Introduction
Application of a controlled mechanical force can
cause orthodontic tooth movement (OTM), which influences several
biological processes. The biological changes to the periodontal
ligament (PDL) during OTM can be divided into the three following
stages: Tissue degeneration, removal of necrotic tissue and new
periodontal attachment (1,2). OTM has been shown to be associated
with the inflammatory response and cytokine production (3). The release of TGF-β and IL-1β during
the inflammatory response can facilitate osteoclast
differentiation, bone remodeling and tooth movement (4,5).
PDL can protect the alveolar bone against tooth root
and is mainly composed of dense connective tissue (6). During orthodontic treatment, PDL is
the initial biological medium that bears the mechanical force
(7). Subsequently, the
microenvironment of PDL cells (PDLCs) changes and the signal
transduction system initiates the transformation of the mechanical
stimulation signals into biochemical signals, participating in the
reconstruction of periodontal tissues, such as PDL, alveolar bone
and cementum (8,9). During that process, cell death, bone
formation, tissue absorption and regeneration promote OTM
development (10). In addition, the
PDL provides nutrition and maintains a balanced metabolism for
periodontal cells and tissues, which in turn regulates periodontal
remodeling (11).
Apoptosis and autophagy are the two main types of
cell self-destruction (12).
Apoptosis is a type I programmed cell death and is characterized by
cell shrinkage, chromatin condensation, DNA degradation and
fragmentation, cell division into apoptotic bodies and, ultimately,
phagocytosis and degradation by phagocytes (13,14).
Autophagy is a type II programmed death and is a highly conserved
intracellular evolutionary process (15). The autophagosome, which is comprised
of a double-membraned structure, encapsulates longevity proteins
and damaged organelles, and fuses with lysosomes (16). These findings indicate that
apoptosis and autophagy may be closely associated with the process
of OTM. However, the dynamic changes of autophagy and apoptosis
with regards to the reconstruction of the PDL during OTM, to the
best of our knowledge, have not been previously investigated.
The present study aims to investigate the dynamic
changes in apoptosis and autophagy and the involvement of PDL and
osteoclasts following OTM. The present study also aims to elucidate
the mechanism of autophagy and apoptosis after OTM. Findings from
the present study may provide potential targets for improving
discomfort and reducing complications after orthodontic
treatment.
Materials and methods
Equipment and reagents
A NiTi spiral tension spring (0.12 mm) was purchased
from Emondi Materials Technology Co., Ltd. The orthodontic ligation
wire (0.2 mm) and dynamometer were purchased from Hangzhou Bioer
Co., Ltd. The antibodies used in the present study were purchased
from Abcam and were as follows: Beclin-1 (cat. no. ab232461;
1:1,500), LC3-II (cat. no. ab51520; 1:1,500), P62 (cat. no.
ab56416; 1:1,000), Bcl-2 (cat. no. ab185002; 1:1,000), Bax (cat.
no. ab32503; 1:1,500) and caspase-3 (cat. no. ab197202; 1:800).
Tartrate-resistant acid phosphatase (TRAP) staining regents were
obtained from Beijing Solarbio Technology Co., Ltd.
OTM animal model
In total, 40 male Sprague Dawley rats (6-8-week-old;
weight, 247±33 g) were purchased from the experimental animal
center of Southwest Medical University and the animal study was
approved by the Ethics Committee of Southwest Medical University
(license no: SCXK-2019-17; Luzhou, China). The rats were housed
under a 12-h light/dark cycle with free access to food and water at
the temperature of 23-25˚C and 40-50% humidity. The rats were
randomly divided into 4 groups, with 10 rats in each group. Rats
from the different groups were treated with orthodontic pressure
for 0 or 1 h, as well as 1 or 7 days. The animals were initially
anesthetized by intraperitoneal injection with xylazine (10 mg/kg)
and ketamine (100 mg/kg) as described previously (17). To check the successful induction of
anesthesia, the palpebral reflex was tested by touching the medial
canthus or the inner corner of the animals' eyes. No palpebral
reflex indicated the successful induction of anesthesia. A nitinol
tension spring was placed between the rat incisor and the right
upper first molar. The two incisors were used as anchorage and a
force of 0.392 N was applied to move the maxillary first molar
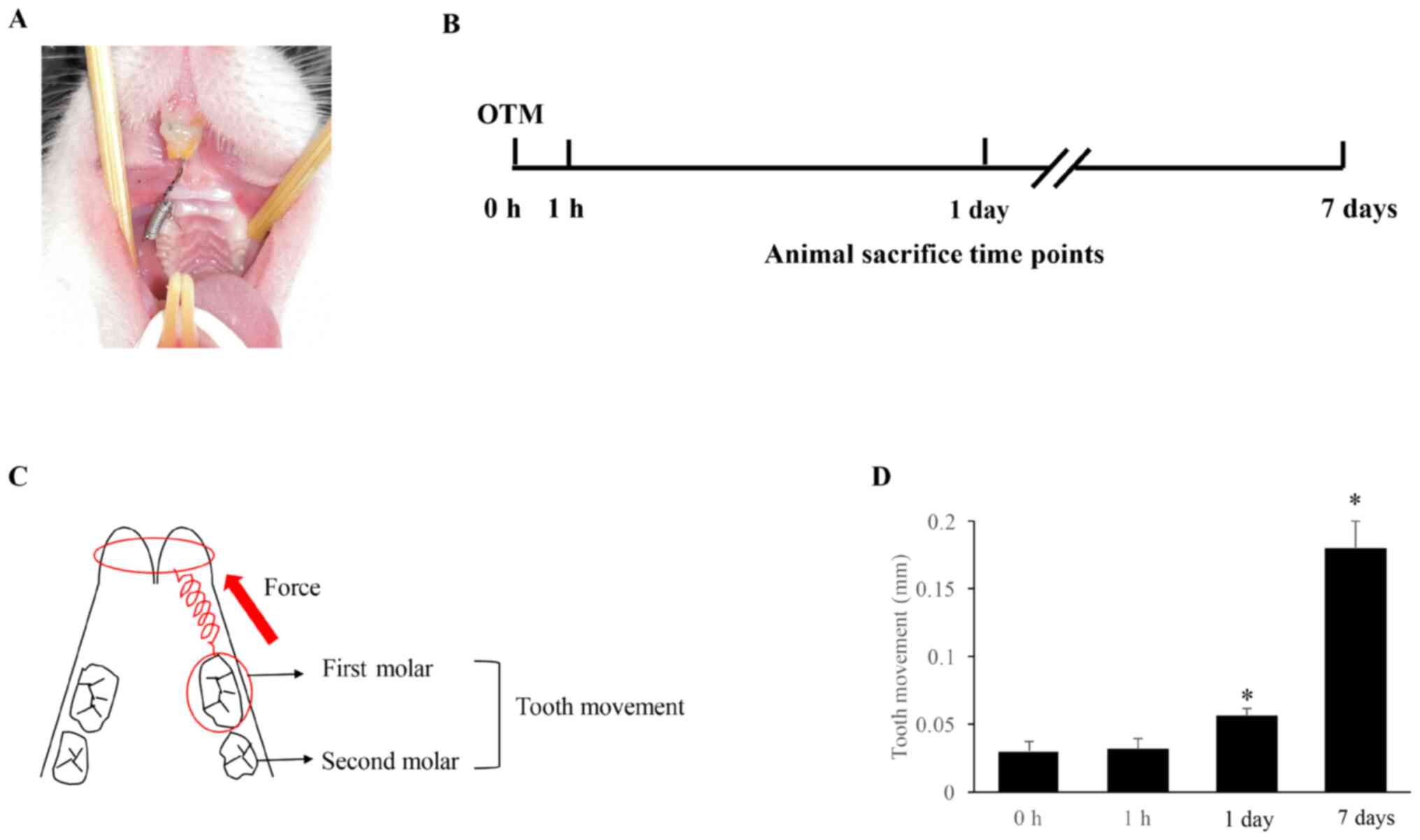
(Fig. 1A). The force application
device for each rat was monitored daily. The animals from the
various groups were euthanized at the end of the testing time
points (0, 1 h, 1 and 7 days). Inhalation of carbon dioxide (used
at a 30% flow displacement rate) was used for animal euthanasia,
which was performed in October 2019. Absence of respiration,
heartbeat and the corneal/palpebral reflex indicated the death of
animal. The right first molar and its surrounding tissues were
dissected and subsequently used for other experiments. The relevant
experiments involving the right first molar were subsequently
performed (Fig. 1B).
Immunohistochemical (IHC)
staining
The prepared tissues were fixed in 4%
paraformaldehyde solution for 48 h at room temperature. The tissues
were buffered in PBS solution for 24 h at room temperature and
transferred into 14% EDTA decalcification solution (pH: 7.3-7.5)
for decalcification at room temperature. The decalcification
solution was replaced daily. Following 6 weeks of decalcification,
the tissues were embedded using paraffin wax and cut into 4-µm
thick slices.
An initial deparaffinization step was conducted as
follows: Slides were incubated in xylene twice (5 min each time),
followed by washes in a descending ethanol gradient. The slides
were then washed using deionized water twice (5 min each time). In
total, 3% hydrogen peroxide (25 min) was used to remove endogenous
peroxidase activity at room temperature. Following washing with PBS
(three times, 5 min/time), goat serum (cat. no. ab7481; Abcam) was
applied for blocking (20 min) at room temperature. The
aforementioned primary antibodies were incubated with the tissues
overnight at 4˚C. Following washing with PBS (three times), the
tissues were incubated with the horseradish peroxidase-conjugated
secondary antibodies (cat. no. ab97051, 1:2,000; cat. no. ab205719,
1:2,000; Abcam) for 4 h at room temperature. Finally, DAB reagents
were used, and slides were counterstained with hematoxylin for 20
sec at room temperature. The relative protein expression was
analyzed using Image Pro-Plus 6.0 (Media Cybernetics, Inc.) with a
light microscope (magnifications, x200 and 400; BX51; Olympus
Corporation). Briefly, the background of images was adjusted
firstly. Five fields per slide were taken for further analysis.
Regions of interest were selected and the intensity of DAB staining
was calculated.
H&E staining
The tissues were prepared as aforementioned.
Following deparaffinization, the tissues were stained with
hematoxylin for 6 min at room temperature. After washing with
running tap water for 5 min, 1% acid alcohol (30 sec) was used for
differentiation. After washing with running tap water again for 1
min, 0.2% ammonia water (30 sec) was used for bluing. After washing
with running tap water for 5 min, slides were rinsed in 95% alcohol
for five times. Finally, the tissues were stained with eosin for 20
sec at room temperature, mounted and observed using light
microscopy (magnifications, x200 and 400; BX51; Olympus
Corporation). PDLCs were counted manually from five different
fields of view per slide.
TRAP staining
The TRAP solution was made according to the
instructions provided by the manufacturer. Following
deparaffinization as aforementioned, the tissues were cultured with
TRAP solution for 1 h at 37˚C and washed with deionized water three
times (5 min/each time). Finally, the tissues were counterstained
with hematoxylin for 3 min at room temperature, dehydrated and
mounted. Tissues were observed using microscopy (magnifications,
x200 and 400; BX51, Olympus Corporation). Osteoclasts were counted
manually from five different fields of view per slide.
Measurement of tooth movement
The mCT system (Rigaku-mCT) was used to measure the
tooth movement as previously described (18). After euthanizing at the end of
testing time points (0, 1 h, 1, and 7 days) before dissection, the
mCT system was used to detect the distance between the mesial
aspect of the second molar and the enamel on the most distal aspect
of the first molar.
Western blot analysis
The tissue samples surrounding first molar were
homogenized and then lysed using Protein Lysis Buffer (Promega
Corporation). Protein concentration was measured using
bicinchoninic acid protein assay method (Nanjing Jiancheng
Bioengineering Institute). The same amount of protein (30 µg) was
separated using 10% SDS-PAGE. The proteins were transferred to a
nitrocellulose membrane (EMD Millipore). Following blocking with 5%
non-fat milk prepared with TBS-Tween 20 (0.1%) at room temperature
for 2 h, the membranes were incubated with the primary antibodies
aforementioned overnight at 4˚C. After washing, the membranes were
incubated with the horseradish peroxidase-conjugated secondary
antibodies (Goat anti-rabbit IgG; cat. no. ab97051; 1:2,000; Goat
anti-mouse IgG, cat. no. ab205719, 1:2,000; Abcam) aforementioned
for 2 h at room temperature. SuperSignal™ West Femto Maximum
Sensitivity Substrate (Thermo Fisher Scientific, Inc.) was used as
visualization reagent. ImageJ software 1.53 version (National
Institutes of Health) was used to analyze the protein band
intensity.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated using the TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and the
extracted RNA was reverse-transcribed into cDNA using SuperScript™
II Reverse Transcriptase kit (Invitrogen; Thermo Fisher Scientific,
Inc.). The full temperature protocol for the reverse transcription
was listed as follows: Primer annealing (65˚C, 5 min), DNA
polymerization (45˚C, 50 min), enzyme deactivation (85˚C, 5 min).
RT-qPCR was conducted using SYBR Premix Ex Taq™ II kit (Takara Bio,
Inc.). The primer sequences used are as follows: GAPDH forward,
5'-ATGGGGAAGGTGAAGGTCG-3' and reverse,
5'-TCGGGGTCATTGATGGCAACAATA-3'; Bax forward,
5'-AGACAGGGGCCTTTTTGCTAC-3' and reverse, 5'-AATTCGCCGGAGACACTCG-3';
Bcl-2 forward, 5'-GTGAACTGGGGGAGGATTGT-3' and reverse,
5'-GGAGAAATCAAACAGAGGCCG-3'; caspase-3 forward,
5'-CTCGCTCTGGTACGGATGTG-3' and reverse,
5'-TCCCATAAATGACCCCTTCATCA-3'; Beclin-1 forward,
5'-ATGGAGGGGTCTAAGGCGTC-3' and reverse,
5'-TGGGCTGTGGTAAGTAATGGA-3'; LC3-II forward,
5'-GACCGCTGTAAGGAGGTGC-3' and reverse, 5'-AGAAGCCGAAGGTTTCTTGGG-3';
and P62 forward, 5'-GAGGCACCCCGAAACATGG-3' and reverse,
5'-ACTTATAGCGAGTTCCCACCA-3'. GAPDH gene was used as internal
control gene. The thermocycling conditions were set as follows:
Initial denaturation (98˚C, 2 min); 35 cycles for denaturation
(94˚C, 30 sec), annealing (55˚C, 40 sec) and extension (72˚C, 60
sec); Final extension (72˚C, 5 min). The data were analyzed using
the 2-∆∆Cq method (19).
Statistical analysis
The data were analyzed using SPSS 25.0 (IBM Corp.).
One-way ANOVAs followed by post-hoc Tukey's test were used to
analyze the significant differences between the various groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Establishment of the OTM animal
model
To investigate the dynamic changes of autophagy and
apoptosis during PDL reconstruction following OTM induction in
rats, initially an OTM model was established (Fig. 1A). The levels of specific key
autophagy and apoptotic proteins were measured at different time
points post OTM (Fig. 1B).
Moreover, the tooth movement was measured post-OTM induction and
the data indicated that after 1 or 7 days of OTM, there was
significant tooth movement compared with that in group 0 h
(Fig. 1C and D).
Assessment of PDLC and osteoclast
morphology using H&E and TRAP staining
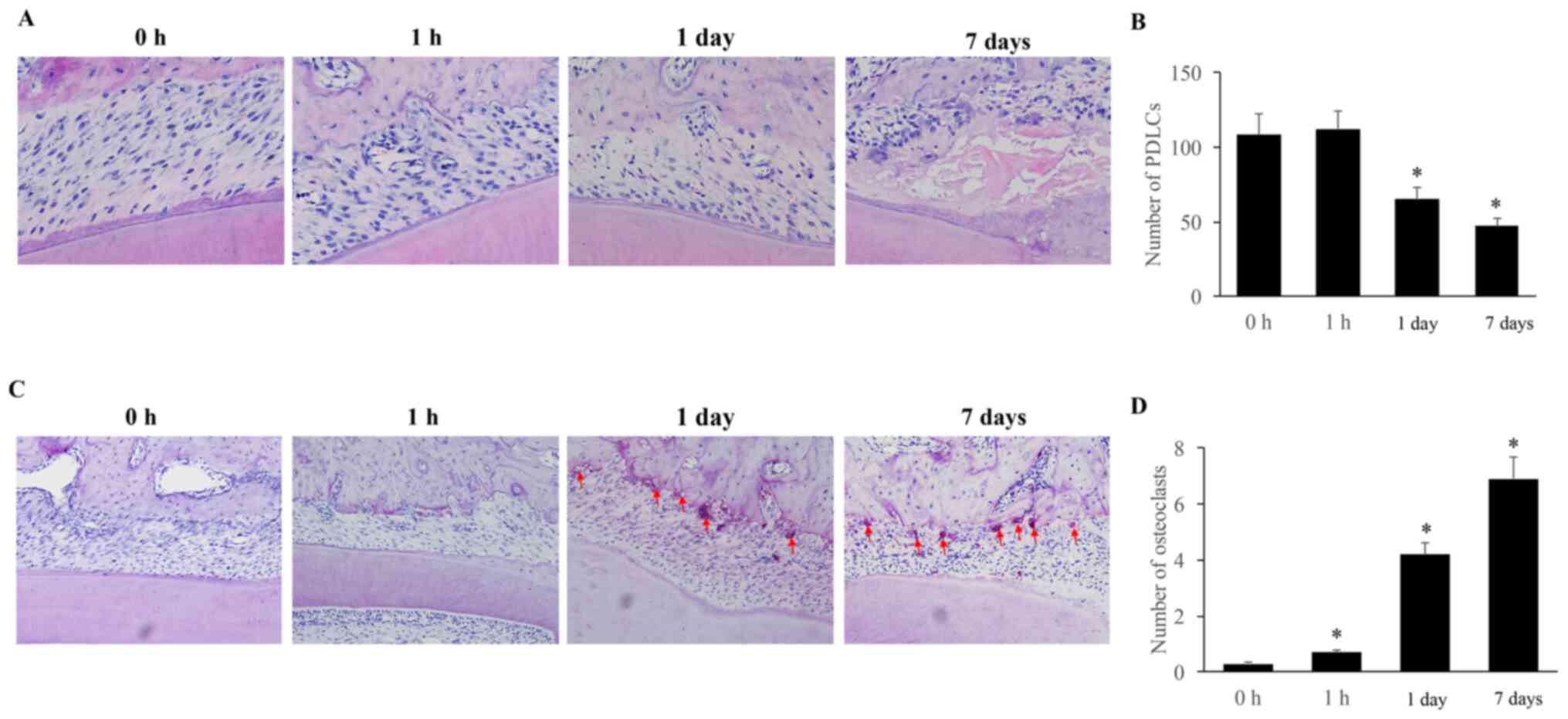
The PDLCs were observed using H&E staining.
PDLCs and PDL fibers in the control group were arranged in a
regular fashion. With the extended treatment times, the PDL gap in
the area where the pressure was exerted in the experimental groups
was gradually narrowed and the local PDL fibers exhibited a
wrinkled-like deformation. The PDLC arrangement was no longer
regular, the blood vessels were compressed and the diameter of the
tube was reduced (Fig. 2A). The
number of PDLCs was slightly increased following 1 h of orthodontic
force and significantly decreased following 1 and 7 days of
treatment (Fig. 2B).
The morphology and quantity of osteoclasts were
investigated using TRAP staining. The alveolar bone, in which
osteoclasts were predominantly located, demonstrated apparent
resorption lacuna (Fig. 2C). The
number of osteoclasts in the pressure area was increased
significantly following 1 h of orthodontic force application and
reached its maximum value on day 7 (Fig. 2D).
Determination of expression levels of
autophagy-related protein
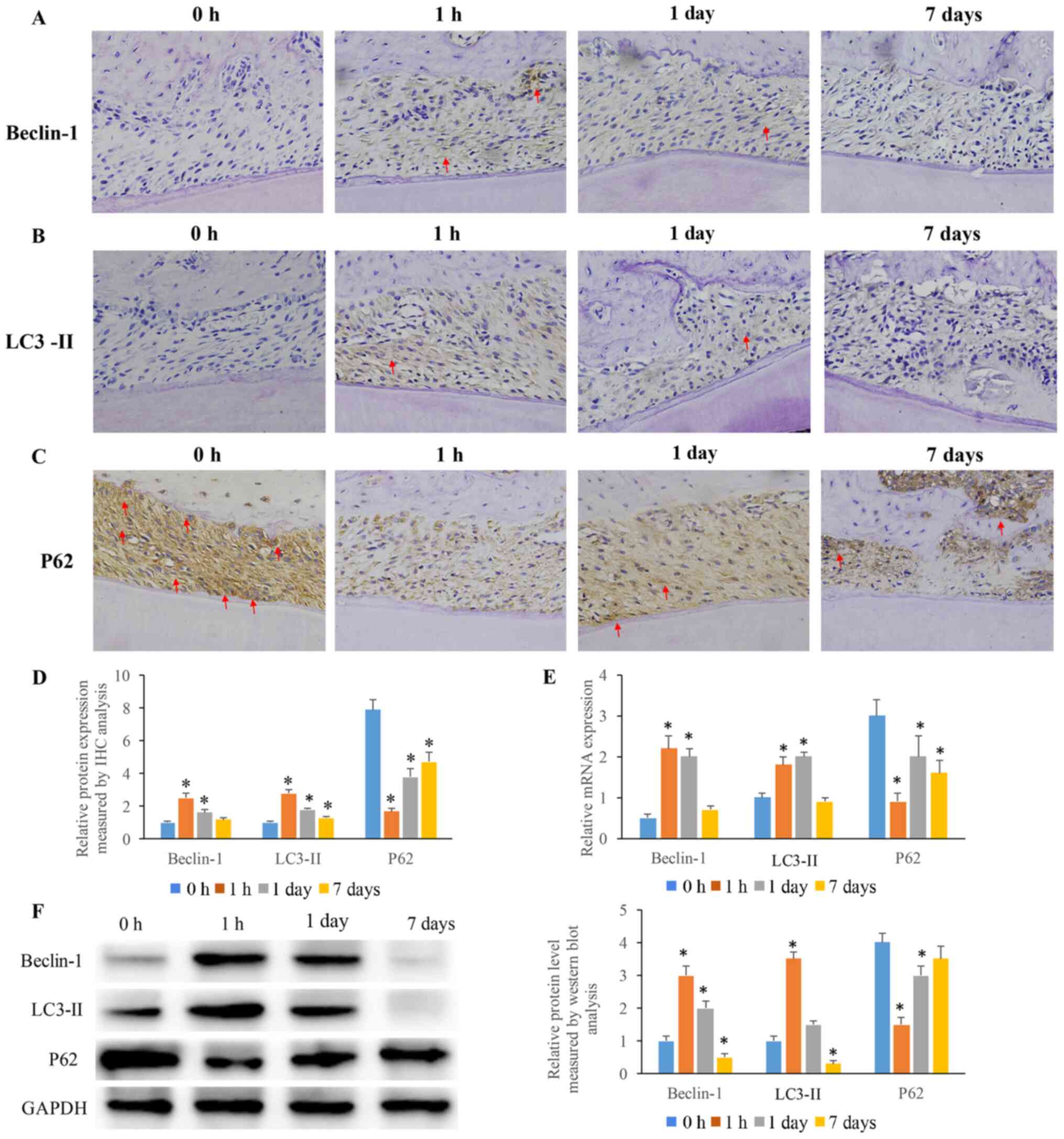
Beclin-1, LC3-II and P62 were mainly expressed in
the cytoplasm of PDLCs (Fig. 3A-C).
The expression levels of Beclin-1 and LC3-II in the experimental
groups reached their peak value following 1 h of treatment compared
with that noted in the 0 h group (Fig.
3D). Subsequently, the expression levels of Beclin-1 and LC3-II
decreased to the lowest levels on the 7th day (Fig. 3D). The changes in the expression
levels of p62 in PDLCs were opposite to those noted for Beclin-1
and LC3-II. The expression levels of p62 in each experimental group
were significantly lower than those noted in the 0 h group
(Fig. 3D). At 1 h post-OTM
treatment, the expression levels of P62 were at their lowest.
Similar findings were observed by measuring the mRNA and protein
expression levels of Beclin-1, LC3-II and p62. The expression
levels of Beclin-1 and LC3-II were significantly increased
following 1 h and 1 day of treatment (Fig. 3E and F). After 7 days, the levels of Beclin-1
and LC3-II reduced to baseline levels, possibly due to adaptation
to continuous stress. However, the expression levels of p62 were
inhibited following 1 h of treatment and were gradually increased
following 1 day of treatment (Fig.
3E and F). After 7 days, the
level of p62 returned to baseline levels, possibly due to
adaptation to continuous stress.
Determination of the expression levels
of apoptotic proteins
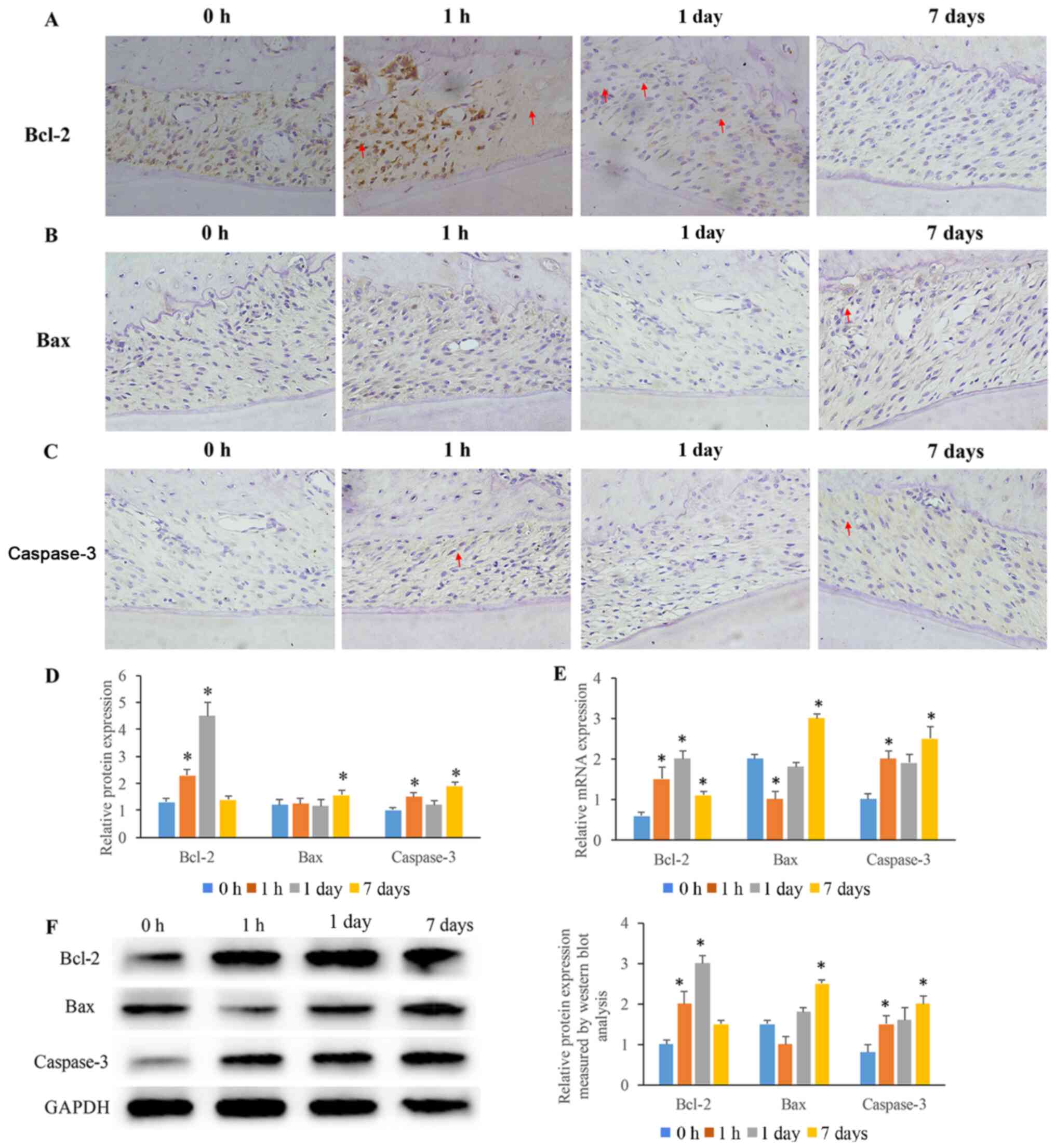
The expression levels of Bcl-2 significantly
increased after induction of OTM (1 h) and reached a peak at the 1
day time-point of OTM treatment (Fig.
4A and D). Subsequently, the
expression levels of Bcl-2 were decreased following 7 days of OTM
induction. However, a significant increase in the expression levels
of Bax was observed only on the 7th day post-OTM (Fig. 4B and D). In addition, the expression levels of
caspase-3 fluctuated during the time course. The expression of
caspase-3 was significantly increased at the 1-h and 7-day
timepoints following OTM induction (Fig. 4C and D). A slight increase was noted following 1
day of OTM induction (Fig. 4C and
D). The protein and mRNA expression
levels of Bcl-2, Bax and caspase-3 were also investigated using
western blotting and RT-qPCR assays, respectively (Fig. 4E and F). Bcl-2 mRNA and protein levels, although
raised, showed a reduction at 7 days, which was no longer
significant for the protein levels compared with those at 0 h.
Caspase-3 expression also stopped showing a significant raise at
the 1 day compared with that at 0 h, reducing from the 1 h
time-point. However, this level was increased again at day 7. The
mRNA levels of Bax were significantly decreased and increased
following 1 h and 7 days of OTM induction, respectively (Fig. 4E and F). Meanwhile, the protein level of Bax was
significantly increased after 7 days.
Discussion
Orthodontic mechanical force can be roughly divided
into traction force and compression force, and it can affect the
biological behavior of PDLCs (20,21).
Moreover, it has been reported that PDL hyalinization, bone density
and bone turnover can affect the rate of OTM (22). Continuous orthodontic pressure can
reduce the diameter of the blood vessels, lead to nutritional
disorders, reduce metabolism of PDLCs and result in PDLC starvation
(23-25).
This nutrient starved environment can further induce autophagy
(26).
Autophagy is a physiological response of cells to
stress and is maintained at a low baseline level under normal
conditions. Several factors, such as mechanical stimulation and
nutritional starvation can affect the induction of autophagy
(23-25).
It has been previously demonstrated that hypoxia and ischemia can
result in an insufficient supply of energy to cells (27). Subsequently, autophagy is activated
and the cell components are decomposed to provide energy, which is
conducive to cell metabolism and survival (25). The induction of autophagy is
involved in the regulation of the differentiation of osteoblasts
(28) and osteoclasts (29), whereby the production and
aggregation of osteoclasts is also associated with autophagy
(30). Appropriate mechanical
tension can promote the osteogenic differentiation of PDL stem
cells. In addition, autophagy can be activated by a compressive
force on PDLCs (23). Autophagy
regulates OTM by negatively modulating osteoclastogenesis and
maintaining bone homeostasis (31).
Sequestosome 1, a marker for autophagy, serves a key role during
the stress adaptation of cells (32). Marked increases in sequestosome 1
levels in the PDL are observed after orthodontic force treatment
(32). In addition,
osteoclastogenesis can be promoted through the expression of
autophagy-mediated RANKL after OTM (33). The active involvement of autophagy
during OTM has been previously shown (31), but the dynamic changes to
autophagy-related protein expression after OTM, to the best of our
knowledge, have not been previously reported.
The induction of apoptosis is characterized by
chromatin condensation, nuclear fragmentation and decomposition of
membrane-bound fragments (34,35).
Caspases cause the degradation of cellular proteins and activate
specific endonucleases, which finally results in DNA fragmentation
(36,37). Hypoxic conditions comprise the main
mechanism by which OTM can induce apoptosis and autophagy (38). It has reported that OTM may cause
hypoxia of periodontal tissues and further trigger the release of
hypoxia-inducible factor-1α, promoting osteogenic differentiation
(1). A previous study indicated
that Oridonin relieves hypoxia-induced apoptosis and autophagy by
targeting the PI3K/AKT/mTOR signaling pathway and by promoting
miR-214 expression (39).
Therefore, induction of OTM may also influence the induction of
apoptosis and autophagy through the regulation of the PI3K/AKT/mTOR
signaling pathway and miR-214. In addition, hypoxia is believed to
be an initiator for orthodontic tissue remodeling after OTM
(40,41). Hypoxia may accelerate the remodeling
of the PDL and bone via inducing an aseptic inflammatory response
(42). Therefore, the association
between the findings of the present study and the aforementioned
targets should be explored further.
Beclin-1 is a key regulator of autophagy and it is
predominantly involved in the formation of the autophagosome
(43). Downregulated or missing
expression of Beclin-1 can result in abnormal autophagy function,
which in turn induces apoptosis (44). Bcl-2 and Bax belong to the same
family of proteins and the Bcl-2/Bax ratio is a key factor that
determines the induction of apoptosis (45). Bcl-2 is not only an apoptosis
suppressor gene, but also serves a regulatory role in the induction
of autophagy by binding to Beclin-1(46). Overexpression of Bcl-2 can inhibit
the expression of Beclin-1 and suppress the formation of the
autophagosome (47). In the present
study, it was found that the levels of Bax and caspase-3 were
gradually increased, but the expression of Bcl-2 was decreased to
almost baseline levels 7 days after OTM. These data indicated that
the induction of apoptosis by OTM was relatively slow compared to
the induction of autophagy.
In the present study, continuous mechanical pressure
appeared to induce autophagy during OTM. The expression levels of
Beclin-1 and LC3-II were significantly increased following 1 h of
OTM induction. However, following 7 days of OTM induction, the
expression levels of Beclin-1 and LC3-II were markedly decreased,
indicating the induction of PDLC apoptosis. Opposite expression
changes of P62 compared to Beclin-1 and LC3-II were observed. The
decrease of Beclin-1 and LC3-II, and the increase of P62 1 day
after OTM may be due to the adaptive change of the cytoskeleton of
PDLCs. However, the proteins levels of Beclin-1 and LC3-II became
significantly reduced compared with those at baseline, where p62
expression was no longer significantly reduced at 7 days.
Long-term orthodontic stress stimulation and gradual
aggravation of starvation can lead to the induction of PDLC
autophagy and the expansion of the apoptotic response. Finally,
hyaline degeneration of the PDL is observed during these processes
(48,49). A previous study suggested that
LC3-II was markedly increased following hypoxia induction, which
indicated that this process was significantly associated with the
increased expression levels of autophagy-related proteins (50). In the present study, OTM treatment
induced the expression of LC3-II following 1 h of OTM induction,
indicating that it could cause the rapid stimulation of
autophagy.
Caspase-3 is the key protease involved in apoptosis
and it inhibits autophagy and promotes apoptosis by inactivating
Beclin-1 (46,51). In addition, caspase-3 is the most
important end shear enzyme involved in the process of apoptosis. It
has been reported that activation of caspase-3 precedes DNA
fragmentation in apoptotic cells (52,53).
In the present study, the expression levels of caspase-3 were
significantly increased following 1 h of OTM induction and were
gradually increased following the 1st day of the experimental
model. The activation of caspase-3 is indicative of the
irreversible stage of apoptosis (54). In the present study, a significant
increase in Bcl-2 expression levels was noted at the early stage
post-OTM, which was accompanied by a gradual increase of both Bax
and caspase-3.
In conclusion, the present study demonstrated a
significant decrease in the number of PDLCs and a significant
increase in the number of osteoclasts on the 1st day following OTM
induction. Autophagy was rapidly initiated 1 h after OTM, whereas
the induction of apoptosis was gradually increased after 7 days.
The present study provided further evidence regarding the dynamic
changes of autophagy and apoptosis during OTM in rats. In addition,
there are some limitations to the present study. Firstly, only 4
time points were set in the animal experiments; therefore, some
detailed changes to autophagy- and apoptosis-related proteins may
have been overlooked. Secondly, further signaling pathways
influencing the autophagy and apoptosis process after OTM need to
be explored.
Acknowledgements
Not applicable.
Funding
The present study was supported by the scientific research fund
project of Sichuan Medical Association (grant no. S18002) and
applied basic research of Luzhou science and technology and Human
Resources Bureau (grant no. 2018-JYJ-37).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MW and XX conceived and designed the experiments;
MW, LZ, FL, LM and QZ performed the experiments, WM and XX wrote
the paper. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
The animal study was approved by the Ethics
Committee of Southwest Medical University (license no:
SCXK-2019-17; Luzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang X, Chen D, Zheng J, Deng L, Chen Z,
Ling J and Wu L: Effect of microRNA-21 on hypoxia-inducible
factor-1α in orthodontic tooth movement and human periodontal
ligament cells under hypoxia. Exp Ther Med. 17:2830–2836.
2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
de Jong T, Bakker AD, Everts V and Smit
TH: The intricate anatomy of the periodontal ligament and its
development: Lessons for periodontal regeneration. J Periodontal
Res. 52:965–974. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Macari S, Ajay Sharma L, Wyatt A, Knowles
P, Szawka RE, Garlet GP, Grattan DR, Dias GJ and Silva TA:
Osteoprotective effects of estrogen in the maxillary bone depend on
ERα. J Dent Res. 95:689–696. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Castroflorio T, Gamerro EF, Caviglia GP
and Deregibus A: Biochemical markers of bone metabolism during
early orthodontic tooth movement with aligners. Angle Orthod.
87:74–81. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Üretürk SE, Saraç M, Fıratlı S, Can ŞB,
Güven Y and Fıratlı E: The effect of low-level laser therapy on
tooth movement during canine distalization. Lasers Med Sci.
32:757–764. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Suttorp CM, Xie R, Lundvig DM,
Kuijpers-Jagtman AM, Uijttenboogaart JT, Van Rheden R, Maltha JC
and Wagener FA: Orthodontic forces induce the cytoprotective enzyme
heme oxygenase-1 in rats. Front Physiol. 7(283)2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Adnan S, Lone MM, Khan FR, Hussain SM and
Nagi SE: Which is the most recommended medium for the storage and
transport of avulsed teeth? A systematic review. Dent Traumatol.
34:59–70. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yan XZ, van den Beucken J, Yuan C, Jansen
JA and Yang F: Spheroid formation and stemness preservation of
human periodontal ligament cells on chitosan films. Oral Dis.
24:1083–1092. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Onizuka S and Iwata T: Application of
periodontal ligament-derived multipotent mesenchymal stromal cell
sheets for periodontal regeneration. Int J Mol Sci.
20(2796)2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Singh A, Gill G, Kaur H, Amhmed M and
Jakhu H: Role of osteopontin in bone remodeling and orthodontic
tooth movement: A review. Prog Orthod. 19(18)2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Xu HY, Nie EM, Deng G, Lai LZ, Sun FY,
Tian H, Fang FC, Zou YG, Wu BL and Ou-Yang J: Periostin is
essential for periodontal ligament remodeling during orthodontic
treatment. Mol Med Rep. 15:1800–1806. 2017.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Ptak GE, Toschi P, Fidanza A, Czernik M,
Zacchini F, Modlinski JA and Loi P: Autophagy and apoptosis:
Parent-of-origin genome-dependent mechanisms of cellular
self-destruction. Open Biol. 4(140027)2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gao L, Loveless J, Shay C and Teng Y:
Targeting ROS-mediated crosstalk between autophagy and apoptosis in
cancer. Adv Exp Med Biol. 1260:1–12. 2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Li M, Tan J, Miao Y, Lei P and Zhang Q:
The dual role of autophagy under hypoxia-involvement of interaction
between autophagy and apoptosis. Apoptosis. 20:769–777.
2015.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Noguchi M, Hirata N, Tanaka T, Suizu F,
Nakajima H and Chiorini JA: Autophagy as a modulator of cell death
machinery. Cell Death Dis. 11(517)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wnuk A and Kajta M: Steroid and xenobiotic
receptor signalling in apoptosis and autophagy of the nervous
system. Int J Mol Sci. 18(2394)2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Alexoff DL, Vaska P, Marsteller D,
Gerasimov T, Li J, Logan J, Fowler JS, Taintor NB, Thanos PK and
Volkow ND: Reproducibility of 11C-raclopride binding in the rat
brain measured with the microPET R4: Effects of scatter correction
and tracer specific activity. J Nucl Med. 44:815–822.
2003.PubMed/NCBI
|
|
18
|
Sahara N, Moriyama K, Iida M and Watanabe
S: Fate of worn-out functional teeth in the upper jaw dentition of
sicyopterus japonicus (Gobioidei: Sicydiinae) during tooth
replacement. Anat Rec (Hoboken). 301:111–124. 2018.PubMed/NCBI View
Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Qin X, Li Q, Chen W, Bai Y, Baban B and
Mao J: The circadian expression of osteogenic factors in
periodontal tissue loading mechanical force: New concepts of the
personalized orthodontic care. EPMA J. 10:13–20. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Li M, Yi J, Yang Y, Zheng W, Li Y and Zhao
Z: Investigation of optimal orthodontic force at the cellular level
through three-dimensionally cultured periodontal ligament cells.
Eur J Orthod. 38:366–372. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Feng L, Yang R, Liu D, Wang X, Song Y, Cao
H, He D, Gan Y, Kou X and Zhou Y: PDL progenitor-mediated PDL
recovery contributes to orthodontic relapse. J Dent Res.
95:1049–1056. 2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
King JS, Veltman DM and Insall RH: The
induction of autophagy by mechanical stress. Autophagy.
7:1490–1499. 2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Ma KG, Shao ZW, Yang SH, Wang J, Wang BC,
Xiong LM, Wu Q and Chen SF: Autophagy is activated in
compression-induced cell degeneration and is mediated by reactive
oxygen species in nucleus pulposus cells exposed to compression.
Osteoarthritis Cartilage. 21:2030–2038. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Baskaran R, Poornima P, Priya LB, Huang CY
and Padma VV: Neferine prevents autophagy induced by hypoxia
through activation of Akt/mTOR pathway and Nrf2 in muscle cells.
Biomed Pharmacother. 83:1407–1413. 2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Kuang Y, Hu B, Feng G, Xiang M, Deng Y,
Tan M, Li J and Song J: Metformin prevents against oxidative
stress-induced senescence in human periodontal ligament cells.
Biogerontology. 21:13–27. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Esumi H, Izuishi K, Kato K, Hashimoto K,
Kurashima Y, Kishimoto A, Ogura T and Ozawa T: Hypoxia and nitric
oxide treatment confer tolerance to glucose starvation in a
5'-AMP-activated protein kinase-dependent manner. J Biol Chem.
277:32791–32798. 2002.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu F, Fang F, Yuan H, Yang D, Chen Y,
Williams L, Goldstein SA, Krebsbach PH and Guan JL: Suppression of
autophagy by FIP200 deletion leads to osteopenia in mice through
the inhibition of osteoblast terminal differentiation. J Bone Miner
Res. 28:2414–2430. 2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Xi G, Rosen CJ and Clemmons DR: IGF-I and
IGFBP-2 stimulate AMPK activation and autophagy, which are required
for osteoblast differentiation. Endocrinology. 157:268–281.
2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Zhao Y, Chen G, Zhang W, Xu N, Zhu JY, Jia
J, Sun ZJ, Wang YN and Zhao YF: Autophagy regulates hypoxia-induced
osteoclastogenesis through the HIF-1α/BNIP3 signaling pathway. J
Cell Physiol. 227:639–648. 2012.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Chen L, Mo S and Hua Y: Compressive
force-induced autophagy in periodontal ligament cells downregulates
osteoclastogenesis during tooth movement. J Periodontol.
90:1170–1181. 2019.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Memmert S, Nogueira AVB, Damanaki A,
Nokhbehsaim M, Rath-Deschner B, Götz W, Gölz L, Cirelli JA, Till A,
Jäger A and Deschner J: Regulation of the autophagy-marker
Sequestosome 1 in periodontal cells and tissues by biomechanical
loading. J Orofac Orthop. 81:10–21. 2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Li W, Zhao J, Sun W, Wang H, Pan Y, Wang L
and Zhang WB: Osteocytes promote osteoclastogenesis via
autophagy-mediated RANKL secretion under mechanical compressive
force. Arch Biochem Biophys. 694(108594)2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kijima M and Mizuta R: Histone H1 quantity
determines the efficiencies of apoptotic DNA fragmentation and
chromatin condensation. Biomed Res. 40:51–56. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Zierler S, Klein B, Furtner T, Bresgen N,
Lütz-Meindl U and Kerschbaum HH: Ultraviolet irradiation-induced
apoptosis does not trigger nuclear fragmentation but translocation
of chromatin from nucleus into cytoplasm in the microglial
cell-line, BV-2. Brain Res. 1121:12–21. 2006.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Verma DK, Gupta S, Biswas J, Joshi N,
Sivarama Raju K, Wahajuddin M and Singh S: Metabolic enhancer
piracetam attenuates the translocation of mitochondrion-specific
proteins of caspase-independent pathway, poly [ADP-Ribose]
polymerase 1 up-regulation and oxidative DNA fragmentation.
Neurotox Res. 34:198–219. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Samejima K, Ogawa H, Ageichik AV, Peterson
KL, Kaufmann SH, Kanemaki MT and Earnshaw WC: Auxin-induced rapid
degradation of inhibitor of caspase-activated DNase (ICAD) induces
apoptotic DNA fragmentation, caspase activation, and cell death: A
cell suicide module. J Biol Chem. 289:31617–31623. 2014.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Luo G, Jian Z, Zhu Y, Zhu Y, Chen B, Ma R,
Tang F and Xiao Y: Sirt1 promotes autophagy and inhibits apoptosis
to protect cardiomyocytes from hypoxic stress. Int J Mol Med.
43:2033–2043. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Gong L, Xu H, Zhang X, Zhang T, Shi J and
Chang H: Oridonin relieves hypoxia-evoked apoptosis and autophagy
via modulating microRNA-214 in H9c2 cells. Artif Cells Nanomed
Biotechnol. 47:2585–2592. 2019.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Romer P, Wolf M, Fanghanel J, Reicheneder
C and Proff P: Cellular response to orthodontically-induced
short-term hypoxia in dental pulp cells. Cell Tissue Res.
355:173–180. 2014.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wolf M, Lossdörfer S, Römer P, Kirschneck
C, Küpper K, Deschner J and Jäger A: Short-term heat pre-treatment
modulates the release of HMGB1 and pro-inflammatory cytokines in
hPDL cells following mechanical loading and affects monocyte
behavior. Clin Oral Investig. 20:923–931. 2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Li Y, Jacox LA, Little SH and Ko CC:
Orthodontic tooth movement: The biology and clinical implications.
Kaohsiung J Med Sci. 34:207–214. 2018.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wang N, Zhang Q, Luo L, Ning B and Fang Y:
β-asarone inhibited cell growth and promoted autophagy via
P53/Bcl-2/Bclin-1 and P53/AMPK/mTOR pathways in human glioma U251
cells. J Cell Physiol. 233:2434–2443. 2018.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Abedin MJ, Wang D, McDonnell MA, Lehmann U
and Kelekar A: Autophagy delays apoptotic death in breast cancer
cells following DNA damage. Cell Death Differ. 14:500–510.
2007.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Huang Y, Fu Z, Dong W, Zhang Z, Mu J and
Zhang J: Serum starvation-induces down-regulation of Bcl-2/Bax
confers apoptosis in tongue coating-related cells in vitro. Mol Med
Rep. 17:5057–5064. 2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Marino G, Niso-Santano M, Baehrecke EH and
Kroemer G: Self-consumption: The interplay of autophagy and
apoptosis. Nat Rev Mol Cell Biol. 15:81–94. 2014.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zhu J, Cai Y, Xu K, Ren X, Sun J, Lu S,
Chen J and Xu P: Beclin1 overexpression suppresses tumor cell
proliferation and survival via an autophagy-dependent pathway in
human synovial sarcoma cells. Oncol Rep. 40:1927–1936.
2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Xu J, Zhao X, Zeng J, Yu JH, Guan S, Xu XM
and Mei L: Role of autophagy in the periodontal ligament
reconstruction during orthodontic tooth movement in rats. J Dent
Sci. 15:351–363. 2020.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Memmert S, Damanaki A, Weykopf B,
Rath-Deschner B, Nokhbehsaim M, Götz W, Gölz L, Till A, Deschner J
and Jäger A: Autophagy in periodontal ligament fibroblasts under
biomechanical loading. Cell Tissue Res. 378:499–511.
2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Dong Y, Wu Y, Zhao GL, Ye ZY, Xing CG and
Yang XD: Inhibition of autophagy by 3-MA promotes hypoxia-induced
apoptosis in human colorectal cancer cells. Eur Rev Med Pharmacol
Sci. 23:1047–1054. 2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Cao C, Wang W, Lu L, Wang L, Chen X, Guo
R, Li S and Jiang J: Inactivation of Beclin-1-dependent autophagy
promotes ursolic acid-induced apoptosis in hypertrophic scar
fibroblasts. Exp Dermatol. 27:58–63. 2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Spiguel LR, Chandiwal A, Vosicky JE,
Weichselbaum RR and Skelly CL: Concomitant proliferation and
caspase-3 mediated apoptosis in response to low shear stress and
balloon injury. J Surg Res. 161:146–155. 2010.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Shimizu H, Ohgoh M, Ikeda M, Nishizawa Y
and Ogura H: Caspase-3-like protease activity-independent apoptosis
at the onset of neuronal cell death in the gerbil hippocampus after
global ischemia. Biol Pharm Bull. 30:1950–1953. 2007.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Thrane C, Kaufmann U, Stummann BM and
Olsson S: Activation of caspase-like activity and poly (ADP-ribose)
polymerase degradation during sporulation in Aspergillus nidulans.
Fungal Genet Biol. 41:361–368. 2004.PubMed/NCBI View Article : Google Scholar
|