Introduction
Nasopharyngeal carcinoma (NPC) is an epithelial
carcinoma that arises from the lining of the nasopharyngeal mucosa
(1). The tumors are usually
observed at the pharyngeal recess in the nasopharynx. Despite
originating from similar cell or tissue lineages, NPC is distinct
from other epithelial head and neck tumors (2). NPC is characterized by poorly or
undifferentiated carcinomas. It differs from other head and neck
squamous cell carcinomas in several ways, including its association
with the Epstein-Barr virus (EBV), increased radio- and
chemosensitivity, and a greater propensity for distant metastases
(3). Compared with other cancers,
NPC is relatively uncommon. The incidence of NPC has a marked
geographical distribution; it is most prevalent in South China,
with an annual incidence of ~30 cases per 100,000 population
(4). NPC is associated with
multiple risk factors, including genetic susceptibility and
environmental factors, such as EBV infection and dietary intake
(5,6).
Proteins of the S100 family are small
Ca2+-binding proteins with a helix-loop-helix (EF-hand
type) binding motif, and have been demonstrated to participate in
the regulation of various intracellular and extracellular
processes, including cell proliferation and differentiation, and
intracellular signaling (7).
Receptor for activated glycation end products (RAGE) is a receptor
for multiple ligands that belongs to the immunoglobulin family. The
dysregulation of RAGE has been indicated to be a critical step in
the development and metastasis of numerous types of tumors,
including head and neck cancer (8,9). RAGE
is able to interact with structurally diverse ligands, and although
these interactions appear to involve oligomerization of the
receptor on the cell surface (10),
the exact mechanism is unknown. Members of the S100 protein family
are known to be RAGE ligands (10).
Several S100 proteins, including S100A12 and S100B, are released
from cells and have the ability to activate RAGE, suggesting that
this may be an important mechanism underlying the extracellular
effects of S100 proteins (11). The
S100P protein is a small isoform of the S100 protein family, the
‘P’ in its name reflecting its original isolation from the placenta
by Becker et al (12) in
1992. S100P is a 95-amino-acid protein, which is encoded by a gene
located on chromosome 4pl. S100P has been reported to interact with
several proteins, both extracellularly and intracellularly, and its
upregulation in various types of human cancer has been shown to be
associated with disease progression, the acquisition of
chemoresistance and poor prognosis (13,14).
In our previous study, the differential expression
of S100P between NPC tissues and the tissues of patients with
benign inflammation was identified using immunohistochemistry, and
S100P was demonstrated to be associated with proliferation and
migration in the C666-1 NPC cell line (15). However, the role of RAGE activation
in S100P signaling has not yet been examined in C666-1 cells and
the mechanism remains unclear.
In the present study, to determine whether the
activation of RAGE by S100P is required for S100P to exert its
effects on cell growth and survival, the activation of RAGE by
S100P was blocked and the effects on cell function and signaling
were investigated using a variety of methods. Mechanisms involving
the activation of extracellular signal-related kinase 1/2 (ERK1/2),
mitogen-activated protein kinase 7 (MAPK7), p38 MAPK, ERK1/2,
NF-κB1 and NF-κB p65 were investigated.
Materials and methods
Patients and tumor samples
In total, 30 patients were included in the study, 15
with NPC and 15 with benign nasopharyngeal inflammation. The
samples were obtained from patients who were hospitalized for
nasopharyngeal biopsy between April 2018 and September 2019 at the
Department of Otolaryngology, Jinshan Hospital, Fudan University
(Shanghai, China). The patients were diagnosed with NPC or benign
nasopharyngeal inflammation through pathological examination. None
of the patients had undergone chemotherapy or radiotherapy, or had
any other tumors. Tissue samples were frozen immediately in liquid
nitrogen and stored at -80˚C until used. The clinical
characteristics of the patients are shown in Table I. The study cohort comprised 16 men
(53.3%) and 14 women (46.7%) with an age range of 25-79 years (mean
age, 50.3±14.9 years; Table I). The
NPC group comprised 10 men and 5 women (mean age, 55.13±15.19
years). The Benign nasopharyngeal inflammation group consisted of 6
men and 9 women (mean age, 45.4±13.88 years). When the collection
of tissue samples took place, the participants signed informed
consent forms to confirm that they agreed to the use of their
tissues in scientific research.
 | Table IClinical characteristics of the
patients. |
Table I
Clinical characteristics of the
patients.
| Characteristic | Total no. of patients
(n=30) | No. of NPC cases
(n=15) | No. of benign
inflammation cases (n=15) |
|---|
| Sex | | | |
|
Male | 16 | 10 | 6 |
|
Female | 14 | 5 | 9 |
| Age (years) | | | |
|
<50 | 14 | 6 | 8 |
|
≥50 | 16 | 9 | 7 |
| Stage | | | |
|
I | | 1 | |
|
II | | 6 | |
|
III | | 6 | |
|
IV | | 2 | |
| Histological
subtype | | | |
|
Non-keratinized | | 15 | |
|
Undifferentiated | | 0 | |
Cell culture and treatments
The C666-1 NPC cell line was purchased from the cell
bank of the Central Laboratory of Xiangya Hospital (Central South
University). The cell line was authenticated by the provider and
tested for Mycoplasma contamination. The C666-1 cell line
was established by Cheung et al (16) from undifferentiated NPC, and
consistently carries EBV in long-term culture. The C666-1 cells
were cultured in RPMI-1640 medium (Sigma-Aldrich; Merck KGaA)
supplemented with 10% certified fetal bovine serum (FBS; Biological
Industries) and maintained in a humidified incubator containing 5%
CO2 at 37˚C. The medium was replaced with fresh culture
medium every 3-4 days. The following three groups of C666-1 cells
were used in subsequent assays: The S100P protein group, treated
with S100P (cat. no. Ag19115; Proteintech Group, Inc.), the FPS-ZM1
group, treated with the RAGE inhibitor FPS-ZM1 (cat. no. 6237;
Tocris Bioscience) (17), and the
untreated control group. The S100P protein and RAGE inhibitor were
separately dissolved in cell culture medium and prepared for
immediate use.
Western blot analysis
Total proteins were extracted from the tissue
samples and cells following lysis with sodium dodecyl sulfate (SDS)
lysis buffer containing phenylmethanesulfonyl fluoride and
proteinase inhibitor (100:1:1; Nanjing KeyGen Biotech Co., Ltd.)
using an automatic grinding machine (JXFSTPRP-24; Jingxin
Technology Co., Ltd.) at 50 Hz for 60 sec. The supernatant was
collected and the protein concentration was determined using a BCA
protein assay (Beyotime Institute of Biotechnology). Western
blotting was performed using a standard method (18). Quantified proteins (50 µg per lane)
were separated by electrophoresis using 8-12% SDS polyacrylamide
gels and electroblotted onto polyvinylidene fluoride membranes. The
membranes were blocked with 5% non-fat milk in tris buffer saline
containing 0.05% Tween-20 for 1 h at room temperature. The
membranes were then incubated sequentially at 4˚C overnight with
primary antibodies. Primary antibodies targeting S100P (cat. no.
11803-1-AP; 1:1,000), RAGE (cat. no. 16346-1-AP; 1:1,000), ERK1/2
(cat. no. 66192, 1:1,000), p38 (cat. no. 66234; 1:1,000), MAPK7
(cat. no. 10036-2-AP; 1:1,000), NF-κB1 (cat. no. 14220-1-AP;
1:1,000), p65 (cat. no. 10745-1-AP; 1:1,000) and GAPDH (cat. no.
10494-1-AP; 1:5,000) were acquired from Proteintech Group, Inc. and
antibodies targeting phosphorylated (p)-ERK1/2 (ab201015; 1:1,000),
p-p38 (ab4822; 1:1,000) and p-MAPK7 (ab5686, 1:1,000) Abcam.
Following this, the membranes were incubated with HRP-conjugated
secondary antibodies (cat. no. SA00001-2; Proteintech; 1:5,000) for
1 h at room temperature. The resulting protein-antibody complexes
were detected using an ECL system (Merck KGaA) and then visualized
using AllDoc version 2.2.1.0 software with the Tanon 4500 imaging
system (Tanon Science and Technology Co., Ltd.).
Cell Counting Kit-8 (CCK-8) assay
The C666-1 cells were seeded at a density of 3,000
cells/well in 96-well plates. After 24 h, the cell viability was
determined using a CCK-8 assay (Dojindo Molecular Technologies,
Inc.), by measurement of the optical density at 450 nm (OD450)
according to the kit protocol. Following this, a range of
concentrations (10, 100, 500, 1,000 and 5,000 ng/ml) of S100P
protein or FPS-ZM1 were added to the wells, an untreated control
group was established, and the cells were cultured for 6 h at 37˚C.
In subsequent experiments, the S100P protein and FPS-ZM1 were used
at a concentration of 1,000 ng/ml, since at this concentration, the
inhibition and proliferation rates were ~50%, respectively. Cell
viability was assessed using the CCK-8 assay after incubation for
6, 12 and 24 h by adding 10 µl CCK-8 solution to each well,
incubating for 30 min to 4 h at 37˚C according to the
manufacturer's protocol, after which the OD450 value was
measured.
Colony formation assay
The C666-1 cells were cultured in 6-well plates at a
density of 2,000 cells/well. After 24 h, 1,000 ng/ml S100P protein
or FPS-ZM1 was added to each well and the cells were cultured for a
further ~14 days at 37˚C. The colonies were then stained with 0.4%
crystal violet (Beyotime Institute of Biotechnology). The numbers
of colonies that contained >50 cells were counted and averaged
(19).
Wound healing assays
The C666-1 cells were cultured in 6-well plates
until they reached confluency. The monolayers were scratched and
the detached cells were washed away using PBS solution. Then, 1,000
ng/ml S100P protein or FPS-ZM1 was added to each well and the cells
were cultured in serum-free RPMI-1640 for 24 h at 37˚C. The width
of the wound was measured at 0 and 24 h with an inverted microscope
equipped with a digital camera (Olympus Corporation).
Transwell migration assay
Transwell chambers (pore size, 8 µm; Corning Life
Sciences) were used for the Transwell migration assay. A total of
2x105 cells suspended in 100 µl serum-free medium with
or without S100P protein or FPS-ZM1 were added to the upper
chamber, and medium containing 20% FBS was added to the lower
chamber. The chambers were incubated for 24 h at 37˚C.
Subsequently, the cells that had migrated through the membrane were
fixed, stained with crystal violet for 20 min at room temperature
and counted in 5 random fields with an inverted microscope equipped
with a digital camera (Olympus Corporation).
Statistical analysis
All experimental assays were conducted as at least
three independent experiments. Data are expressed as the mean ± SD.
Data analysis was performed using SPSS version 26.0 (IBM Corp.).
Comparison of the protein levels between the NPC and benignly
inflamed tissues was performed using independent-sample t-tests.
Kruskal-Wallis and Mann-Whitney U tests were used to analyze the
differences in S100P expression level among or between patients
according to disease stage, age and gender. Comparisons of multiple
groups in other assays were performed by one-way ANOVA followed by
Bonferroni post hoc tests for the multiple comparisons. P<0.05
was considered to indicate a statistically significant
difference.
Results
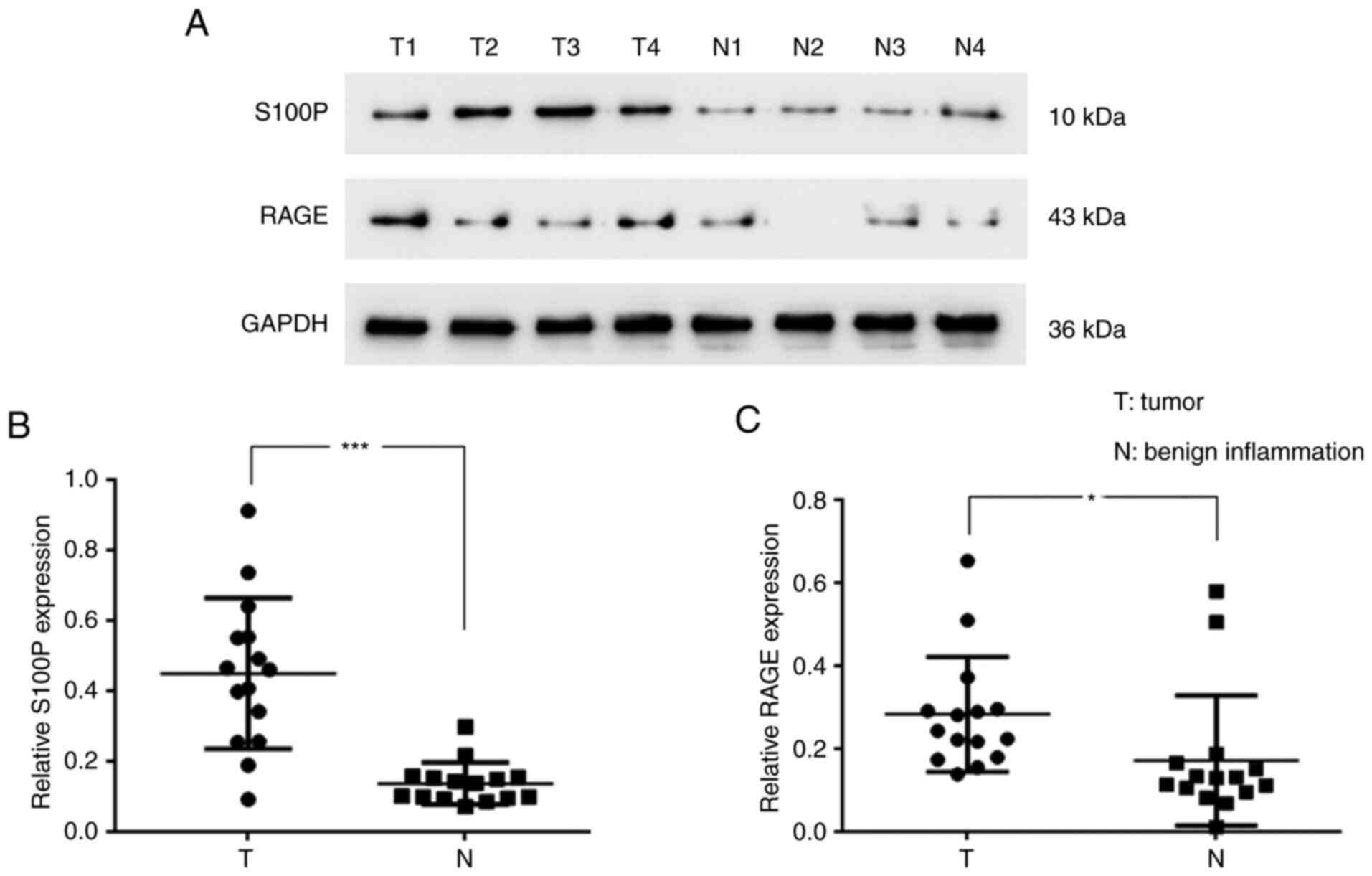
Protein expression of S100P and RAGE
in NPC
To evaluate the levels of S100P and RAGE in
nasopharyngeal tumors, the tumor samples of 15 patients with NPC
and tissues of 15 patients with benign nasopharygeal inflammation
were analyzed. The western blotting results revealed that S100P
(P<0.001; Fig. 1A and B) and RAGE (P<0.05; Fig. 1A and C) were significantly upregulated in the
NPC tissues compared with the tissues with benign inflammation. The
associations of the S100P expression level with disease stage and
the age and gender of the patients were also analyzed. The S100P
expression level was found to differ significantly according to
disease stage when analyzed using a Kruskal-Wallis test (P<0.01;
Table II); however, no significant
difference in S100P expression level was detected according to the
age and gender of the patients when analyzed using a Kruskal-Wallis
and Mann-Whitney U test (Table
II).
| Table IIDifferences in S100P expression
levels according to the disease stage, age and sex of patients. |
Table II
Differences in S100P expression
levels according to the disease stage, age and sex of patients.
| Characteristic | Number of cases
(n=30) | Relative S100P
expression | S100P expression
(P-value) |
|---|
| Sex | | | 0.058 |
|
Male | 16 | 0.327±0.179 | |
|
Female | 14 | 0.254±0.264 | |
| Age (years) | | | 0.42 |
|
<50 | 14 | 0.301±0.272 | |
|
≥50 | 16 | 0.308±0.176 | |
| Stage | | | 0.017 |
|
Benign
inflammation | 15 | 0.137±0.059 | |
|
I | 1 | 0.092 | |
|
II | 6 | 0.417±0.267 | |
|
III | 6 | 0.493±0.138 | 0.005 |
|
IV | 2 | 0.596±0.06 | |
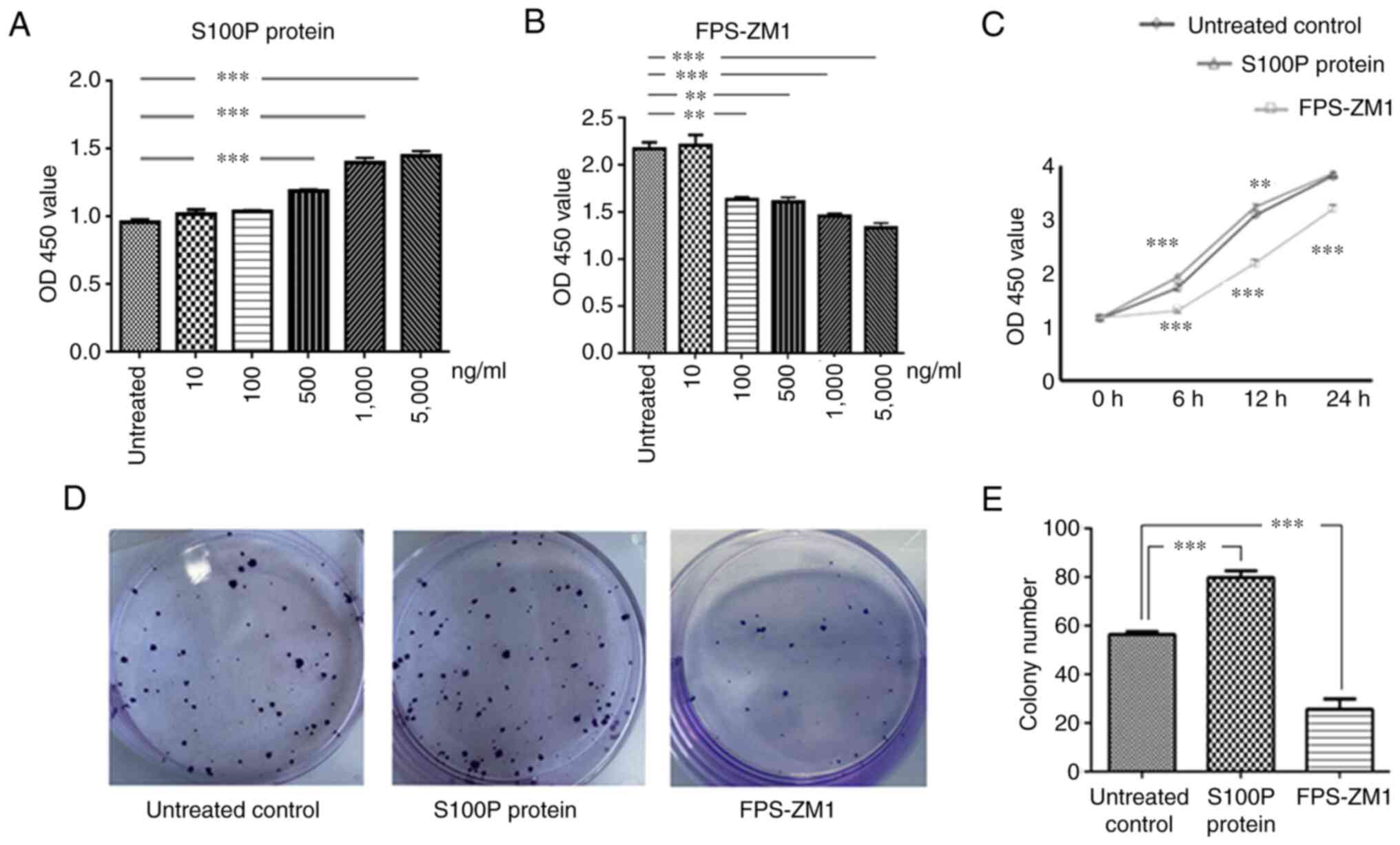
Effects of S100P-RAGE on cell
proliferation
We hypothesized that the effects of S100P in NPC
might be due to its binding with RAGE and the activation of
autocrine signaling mechanisms. Therefore, three groups of C666-1
cells were analyzed, namely the S100P protein group, the FPS-ZM1
group and the untreated control group. These groups were analyzed
in cell viability and migration assays to identify the association
between S100P and RAGE.
Different concentrations of S100P protein and the
RAGE inhibitor FPS-ZM1 were added to C666-1 cells to test their
effects on cell viability. The effects of S100P protein and FPS-ZM1
on cell viability were observed to be concentration-dependent
(P<0.001 and P<0.01, respectively; Fig. 2A and B). Furthermore, the effects of S100P
protein and FPS-ZM1 on cell viability were found to be
time-dependent. Compared with that in the untreated control group,
cell viability was significantly affected at 6 and 12 h following
treatment with S100P protein or FPS-ZM1 at a concentration of 1,000
ng/ml (P<0.01 and P<0.001, respectively; Fig. 2C). The 1,000 ng/ml concentration was
selected for use in subsequent experiments on the basis that it is
the concentration at which the inhibition and proliferation rates
for S100P and FPS-ZM1 are ~50%, respectively (data not shown). The
colony forming assay revealed that the ability of C666-1 cells to
form colonies was increased by treatment with S100P protein and
decreased by treatment with the RAGE inhibitor FPS-ZM1 (P<0.001;
Fig. 2D and E).
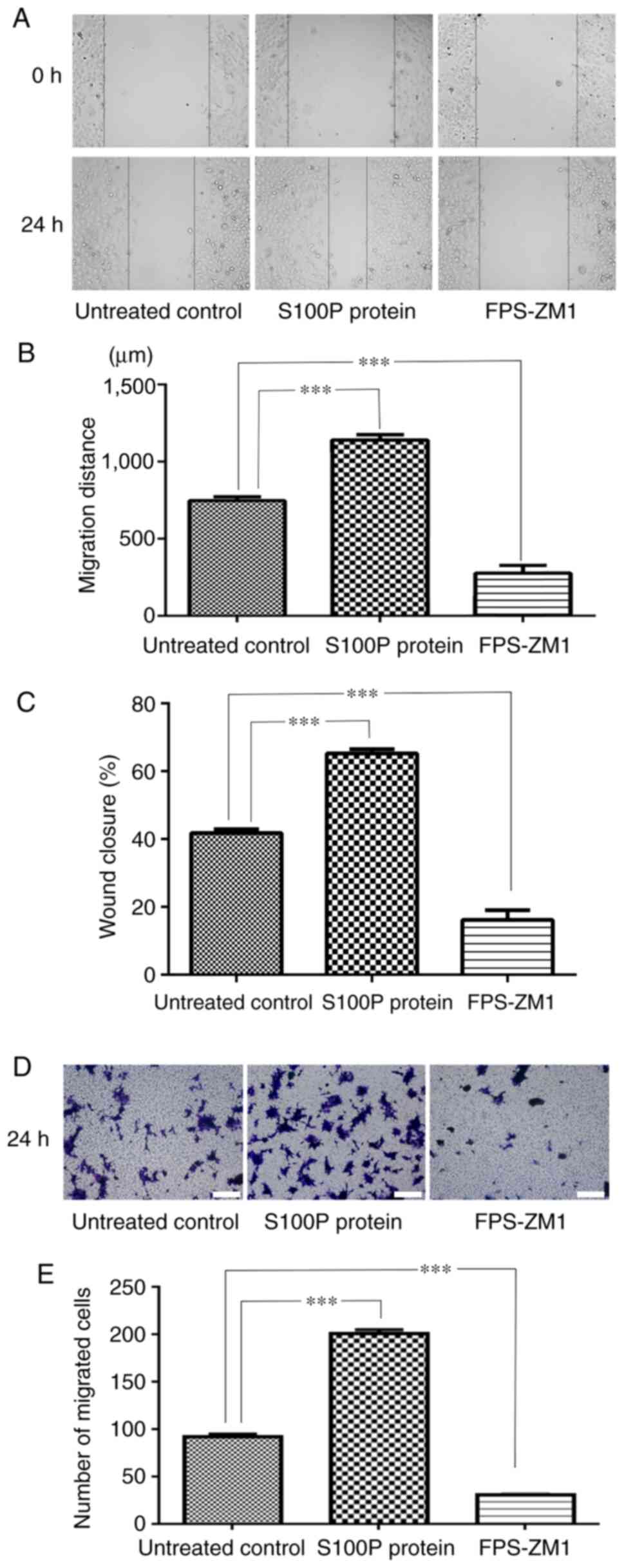
Effects of S100P-RAGE on cell
migration
The effects of S100P and FPS-ZM1 on the migration of
C666-1 cells were assessed by wound healing and Transwell assays.
As presented in Fig. 3A and
B, measurements made 24 h following
wounding of the cell monolayer revealed that wound closure was
induced in the S100P protein group and delayed in FPS-ZM1 group
compared with the untreated control group. The migrated distances
of the cells in the untreated control group, S100P protein group
and FPS-ZM1 group were 746.67±55.28, 1,139.39±77.61 and
276.36±112.21 µm, respectively (P<0.001; Fig. 3A and B). The percentage of wound closure in the
untreated control, S100P protein and FPS-ZM1 groups was calculated
to be 41.79, 65.23 and 16.17%, respectively (P<0.001; Fig. 3C). The Transwell assay revealed that
the addition of S100P protein significantly increased the migration
ability of the C666-1 cells, while the addition of the RAGE
inhibitor FPS-ZM1 reduced the migration ability of the C666-1
cells. The numbers of migrated cells in the untreated control,
S100P protein and FPS-ZM1 groups were 91.8±6.72, 200.8±9.26 and
30.4±2.3, respectively (P<0.001; Fig. 3D and E).
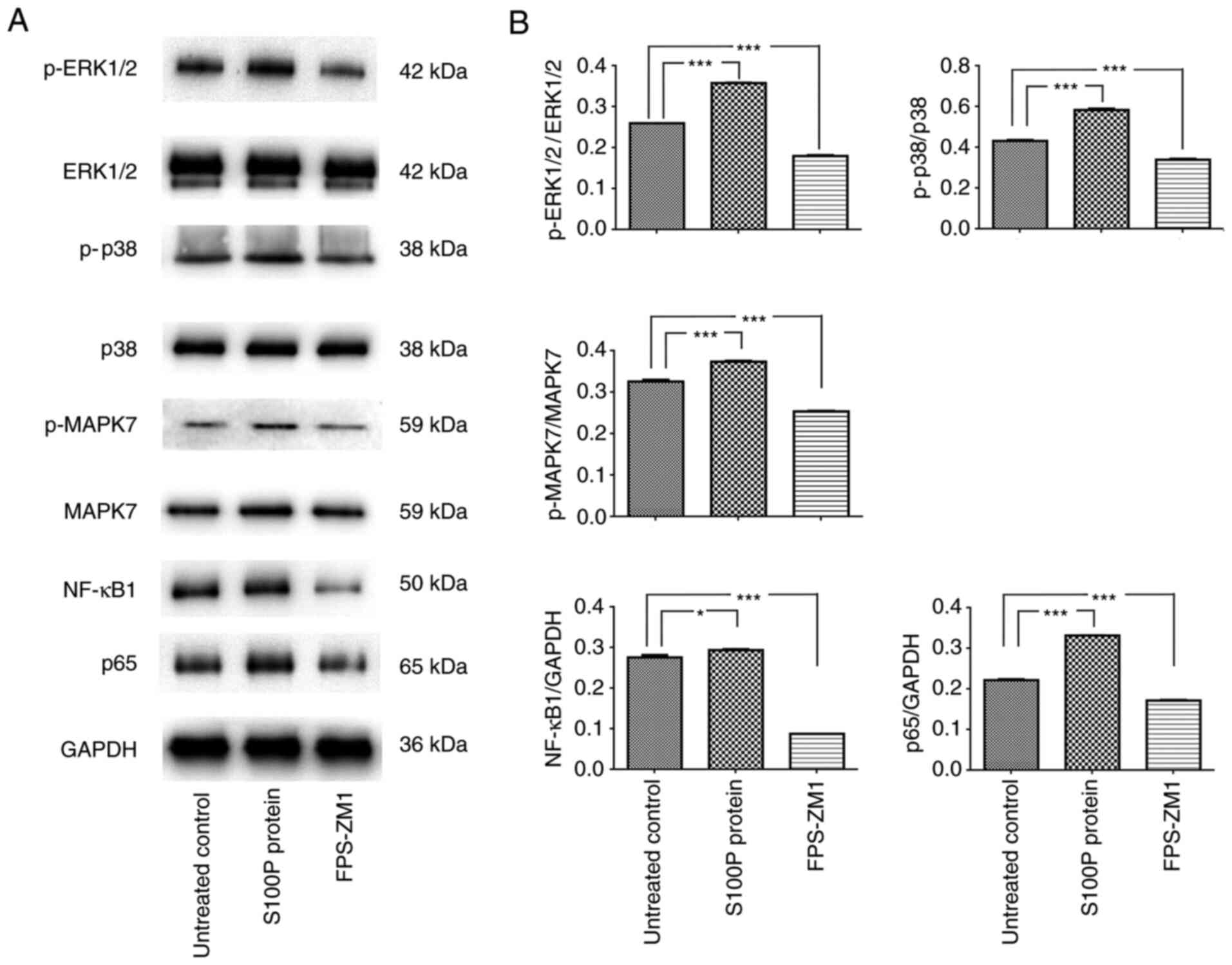
S100P-RAGE activates MAPK and NF-κB
signaling
The MAPK signaling pathway is reported to be
associated with cell proliferation, differentiation, migration,
senescence and apoptosis (20). The
activation of NF-κB is often associated with increased cell
survival (21). Therefore, whether
extracellular S100P activates these transcription factors was
investigated. Consistent with the effects on cell proliferation and
migration, blockade of the S100P-RAGE interaction inhibited the
effects of S100P on MAPK and NF-κB signaling. Following treatment
of the cells with S100P protein, the western blotting results
revealed that the p-ERK1/2/ERK1/2, p-p38/p38 and p-MAPK7/MAPK7
ratios and NF-κB1 and p65 expression levels were significantly
increased compared with those in the untreated cells (NF-κB1,
P<0.05; other proteins, P<0.001). Conversely, following
treatment with the RAGE inhibitor FPS-ZM1, the p-ERK1/2/ERK1/2,
p-p38/p38 and p-MAPK7/MAPK7 ratios and NF-κB1 and p65 expression
levels were decreased compared with those in the untreated cells
(all P<0.001; Fig. 4). These
results indicate that the S100P-RAGE interaction activates MAPK and
NF-κB signaling in NPC cells.
Discussion
The gold standard treatment for NPC is radiation
therapy, which is a local treatment for the cure or palliative
treatment of tumors (22). The cure
rate of early-stage NPC is >90% (23). However, the efficacy of radiation
therapy in advanced stages of NPC is limited owing to radiation
resistance (24). Increased tumor
volumes, tumor hypoxia and the dysregulation of genes can cause
tumor cells to become tolerant to radiation, and thus reduce their
sensitivity (25). Therefore, the
early diagnosis of NPC and the development of novel treatment
approaches are particularly important.
In a previous study, the effect of S100P expression
on the proliferation and migration of C666-1 cells was investigated
by knocking down S100P expression via infection with S100P small
interfering RNA (siRNA) (15).
Following S100P knockdown, the downregulation, proliferation and
migration of the cells were significantly decreased (15). In addition, it was observed that
RAGE expression was downregulated in the cells transfected with
S100P siRNA, as compared with the untreated and negative
siRNA-transfected controls (15).
Notably, certain S100 proteins have been identified to serve a role
in tumors by interacting with receptors, suggesting that
extracellular S100 proteins have important effects (11). In addition, some S100 proteins have
been demonstrated to interact with RAGE in vitro, triggering
RAGE-dependent signal transduction in cell-based assays. For
example, Arumugam et al (26) demonstrated that S100P triggers the
activation of NF-κB in a RAGE-dependent manner through a MAPK
signaling pathway in BxPC3 and SW480 adenocarcinoma cells. However,
the mechanism remains incompletely explored. In the present study,
to determine whether the activation of RAGE by S100P is required
for the effects of S100P on the growth and migration of C666-1
cells, the activation of RAGE by S100P was blocked and its effects
on cell proliferative and migratory behavior and downstream
signaling were investigated. In particular, to elucidate the
molecular mechanisms underlying the activity and function of S100P
in NPC, the role of S100P as an activator of signaling pathways
known to affect the development and progression of numerous types
of cancer was assessed. The most frequently reported signaling
pathways in which S100P participates include ERK1/2, NF-κB and
PI3K/AKT (27). Arumugam et
al (26) revealed that
exogenous S100P increased the survival of NIH3T3 cells and
simultaneously activated ERK in the cells. The MAPK signaling
pathway comprises distinct ERK1/2, JNK1/2/3, p38 MAPK and ERK5
pathways (28). The MAPK/ERK
pathway is reported to be associated with cell proliferation,
differentiation, migration, senescence and apoptosis (20). The present study demonstrated that
the S100P-RAGE interaction is associated with MAPK activation in
C666-1 cells.
S100P has been reported to exert autocrine effects
via RAGE that stimulate cell proliferation and survival via the
NF-κB pathway (26). The
transcription factor NF-κB is a nuclear factor that binds to the
enhancer element of the immunoglobulin κ light-chain of activated B
cells (29). The NF-κB
transcription factor family has five members, known as p65 (RelA),
RelB, c-Rel, NF-κB1 and NF-κB2. The constitutive activation of
NF-κB has been demonstrated to lead to the promotion of cell
proliferation, angiogenesis, invasion and metastasis (21). Consistent with this, the present
study detected a significant increase in NF-κB activation in
NPC.
The inhibition of S100P, using antisense mRNA
retroviral transfection for example, has been shown to decrease
cellular motility and metastatic potential in colon, gastric and
breast cancer cell lines (30-32).
The expression of S100P has been identified to be associated with
the resistance of cancers to several chemotherapeutic agents, and
its silencing sensitizes cancer cells to cisplatin and oxaliplatin
in vitro (33,34). In the present study, the ability of
extracellular S100P to activate MAPK and NF-κB signaling was
investigated. Notably, blockade of the S100P-RAGE interaction
inhibited the effects of S100P on MAPK and NF-κB signaling, which
is similar to its effects on cell proliferation and migration.
The present study has potential limitations,
including the relatively small number of clinical samples and the
use of only one cell line. However, NPC is associated with multiple
risk factors, including EBV infection (5,6), and
the C666-1 cell line consistently carries EBV in long-term cultures
(16). Therefore, only the C666-1
cell line was used. Further studies using primary cultures of NPC
cells and in vivo experiments are required in the future.
Also, although significant data were obtained in the experiments in
the present study, the potential of S100P-RAGE blockade as a
therapeutic treatment for NPC requires further examination in a
larger number of patients.
In conclusion, the present results confirm and
extend those of our previous study, and suggest that extracellular
S100P may participate in the proliferation and migration of C666-1
cells by binding to RAGE and activating the MAPK and NF-κB
signaling pathways. Therefore, blocking S100P-RAGE function might
also be expected to improve the response of NPC to therapeutic
treatments.
Acknowledgements
Not applicable.
Funding
This study was supported by funding from the Jinshan District
Science and Technology Innovation project (grant no. 2017-3-07) and
the Qihang Plan of Jinshan Hospital of Fudan University (grant no.
2018-JSYYQH-02).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HJ performed the diagnostic investigations and
biopsy treatments. CW performed the experiments and wrote the
manuscript. XW and CW confirm the authenticity of all the raw data.
XW performed the statistical analysis and analyzed the data. AH
performed and guided experiment operations. YW collected and
analyzed pathological data. All the authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
All experimental procedures and protocols were
approved by the Ethics Committee of Jinshan Hospital Affiliated to
Fudan University (approval no. 2018-05-01). The participants signed
informed consent forms for use of their tissues in scientific
research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tian Y, Tang L, Yi P, Pan Q, Han Y, Shi Y,
Rao S, Tan S, Xia L, Lin J, et al: miRNAs in radiotherapy
resistance of nasopharyngeal carcinoma. J Cancer. 11:3976–3985.
2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Chen YP, Chan ATC, Le QT, Blanchard P, Sun
Y and Ma J: Nasopharyngeal carcinoma. Lancet. 394:64–80.
2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lee AW, Ng WT, Chan YH, Sze H, Chan C and
Lam TH: The battle against nasopharyngeal cancer. Radiother Oncol.
104:272–278. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Cao SM, Simons MJ and Qian CN: The
prevalence and prevention of nasopharyngeal carcinoma in China.
Chin J Cancer. 30:114–119. 2011.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Tu C, Zeng Z, Qi P, Li X, Yu Z, Guo C,
Xiong F, Xiang B, Zhou M, Gong Z, et al: Genome-wide analysis of 18
epstein-barr viruses isolated from primary nasopharyngeal carcinoma
biopsy specimens. J Virol. 91:e00301–e00317. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Lam WKJ and Chan JYK: Recent advances in
the management of nasopharyngeal carcinoma. F1000Res.
7(F1000)2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Nakayama H, Ohuchida K, Yonenaga A, Sagara
A, Ando Y, Kibe S, Takesue S, Abe T, Endo S, Koikawa K, et al:
S100P regulates the collective invasion of pancreatic cancer cells
into the lymphatic endothelial monolayer. Int J Oncol. 55:211–222.
2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Sasahira T, Kirita T, Bhawal UK, Yamamoto
K, Ohmori H, Fujii K and Kuniyasu H: Receptor for advanced
glycation end products (RAGE) is important in the prediction of
recurrence in human oral squamous cell carcinoma. Histopathology.
51:166–172. 2007.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Choi J, Lee MK, Oh KH, Kim YS, Choi HY,
Baek SK, Jung KY, Woo JS, Lee SH and Kwon SY: Interaction effect
between the receptor for advanced glycation end products (RAGE) and
high-mobility group box-1 (HMGB-1) for the migration of a squamous
cell carcinoma cell line. Tumori. 97:196–202. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
10
|
Leclerc E, Fritz G, Vetter SW and Heizmann
CW: Binding of S100 proteins to RAGE: An update. Biochim Biophys
Acta. 1793:993–1007. 2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Hofmann MA, Drury S, Fu C, Qu W, Taguchi
A, Lu Y, Avila C, Kambham N, Bierhaus A, Nawroth P, et al: RAGE
mediates a novel proinflammatory axis: A central cell surface
receptor for S100/calgranulin polypeptides. Cell. 97:889–901.
1999.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Becker T, Gerke V, Kube E and Weber K:
S100P, a novel Ca(2+)-binding protein from human placenta. cDNA
cloning, recombinant protein expression and Ca2+ binding
properties. Eur J Biochem. 207:541–547. 1992.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Prica F, Radon T, Cheng Y and
Crnogorac-Jurcevic T: The life and works of S100P-from conception
to cancer. Am J Cancer Res. 6:562–576. 2016.PubMed/NCBI
|
|
14
|
Arumugam T, Simeone DM, Van Golen K and
Logsdon CD: S100P promotes pancreatic cancer growth, survival, and
invasion. Clin Cancer Res. 11:5356–5364. 2005.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Liu Y, Wang C, Shan X, Wu J, Liu H, Liu H,
Zhang J, Xu W, Sha Z, He J and Fan J: S100P is associated with
proliferation migration in nasopharyngeal carcinoma. Oncol Lett.
14:525–532. 2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Cheung ST, Huang DP, Hui AB, Lo KW, Ko CW,
Tsang YS, Wong N, Whitney BM and Lee JC: Nasopharyngeal carcinoma
cell line (C666-1) consistently harbouring Epstein-Barr virus. Int
J Cancer. 83:121–126. 1999.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hudson BI and Lippman ME: Targeting RAGE
signaling in inflammatory disease. Annu Rev Med. 69:349–364.
2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Mahmood T and Yang PC: Western blot:
Technique, theory, and trouble shooting. N Am J Med Sci. 4:429–434.
2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Franken NA, Rodermond HM, Stap J, Haveman
J and van Bree C: Clonogenic assay of cells in vitro. Nat Protoc.
1:2315–2319. 2006.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Pramanik KC, Makena MR, Bhowmick K and
Pandey MK: Advancement of NF-κB signaling pathway: A novel target
in pancreatic cancer. Int J Mol Sci. 19(3890)2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Huang T, Yin L, Wu J, Gu JJ, Wu JZ, Chen
D, Yu HL, Ding K, Zhang N, Du MY, et al: MicroRNA-19b-3p regulates
nasopharyngeal carcinoma radiosensitivity by targeting
TNFAIP3/NF-κB axis. J Exp Clin Cancer Res. 35(188)2016.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Chan AT: Current treatment of
nasopharyngeal carcinoma. Eur J Cancer. 47 (Suppl 3):S302–S303.
2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Yang H, Zhang G, Che X and Yu S: Slug
inhibition increases radiosensitivity of nasopharyngeal carcinoma
cell line C666-1. Exp Ther Med. 15:3477–3482. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Barker HE, Paget JT, Khan AA and
Harrington KJ: The tumour microenvironment after radiotherapy:
Mechanisms of resistance and recurrence. Nat Rev Cancer.
15:409–425. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
26
|
Arumugam T, Simeone DM, Schmidt AM and
Logsdon CD: S100P stimulates cell proliferation and survival via
receptor for activated glycation end products (RAGE). J Biol Chem.
279:5059–5065. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Parkkila S, Pan PW, Ward A, Gibadulinova
A, Oveckova I, Pastorekova S, Pastorek J, Martinez AR, Helin HO and
Isola J: The calcium-binding protein S100P in normal and malignant
human tissues. BMC Clin Pathol. 8(2)2008.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Stecca B and Rovida E: Impact of ERK5 on
the hallmarks of cancer. Int J Mol Sci. 20(1426)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Sen R and Baltimore D: Multiple nuclear
factors interact with the immunoglobulin enhancer sequences. Cell.
46:705–716. 1986.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Jiang L, Lai YK, Zhang J, Wang H, Lin MC,
He ML and Kung HF: Targeting S100P inhibits colon cancer growth and
metastasis by Lentivirus-mediated RNA interference and proteomic
analysis. Mol Med. 17:709–716. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ning X, Sun S, Hong L, Liang J, Liu L, Han
S, Liu Z, Shi Y, Li Y, Gong W, et al: Calcyclin-binding protein
inhibits proliferation, tumorigenicity, and invasion of gastric
cancer. Mol Cancer Res. 5:1254–1262. 2007.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Beissel B, Silva ID, Pesquero JB, Russo J,
Schor N and Bellini MH: S-phase reduction in T47D human breast
cancer epithelial cells induced by an S100P antisense-retroviral
construct. Oncol Rep. 17:611–615. 2007.PubMed/NCBI
|
|
33
|
Zhang YW, Zheng Y, Wang JZ, Lu XX, Wang Z,
Chen LB, Guan XX and Tong JD: Integrated analysis of DNA
methylation and mRNA expression profiling reveals candidate genes
associated with cisplatin resistance in non-small cell lung cancer.
Epigenetics. 9:896–909. 2014.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Tang H, Liu YJ, Liu M and Li X:
Establishment and gene analysis of an oxaliplatin-resistant colon
cancer cell line THC8307/L-OHP. Anticancer Drugs. 18:633–639.
2007.PubMed/NCBI View Article : Google Scholar
|