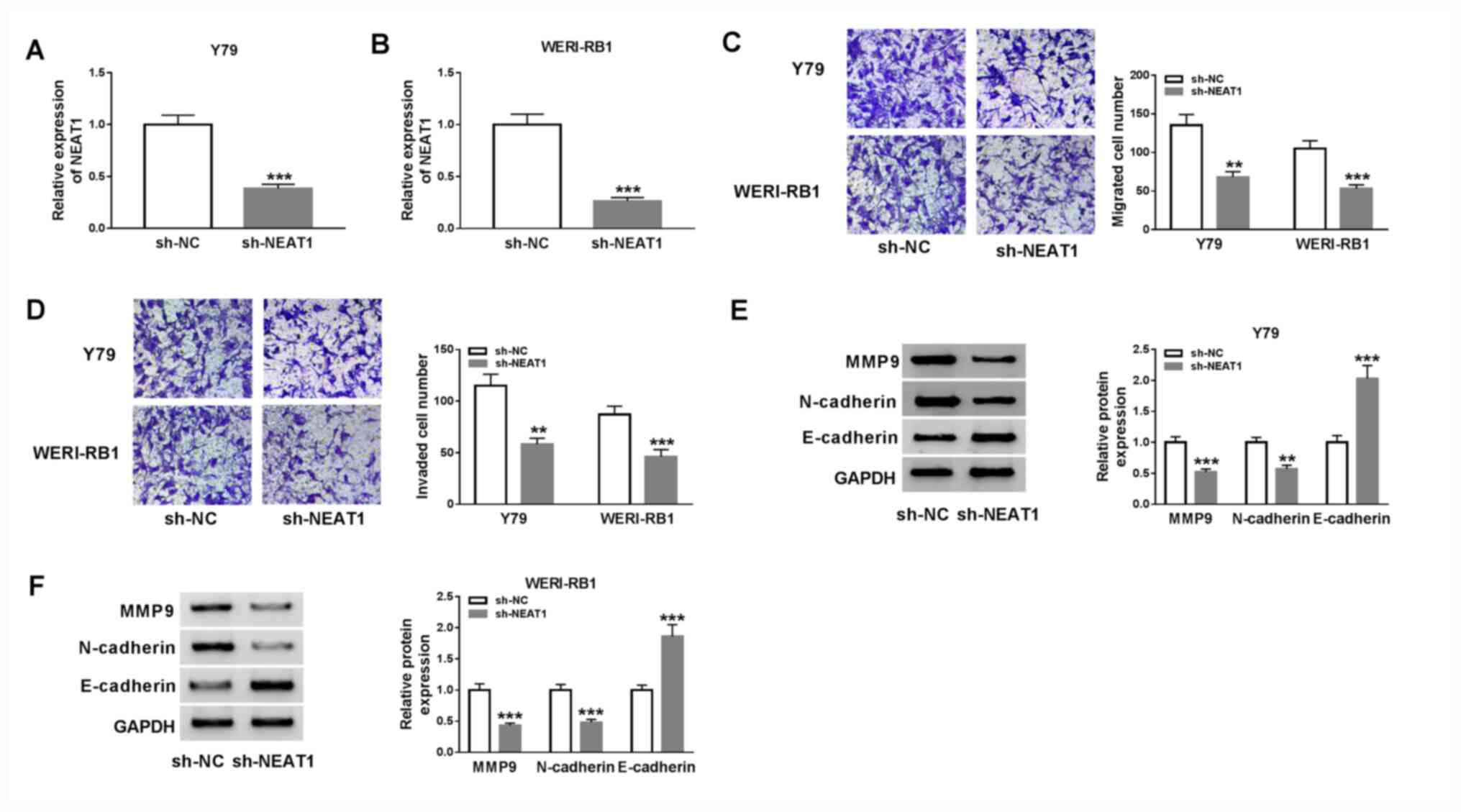
Subsequently, a Transwell assay was performed to
examine cell migration and invasion. The results demonstrated that
migration (Y79 cells, P<0.01; WERI-RB1 cells, P<0.001) and
invasion (Y79 cells, P<0.01; WERI-RB1 cells, P<0.001) of Y79
and WERI-RB1 cells were significantly decreased after the knockdown
of NEAT1 (Fig. 2C and D).
It has been previously reported that the migratory
and invasive phenotypes of tumor cells are gained via the process
of epithelial-mesenchymal transition (EMT) (27,28).
Therefore, the effects of NEAT1 on the protein expression levels of
EMT markers, including MMP9, N-cadherin and E-cadherin, were
examined via western blot analysis. The results indicated that
NEAT1 knockdown resulted in a significant decrease in MMP9 (Y79
cells, P<0.001; WERI-RB1 cells, P<0.001) and N-cadherin (Y79
cells, P<0.01; WERI-RB1 cells, P<0.001) expression levels,
and a significant increase in E-cadherin (Y79 cells, P<0.001;
WERI-RB1 cells, P<0.001) expression in both Y79 and WERI-RB1
cells (Fig. 2E and F). Collectively, it was indicated that
knockdown of NEAT1 may suppress cell migration, invasion and the
EMT pathway in RB.
NEAT1 negatively regulates miR-24-3p
expression via a direct interaction in RB cells
Based on the aberrant upregulation of NEAT1 and
downregulation of miR-24-3p in RB cells, it was speculated that
miR-24-3p may represent a target of NEAT1. According to the
bioinformatics software starBase v2.0, NEAT1 was predicted to
contain the putative binding sites of miR-24-3p (Fig. 3A), indicating that miR-24-3p may be
targeted by NEAT1.
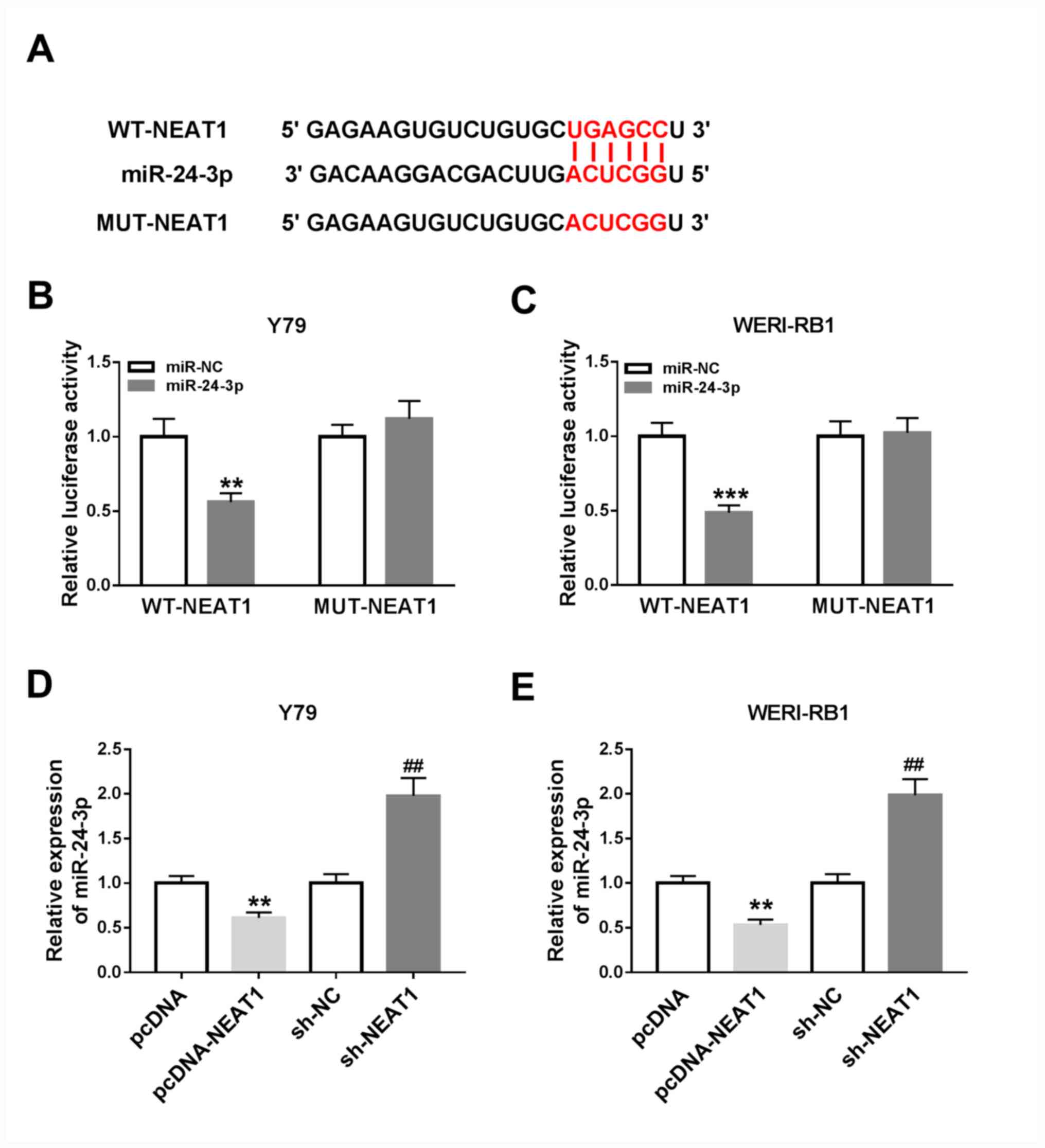 | Figure 3NEAT1 directly targets miR-24-3p and
negatively regulates miR-24-3p expression in RB cells. (A)
Predicted binding sites between miR-24-3p and NEAT1. Luciferase
activities in (B) Y79 and (C) WERI-RB1 cells co-transfected with
miR-24-3p or miR-NC and WT-NEAT1 or MUT-NEAT1.
**P<0.01 vs. miR-NC, ***P<0.001 vs.
miR-NC. The expression level of miR-24-3p in (D) Y79 and (E)
WERI-RB1 cells transfected with pcDNA, pcDNA-NEAT1, sh-NC or
sh-NEAT1 was detected via reverse transcription-quantitative PCR.
**P<0.01 vs. pcDNA, ##P<0.01 vs. sh-NC.
miR, microRNA; NEAT1, nuclear paraspeckle assembly transcript 1;
RB, retinoblastoma; sh, short hairpin RNA; NC, negative control;
EMT, epithelial-to-mesenchymal transition; WT, wild-type; MUT,
mutant. |
To assess this prediction, a dual-luciferase
reporter assay was conducted. It was revealed that the luciferase
activities were significantly inhibited in Y79 and WERI-RB1 cells
co-transfected with WT-NEAT1 and miR-24-3p, compared with cells
co-transfected with WT-NEAT1 and miR-NC (Y79 cells, P<0.01;
WERI-RB1 cells, P<0.001), while the luciferase activities were
not affected in the MUT-NEAT1 groups (Fig. 3B and C).
It was demonstrated that pcDNA-NEAT1 transfection
resulted in a significant increase in NEAT1 expression in Y79 and
WERI-RB1 cells, suggesting that pcDNA-NEAT1 was successfully
transfected (Fig. S1A).
Subsequently, the expression of miR-24-3p in pcDNA-NEAT1-, pcDNA-,
sh-NC- or sh-NEAT1-transfected Y79 and WERI-RB1 cells was detected
via RT-qPCR. It was revealed that miR-24-3p expression was
significantly decreased following NEAT1 overexpression and
significantly increased following NEAT1 knockdown in both Y79 and
WERI-RB1 cells (all P<0.01; Fig.
3D and E). Thus, it was
speculated that NEAT1 negatively regulates miR-24-3p expression by
directly interaction.
miR-24-3p inhibition restores the
inhibitory effects of NEAT1 silencing on cell migration, invasion
and the EMT process in RB cells
To further examine whether NEAT1 regulates RB cell
migration, invasion and EMT, miR-24-3p mimic and anti-miR-24-3p
were successfully transfected into Y79 and WERI-RB1 cells (Fig. S1B and C).
Next, Y79 and WERI-RB1 cells were transfected with
either miR-24-3p, miR-NC, sh-NEAT1, sh-NC, sh-NEAT1 + anti-NC or
sh-NEAT1 + anti-miR-24-3p. The Transwell assay results indicated
that miR-24-3p overexpression or NEAT1 knockdown significantly
suppressed migration and invasion abilities in Y79 and WERI-RB1
cells (all P<0.001). Moreover, inhibition of miR-24-3p could
eliminate the inhibitory effects of NEAT1 knockdown on the
migration and invasion of these cells (all P<0.01; Fig. 4A and B).
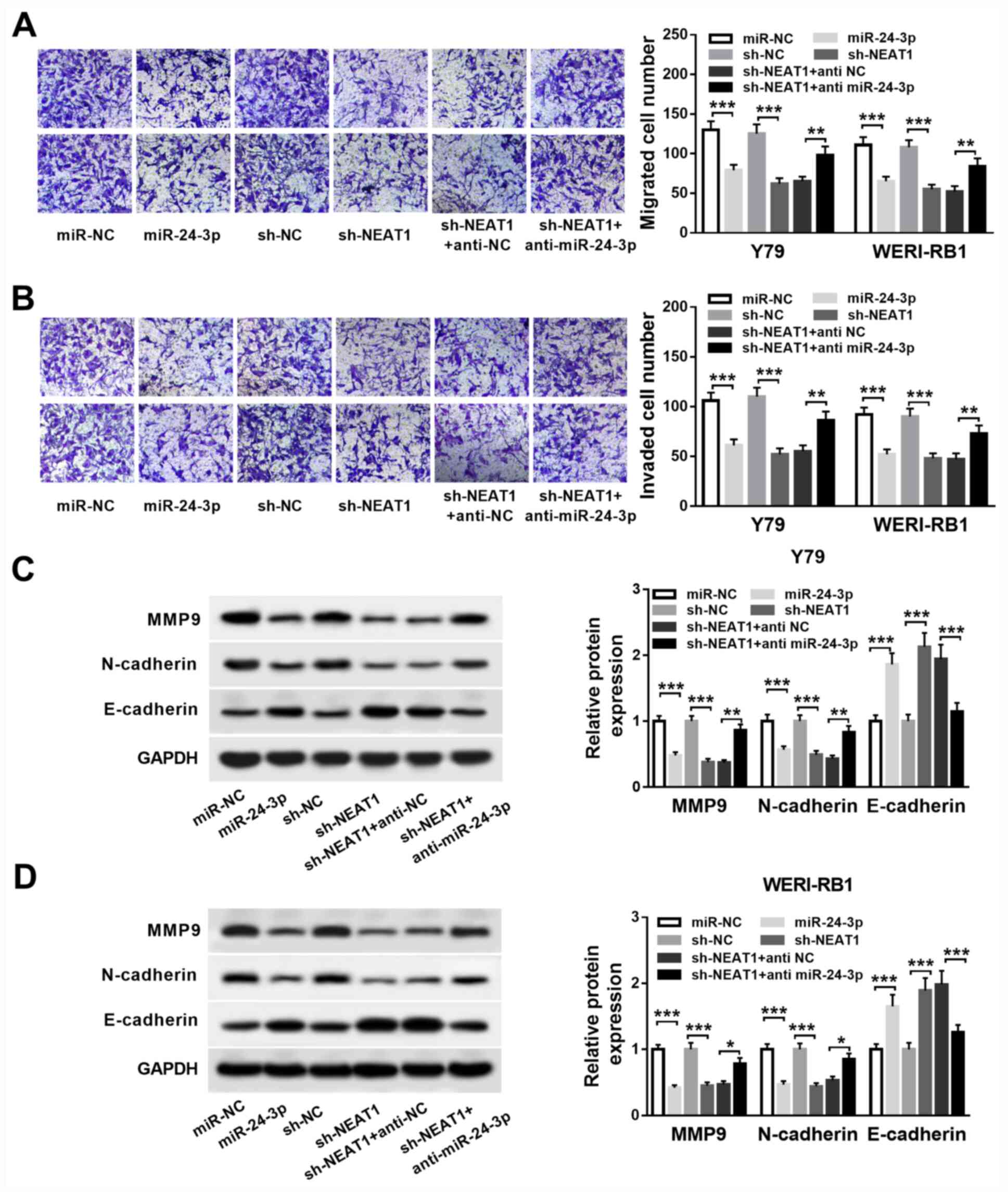 | Figure 4NEAT1 represses RB cell migration,
invasion and EMT via targeting miR-24-3p. Y79 and WERI-RB1 cells
were treated with miR-24-3p, miR-NC, sh-NEAT1, sh-NC, sh-NEAT1 +
anti NC or sh-NEAT1 + anti miR-24-3p. (A) Migrated and (B) invaded
cell numbers of Y79 and WERI-RB1 cells were evaluated by Transwell
assay. Protein expression levels of MMP9, N-cadherin and E-cadherin
in (C) Y79 and (D) WERI-RB1 cells were detected by western blot
analysis. *P<0.05, **P<0.01 and
***P<0.001. miR, microRNA; NEAT1, nuclear paraspeckle
assembly transcript 1; RB, retinoblastoma; MMP, matrix
metalloproteinase; sh, short hairpin RNA; NC, negative control;
EMT, epithelial-to-mesenchymal transition. |
Western blotting was then conducted to detect the
protein expression levels of EMT markers MMP9, N-cadherin and
E-cadherin. It was demonstrated that the expression levels of MMP9
and N-cadherin were significantly decreased (all P<0.001), while
the expression of E-cadherin was significantly increased (all
P<0.001) via miR-24-3p overexpression or NEAT1 knockdown in Y79
and WERI-RB1 cells. Furthermore, co-transfection with the miR-24-3p
inhibitor reversed the effects of NEAT1 knockdown on MMP9 (Y79
cells, P<0.01; WERI-RB1 cells, P<0.05), N-cadherin (Y79
cells, P<0.01; WERI-RB1 cells, P<0.05) and E-cadherin (Y79
cells, P<0.001; WERI-RB1 cells: P<0.001) expression levels in
Y79 and WERI-RB1 cells (Fig. 4C and
D). Therefore, NEAT1 knockdown may
suppress cell migration, invasion and the EMT process via targeting
miR-24-3p in RB.
NEAT1 regulates LRG1 expression via
sponging miR-24-3p in RB cells
To investigate the potential mechanism of miR-24-3p
in RB cell migration and invasion, an online software miRcode was
used to predict the potential binding sites of miR-24-3p. LRG1 was
predicted to be a target gene of miR-24-3p (Fig. 5A). Then, the mRNA expression level
of LRG1 was detected via RT-qPCR and it was demonstrated that LRG1
mRNA was significantly upregulated in RB tissues compared with
corresponding healthy tissues (P<0.001; Fig. 5B). Furthermore, there was an inverse
correlation between LRG1 mRNA expression and miR-24-3p expression
in RB tissues (P<0.01; r=-0.585; Fig. 5C). Western blotting results
indicated that the protein expression level of LRG1 was also
elevated in RB tissues compared with healthy tissues (P<0.001;
Fig. 5D).
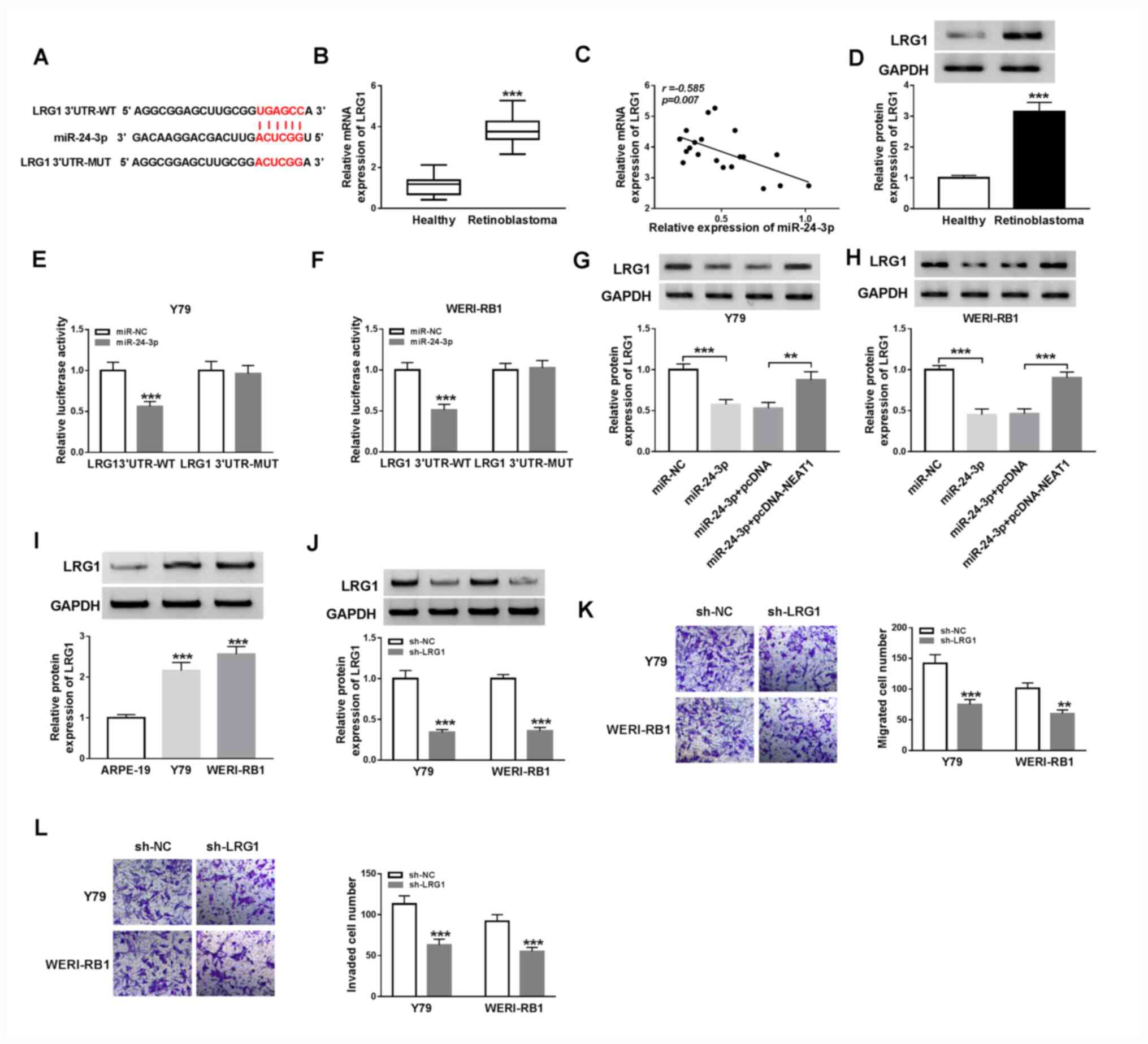 | Figure 5NEAT1 upregulates LRG1 expression via
miR-24-3p in RB cells. (A) Potential binding sites between
miR-24-3p and the 3'UTR of LRG1. (B) mRNA expression levels of LRG1
in RB tissues and healthy tissues were determined via reverse
transcription-quantitative PCR. ***P<0.001 vs.
healthy. (C) Correlation between LRG1 and miR-24-3p was determined
using Pearson's correlation analysis. (D) Protein expression levels
of LRG1 in RB tissues and healthy tissues were detected via western
blot analysis. ***P<0.001 vs. healthy. Luciferase
activities were detected in (E) Y79 and (F) WERI-RB1 cells
co-transfected with LRG1 3'UTR-WT or LRG1 3'UTR-MUT and miR-NC or
miR-24-3p. ***P<0.001 vs. miR-NC. Protein expression
levels of LRG1 in (G) Y79 and (H) WERI-RB1 cells treated with
miR-24-3p, miR-24-3p + pcDNA-NEAT1 or the corresponding controls
were measured via western blotting ***P<0.001 vs.
miR-NC, **P<0.01 and ***P<0.001 vs.
miR-24-3p+pcDNA. (I) Protein expression level of LRG1 in ARPE-19,
Y79 and WERI-RB1 cells was detected via western blotting.
***P<0.001 vs. ARPE-19. (J) LRG1 protein expression
in Y79 and WERI-RB1 cells transfected with sh-NC or sh-LRG1.
***P<0.001 vs. sh-NC. (K) Migratory and (L) invasive
abilities of Y79 and WERI-RB1 cells were evaluated via Transwell
assays. **P<0.01 and ***P<0.001 vs.
sh-NC. miR, microRNA; NEAT1, nuclear paraspeckle assembly
transcript 1; RB, retinoblastoma; sh, short hairpin RNA; NC,
negative control; WT, wild-type; MUT, mutant; LRG1, leucine-rich
α-2-glycoprotein; UTR, untranslated region. |
To assess the targeting association between
miR-24-3p and LRG1, a dual-luciferase reporter assay was performed
by transfecting miR-24-3p or miR-NC and LRG1 3'UTR-WT or LRG1
3'UTR-MUT into Y79 and WERI-RB1 cells. The luciferase activities
were significantly suppressed in Y79 and WERI-RB1 cells
co-transfected with LRG1 3'UTR-WT and miR-24-3p compared with LRG1
3'UTR-WT and miR-NC (both P<0.001), while luciferase activities
were not significantly affected in the LRG1 3'UTR-MUT groups
(Fig. 5E and F).
Subsequently, the protein expression levels of LRG1
in Y79 and WERI-RB1 cells transfected with miR-24-3p, miR-24-3p +
pcDNA-NEAT1 or the corresponding controls was examined via western
blot analysis. It was demonstrated that miR-24-3p overexpression
significantly suppressed the expression of LRG1 (both P<0.001),
while NEAT1 overexpression reversed this effect (Y79 cells,
P<0.05; WERI-RB1 cells, P<0.001; Fig. 5G and H). Moreover, the protein expression level
of LRG1 was elevated in Y79 and WERI-RB1 cells compared with
ARPE-19 cells (both P<0.001; Fig.
5I). sh-LRG1 was then transfected into Y79 and WERI-RB1 cells
before transfection efficiency was evaluated by western blotting.
sh-LRG1 transfection led to a significant reduction in LRG1 protein
expression in Y79 and WERI-RB1 cells compared with that in the
sh-NC groups (both P<0.001; Fig.
5J). In addition, it was discovered that knockdown of LRG1
caused a significant decrease in the migration (both P<0.001)
and invasion (Y79 cells, P<0.01; WERI-RB1 cells, P<0.001)
abilities of Y79 and WERI-RB1 cells (Fig. 5K and L). Thus, it was indicated that NEAT1 may
positively regulate LRG1 expression via the targeting of miR-24-3p
in RB cells.
NEAT1 modulates RB cell migration,
invasion and EMT by upregulating LRG1 expression via sponging
miR-24-3p
In order to examine the regulatory effects of NEAT1,
miR-24-3p and LRG1 in RB progression, pcDNA-LRG1, pcDNA-LRG1 +
miR-24-3p, pcDNA-LRG1 + miR-24-3p + pcDNA-NEAT1 or their matched
controls were transfected into Y79 and WERI-RB1 cells. The
transfection efficiency of pcDNA-LRG1 was determined via western
blotting, and it was identified that the protein expression level
of LRG1 was significantly increased in Y79 and WERI-RB1 cells,
which demonstrated the success of transfection (Fig. S1D).
Transwell assay results suggested that the migratory
(both P<0.001) and invasive (both P<0.001) abilities of Y79
and WERI-RB1 cells were significantly enhanced following pcDNA-LRG1
transfection, and the effects were reversed following miR-24-3p
overexpression (all P<0.01). However, the effects of pcDNA-LRG1
+ miR-24-3p on cell migration (both P<0.001) and invasion (Y79
cells, P<0.05; WERI-RB1 cells, P<0.05) were reversed by the
overexpression of NEAT1 (Fig. 6A
and B).
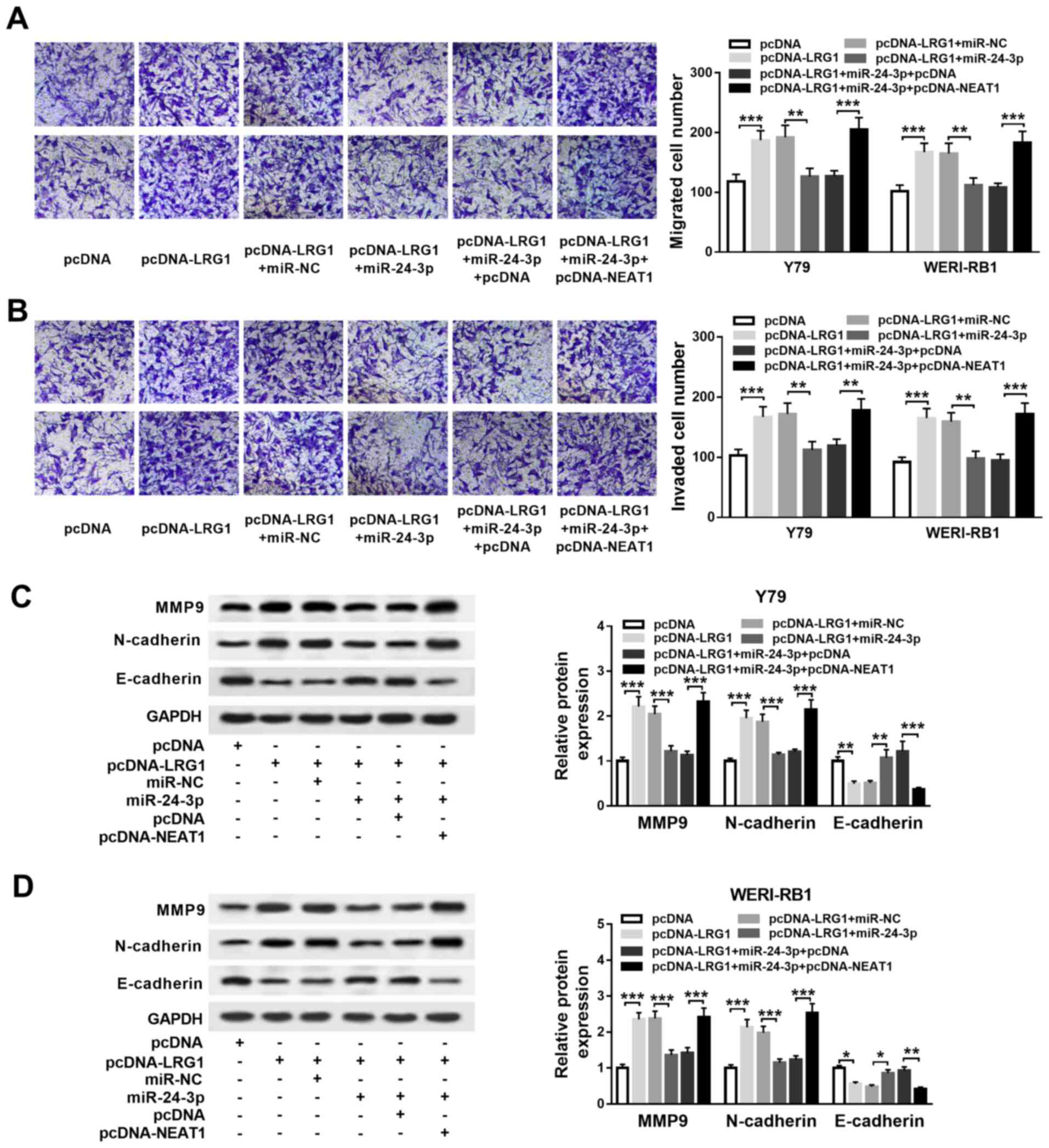 | Figure 6NEAT1 affects RB cell migration,
invasion and EMT by upregulating LRG1 expression via the sponging
of miR-24-3p. Y79 and WERI-RB1 cells were divided into six groups
and transfected with the following: pcDNA-LRG1, pcDNA, pcDNA-LRG1 +
miR-24-3p, pcDNA-LRG1 + miR-NC, pcDNA-LRG1 + miR-24-3p +
pcDNA-NEAT1 or pcDNA-LRG1 + miR-24-3p + pcDNA. (A) Migration and
(B) invasion of Y79 and WERI-RB1 cells were evaluated via a
Transwell assay. MMP9, N-cadherin and E-cadherin protein levels in
(C) Y79 and (D) WERI-RB1 cells were determined via a western blot
assay. *P<0.05, **P<0.01 and
***P<0.001. miR, microRNA; NEAT1, nuclear paraspeckle
assembly transcript 1; RB, retinoblastoma; MMP, matrix
metalloproteinase; sh, short hairpin RNA; NC, negative control;
EMT, epithelial-to-mesenchymal transition; LRG1, leucine-rich
α-2-glycoprotein. |
The expression levels of MMP9, N-cadherin and
E-cadherin were then determined via western blotting. It was
indicated that MMP9 (both P<0.001)and N-cadherin expression
levels were significantly elevated (both P<0.001), while
E-cadherin expression was significantly reduced (Y79 cells,
P<0.01; WERI-RB1 cells, P<0.05) in pcDNA-LRG1-treated Y79 and
WERI-RB1 cells; these effects on MMP9 (both P<0.001), N-cadherin
(both P<0.001) and E-cadherin (Y79 cells, P<0.01; WERI-RB1
cells, P<0.05) levels were restored in pcDNA-LRG1 +
miR-24-3p-treated cells. Furthermore, NEAT1 overexpression
decreased the impact of pcDNA-LRG1 + miR-24-3p on the expression
levels of MMP9 (both P<0.001), N-cadherin (both P<0.001) and
E-cadherin (Y79 cells, P<0.001; WERI-RB1 cells, P<0.01;
Fig. 6C and D). Collectively, the results indicated
that NEAT1 regulates cell migration, invasion and EMT via the
miR-24-3p/LRG1 axis in RB.
Discussion
RB is an aggressive eye cancer associated with poor
prognosis in infants and children (2). Previous studies have reported that
lncRNAs serve important roles in RB progression (10,29,30).
However, understanding of the role lncRNA NEAT1 in RB is limited.
The present results suggest that NEAT1 is significantly upregulated
in RB tissues and cells, compared with corresponding healthy
tissues and cells. Furthermore, it was demonstrated that NEAT1
promotes RB cell migration, invasion and EMT by upregulating LRG1
expression via the sponging of miR-24-3p.
It has been revealed that the aberrant expression of
NEAT1 regulates the development of different cancer types. For
instance, Liu et al (31)
reported that NEAT1 was significantly increased in renal cell
carcinoma (RCC), and that NEAT1 knockdown suppressed RCC cell
proliferation, migration and invasion. Moreover, Ji et al
(32) reported that there is a high
expression of NEAT1 in osteosarcoma and that NEAT1 overexpression
accelerates osteosarcoma cell proliferation and inhibits cell
apoptosis. Wang et al (33)
revealed that there is a high expression of NEAT1 in RB tissues
compared with healthy retinal tissues, and NEAT1 silencing
represses RB cell proliferation, while inducing cell cycle arrest
and apoptosis. In line with the findings from Wang et al
(33), the present results indicate
that NEAT1 expression is elevated in RB tissues and cell lines.
Furthermore, two cell lines (Y79 and WERI-RB1) were selected to
investigate the roles of NEAT1 in RB. It was demonstrated that
NEAT1 knockdown significantly suppresses the migratory and invasive
abilities of these cells. Thus, it was speculated that NEAT1 may
function as an oncogene in RB.
Previous studies have demonstrated that lncRNAs
exert their biological functions via serving as miRNA sponges
(34). To the best of our
knowledge, the present study was the first to identified that NEAT1
binds miR-24-3p in Y79 and WERI-RB1 cells. miR-24 is downregulated
and function as a tumor suppressor in various cancer types, such as
osteosarcoma (35), gastric
(36) and bladder cancer (37). Moreover, miR-24 expression is
decreased in RB and its overexpression represses RB cell
proliferation, migration and invasion abilities (17,18).
In line with these previous findings, the present results suggested
that miR-24-3p expression was significantly decreased and
negatively modulated by NEAT1 in RB. Furthermore, overexpression of
miR-24-3p suppressed RB cell migration and invasion, and inhibition
of miR-24-3p restored the inhibitory effects of NEAT1 knockdown on
cell migration and invasion in Y79 and WERI-RB1 cells. Therefore,
it was indicated that miR-24-3p serves as a tumor inhibitor in RB,
and that NEAT1 regulates cell migration and invasion via sponging
miR-24-3p in RB.
It has been revealed that LRG1 is aberrantly
upregulated in several types of cancer, including RB (25). In addition, the present results were
consistent with these previous findings, and demonstrated that LRG1
expression was significantly elevated in RB tissues and cells. It
was also demonstrated that LRG1 is a target of miR-24-3p, and the
mRNA expression level of LRG1 was negatively correlated with
miR-24-3p expression. Furthermore, the roles of LRG1 in RB were
examined and it was identified that LRG1 overexpression promoted
the migration and invasion of Y79 and WERI-RB1 cells, while these
effects were reversed by miR-24-3p overexpression.
The EMT process results in increased mobility and
invasiveness of cancer cells, which is considered as a key early
step in cancer progression and metastasis (38,39).
Moreover, Li et al (40)
reported that NEAT1 induces EMT in breast cancer. Lu et al
(41) also reported that
overexpression of NEAT1 is associated with the EMT process via the
miR-204/zinc finger E-box binding homeobox 1 axis in nasopharyngeal
carcinoma. The present results suggest that knockdown of NEAT1 may
suppress MMP9 and N-cadherin expression, as well as promote
E-cadherin expression; however, these effects were reversed by
miR-24-3p knockdown. Furthermore, the effects of LRG1
overexpression on EMT markers was reversed by co-transfection with
an miR-24-3p mimic, while the effects were further strengthened by
the overexpression of NEAT1 in RB cells. Therefore, the present
results indicated that NEAT1 facilitates the EMT pathway by
upregulating LRG1 via sponging miR-24-3p in RB cells. However, the
present study did not conduct in vivo experiments on
animals, and thus these will be performed in future studies through
transfecting sh-NEAT1 into nude mice and then monitoring the volume
and weight of xenograft tumors.
In conclusion, the present results suggested that
NEAT1 expression is elevated in RB. It was also demonstrated that
NEAT1 knockdown represses RB progression via suppressing cell
migration, invasion and the EMT process. Moreover, the present
study established a novel regulatory network; the
NEAT1/miR-24-3p/LRG1 axis, which was revealed to regulate RB
progression via functional and mechanism analysis. Collectively,
the findings indicated that NEAT1 may represent a potential
therapeutic target for RB treatment.
Supplementary Material
Transfection efficiency of
pcDNA-NEAT1, miR-24-3p, anti-miR-24-3p and pcDNA-LRG1. (A)
Expression of NEAT1 in Y79 and WERI-RB1 cells transfected with
pcDNA or pcDNA-NEAT1 was determined via RT-qPCR.
***P<0.001 vs. pcDNA. (B) Expression of miR-24-3p in
Y79 and WERI-RB1 cells transfected with miR-NC or miR-24-3p was
determined by RT-qPCR. ***P<0.001 vs. miR-NC. (C)
Expression of miR-24-3p in Y79 and WERI-RB1 cells transfected with
anti-NC or anti miR-24-3p was determined by RT-qPCR and
***P<0.001 vs. anti-NC. (D) Protein expression of
LRG1 in Y79 and WERI-RB1 cells transfected with pcDNA or pcDNA-LRG1
was determined via western blotting. ***P<0.001 vs.
pcDNA. miR, microRNA; NEAT1, nuclear paraspeckle assembly
transcript 1; NC, negative control; RT-qPCR, reverse
transcription-quantitative PCR; LRG1, leucine-rich
α.2.glycoprotein.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LaL conceived and designed the study, and drafted
the first draft of the manuscript. All experiments were completed
by all authors. QH, YW, LuL and JL analyzed and collated the
results. All authors reviewed and critiqued the manuscript, and
agreed to the final submission of the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Written informed consent was obtained from every
patient and the study was approved by the Ethics Committee of
Renmin Hospital, Hubei University of Medicine (Shiyan, Hubei).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dimaras H, Kimani K, Dimba EA, Gronsdahl
P, White A, Chan HS and Gallie BL: Retinoblastoma. Lancet.
379:1436–1446. 2012.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Kivelä T: The epidemiological challenge of
the most frequent eye cancer: Retinoblastoma, an issue of birth and
death. Br J Ophthalmol. 93:1129–1131. 2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Cimino PJ, Robirds DH, Tripp SR, Pfeifer
JD, Abel HJ and Duncavage EJ: Retinoblastoma gene mutations
detected by whole exome sequencing of merkel cell carcinoma. Mod
Pathol. 27:1073–1087. 2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Khan AA, Bukhari MH and Mehboob R:
Association of retinoblastoma with clinical and histopathological
risk factors. Nat Sci. 5:437–444. 2013.
|
|
5
|
Abramson DH, Shields CL, Munier FL and
Chantada GL: Treatment of retinoblastoma in 2015: Agreement and
disagreement. JAMA Ophthalmol. 133:1341–1347. 2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Peng WX, Koirala P and Mo YY:
LncRNA-mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Fan Q, Yang L, Zhang X, Peng X, Wei S, Su
D, Zhai Z, Hua X and Li H: The emerging role of exosome-derived
non-coding RNAs in cancer biology. Cancer Lett. 414:107–115.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Gutschner T and Diederichs S: The
hallmarks of cancer: A long non-coding RNA point of view. RNA Biol.
9:703–719. 2012.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yang Y and Peng XW: The silencing of long
non-coding RNA ANRIL suppresses invasion, and promotes apoptosis of
retinoblastoma cells through the ATM-E2F1 signaling pathway. Biosci
Rep. 38(BSR20180558)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhang H, Zhong J, Bian Z, Fang X, Peng Y
and Hu Y: Long non-coding RNA CCAT1 promotes human retinoblastoma
SO-RB50 and Y79 cells through negative regulation of miR-218-5p.
Biomed Pharmacother. 87:683–691. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang A, Shang W, Nie Q, Li T and Li S:
Long non-coding RNA H19 suppresses retinoblastoma progression via
counteracting miR-17-92 cluster. J Cell Biochem. 119:3497–3509.
2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yu X, Li Z, Zheng H, Chan MT and Wu WK:
NEAT 1: A novel cancer-related long non-coding RNA. Cell Prolif.
50(e12329)2017.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li X, Wang S, Li Z, Long X, Guo Z, Zhang
G, Zu J, Chen Y and Wen L: The lncRNA NEAT1 facilitates cell growth
and invasion via the miR-211/HMGA2 axis in breast cancer. Int J
Biol Macromol. 105:346–353. 2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Guo H, Yang S, Zhao S, Li L, Yan M and Fan
M: LncRNA NEAT1 regulates cervical carcinoma proliferation and
invasion by targeting AKT/PI3K. Eur Rev Med Pharmacol Sci.
22:4090–4097. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Lü J, Qian J, Chen F, Tang X, Li C and
Cardoso WV: Differential expression of components of the microRNA
machinery during mouse organogenesis. Biochem Biophys Res Commun.
334:319–323. 2005.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Zhao JJ, Yang J, Lin J, Yao N, Zhu Y,
Zheng J, Xu J, Cheng JQ, Lin JY and Ma X: Identification of miRNAs
associated with tumorigenesis of retinoblastoma by miRNA microarray
analysis. Childs Nerv Syst. 25:13–20. 2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
To KH, Pajovic S, Gallie BL and Thériault
BL: Regulation of p14ARF expression by miR-24: A potential
mechanism compromising the p53 response during retinoblastoma
development. BMC Cancer. 12(69)2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yu F, Pang G and Zhao G: ANRIL acts as
onco-lncRNA by regulation of microRNA-24/c-Myc, MEK/ERK and
Wnt/β-catenin pathway in retinoblastoma. Int J Biol Macromol.
128:583–592. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Takahashi N, Takahashi Y and Putnam FW:
Periodicity of leucine and tandem repetition of a 24-amino acid
segment in the primary structure of leucine-rich alpha
2-glycoprotein of human serum. Proc Natl Acad Sci USA.
82:1906–1910. 1985.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Serada S, Fujimoto M, Terabe F, Iijima H,
Shinzaki S, Matsuzaki S, Ohkawara T, Nezu R, Nakajima S, Kobayashi
T, et al: Serum leucine-rich alpha-2 glycoprotein is a disease
activity biomarker in ulcerative colitis. Inflamm Bowel Dis.
18:2169–2179. 2012.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang X, Abraham S, McKenzie JAG, Jeffs N,
Swire M, Tripathi VB, Luhmann UFO, Lange CAK, Zhai Z, Arthur HM, et
al: LRG1 promotes angiogenesis by modulating endothelial TGF-β
signalling. Nature. 499:306–311. 2013.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Nakajima M, Miyajima M, Ogino I, Watanabe
M, Miyata H, Karagiozov KL, Arai H, Hagiwara Y, Segawa T, Kobayashi
K and Hashimoto Y: Leucine-rich α-2-glycoprotein is a marker for
idiopathic normal pressure hydrocephalus. Acta Neurochir (Wien).
153:1339–1346. 2011.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lindén M, Lind SB, Mayrhofer C, Segersten
U, Wester K, Lyutvinskiy Y, Zubarev R, Malmström PU and Pettersson
U: Proteomic analysis of urinary biomarker candidates for nonmuscle
invasive bladder cancer. Proteomics. 12:135–144. 2012.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Andersen JD, Boylan KL, Jemmerson R,
Geller MA, Misemer B, Harrington KM, Weivoda S, Witthuhn BA,
Argenta P, Vogel RI and Skubitz AP: Leucine-rich
alpha-2-glycoprotein-1 is upregulated in sera and tumors of ovarian
cancer patients. J Ovarian Res. 3(21)2010.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Amer R, Tiosano L and Pe'er J:
Leucine-rich α-2-glycoprotein-1 (LRG-1) expression in
retinoblastoma. Invest Ophthalmol Vis Sci. 59:685–692.
2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Yilmaz M and Christofori G: EMT, the
cytoskeleton, and cancer cell invasion. Cancer Metastasis Rev.
28:15–33. 2009.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wang JX, Yang Y and Li K: Long noncoding
RNA DANCR aggravates retinoblastoma through miR-34c and miR-613 by
targeting MMP-9. J Cell Physiol. 233:6986–6995. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Hu C, Liu S, Han M, Wang Y and Xu C:
Knockdown of lncRNA XIST inhibits retinoblastoma progression by
modulating the miR-124/STAT3 axis. Biomed Pharmacother.
107:547–554. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Liu F, Chen N, Gong Y, Xiao R, Wang W and
Pan Z: The long non-coding RNA NEAT1 enhances
epithelial-to-mesenchymal transition and chemoresistance via the
miR-34a/c-Met axis in renal cell carcinoma. Oncotarget.
8:62927–62938. 2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ji S, Wang S, Zhao X and Lv L: Long
noncoding RNA NEAT1 regulates the development of osteosarcoma
through sponging miR-34a-5p to mediate HOXA13 expression as a
competitive endogenous RNA. Mol Genet Genomic Med.
7(e673)2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang L, Yang D, Tian R and Zhang H: NEAT1
promotes retinoblastoma progression via modulating miR-124. J Cell
Biochem. 120:15585–15593. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wang P, Ning S, Zhang Y, Li R, Ye J, Zhao
Z, Zhi H, Wang T, Guo Z and Li X: Identification of
lncRNA-associated competing triplets reveals global patterns and
prognostic markers for cancer. Nucleic Acids Res. 43:3478–3489.
2015.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Song L, Yang J, Duan P, Xu J, Luo X, Luo
F, Zhang Z, Hou T, Liu B and Zhou Q: MicroRNA-24 inhibits
osteosarcoma cell proliferation both in vitro and in vivo by
targeting LPAATβ. Arch Biochem Biophys. 535:128–135.
2013.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Duan Y, Hu L, Liu B, Yu B, Li J, Yan M, Yu
Y, Li C, Su L, Zhu Z, et al: Tumor suppressor miR-24 restrains
gastric cancer progression by downregulating RegIV. Mol Cancer.
13(127)2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Zhang S, Zhang C, Liu W, Zheng W, Zhang Y,
Wang S, Huang D, Liu X and Bai Z: MicroRNA-24 upregulation inhibits
proliferation, metastasis and induces apoptosis in bladder cancer
cells by targeting CARMA3. Int J Oncol. 47:1351–1360.
2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Bates RC and Mercurio AM: The
epithelial-mesenchymal tansition (EMT) and colorectal cancer
progression. Cancer Biol Ther. 4:371–376. 2005.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Li L and Li W: Epithelial-mesenchymal
transition in human cancer: Comprehensive reprogramming of
metabolism, epigenetics, and differentiation. Pharmacol Ther.
150:33–46. 2015.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Li X, Wang S, Li Z, Long X, Guo Z, Zhang
G, Zu J, Chen Y and Wen L: NEAT1 induces epithelial-mesenchymal
transition and 5-FU resistance through the miR-129/ZEB2 axis in
breast cancer. FEBS Lett. 591(570)2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Lu Y, Li T, Wei G, Liu L, Chen Q, Xu L,
Zhang K, Zeng D and Liao R: The long non-coding RNA NEAT1 regulates
epithelial to mesenchymal transition and radioresistance in through
miR-204/ZEB1 axis in nasopharyngeal carcinoma. Tumour Biol.
37:11733–11741. 2016.PubMed/NCBI View Article : Google Scholar
|