Introduction
Glutamate-induced excitotoxicity is one of the major
pathological mechanisms associated with a number of chronic
neurodegenerative disorders, such as Alzheimer's disease and
Parkinson's disease in the brain (1-3)
and glaucoma, retinitis pigmentosa and age-associated macular
degeneration in the eyes (4-6).
Excessive accumulation of extracellular glutamate may lead to the
hyperexcitability of neurons, calcium overload and increased
intracellular oxygen free radicals, resulting in DNA injury and
apoptosis (7,8). Massive death of neurons inevitably
leads to the dysfunction of nervous system, irreversible vision
loss and is life threatening; therefore, minimizing neuronal
apoptosis is the most direct and effective therapeutic approach for
patients with the aforementioned diseases (9,10).
Small-molecular chemicals, including glutamate
antagonists, calcium channel blockers and antioxidant/radical
scavenging agents, are primarily used to provide pharmacological
neuroprotection (11-13);
however, the unexpected side effects associated with their low
target specificity have impeded their wider application (14). Supplementation of downregulated
neurotrophic factors is another approach to neuroprotection, such
as growth factors and erythropoietin (15-17).
Nevertheless, it requires repeated invasive administration, which
is not suitable for long-term therapy in chronic diseases, such as
glaucoma and diabetic retinopathy (18,19).
In contrast, small-molecule bioactive peptides are becoming a
hotspot for research, due to their relatively high membrane
permeability, low immunogenicity and convenient synthesis and
modification (20-22).
FK18 is a small peptide composed of 18 amino acids
that is derived from basic fibroblast growth factor (bFGF) using
bioinformatics screening methods (23). The sequence is located in the
conserved receptor-binding domain of human bFGF, which is known for
its neuroprotective activity against a series of cerebral and
ocular diseases, such as cerebral ischemia and light damage in eyes
(24-26).
SH-SY5Y cells were chosen in the current study as they share
similar morphological, neurochemical and electrophysiological
properties to neurons. The aim of the present study was to examine
the protective effect of FK18 on neurodegenerative diseases and
further investigated its mechanisms.
Materials and methods
Preparation of peptides
FK18 peptide (sequence, FFFERLESNNYNTYSRK) and a
scrambled negative control peptide, SCpep (sequence,
SFLNKTNFREFRNYSEYF), were prepared using a high-efficiency
solid-phase method by China Peptides Co., Ltd. All the peptides had
a purity >98% as tested by high-performance liquid
chromatography (Shimadzu Corporation) and were freeze-dried and
stored at -20˚C until further use.
Cell culture and treatment
SH-SY5Y cells were purchased from the Shanghai Cell
Bank at the Chinese Academy of Sciences (cat. no. SCSP-5014) and
were authenticated using STR profiling. The cells were maintained
in a DMEM/F-12 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Gibco; Thermo
Fisher Scientific, Inc.) at 37˚C in a humidified atmosphere
containing 5% CO2. Cells between the fourth and the
seventh passages were used in the present study, to ensure their
stable properties, including morphology, function and viability.
Cells were seeded in 96-well plates to test cell viability, in
24-well plates to conduct TUNEL assay, in 6-well plates to perform
flow cytometry and 10-cm dishes to perform western blotting. A cell
density of 1x106/ml was used for each type of plate in
following experiments. Excitotoxicity was induced by exposing the
cultures to different concentrations of glutamate (4, 6, 8 and 10
mM; Sigma-Aldrich; Merck KGaA) for 24 h at 37˚C.
To determine the effect of FK18, its dose-response
effect was tested, and the result demonstrated a plateau of
protection reached under the dose of 10 µg/ml (data not shown),
which was similar to that against oxygen and glucose
deprivation-induced injury as previously reported (23). Hence, 10 µg/ml was selected as the
dosage in subsequent experiments. The cells were pretreated with
normal saline, 10 µg/ml FK18, or recombinant human bFGF (100 ng/ml;
R&D Systems, Inc.) 2 h before, at the same time or 30 min after
the induction of excitotoxicity at 37˚C. The timepoint was chosen
according to preliminary experiment results (data not shown). For
the detection of Akt, phospho-Akt, Erk1/2, and phospho-Erk1/2, the
cells were treated with either LY294002 (20 µM; Beyotime Institute
of Biotechnology), a PI3K inhibitor, or U0126, a highly selective
inhibitor of MEK1/2 (20 µM; Beyotime Institute of Biotechnology ),
2 h prior to the addition of FK18.
Cell viability assay
Viability of SH-SY5Y cells was quantitatively
evaluated using an MTS kit (Promega Corporation). Briefly, 20 µl
MTS was added to each well of a 96-well plate for 2 h at 37˚C.
Absorbance was measured photometrically at 490 nm and cell
viability was expressed as a percentage of the optical density
measured in the control group treated with normal saline.
Annexin V staining and flow cytometry
for measuring apoptosis
The percentage of SH-SY5Y apoptotic cells following
glutamate injury was determined by flow cytometry using an Annexin
V-FITC/propidium iodide (PI) Detection kit (BD Pharmingen; BD
Biosciences), according to the manufacturer's instructions.
Briefly, the cells were digested with 0.05% trypsin and collected
by centrifugation (1,000 x g for 5 min) at room temperature. The
pellets were washed in PBS and resuspended in binding buffer. A 100
µl volume of the cell suspension (~1x105 cells) was
treated in the dark with 5 µl FITC Annexin V and 5 µl PI at room
temperature for 15 min. The stained cells were analyzed using a
FACSCalibur (Becton-Dickinson and Company) and FlowJo version 10.4
(Becton-Dickinson and Company). Both early and late apoptotic cells
were included in the final analysis.
TUNEL assay
TUNEL staining was performed according to the
manufacturer's instructions for 1 h at 37 ˚C (DeadEnd™ Fluorometric
TUNEL System; Promega Corporation) to detect apoptotic cells. For
the in vitro experiments, following glutamate treatment,
SH-SY5Y cells were fixed with 4% PFA for 25 min at 4˚C. After
fixation, the cells were permeabilized with 0.2% Triton X-100 for 5
min. The nuclei were imaged after counterstaining with 0.5 µg/ml
DAPI at room temperature for 10 min. TUNEL-positive cells were
viewed using a laser scanning confocal microscope (magnification,
x20; Zeiss LSM 510; Carl Zeiss AG) in six random fields (at least
100 DAPI-positive cells per field).
Western blotting
SH-SY5Y cells were lysed in RIPA lysis buffer
containing PMSF and phosphatase inhibitor (v/v=98:1:1) for 5, 10,
20, 30 and 60 min after 8 mM glutamate addition for detecting
phosphorylated related proteins. For detection of
apoptosis-associated markers, cell lysates were prepared 24 h after
8 mM glutamate addition. The cell lysates were then prepared and
the amount of protein in each lysate was determined using a
bicinchoninic acid (BCA) protein assay kit (Beyotime Institute of
Biotechnology). The mass of protein loaded per lane was 10 µl. The
extracted proteins were subjected to either 10 or 12% SDS-PAGE and
transferred onto PVDF membranes electrophoretically. After blocking
non-specific sites with 5% non-fat milk for 1 h at 37 ˚C, the
membrane was incubated with primary antibodies (all from Cell
Signaling Technology, Inc.) against phosphorylated-(p) Akt
(1:1,000; cat. no. 4060S), Akt (1:1,000; cat. no. 4691S), p-Erk
(1:1,000; cat. no. 4370S), Erk (1:1,000; cat. no. 4695S), Bcl-2
(1:1,000; cat. no. 4223S), Bax (1:1,000; cat. no. 5023S), cleaved
caspase-3 (1:1,000; cat. no. 9664S), full-length caspase-3
(1:1,000; cat. no. 14220S) or GAPDH (1:1,000; cat. no. 2118L)
overnight at 4˚C and detected with HRP-conjugated anti-rabbit IgG
secondary antibody (1:10;000; cat. no. 111-035-003; Jackson
ImmunoResearch Laboratories, Inc.) for 2 h at room temperature.
Bands were visualized using an enhanced chemiluminescence detection
system (Merck KGaA), and band densities were quantified using Adobe
Photoshop version 19.1.8 (Adobe Systems, Inc.).
Statistical analysis
All experiments were performed at least three times.
Data are expressed as the mean ± SD and analyzed using IBM SPSS
statistics version 24.0 (IBM Corp). The distribution of data was
examined using the Kolmogorov-Smirnov test. Differences were
evaluated by one-way ANOVA followed by a post hoc Tukey's HSD test.
Non-parametric data were analyzed using the Kruskal-Wallis test and
Bonferroni's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
FK18 improved cell viability in
SH-SY5Y cells after glutamate injury
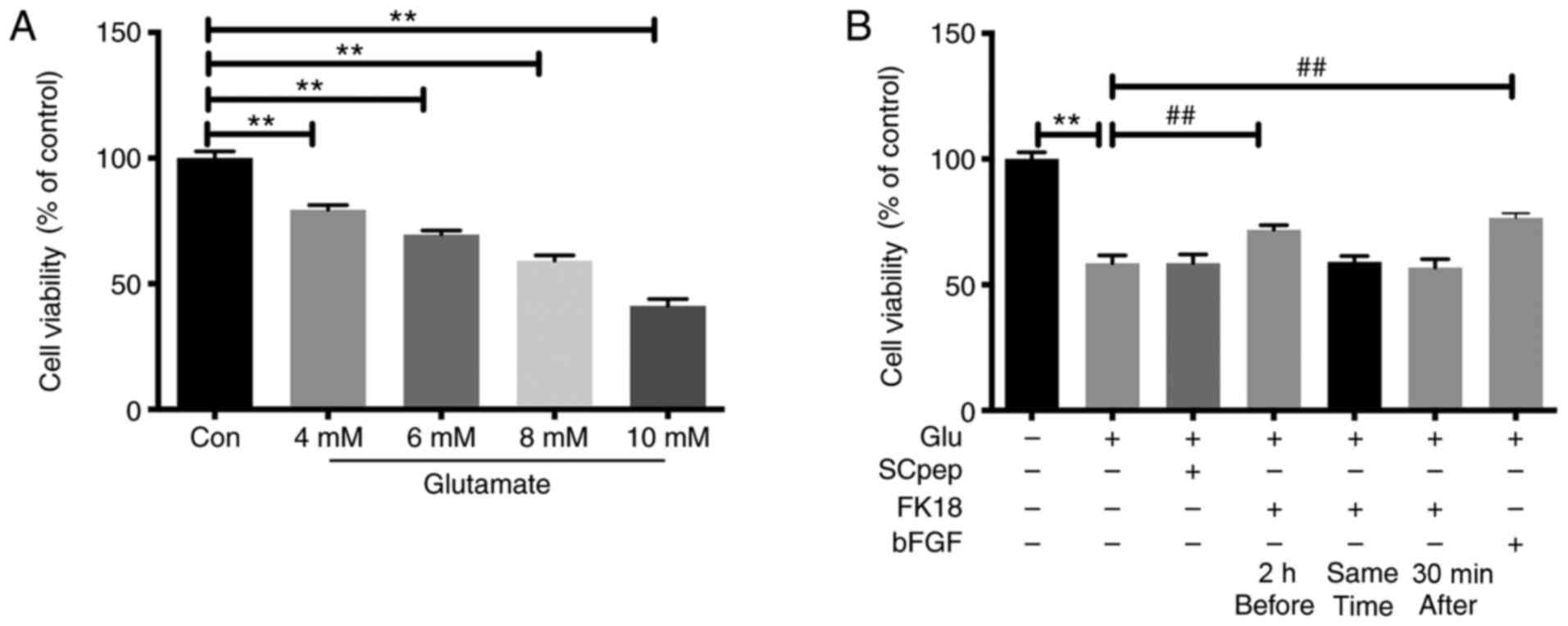
The cell viability of SH-SY5Y cells decreased
gradually as the concentration of glutamate increased, with the
survival rates being 79.48±1.84, 69.58±1.59, 59.16±2.21 and
41.18±2.77% after exposure to 4, 6, 8 and 10 mM of glutamate,
respectively (Fig. 1A). The insult
of 8 mM glutamate was chosen for the present study. Pretreatment
with FK18 dramatically increased cell viability, with an increase
in cell viability from 58.50±3.22 to 71.78 ± 1.87% (P<0.01;
Fig. 1B). However, when cells were
exposed to FK18, either simultaneously or 30 min after adding
glutamate, the neuroprotective effect was lost (Fig. 1B).
FK18 attenuated glutamate-induced cell
apoptosis in SH-SY5Y cells
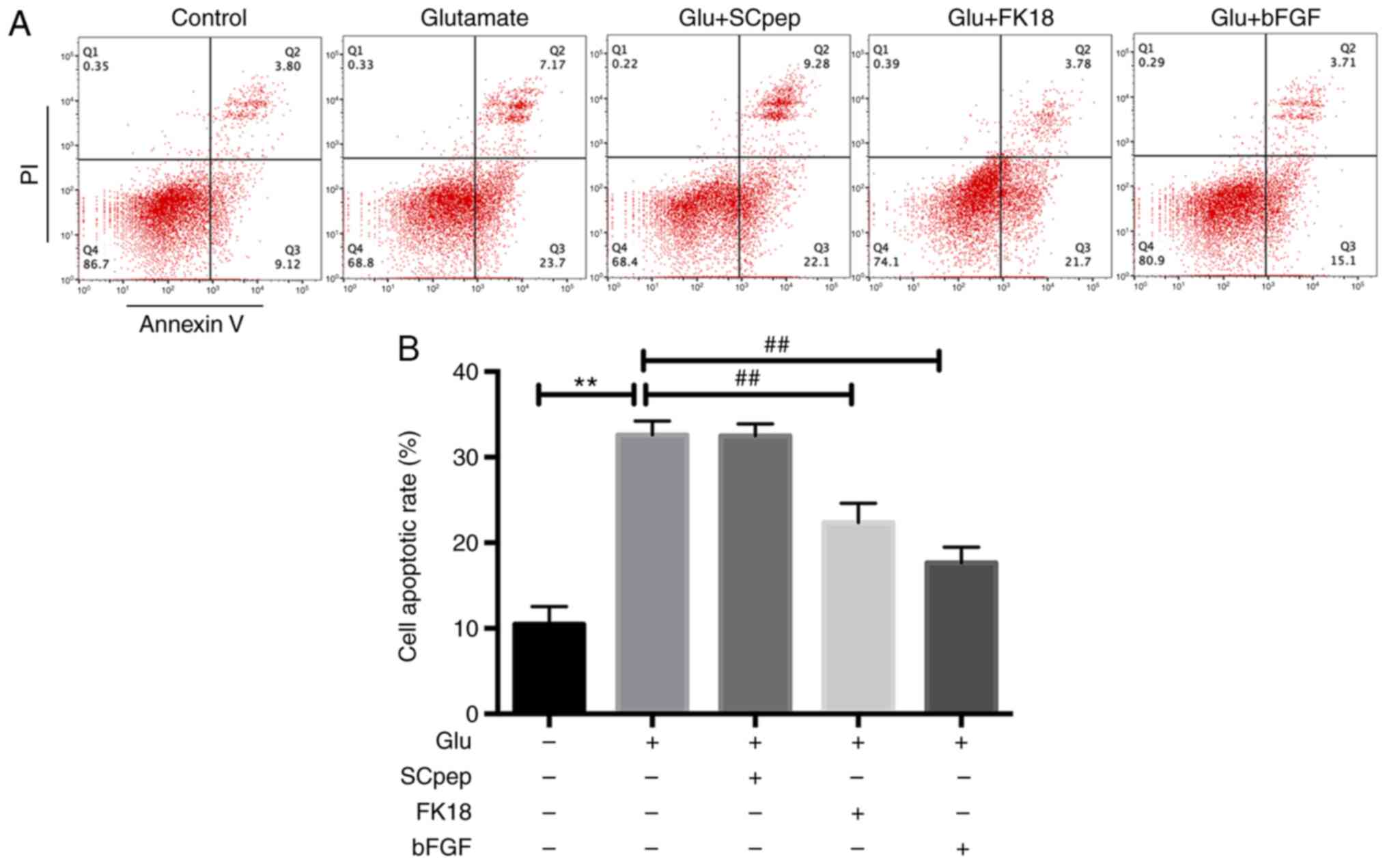
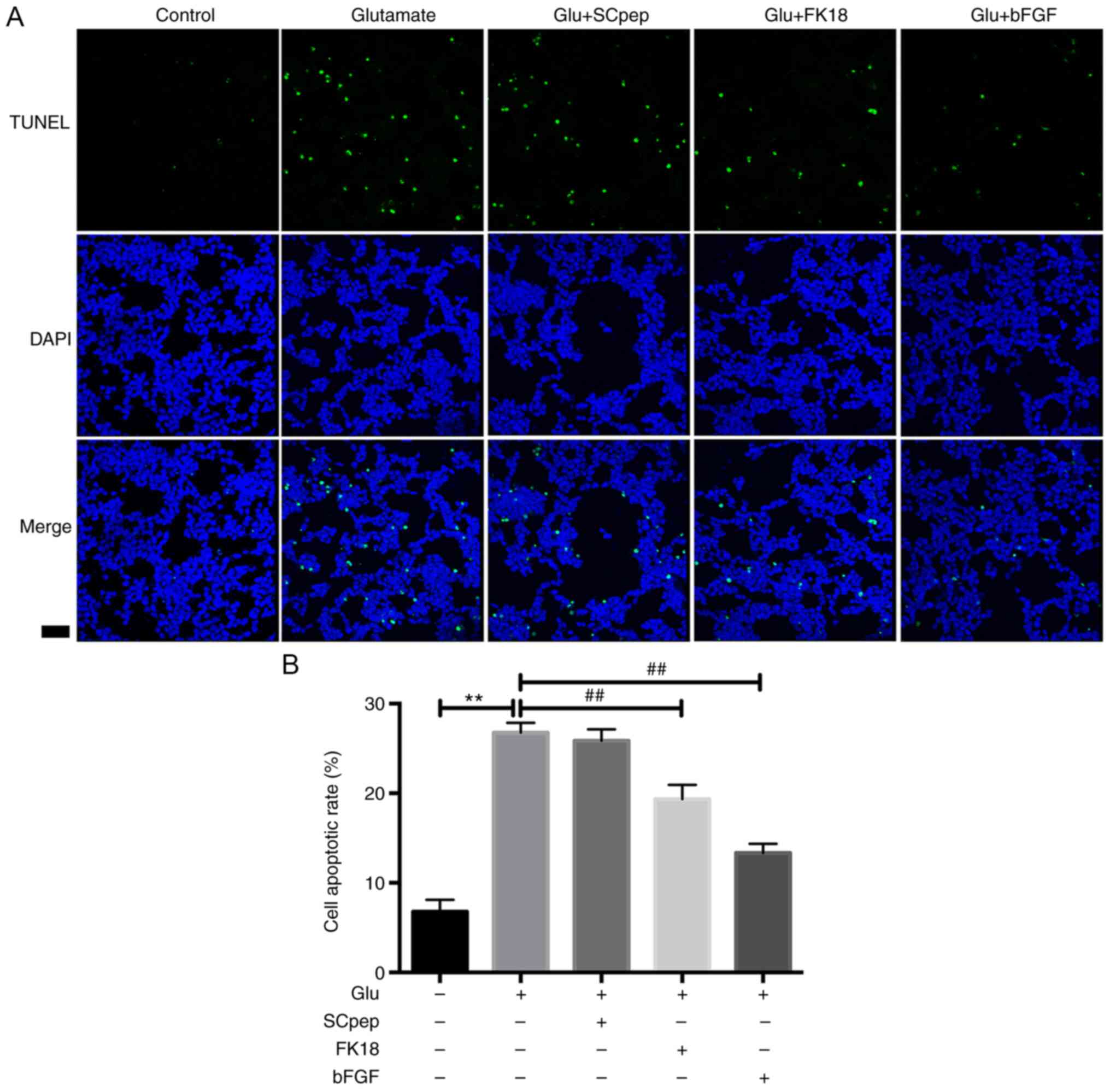
FK18 protective ability against glutamate-induced
apoptosis was assessed using flow cytometry and TUNEL assay. The
flow cytometry results demonstrated that pretreatment with FK18
reduced cell apoptosis from 32.58±1.63 to 22.35±2.26% in the
glutamate group (Fig. 2A and
B). The TUNEL assay demonstrated
that the percentage of TUNEL-positive cells decreased from
26.77±1.10 to 19.34±1.60% in the glutamate group following the
application of FK18 (Fig. 3A and
B). The bFGF group demonstrated a
decrease in apoptosis to 17.62±1.83 and 13.33±1.03% compared with
the glutamate group according to the flow cytometry and the TUNEL
assay, respectively. The SCpep group demonstrated no significant
difference compared with the glutamate group (Figs. 2 and 3).
Effect of FK18 on phosphorylation of
Akt and Erk in SH-SY5Y cells
To investigate the potential mechanisms underlying
the protective effects of FK18, the present study investigated
p-Akt and p-Erk expression with the treatment of glutamate or FK18
at different time points using western blotting. Treatment with 8
mM glutamate decreased the basal p-Akt level in a time-dependent
manner by 4.17, 9.38, 12.50, 13.50 and 21.88% compared with the
control group after 5, 10, 20, 30 and 60 min of treatment,
respectively, while it significantly increased the basal p-Erk
level in a time-dependent manner 1.12-, 1.23-, 1.37-, 1.43- and
1.58-fold, respectively (Fig.
4A-C). The decrease of Akt phosphorylation reached significance
60 min after the glutamate injury when compared with the control
group, while the increase of Erk phosphorylation reached
significance 20 min and peaked at 60 min, following glutamate
injury (Fig. 4A-C). FK18
pretreatment maintained a relatively high level of p-Akt and a low
level of p-Erk compared with the glutamate group (Fig. 4A-C). The addition of LY294002
significantly blocked the Akt phosphorylation caused by FK18 and
promoted the Erk phosphorylation that had been decreased by FK18
(Fig. 4D-F). The addition of U0126
had no effect on the p-Akt level, but significantly downregulated
p-Erk expression (Fig. 4D-F)
compared with the FK18 group. LY294002 abolished the
neuroprotective effect of FK18, with a decrease in cell viability
from 76.23±4.58 to 61.83±2.40%, while U0126 facilitated the effect
of FK18, increasing cell viability from 76.23±4.58 to 88.23±3.33%
(Fig. 4G).
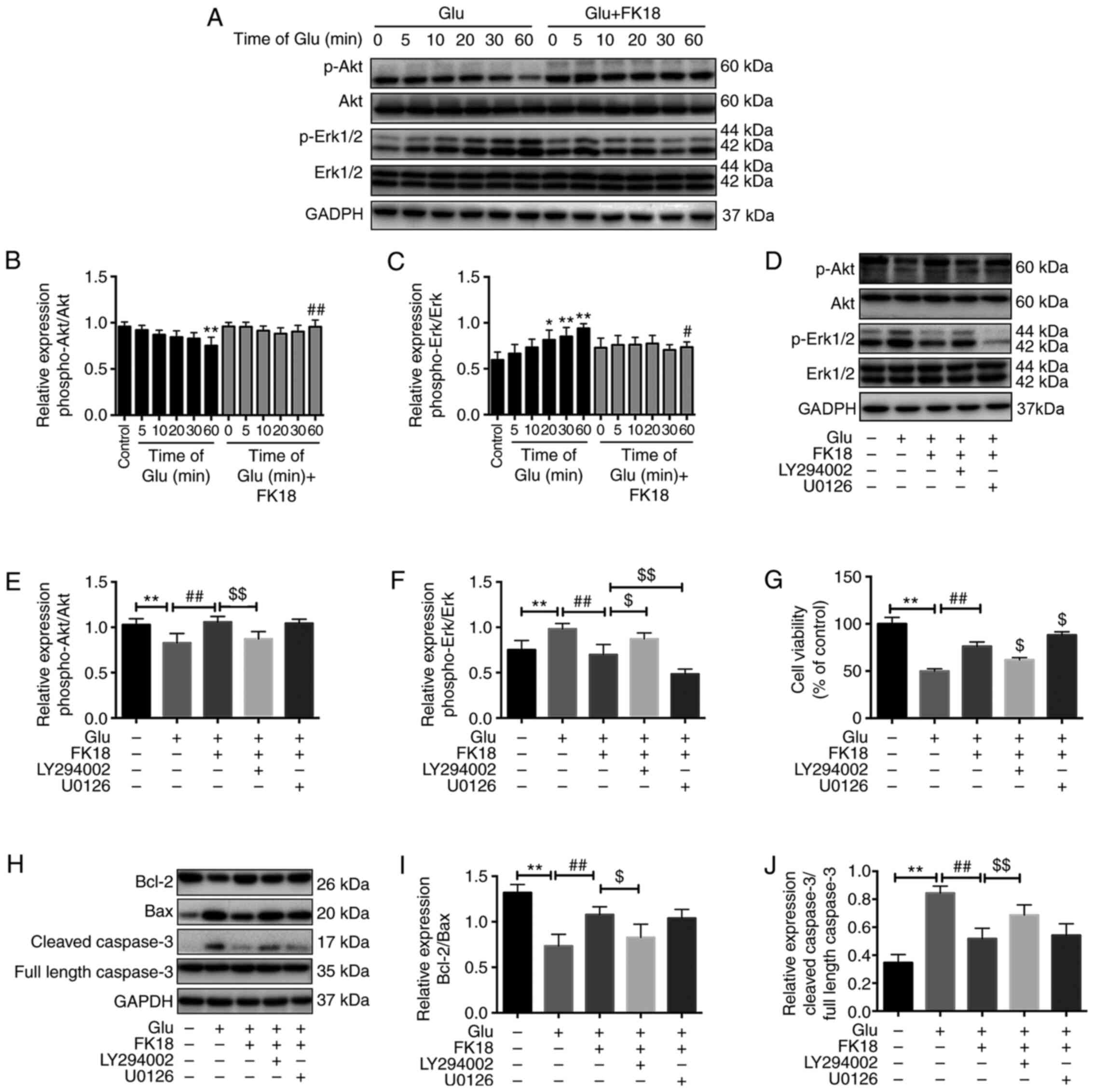 | Figure 4Effect of FK18 on Akt, Erk
phosphorylation and apoptosis-associated proteins in SH-SY5Y cells.
(A) Representative western blot image demonstrating the effect of
FK18 on p-Akt and p-Erk expression following the induction of
glutamate in a time-dependent manner. (B and C) Semi-quantification
of the relative expression of p-Akt/total Akt and p-Erk/total Erk
following glutamate injury in a time-dependent manner. (D)
Representative western blot image demonstrating the effect of FK18
on p-Akt and p-Erk expression in the presence or absence of
LY294002 or U0126. (E and F) Semi-quantification of the relative
expression of p-Akt/total Akt and p-Erk/total Erk in the presence
or absence of LY294002 and U0126. (G) Cell viability of SH-SY5Y
cells following glutamate injury in the presence or absence of
LY294002 or U0126, as determined by MTS assay. (H) Representative
western blots demonstrating the expression of Bcl-2, Bax and
caspase-3 following glutamate injury, with or without FK18
pretreatment, in the presence or absence of LY294002 or U0126. (I)
Bcl-2/Bax protein expression ratio following various treatments.
(J) Semi-quantification of cleaved caspase-3 protein expression
expressed as a ratio full length caspase-3 expression. The data is
expressed as the mean ± SD. *P<0.05 and
**P<0.01 vs. control group; #P<0.05 and
##P<0.01 vs. glutamate group; $P<0.05
and $$P<0.01 vs. glutamate-plus-FK18 group. Control
group, treated with normal saline; Glu, glutamate; bFGF, basic
fibroblast factor; PI, propidium iodide; p, phosphorylated;
LY294002, Akt pathway inhibitor; U0126, Erk pathway inhibitor. |
Effect of FK18 on apoptosis-associated
proteins in SH-SY5Y cells
Bcl-2/Bax ratio was a sensitive determinant of the
regulatory effects of glutamate, with or without FK18 and pathway
inhibitor pretreatment (27). The
ratio significantly decreased following glutamate injury, whereas
it significantly increased following the addition of FK18 and was
further inhibited by the effect of LY294002, but not affected by
U0126 compared with the FK18 group (Fig. 3H and I). The relative protein expression level
of cleaved caspase-3 significantly increased following glutamate
injury, whereas FK18 pretreatment suppressed this change. The
addition of LY294002 reversed this effect, but U0126 was not able
to do so (Fig. 3H and J).
Discussion
The present study demonstrated, for the first time
that FK18 protects against neuronal cell death induced by glutamate
in SH-SY5Y cells. The findings of the present study suggested that
the FK18 neuroprotective role is mediated via the Akt and Erk
pathways, which may be affected by Akt and Erk phosphorylation, but
independently of the caspase apoptotic pathway.
The human neuroblastoma cell line SH-SY5Y possesses
similar morphological, neurochemical and electrophysiological
properties to neurons and has been widely used for researching the
disease pathogenesis and mechanism underlying drugs in the nervous
system (28-30).
Glutamate-induced injury of SH-SY5Y cells provides a rapid and
sensitive in vitro model of neuronal excitotoxicity, with
concentrations of glutamate ranging from 2-100 mM (28,31-33).
In the present study, it was demonstrated that the injurious effect
of glutamate was concentration-dependent using MTS assay; 8 mM of
glutamate decreased cell viability by ~50% and was the most
suitable concentration for the subsequent observation of drug
effects.
The results of the present study demonstrated that
FK18 protected against neuronal excitatory injury in SH-SY5Y cells.
However, the neuroprotective effect of FK18 only occurred when the
peptide was applied before the excitotoxic stimulus and did not
occur when it was added simultaneously or after glutamate. This is
a common difficulty in other neuroprotective agents, such as
neuropeptide Y (34). As for
peptides, it may be improved by chemical modification and
optimization of its sequence and structure, or by the development
of a targeted drug delivery system (35). In addition, the neuroprotective
effects of FK18 against excitotoxicity in vitro were
relatively weak in the present study, which could be associated
with the non-specific mechanism of FK18 against excitotoxic injury.
The aforementioned anti-excitotoxic effect of FK18 may be a part of
its neuroprotection and other mechanisms that may be involved need
further investigation.
The Akt pathway is a central signal transduction
pathway involved in cell proliferation, survival and metabolism
(36,37). A previous study demonstrated that
Akt activity is responsible for as much as 80% of
neurotrophin-regulated cell survival, indicating that Akt is the
major survival-promoting protein for neurons (38). Akt signaling inhibits apoptosis by
regulating the expression of Bcl-2 and Bax and finally, the
expression of caspase-3 (39,40).
The findings of the present study were consistent with those of the
aforementioned studies. As demonstrated by western blotting in the
present study, FK18 activated the Akt pathway, significantly
promoted Akt phosphorylation and increased the cell viability of
SH-SY5Y cells. With the addition of the Akt inhibitor LY294002,
protection of SH-SY5Y cells by FK18 was significantly reversed. In
addition, the present study revealed that the expression of the
Bcl-2/Bax ratio decreased significantly after exposure to glutamate
and that this process could be reversed in the presence of FK18
(which was accompanied by lower cleaved-caspase-3 activity). When
adding the Akt inhibitor LY294002 to FK18, this effect was
eliminated in the present study.
The Erk signaling pathway is not only involved in
synaptic plasticity and neuronal development under physiological
conditions (41,42), but is associated with cell apoptosis
and neurodegeneration under pathological conditions (43,44).
In the present study, a role that the Erk pathway serves was
revealed, albeit a smaller one, in the protective effect of FK18.
When glutamate-injured SH-SY5Y cells were treated with FK18, Erk1/2
phosphorylation was significantly suppressed, this decrease was
further promoted by the addition of the Erk1/2 inhibitor U0126. In
the present study, cell viability increased by FK18 was further
improved by U0126; however, there were no significant changes of
the Bcl-2/Bax ratio and cleaved-caspase-3 with the addition of
U0126 compared with the FK18 group, suggesting that the
Erk-mediated neuroprotective effects of FK18 may be independent of
caspase-3. Previous studies have also demonstrated that persistent
active Erk translocates to the nucleus, causing cell apoptosis by
regulating gene expression and cell differentiation in a
caspase-3-independent manner (45,46).
In addition, cross-talk between the Akt and Erk pathways was
observed in the present study, with the Akt inhibitor LY294002
promoting the phosphorylation of Erk. Other studies have also
suggested a negative regulatory effect of Akt on the Erk pathway
(47,48), although the exact mechanisms need to
be further investigated.
Several limitations should be acknowledged. First,
the effects of FK18 in animal disease models have not been
evaluated in the current study, which needs further investigation.
Second, the specific target of FK18 has not been fully elucidated;
its effects on the Akt and Erk pathways as suggested in the current
study might provide some clues for future investigation Third, the
protection of FK18 against excitotoxic injury is relatively weak,
future modifications of the peptide would be helpful to improve its
effect.
In conclusion, the present study extended present
knowledge of the application of FK18 to neuronal excitatory
diseases by demonstrating the protective effects of FK18 against
excitotoxic injury in SH-SY5Y cells. In addition, the present study
elucidated the mechanism of FK18 by demonstrating that apart from
activating the Akt pathway, FK18 could exert its neuroprotective
effect through the suppression of the Erk pathway, which
cross-talks with the Akt pathway, but involves a
caspase-3-independent mechanism. The findings of the present study
indicated that FK18, a novel peptide derived from human bFGF may be
a promising therapeutic agent for the inhibition of neuronal death
in multiple neurological diseases involving excitotoxicity,
including Alzheimer's disease and Parkinson's disease in the brain
and glaucoma, retinitis pigmentosa and age-associated macular
degeneration in the eyes.
Acknowledgements
Not applicable.
Funding
This work was financially supported by the Shanghai Sailing
Program (grant no. 18YF1420200) and the Program of the National
Natural Science Foundation of China (grant no. 81970812).
Availability of data and methods
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SX and MM designed the study. SX drafted the
manuscript. SX, YX, FW and QG were responsible for the collection
and analysis of the experimental data. XH and XX made substantial
contributions to conception and design of the research, were
involved in revising the manuscript critically for important
intellectual content and approved the final version to be
published. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lipton SA and Rosenberg PA: Excitatory
amino acids as a final common pathway for neurologic disorders. N
Engl J Med. 330:613–622. 1994.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wang R and Reddy PH: Role of Glutamate and
NMDA Receptors in Alzheimer's Disease. J Alzheimers Dis.
57:1041–1048. 2017.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Pisanò CA, Brugnoli A, Novello S, Caccia
C, Keywood C, Melloni E, Vailati S, Padoani G and Morari M:
Safinamide inhibits in vivo glutamate release in a rat model of
Parkinson's disease. Neuropharmacology. 167(108006)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Connaughton V: Glutamate and Glutamate
Receptors in the Vertebrate Retina. In: Webvision: The Organization
of the Retina and Visual System. Kolb H, Fernandez E and Nelson R
(eds) University of Utah Health Sciences Center Copyright,
Webvision, Salt Lake City, UT, 1995.
|
|
5
|
Charles-Messance H, Blot G, Couturier A,
Vignaud L, Touhami S, Beguier F, Siqueiros L, Forster V, Barmo N,
Augustin S, et al: IL-1β induces rod degeneration through the
disruption of retinal glutamate homeostasis. J Neuroinflammation.
17(1)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Delyfer MN, Forster V, Neveux N, Picaud S,
Léveillard T and Sahel JA: Evidence for glutamate-mediated
excitotoxic mechanisms during photoreceptor degeneration in the rd1
mouse retina. Mol Vis. 11:688–696. 2005.PubMed/NCBI
|
|
7
|
Reynolds IJ and Hastings TG: Glutamate
induces the production of reactive oxygen species in cultured
forebrain neurons following NMDA receptor activation. J Neurosci.
15:3318–3327. 1995.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kruman II and Mattson MP: Pivotal role of
mitochondrial calcium uptake in neural cell apoptosis and necrosis.
J Neurochem. 72:529–540. 1999.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Ginsberg MD: Neuroprotection for ischemic
stroke: Past, present and future. Neuropharmacology. 55:363–389.
2008.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Osborne NN, Casson RF, Wood JP, Chidlow G,
Graham M and Melena J: Retinal ischemia: Mechanisms of damage and
potential therapeutic strategies. Prog Retin Eye Res. 23:91–147.
2004.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Binda NS, Carayon CP, Agostini RM,
Pinheiro AC, Cordeiro MN, Silva MA, Silva JF, Pereira EM, da Silva
CA Jr, Castro CJ Jr, et al: PhTx3-4, a spider toxin calcium channel
blocker, reduces NMDA-induced injury of the retina. Toxins (Basel).
8(70)2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yiğit U, Erdenöz S, Uslu U, Oba E, Cumbul
A, Cağatay H, Aktaş S and Eskicoğlu E: An immunohistochemical
analysis of the neuroprotective effects of memantine, hyperbaric
oxygen therapy, and brimonidine after acute ischemia reperfusion
injury. Mol Vis. 17:1024–1033. 2011.PubMed/NCBI
|
|
13
|
Stankowska DL, Dibas A, Li L, Zhang W,
Krishnamoorthy VR, Chavala SH, Nguyen TP, Yorio T, Ellis DZ and
Acharya S: Hybrid compound SA-2 is neuroprotective in animal models
of retinal ganglion cell death. Invest Ophthalmol Vis Sci.
60:3064–3073. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Craik DJ, Fairlie DF, Liras S, Liras S and
Price D: The future of peptide-based drugs. Chem Biol Drug Des.
81:136–147. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Jiang Y, Wei N, Lu T, Zhu J, Xu G and Liu
X: Intranasal brain-derived neurotrophic factor protects brain from
ischemic insult via modulating local inflammation in rats.
Neuroscience. 172:398–405. 2011.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Larpthaveesarp A, Georgevits M, Ferriero
DM and Gonzalez FF: Delayed erythropoietin therapy improves
histological and behavioral outcomes after transient neonatal
stroke. Neurobiol Dis. 93:57–63. 2016.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhao YZ, Lin M, Lin Q, Yang W, Yu XC, Tian
FR, Mao KL, Yang JJ, Lu CT and Wong HL: Intranasal delivery of bFGF
with nanoliposomes enhances in vivo neuroprotection and neural
injury recovery in a rodent stroke model. J Control Release.
224:165–175. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Harrell CR, Fellabaum C, Arsenijevic A,
Markovic BS, Djonov V and Volarevic V: Therapeutic potential of
mesenchymal stem cells and their secretome in the treatment of
glaucoma. Stem Cells Int. 2019(7869130)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Minhas G, Prabhakar S, Morishita R,
Shimamura M, Bansal R and Anand A: Transplantation of
lineage-negative stem cells in pterygopalatine artery ligation
induced retinal ischemia-reperfusion injury in mice. Mol Cell
Biochem. 429:123–136. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Cervia D, Catalani E and Casini G:
Neuroprotective peptides in retinal disease. J Clin Med.
8(1146)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Sun Q, Shen Y, Su L and Xu X: Inhibition
of pathological retinal neovascularization by a small peptide
derived from human tissue-type plasminogen kringle 2. Front
Pharmacol. 10(1639)2020.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Niu T, Cheng L, Wang H, Zhu S, Yang X, Liu
K, Jin H and Xu X: KS23, a novel peptide derived from adiponectin,
inhibits retinal inflammation and downregulates the proportions of
Th1 and Th17 cells during experimental autoimmune uveitis. J
Neuroinflammation. 16(278)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Xiong S, Xu Y, Ma M, Wang H, Wei F, Gu Q
and Xu X: Neuroprotective effects of a novel peptide, FK18, under
oxygen-glucose deprivation in SH-SY5Y cells and retinal ischemia in
rats via the Akt pathway. Neurochem Int. 108:78–90. 2017.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Fernández-Sánchez MT and Novelli A: Basic
fibroblast growth factor protects cerebellar neurons in primary
culture from NMDA and non-NMDA receptor mediated neurotoxicity.
FEBS Lett. 335:124–131. 1993.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ma J, Qiu J, Hirt L, Dalkara T and
Moskowitz MA: Synergistic protective effect of caspase inhibitors
and bFGF against brain injury induced by transient focal ischaemia.
Br J Pharmacol. 133:345–350. 2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
O'Driscoll C, O'Connor J, O'Brien CJ and
Cotter TG: Basic fibroblast growth factor-induced protection from
light damage in the mouse retina in vivo. J Neurochem. 105:524–536.
2008.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619.
1993.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Taveira M, Sousa C, Valentão P, Ferreres
F, Teixeira JP and Andrade PB: Neuroprotective effect of steroidal
alkaloids on glutamate-induced toxicity by preserving mitochondrial
membrane potential and reducing oxidative stress. J Steroid Biochem
Mol Biol. 140:106–115. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Ito S, Ménard M, Atkinson T, Brown L,
Whitfield J and Chakravarthy B: Relative expression of the p75
neurotrophin receptor, tyrosine receptor kinase A, and insulin
receptor in SH-SY5Y neuroblastoma cells and hippocampi from
Alzheimer's disease patients. Neurochem Int. 101:22–29.
2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu W, Ma H, DaSilva NA, Rose KN, Johnson
SL, Zhang L, Wan C, Dain JA and Seeram NP: Development of a
neuroprotective potential algorithm for medicinal plants. Neurochem
Int. 100:164–177. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gao M, Zhang WC, Liu QS, Hu JJ, Liu GT and
Du GH: Pinocembrin prevents glutamate-induced apoptosis in SH-SY5Y
neuronal cells via decrease of bax/bcl-2 ratio. Eur J Pharmacol.
591:73–79. 2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Hu Y, Li J, Liu P, Chen X, Guo DH, Li QS
and Rahman K: Protection of SH-SY5Y neuronal cells from
glutamate-induced apoptosis by 3,6'-disinapoyl sucrose, a bioactive
compound isolated from Radix Polygala. J Biomed Biotechnol.
2012:1–5. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Xu MF, Xiong YY, Liu JK, Qian JJ, Zhu L
and Gao J: Asiatic acid, a pentacyclic triterpene in Centella
asiatica, attenuates glutamate-induced cognitive deficits in mice
and apoptosis in SH-SY5Y cells. Acta Pharmacol Sin. 33:578–587.
2012.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Santos-Carvalho A, Elvas F, Alvaro AR,
Ambrósio AF and Cavadas C: Neuropeptide Y receptors activation
protects rat retinal neural cells against necrotic and apoptotic
cell death induced by glutamate. Cell Death Dis.
4(e636)2013.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Ahrens VM, Bellmann-Sickert K and
Beck-Sickinger AG: Peptides and peptide conjugates: Therapeutics on
the upward path. Future Med Chem. 4:1567–1586. 2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Brazil DP, Yang ZZ and Hemmings BA:
Advances in protein kinase B signalling: AKTion on multiple fronts.
Trends Biochem Sci. 29:233–242. 2004.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Manning BD and Cantley LC: AKT/PKB
signaling: Navigating downstream. Cell. 129:1261–1274.
2007.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Hanada M, Feng J and Hemmings BA:
Structure, regulation and function of PKB/AKT - a major therapeutic
target. Biochim Biophys Acta. 1697:3–16. 2004.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lim JY, Park SI, Oh JH, Kim SM, Jeong CH,
Jun JA, Lee KS, Oh W, Lee JK and Jeun SS: Brain-derived
neurotrophic factor stimulates the neural differentiation of human
umbilical cord blood-derived mesenchymal stem cells and survival of
differentiated cells through MAPK/ERK and PI3K/Akt-dependent
signaling pathways. J Neurosci Res. 86:2168–2178. 2008.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Luo C, Huang Q, Yuan X, Yang Y, Wang B,
Huang Z, Tang L and Sun H: Abdominal paracentesis drainage
attenuates severe acute pancreatitis by enhancing cell apoptosis
via PI3K/AKT signaling pathway. Apoptosis. 25:290–303.
2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Sweatt JD: Mitogen-activated protein
kinases in synaptic plasticity and memory. Curr Opin Neurobiol.
14:311–317. 2004.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Fukunaga K and Miyamoto E: Role of MAP
kinase in neurons. Mol Neurobiol. 16:79–95. 1998.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Stanciu M, Wang Y, Kentor R, Burke N,
Watkins S, Kress G, Reynolds I, Klann E, Angiolieri MR, Johnson JW,
et al: Persistent activation of ERK contributes to
glutamate-induced oxidative toxicity in a neuronal cell line and
primary cortical neuron cultures. J Biol Chem. 275:12200–12206.
2000.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Chu CT, Levinthal DJ, Kulich SM, Chalovich
EM and DeFranco DB: Oxidative neuronal injury. The dark side of
ERK1/2. Eur J Biochem. 271:2060–2066. 2004.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Subramaniam S, Zirrgiebel U, von Bohlen
Und Halbach O, Strelau J, Laliberté C, Kaplan DR and Unsicker K:
ERK activation promotes neuronal degeneration predominantly through
plasma membrane damage and independently of caspase-3. J Cell Biol.
165:357–369. 2004.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Kolch W: Meaningful relationships: The
regulation of the Ras/Raf/MEK/ERK pathway by protein interactions.
Biochem J. 351:289–305. 2000.PubMed/NCBI
|
|
47
|
Wennström S and Downward J: Role of
phosphoinositide 3-kinase in activation of ras and
mitogen-activated protein kinase by epidermal growth factor. Mol
Cell Biol. 19:4279–4288. 1999.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Rommel C, Clarke BA, Zimmermann S, Nuñez
L, Rossman R, Reid K, Moelling K, Yancopoulos GD and Glass DJ:
Differentiation stage-specific inhibition of the Raf-MEK-ERK
pathway by Akt. Science. 286:1738–1741. 1999.PubMed/NCBI View Article : Google Scholar
|