Introduction
Curcumin is a phenolic pigment extracted from
turmeric (Curcuma longa) rhizomes with anti-inflammatory,
antioxidant and anti-cancer properties (1-5).
In the gastrointestinal tract, curcumin plays important
anti-inflammatory and tumor-suppressive roles (6-8).
Our previous study suggested that gastrointestinal food propulsion
was delayed by exogenous nitric oxide (NO) and atropine in mice and
that curcumin could attenuate this effect (9). However, the mechanism of action of
curcumin remains unclear.
NO synthase (NOS) generates NO using L-arginine as a
precursor in vivo. NO is non-cholinergic and non-adrenergic
neurotransmitter that inhibits gastrointestinal motility by
relaxing gastrointestinal smooth muscles (10). Atropine is an antagonist of
acetylcholine receptors (AChRs) that inhibits the contracting
effect of ACh on gastrointestinal smooth muscles (11). Moreover, NO also can inhibit
gastrointestinal motility by disrupting the activity of ICC
(12) and ACh can stimulate ICC to
regulate gastrointestinal motility by combining with the
corresponding neurotransmitter receptor (13). ICC are mesenchymal cells that serve
a key role in normal gastrointestinal motility (14,15).
ICC mediate the interaction between the autonomic nervous system
and smooth muscle cells of the digestive tract. Thus, ICC act as
the ‘pacemaker’ of the gastrointestinal tract by generating
spontaneous electrical slow waves and regulating rhythmic
peristalsis (16). A previous study
reported that ICC depletion and damage, network disruption
(17,18) and the decreased expression of
calcium-activated chloride channel (19) may lead to aberrant slow wave
initiation and conduction. The c-kit tyrosine kinase receptor is an
established marker of ICC (20).
Indeed, c-kit and its ligand, stem cell factor (SCF), regulate ICC
proliferation, development and functional maintenance (21). Insufficient c-kit/SCF signaling
contributes to intestinal motility dysfunction induced by hypoxia
in neonatal mice (22). Anoctamin 1
(Ano1) is a calcium-activated chloride channel identified as a
selective molecular marker ICC in the human and murine
gastrointestinal tracts (23). Slow
gastrointestinal contraction depends on Ano1 activation in ICC
(24). In addition, the gap
junction protein connexin 43 (CX43) also serves a key role in the
transmission and regulation of gastrointestinal motility (25,26).
The aim of the present study was to determine the
mechanisms through which curcumin improves gastrointestinal
motility using L-arginine and atropine-induced murine models of
functional gastric emptying disorder. In addition to NO and ACh
levels, the expression levels of several ICC markers, including
C-kit, Ano1 and CX43, were also examined in the stomach to
determine whether curcumin alters ICC.
Materials and methods
Chemicals
Curcumin (Sigma-Aldrich; Merck KGaA) was mixed in a
5% gum arabic solution to a concentration of 25 g/l for
intragastric administration. L-arginine (Sigma-Aldrich; Merck KGaA)
was mixed with a 5% gum arabic suspension for a final concentration
of 250 g/l. Atropine Sulfate Injection (Tianjin Pharmaceutical
Group Xinzheng Co., Ltd.) was diluted with saline to a
concentration of 0.075 g/l.
Animals
Because hormone secretion in female mice may affect
experimental results, male mice were exclusively used in the
present study. A total of 40 male Kunming mice (3 weeks) weighing
19-22 g were supplied by Jinan Pengyue Experimental Animal Co. Ltd.
The mice were housed at 18-22˚C and 50-60% humidity, in a 12-h
light/dark cycle with free access to food and water. They were
allowed to acclimatize to their surroundings for a week before
experimentation. All animal experiments were approved by The Animal
Ethics Committee of Qingdao University (approval no. QYFY WZLL
2017-10-16).
L-arginine-induced model
establishment
Mice were randomly divided into normal (untreated)
control, curcumin, L-arginine and curcumin + L-arginine groups
(n=10 mice/group). Curcumin (200 mg/kg) was administered
intragastrically in a 0.2-ml volume once per day for 15 days, as
previously described (9). Mice in
the curcumin-untreated groups received gum arabic suspension alone.
From the 11th day, 2 g/kg L-arginine (2 g/kg, 0.2 ml)
suspension was administered intragastrically in a 0.2-ml volume
once per day for 5 consecutive days. Mice in L-arginine-untreated
groups received gum arabic suspension.
Atropine-induced model
establishment
Similar to the L-arginine model, mice were randomly
divided into normal control, curcumin, atropine and curcumin +
atropine groups (n=10 mice/group). Curcumin was administered as
described for the L-arginine-induced model. On the 16th
day, 0.5 mg/kg atropine sulfate was injected intraperitoneally in a
0.2-ml volume. Mice in atropine-untreated groups received 0.2 ml
physiological saline.
Gastric emptying rate
All mice were fasted for 24 h. A volume of 0.8 ml
semi-fluid gum arabic solution was administered intragastrically to
mice for gastric emptying test. The animals were sacrificed by
cervical dislocation 20 min later. The stomach was removed and
weighed to determine the total weight (T), then cut along the
greater curvature to remove the contents. The stomach was then
weighed again to obtain the net weight (t). The gastric emptying
rate (%) was calculated according to the following formula [1 -
(X/Y)] x 100, where X = T - t, and Y is volume of semi-fluid
solution.
NO content, acetylcholinesterase
(AChE) and NOS activity measurement
Total of 4 g of gastric tissue were ground into a
10% homogenate solution at 4˚C in lysis buffer according to the
manufacturer's instructions (NO assay kit, cat. no. A102-1; AchE
assay kit, cat. no. A024-1-1; NOS typed assay kit, cat. no. A014-1;
all from Nanjing Jiancheng Bioengineering Institute). After a
10-min centrifugation at 2,500-3,500 x g and 4˚C, the supernatant
was collected and combined with the color development reaction
solution from the aforementioned kits. The samples were then
transferred to a 96-well plate and absorbance was measured with an
automatic enzyme marker to calculate NO content (550 nm) and the
activities of NOS (530 nm) and AChE (412 nm).
Reverse transcription-quantitative PCR
(RT-qPCR)
RT-qPCR was carried out to measure the expression of
the AChE, AChR, c-kit, CX43 and Ano1 genes. Total RNA was extracted
from mouse gastric tissue using TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.). A total of 1 µg RNA was reverse
transcribed at 37˚C for 15 min and 85˚C for 5 sec, using the
PrimeScript RT reagent kit (Takara Bio, Inc.). qPCRs were carried
out using SYBR Premix Ex Taq II (Takara Bio, Inc.) on a 7500 Fast
Sequence Detection System (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
Initial denaturation at 95˚C for 30 sec; followed by 40 cycles at
95˚C for 10 sec, 60˚C for 20 sec and 72˚C for 30 sec. All reactions
were set up in triplicate for each sample. Primers were purchased
from Sangon Biotech Co., Ltd. (Table
I). The expression levels of mRNA were calculated using the
2-ΔΔCq method (27).
Gene expression was normalized to a GAPDH internal control, and
data are presented as n-fold expression difference of each sample
relative to the respective control group.
 | Table IPrimer sequences used for reverse
transcription-quantitative PCR. |
Table I
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5'→3') |
|---|
| AchE | F:
ACCTGCTTCTCCCACACCT |
| | R:
GGTTCCCACTCGGTAGTTCA |
| AchR | D:
CATCTTGCTGGCTTTCATCA |
| | R:
CACAGCCAGTAGCCCAGATT |
| C-kit | F:
AGCGTGTGTAAATCGTGTTTG |
| | R:
ACATTCAGCATTCCTCCCATA |
| Ano1 | F:
AGCGGAAGCAGCGCTATGA |
| | R:
GGGTGACAAAGCCGAACTGAA |
| CX43 | F:
ACCCAACAGCAGCAGACTTTGA |
| | R:
GCTTGGACCTTGTCCAGCAG |
| GAPDH | F:
CGGAGTCAACGGATTTGGTCGTAT |
| | R:
AGCCTTCTCCATGGTGGTGAAGAC |
Western blotting
Proteins were extracted from stomach tissue by
homogenization at 4˚C in RIPA lysis buffer supplemented with
phenylmethylsulphonyl fluoride (Beijing Solarbio Science &
Technology Co., Ltd.). The lysate was centrifuged at 10,000 x g and
4˚C for 5 min, and protein concentration in the supernatant was
quantified using a bicinchoninic acid assay kit (Beyotime Institute
of Biotechnology). Protein samples (15 µg) were loaded into each
lane of a 5% stacking gel and 8% resolving gel (Beijing Solarbio
Science & Technology Co., Ltd.) and separated by SDS-PAGE.
Proteins were electrotransferred to PVDF membranes. The membranes
were then blocked in 5% milk for 1 h at room temperature, then
incubated at 37˚C for 1 h with the following primary antibodies
against the following proteins purchased from BIOSS: i) CX43
(1:500), cat. no. bs-0651R; ii) c-kit (1:500), cat. no. bs-0672R;
iii) ano1 (1:300), cat. no. bs-9061R; and iv) GAPDH (1:500), cat.
no. bs-0755R. After three washes with 1X TBS + 0.1% Tween-20, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibody (goat anti-rabbit IgG (H+L)/HRP antibody
(1:3,000), cat. no. bs-40295G-HRP; Beijing Biosynthesis
Biotechnology Co., Ltd.) for 1 h at room temperature. Protein bands
were visualized with ECL Western Blotting Substrate (Thermo Fisher
Scientific, Inc.). Densitometry was carried out using ImageJ
software (version no. 1.52; National Institutes of Health).
Statistical analysis
All assays in the present study were repeated three
times. Data analysis was performed using SPSS 17.0 (SPSS, Inc.).
Data are expressed as the mean ± SD. Multigroup comparisons were
analyzed using ANOVA, followed by Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
Changes in gastric emptying rate in
the L-arginine model
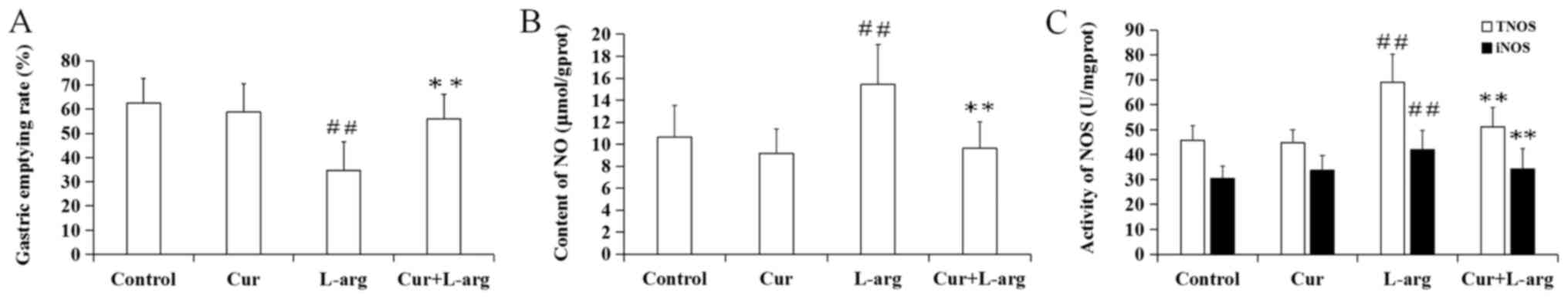
Following L-arginine treatment, the gastric emptying
rate significantly decreased, compared with the control (P<0.01;
Fig. 1A). However, curcumin
pre-treatment significantly improved the delayed gastric emptying
rate caused by L-arginine (P<0.01). In addition, the
administration of curcumin alone had no significant effect on the
gastric emptying rate, compared with control mice (P>0.05).
Changes in NO content, total NOS
(TNOS) and inducible NOS (iNOS) activity in the L-arginine
model
L-arginine administration significantly increased NO
content, as well as TNOS and iNOS activities compared with the
control group (P<0.01). However, pretreatment with curcumin led
to a decrease in all three parameters compared with
L-arginine-treated mice (P<0.01). Moreover, curcumin alone had
no significant effect on NO content, TNOS activity and iNOS
activity compared with control mice (P>0.05, respectively;
Fig. 1B and C).
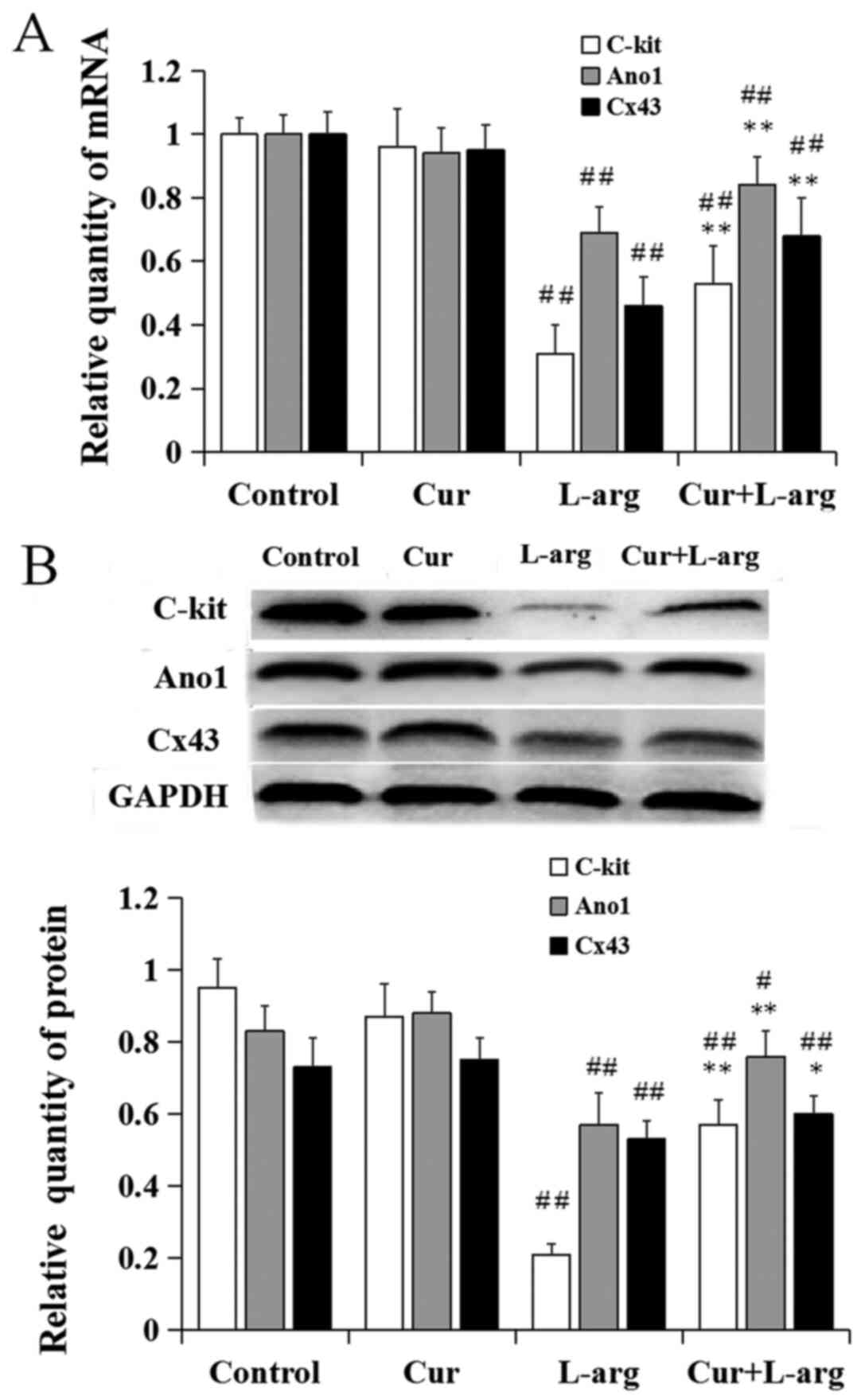
Changes in the expression of ICC
markers in the L-arginine model
Following the administration of L-arginine, the
relative mRNA levels of c-kit, ano1 and CX43 significantly
decreased compared with control (P<0.01). Pretreatment with
curcumin increased c-kit, ano1 and CX43 mRNA expression compared
with L-arginine alone (P<0.01). There was no difference in gene
expression of these markers following treatment with curcumin alone
compared with controls (P>0.05, respectively; Fig. 2A). Similar results for protein
expression levels were demonstrated by western blot analysis
(Fig. 2B).
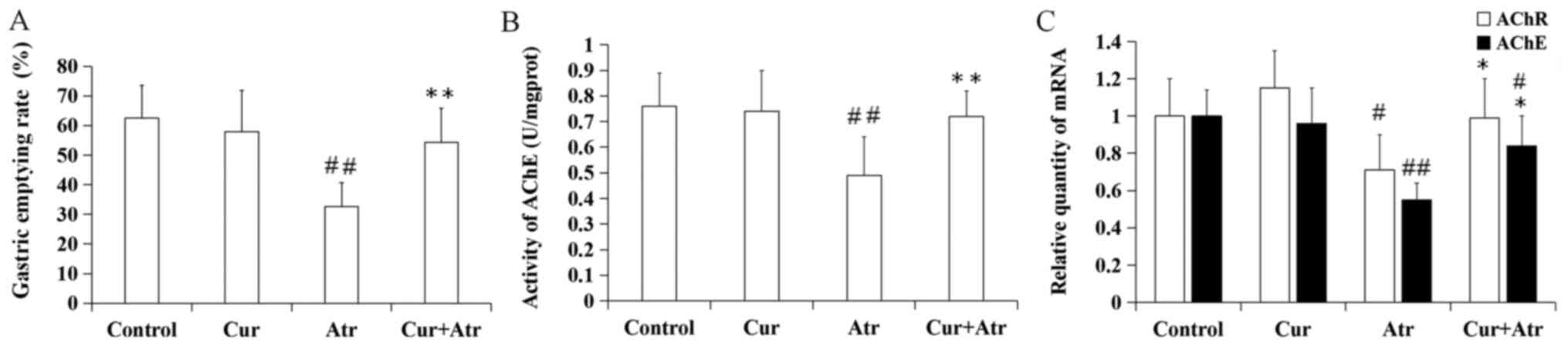
Changes in gastric emptying rates in
the atropine model
The gastric emptying rate significantly decreased
following atropine treatment compared with the control group
(P<0.01). However, curcumin pre-treatment significantly improved
the delayed gastric emptying rate induced by atropine (P<0.01).
Curcumin alone had no significant effect compared with control mice
(P>0.05; Fig. 3A).
Changes in AChE and AChR levels in the
atropine model
Following the administration of atropine, the
activity of AChE (P<0.01), and the relative mRNA expression
levels of AChE (P<0.01) and AChR (P<0.05) significantly
decreased compared with the control group. Pretreatment with
curcumin increased AChE activity (P<0.01) and mRNA expression of
AChE (P<0.05) and AChR (P<0.05) compared with atropine alone.
Curcumin alone had no significant effect on AChE activity
(P>0.05) and gene expression of AChE (P>0.05) and AChR
(P>0.05) (Fig. 3B and C, respectively).
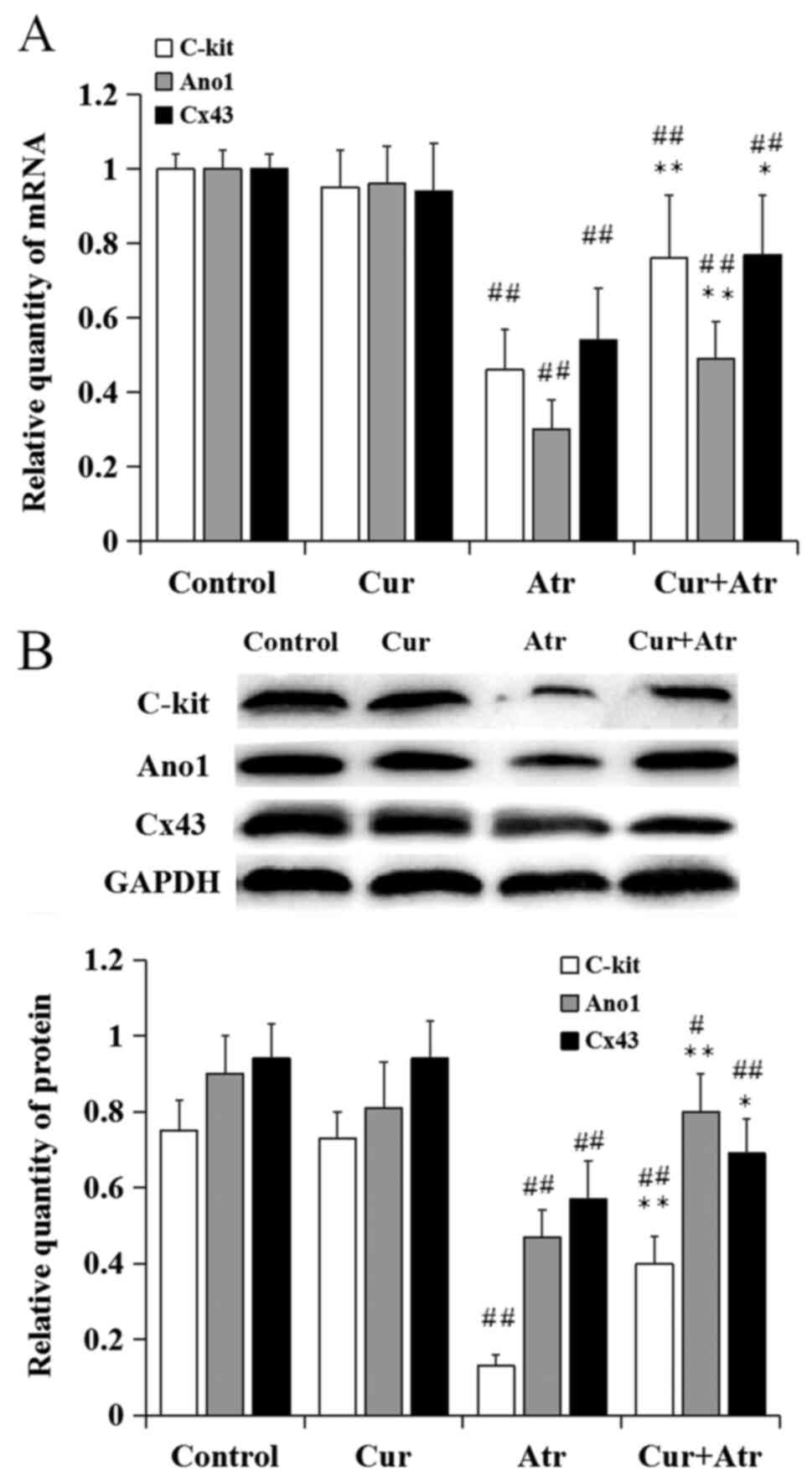
Changes in the expression of ICC
markers in the atropine model
Following atropine treatment, the relative mRNA
levels of c-kit, ano1 and CX43 significantly decreased, compared
with control (P<0.01). However, curcumin pre-treatment
significantly increased the mRNA levels of c-kit (P<0.01), ano1
(P<0.01) and CX43 (P<0.05) compared with atropine alone.
Curcumin alone had no significant effect on the expression of ICC
markers compared with control mice (P>0.05; Fig. 4A). Similar results of the protein
expression levels were demonstrated by western blot analysis
(Fig. 4B).
Discussion
NO acts as an inhibitory neurotransmitter that
relaxes gastrointestinal smooth muscles. However, excessive NO
production can induce gastrointestinal motility dysfunction
(10). Previous studies have
reported that curcumin can inhibit iNOS activity, thereby reducing
NO production in an in vitro model of inflamed human
intestinal mucosa (28,29). In the present study, pretreatment
with curcumin improved the gastric emptying rate in vivo using a
murine model of gastric emptying disorders. In the
L-arginine-induced model, curcumin pre-treatments decreased NO
content and inhibited iNOS activity. It has been reported that
curcumin promotes the degradation of iNOS and suppresses its enzyme
activities in various types of cells (30,31).
However, to the best of our knowledge, there are no previous
reports about curcumin improving gastrointestinal dysfunction by
inhibiting the activity of iNOS. The present findings provide
additional evidence that curcumin ameliorates functional
gastrointestinal dysfunction through the inhibition of iNOS
activity.
ACh is a neurotransmitter that can induce
contraction of gastrointestinal smooth muscles by binding to the
AChR. Once released into the synapse, ACh is rapidly hydrolyzed by
AChE to choline and acetic acid (32), and AChE activity can be measured by
detecting the amount of choline in tissues. A previous study
suggested that ACh release could increase AChE mRNA levels
(33). Thus, the activity and mRNA
level of AChE can reflect the release of ACh. In the
atropine-induced mouse model of gastric emptying disorders,
curcumin pre-treatment led to an increase in the activity and mRNA
levels of AChE, suggesting that curcumin improved gastric emptying
rate by stimulating ACh release in gastric tissues. This was also
confirmed by the increased expression of AChR mRNA. The findings of
the present study are in agreement with previous studies carried
out in animal models. For example, curcumin increases AChE activity
in the mice brain (34). Moreover,
curcumin promoted ACh release in rat brain tissue (35). Nevertheless, how curcumin promotes
ACh release is not fully understood.
Gastrointestinal motility is regulated by
NO-mediated inhibition (12) and
ACh-mediated stimulation (13) of
ICC. In the present study, changes in the expression levels of ICC
markers were also evaluated in murine gastric tissue. In both the
L-arginine and atropine models, c-kit, ano1 and CX43 mRNA levels
significantly decreased. However, pretreatment with curcumin
inhibited the downregulation of these markers, suggesting that
functional gastric emptying disorders caused by L-arginine and
atropine may be related to ICC dysfunction. Thus, curcumin may
alleviate functional gastric emptying disorders partly through an
increase of related ICC signaling.
Through inhibiting exogenous NO production,
promoting ACh release and improving signaling of ICC, curcumin may
improve gastric emptying in the L-arginine and atropine mouse
models, indicating that curcumin may promote delayed gastric
emptying to some degree. Curcumin did not significantly affect the
gastric emptying rate when administered alone. It is possible that
a dose of 200 mg/kg curcumin intragastrically administered was not
sufficient to affect the normal gastrointestinal tract. All indexes
were observed 24 h after last application of curcumin. This time
point fully ruled out the direct stimulation of curcumin in the
stomach, which could more reasonably explain the effects of
curcumin pre-treatment for more than 10 days. The dose and days of
curcumin were chosen according to our previous study (9).
The use of curcumin has also been assessed in
clinical trials, and early-phase trials have highlighted its safety
and efficacy in patients (36). A
previous study suggesting that high doses of curcumin were
well-tolerated orally (37).
However, the experimental biological benefits of curcumin have not
yet been replicated in clinical trials, owing to its low
bioavailability following oral administration (38). Nonetheless, since curcumin
metabolites are detectable in the plasma following oral
administration (38,39), it is possible that the effects of
curcumin are also mediated by its metabolites. In addition, several
strategies have been employed to improve the solubility of curcumin
(40,41), which could improve its use and
applicability.
In conclusion, the present study demonstrated that
curcumin could reduce gastric emptying dysfunction. Thus, in
addition to anti-inflammatory and antioxidant properties, curcumin
could affect gastrointestinal peristalsis through inhibiting
exogenous NO production, promoting ACh release and improving
signaling of ICC. These findings identified a potential application
for curcumin in the prevention of gastrointestinal disorders.
However, the mechanism through which curcumin might exert its
effect on gastric tissue, including ICC signal transduction,
remains unclear. Owing to the complexity of regulation of
gastrointestinal motility, other mechanisms of curcumin should be
further examined.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by The Natural Science
Foundation of Shandong Province (grant no. ZR2014HM033).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PL, BL and XY contributed to the experimental
design, carried out experiments and wrote the manuscript. JY, FS,
HZ and JX carried out experiments and data analysis. All authors
agreed to be accountable for all aspects of the work in ensuring
that questions related to the accuracy or integrity of any part of
the work are appropriately investigated and resolved. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The Qingdao University (Qingdao, China; approval no. QYFY WZLL
2017-10-16).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bhawana Basniwal RK, Buttar HS, Jain VK
and Jain N: Curcumin nanoparticles: preparation, characterization,
and antimicrobial study. J Agric Food Chem. 59:2056–2061.
2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Tsuda T: Curcumin as a functional
food-derived factor: Degradation products, metabolites,
bioactivity, and future perspectives. Food Funct. 9:705–714.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Daily JW, Yang M and Park S: Efficacy of
turmeric extracts and curcumin for alleviating the symptoms of
joint arthritis: A systematic review and meta-analysis of
randomized clinical trials. J Med Food. 19:717–729. 2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Hussain Z, Thu HE, Amjad MW, Hussain F,
Ahmed TA and Khan S: Exploring recent developments to improve
antioxidant, anti-inflammatory and antimicrobial efficacy of
curcumin: A review of new trends and future perspectives. Mater Sci
Eng C. 77:1316–1326. 2017.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yue GG, Kwok HF, Lee JK, Jiang L, Wong EC,
Gao S, Wong HL, Li L, Chan KM, Leung PC, et al: Combined therapy
using bevacizumab and turmeric ethanolic extract (with absorbable
curcumin) exhibited beneficial efficacy in colon cancer mice.
Pharmacol Res. 111:43–57. 2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhang X, Wu J, Ye B, Wang Q, Xie X and
Shen H: Protective effect of curcumin on TNBS-induced intestinal
inflammation is mediated through the JAK/STAT pathway. BMC
Complement Altern Med. 16(299)2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Murphy EA, Davis JM, McClellan JL, Gordon
BT and Carmichael MD: Curcumin's effect on intestinal inflammation
and tumorigenesis in the ApcMin/+ mouse. J Interferon
Cytokine Res. 31:219–226. 2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Roberts JL, Poklepovic A and Booth L:
Curcumin interacts with sildenafil to kill GI tumor cells via
endoplasmic reticulum stress and reactive oxygen/ nitrogen species.
Oncotarget. 8:99451–99469. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Yu J, Xu WH, Sun W, Sun Y, Guo ZL and Yu
XL: Curcumin alleviates the functional gastrointestinal disorders
of mice in vivo. J Med Food. 20:1176–1183. 2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Garella R, Squecco R and Baccari MC:
Site-related effects of relaxin in the gastrointestinal tract
through nitric oxide signaling: An updated report. Curr Protein
Pept Sci. 18:1254–1262. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Eglen RM: Muscarinic receptors and
gastrointestinal tract smooth muscle function. Life Sci.
68:2573–2578. 2001.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kaji N, Nakayama S, Horiguchi K, Iino S,
Ozaki H and Hori M: Disruption of the pacemaker activity of
interstitial cells of Cajal via nitric oxide contributes to
postoperative ileus. Neurogastroenterol Motil.
30(13334)2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Sung TS, Hwang SJ, Koh SD, Bayguinov Y,
Peri LE, Blair PJ, Webb TI, Pardo DM, Rock JR, Sanders KM, et al:
The cells and conductance mediating cholinergic neurotransmission
in the murine proximal stomach. J Physiol. 596:1549–1574.
2018.PubMed/NCBI View
Article : Google Scholar
|
|
14
|
Al-Shboul OA: The importance of
interstitial cells of cajal in the gastrointestinal tract. Saudi J
Gastroenterol. 19:3–15. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Iino S, Ward SM and Sanders KM:
Interstitial cells of Cajal are functionally innervated by
excitatory motor neurones in the murine intestine. J Physiol.
556:521–530. 2004.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Rumessen JJ and Thuneberg L: Pacemaker
cells in the gastrointestinal tract: Interstitial cells of Cajal.
Scand J Gastroenterol Suppl. 216:82–94. 1996.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Chang IY, Glasgow NJ, Takayama I,
Horiguchi K, Sanders KM and Ward SM: Loss of interstitial cells of
Cajal and development of electrical dysfunction in murine small
bowel obstruction. J Physiol. 536:555–568. 2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang TH, Angeli TR, Ishida S, Du P,
Gharibans A, Paskaranandavadivel N, Imai Y, Miyagawa T, Abell TL,
Farrugia G, et al: The influence of interstitial cells of Cajal
loss and aging on slow wave conduction velocity in the human
stomach. Physiol Rep. 8(e14659)2021.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Sanders KM: Spontaneous electrical
activity and rhythmicity in gastrointestinal smooth muscles. Adv
Exp Med Biol. 1124:3–46. 2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Tamada H and Kiyama H: Existence of c-Kit
negative cells with ultrastructural features of interstitial cells
of Cajal in the subserosal layer of the W/W(v) mutant mouse colon.
J Smooth Muscle Res. 51:1–9. 2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Tan YY, Ji ZL, Zhao G, Jiang JR, Wang D
and Wang JM: Decreased SCF/c-kit signaling pathway contributes to
loss of interstitial cells of Cajal in gallstone disease. Int J
Clin Exp Med. 7:4099–4106. 2014.PubMed/NCBI
|
|
22
|
Ren H, Han J, Li Z and Xiong Z: Stem cell
factor/kit signal insufficiency contributes to hypoxia-induced
intestinal motility dysfunctions in neonatal mice. Dig Dis Sci.
62:1193–1203. 2017.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Gomez-Pinilla PJ, Gibbons SJ, Bardsley MR,
Lorincz A, Pozo MJ, Pasricha PJ, Van de Rijn M, West RB, Sarr MG,
Kendrick ML, et al: Ano1 is a selective marker of interstitial
cells of Cajal in the human and mouse gastrointestinal tract. Am J
Physiol Gastrointest Liver Physiol. 296:G1370–G1381.
2009.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Drumm BT, Hennig GW, Battersby MJ,
Cunningham EK, Sung TS, Ward SM, Sanders KM and Baker SA:
Clustering of Ca2+ transients in interstitial cells of
Cajal defines slow wave duration. J Gen Physiol. 149:703–725.
2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Nemeth L, Maddur S and Puri P:
Immunolocalization of the gap junction protein Connexin43 in the
interstitial cells of Cajal in the normal and Hirschsprung's
disease bowel. J Pediatr Surg. 35:823–828. 2000.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zhang G, Xie S, Hu W, Liu Y, Liu M, Liu M
and Chang X: Effects of electroacupuncture on interstitial cells of
Cajal (ICC) ultrastructure and Connexin 43 protein expression in
the gastrointestinal tract of functional dyspepsia (FD) rats. Med
Sci Monit. 22:2021–2027. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Somchit M, Changtam C, Kimseng R, Utaipan
T, Lertcanawanichakul M, Suksamrarn A and Chunglok W:
Demethoxycurcumin from Curcuma longa rhizome suppresses iNOS
induction in an in vitro inflamed human intestinal mucosa model.
Asian Pac J Cancer Prev. 15:1807–1810. 2014.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Dai C, Li B, Zhou Y, Li D, Zhang S, Li H,
Xiao X and Tang S: Curcumin attenuates quinocetone induced
apoptosis and inflammation via the opposite modulation of Nrf2/HO-1
and NF-κB pathway in human hepatocyte L02 cells. Food Chem Toxicol.
95:52–63. 2016.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Ben P, Liu J, Lu C, Xu Y, Xin Y, Fu J,
Huang H, Zhang Z, Gao Y, Luo L, et al: Curcumin promotes
degradation of inducible nitric oxide synthase and suppresses its
enzyme activity in RAW 264.7 cells. Int Immunopharmacol.
11:179–186. 2011.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li M, Wang L, Liu H, Su B, Liu B, Lin W,
Li Z and Chang L: Curcumin inhibits HeLa cell invasion and
migration by decreasing inducible nitric oxide synthase. Nan Fang
Yi Ke Da Xue Xue Bao. 33:1752–1756. 2013.PubMed/NCBI(In Chinese).
|
|
32
|
Trang A and Khandhar PB: Physiology,
Acetylcholinesterase. StatPearls, 2020. https://www.ncbi.nlm.nih.gov/books/NBK539735/.
Accessed July 10, 2020.
|
|
33
|
Cresnar B, Crne-Finderle N, Breskvar K and
Sketelj J: Neural regulation of muscle acetylcholinesterase is
exerted on the level of its mRNA. J Neurosci Res. 38:294–299.
1994.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Abu-Taweel GM: Effects of curcumin on the
social behavior, blood composition, reproductive hormones in plasma
and brain acetylcholinesterase in cadmium intoxicated mice. Saudi J
Biol Sci. 23:219–228. 2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Liu ZJ, Li ZH, Liu L, Tang WX, Wang Y,
Dong MR and Xiao C: Curcumin attenuates beta-amyloid-induced
neuroinflammation via activation of peroxisome
proliferator-activated receptor-gamma function in a rat model of
Alzheimer's disease. Front Pharmacol. 7(261)2016.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Cruz-Correa M, Hylind LM, Marrero JH,
Zahurak ML, Murray-Stewart T, Casero RA Jr, Montgomery EA,
Iacobuzio-Donahue C, Brosens LA, Offerhaus GJ, et al: Efficacy and
safety of curcumin in treatment of intestinal adenomas in patients
with familial adenomatous polyposis. Gastroenterology. 155:668–673.
2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Sharma RA, McLelland HR, Hill KA, Ireson
CR, Euden SA, Manson MM, Pirmohamed M, Marnett LJ, Gescher AJ and
Steward WP: Pharmacodynamic and pharmacokinetic study of oral
Curcuma extract in patients with colorectal cancer. Clin
Cancer Res. 7:1894–1900. 2001.PubMed/NCBI
|
|
38
|
Wang L, Li W, Cheng D, Guo Y, Wu R, Yin R,
Li S, Kuo HC, Hudlikar R, Yang H, et al: Pharmacokinetics and
pharmacodynamics of three oral formulations of curcumin in rats. J
Pharmacokinet Pharmacodyn. 47:131–144. 2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lao CD, Ruffin MT IV, Normolle D, Heath
DD, Murray SI, Bailey JM, Boggs ME, Crowell J, Rock CL and Brenner
DE: Dose escalation of a curcuminoid formulation. BMC Complement
Altern Med. 6(10)2006.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Kurniawansyah F, Quachie L, Mammucari R
and Foster NR: Improving the dissolution properties of curcumin
using dense gas antisolvent technology. Int J Pharm. 521:239–248.
2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Peng S, Li Z, Zou L, Liu W, Liu C and
McClements DJ: Improving curcumin solubility and bioavailability by
encapsulation in saponin-coated curcumin nanoparticles prepared
using a simple pH-driven loading method. Food Funct. 9:1829–1839.
2018.PubMed/NCBI View Article : Google Scholar
|