Introduction
Osteoporosis is a ‘silent disease’ that results in
fragile bones that are prone to fracture and lacks obvious symptoms
at the beginning of the pathological process (1). The loss of bone, decrease in bone
mass, destruction of bone microstructure and increase in bone
brittleness can lead to systemic bone disease in patients with
osteoporosis (2). Osteoporosis can
be classified as primary or secondary according to its etiology.
The etiology of primary osteoporosis is still not clear, but
secondary osteoporosis is often caused by endocrine metabolic
disease (such as hyperthyroidism and hyperparathyroidism) and can
also be caused by certain drugs (such as glucocorticoids) that
affect bone metabolism (3-5).
In general, adequate calcium intake, a healthy diet and physical
activity contribute to good bone health and decrease the risk of
osteoporosis (6-8).
However, age is associated with the onset of osteoporosis, which is
why this disease is considerably more common in older individuals
(9). In China, the prevalence of
osteoporosis in the elderly (>60 years old) is 36%, amongst
which, the incidence is ~23% in males and ~49% in females (10-11).
Fractures are a serious consequence of osteoporosis. In 2010, the
number of fractures caused by osteoporosis in China reached 2.23
million, of which 1.11 million were spinal fractures and 360,000
were pelvic fractures (12).
According to various reports, osteoporosis has become one important
factor affecting the quality of life of middle-aged and elderly
individuals (13,14). Therefore, it is necessary to
identify diagnostic markers and therapeutic targets for improved
diagnosis and treatment of osteoporosis.
Long non-coding RNA (lncRNA) serves a role in cell
proliferation, migration and differentiation, among numerous other
processes (15,16). lncRNA KCNQ1 opposite
strand/antisense transcript 1 (Kcnq1ot1) is an imprinted antisense
lncRNA in the human KCNQ1 locus and is associated with bone
formation (17). Additionally,
Kcnq1ot1 has been shown to facilitate osteogenic differentiation of
human bone marrow-derived mesenchymal stem cells via the microRNA
(miRNA or miR)-320a/Smad5 axis (18). Furthermore, miR-98-5p has been
reported to participate in myocardial differentiation of bone
marrow mesenchymal stem cells via the regulation of T-box
transcription factor 5 (Tbx5) (19). In addition, this miRNA may target
high mobility group AT-hook 2 to inhibit the osteogenic
differentiation of MC3T3-E1 osteoblasts, thereby obstructing bone
regeneration (20). Thus, it was
hypothesized that Kcnq1ot1 may affect osteogenic differentiation by
binding to miR-98-5p. The aim of the present study was to verify
this hypothesis to clarify the role of miR-98-5p in osteogenic
differentiation and identify novel therapeutic targets for the
treatment of osteoporosis.
Materials and methods
Bioinformatics analysis
starBase database (starbase.sysu.edu.cn/index.php/) was used to predict
the binding site of Kcnq1ot1 and miR-98-5p. miR-98-5p and Tbx5
target site was predicted using TargetScan (targetscan.org/vert_72/).
Cell culture
The MC3T3-E1 mouse pre-osteoblast cell line was
sourced from the American Type Culture Collection (cat. no.
CRL-2594) and cultured in α-Minimum Essential Medium (cat. no.
A1049001) supplemented with ribonucleosides, deoxyribonucleosides,
2 mM L-glutamine and 1 mM sodium pyruvate and 10% FBS (all Gibco;
Thermo Fisher Scientific, Inc.) in the absence of ascorbic acid.
Cells were cultured at 37˚C in a humidified incubator with 5%
CO2 and the medium was replaced every 2-3 days.
Induction of osteogenic
differentiation
Osteogenic differentiation of MC3T3-E1 cells was
induced by incubation with osteogenesis-inducing medium at 80%
confluence for 14 days at 37˚C, as previously described (18). The medium consisted of 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.), 5 mM L-glycerophosphate
(Sigma-Aldrich; Merck KGaA), 100 nM dexamethasone (AmyJet
Scientific, Inc.) and 50 mg/ml ascorbic acid (Shanghai Aladdin
Biochemical Technology Co., Ltd.).
Cell transfection
Short hairpin (sh)RNA plasmid for Kcnq1ot1
(sh-Kcnq1ot1; 5'-GCAGAACCAUCGAUGGUGCGU-3'), shRNA targeting Tbx5
(sh-Tbx5; 5'-CGGCUGCUAGUGUCUAUGUUU-3'), shRNA-negative control
(sh-NC; 5'-AGUGCUGCGCACGUGUCUCAU-3'), pcDNA3.1(+)/Kcnq1ot1 plasmid
(pc-Kcnq1ot1; 5'-GGGGTACCCCAGGTGACAAGGTGCAGGCGC-3'), pcDNA3.1
(5'-AUCUCCGGGGUUUACGUAUAC-3'), miR-98-5p antagomir
(antagomiR-98-5p; 5'-AACAAUACAACUUACUACCUCA-3'), antagomiR-NC
(5'-UCACAACCUCCUAGAAAGAGUAGA-3'), miR-98-5p agomir (agomiR-98-5p;
5'-UGAGGUAGUAAGUUGUAUUGUU-3') and agomiR-NC
(5'-UCGCUUGGUGCAGGUCGGG-3') were constructed by Shanghai
GenePharma, Co., Ltd. A final concentration of 100 nM shRNA, pcDNA,
antagomir or agomir of miR-98-5p and NC was transfected into
MC3T3-E1 cells using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) at 37˚C for 48 h. At transfection,
cells were harvested, and reverse transcription-quantitative PCR
(RT-qPCR) was performed to assess the transfection efficiency. On
the 7th day of osteogenic differentiation, transfection as
aforementioned was performed to maintain the altered gene
expression.
Alkaline phosphatase (ALP) activity
assay
ALP activity was measured to assess differentiation
of MC3T3-E1 cells. Briefly, cells were seeded into 12-well plates
at a density of 4x104 cells/ml 37˚C for 7 days.
Subsequently, the cells were fixed in 4% paraformaldehyde for 30
min at room temperature and treated with 0.3 nitro-blue tetrazolium
and 0.15 mg/ml 5-bromo-4-chloro-3-indolyl phosphate (Sigma-Aldrich;
Merck KGaA) at room temperature for 2 h. Cells were then washed
with deionized water and observed under an inverted light
microscope (Nikon Corporation; magnification, x200) at a wavelength
of 405 nm.
Western blotting
Total protein was extracted from MC3T3-E1 cells
using RIPA lysis buffer (Beyotime Institute of Biotechnology). The
protein concentration was determined using a BCA assay kit
(Beyotime Institute of Biotechnology). A total of 30 µg
protein/well was resolved using 10% SDS-PAGE and transferred to a
PVDF membrane. Subsequently, 5% non-fat milk was used to block the
membrane at 37˚C for 1 h, followed by incubation at room
temperature for 1 h with primary antibodies as follows:
Runt-related transcription factor 2 (RUNX2; 1:1,000; cat. no.
12556; Cell Signaling Technology, Inc.), collagen type I α 1
(COL1A1; 1:1,000; cat. no. ab34710; Abcam), osteopontin (1:1,000,
cat. no. ab214050; Abcam), osteocalcin (1:1,000, cat. no. ab133612;
Abcam), Tbx5 (cat. no. ab259980; Abcam) and GAPDH (1:2,500, cat.
no. ab9485; Abcam). The membrane was then incubated with
horseradish peroxidase-conjugated anti-rabbit IgG secondary
antibody (1:2,000, cat. no. #5127; Cell Signaling Technology, Inc.)
at room temperature for 2 h. The bands were visualized by using an
enhanced chemiluminescence (ECL) reagent kit (Shanghai Yeasen
Biotechnology Co., Ltd.) and semi-quantified with Image J software
(Version 1.49; National Institutes of Health).
RT-qPCR
Total RNA was extracted from MC3T3-E1 cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.). RT
was performed to synthesize cDNA from total RNA, using a
PrimeScript RT Reagent kit, according to the manufacturer's
protocol (Takara Bio, Inc.). A SYBR green PCR Master Mix kit (cat.
no. SR1110; Beijing Solarbio Science & Technology Co., Ltd.)
was used for cDNA amplification by qPCR in accordance with the
manufacturer's instructions on an AFD9600 PCR system (Hangzhou AGS
BioTech Co., Ltd.). The primer sequences for PCR were as follows:
Kcnq1ot1 forward, 5'-ACTCACTCACTCACTCACT-3' and reverse,
5'-CTGGCTCCTTCTATCACATT-3'; miR-98-5p forward,
5'-ATCCAGTGCGTGTCGTG-3' and reverse, 5'-TGCTTGAGGTAGTAAGTTG-3';
Tbx5 forward, 5'-AAGTAAAGAATATCCCGTGGTC-3' and reverse,
5'-AGACTCGCTGCTGAAAGG-3'; GAPDH forward, 5'-GGGAAACTGTGGCGTGAT-3'
and reverse, 5'-GAGTGGGTGTCGCTGTTGA-3' and U6 forward,
5'-CTCGCTTCGGCAGCACATATA-3' and reverse,
5'-ACGCTTCACGAATTTGAGTGTC-3'. The thermocycling conditions were as
follows: 94˚C for 60 sec, followed by 40 cycles of 94˚C for 30 sec,
60˚C for 30 sec and 72˚C for 60 sec. The 2-ΔΔCq method
(21) was used to calculate
relative gene expression. GAPDH was used as an internal reference
for Kcnq1ot1 and Tbx5, while U6 was used as the control gene for
miR-98-5p.
Dual luciferase reporter assay
The wild-type (Kcnq1ot1-WT or Tbx5-WT) and mutant
(Kcnq1ot1-MUT or Tbx5-MUT) 3'-untranslated regions were cloned into
a pmirGLO vector (Shanghai GenePharma Co., Ltd.). For luciferase
reporter analysis, MC3T3-E1 cells were co-transfected with
luciferase reporter vectors, agomiR-98-5p
(5'-UGAGGUAGUAAGUUGUAUUGUU-3') and agomiR-NC
(5'-UCGCUUGGUGCAGGUCGGG-3') using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). At 48 h after transfection, the
relative luciferase activities were measured by using a
Dual-Luciferase Reporter Assay (Promega Corporation) and normalized
to Renilla luciferase reporter activity according to the
manufacturer's protocol.
Alizarin red S (ARS) staining
assay
An ARS staining kit (GuideChem) was used to detect
the formation of mineralized nodules in MC3T3-E1 osteoblasts.
Following induction of osteogenesis and cell fixation with 95%
ethanol for 20 min at room temperature, the cells were washed with
PBS (Shanghai Aladdin Biochemical Technology Co., Ltd.) three times
and stained with ARS staining solution for 30 min at room
temperature. The cells were observed and images were obtained using
a light microscope (magnification, x200).
Statistical analysis
All experiments were performed independently three
times. GraphPad Prism version 6.0 (GraphPad Software, Inc.) was
used to analyze the data. Data are presented as the mean ± SD.
One-way ANOVA followed by Tukey's post hoc test was used for
comparisons between multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Kcnq1ot1 expression is increased and
miR-98-5p expression is decreased during osteogenic
differentiation
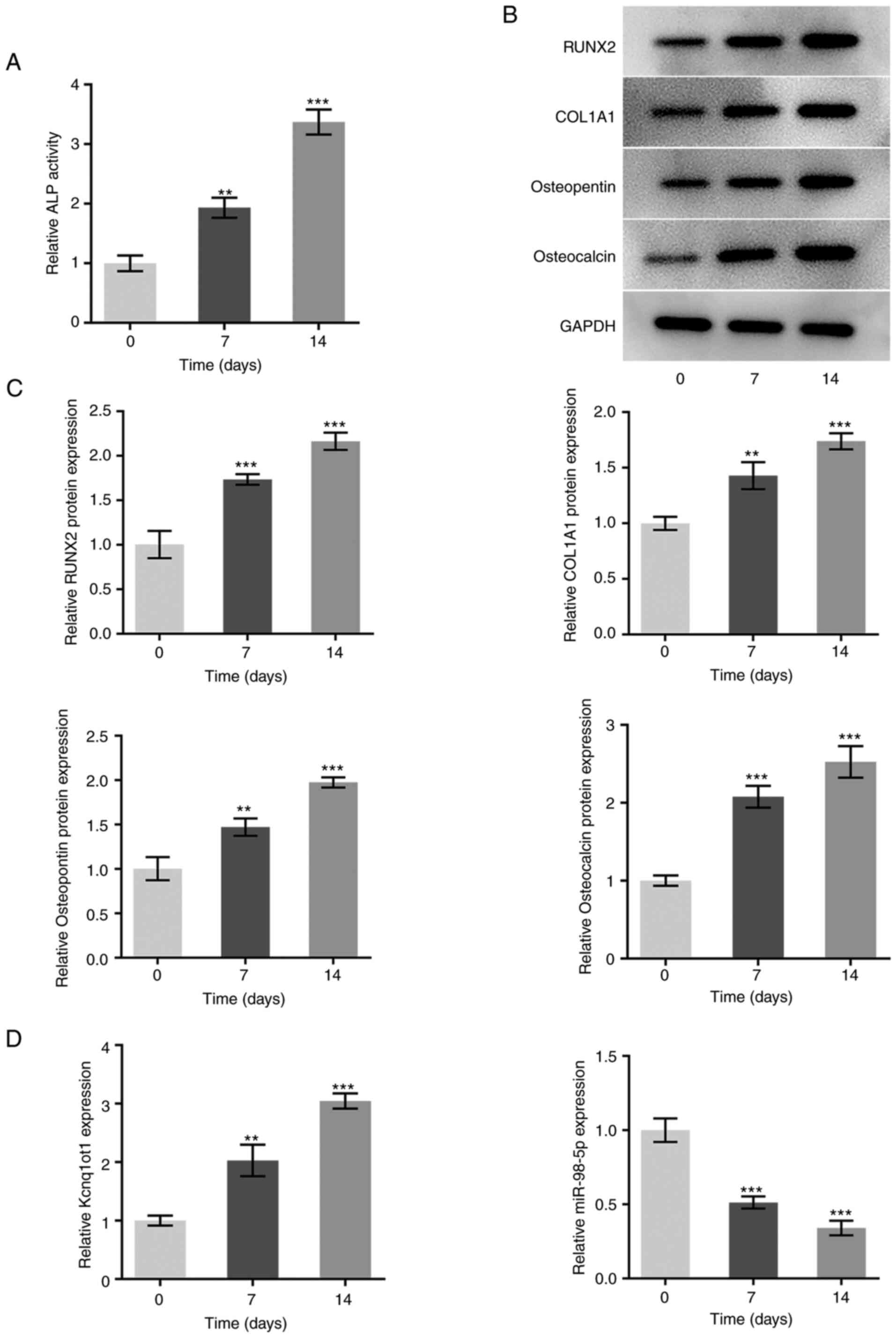
A time-dependent increase in the relative activity
of ALP was observed during the incubation of MC3T3-E1 osteoblasts
in osteogenesis-inducing medium, indicating the occurrence of
osteogenic differentiation (Fig.
1A). The protein expression levels of osteogenic
differentiation-associated proteins, RUNX2, COL1A1, osteopontin and
osteocalcin, were also increased in a time-dependent manner
following incubation of MC3T3-E1 cells in osteogenesis-inducing
medium (Fig. 1B and C). The expression of Kcnq1ot1 was
elevated, and that of miR-98-5p was decreased as the duration of
osteogenesis induction increased (Fig.
1D). These results suggested that Kcnq1ot1 expression increased
and miR-98-5p expression decreased during osteogenic
differentiation.
Kcnq1ot1 targets and inhibits
miR-98-5p
According to starBase, Kcnq1ot1 was predicted to
bind to miR-98-5p (Fig. 2A). A
dual luciferase reporter assay showed decreased luciferase activity
in MC3T3-E1 cells co-transfected with agomiR-98-5p and Kcnq1ot1-WT
compared with that in cells co-transfected with agomiR-98-5p and
Kcnq1ot1-MUT, which verified the binding between Kcnq1ot1 and
miR-98-5p (Fig. 2B). Furthermore,
following successful knockdown of Kcnq1ot1 (Fig. 2C), the expression of miR-98-5p was
significantly upregulated compared with the control group (Fig. 2D). These results suggest that
Kcnq1ot1 may target and inhibit miR-98-5p in MC3T3-E1 cells.
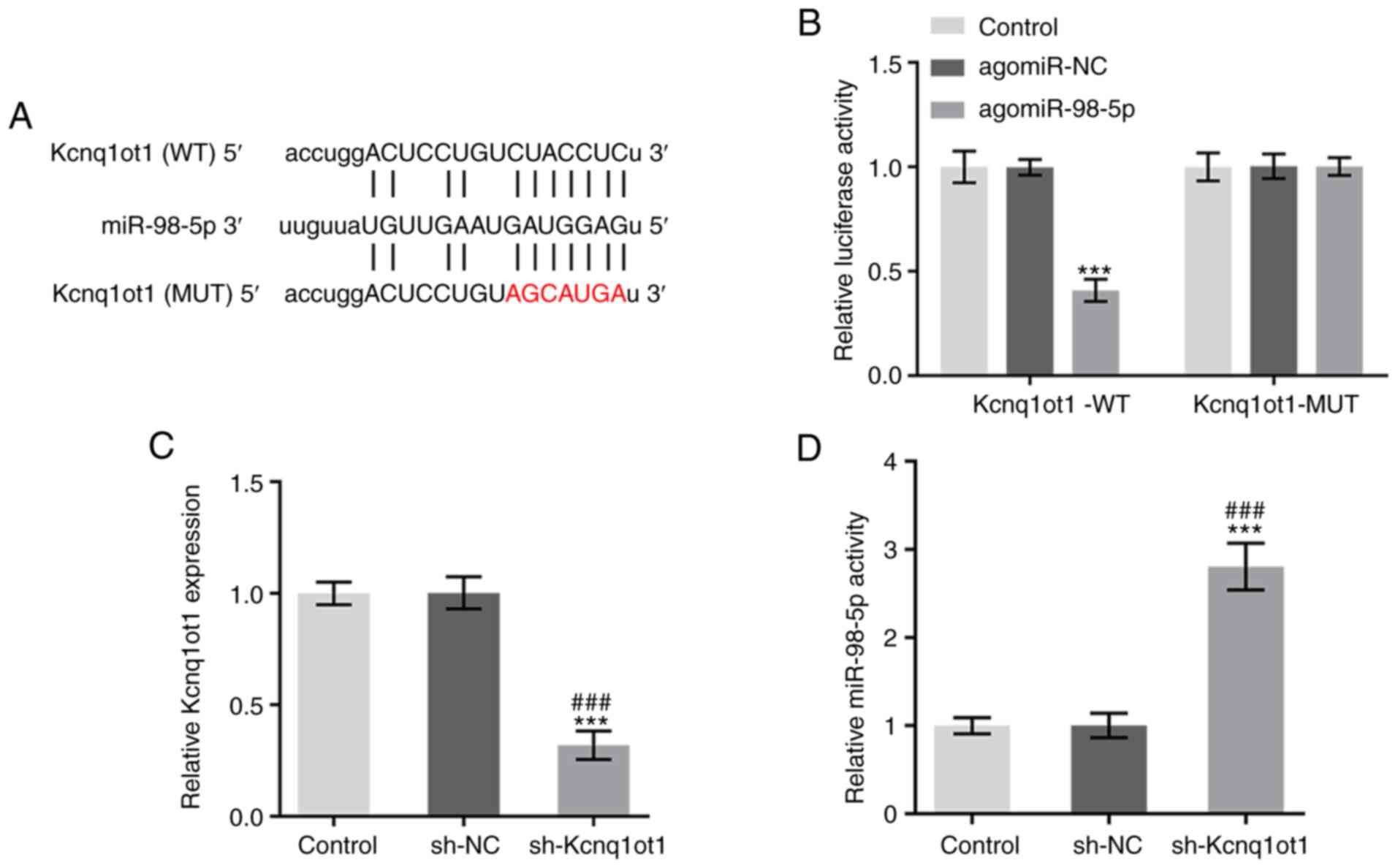 | Figure 2Kcnq1ot1 targets and inhibits
miR-98-5p. (A) StarBase prediction of the shared binding sites
between Kcnq1ot1 and miR-98-5p. (B) Relative luciferase activity in
MC3T3-E1 cells transfected with control, Kcnq1ot1-WT + agomiR-NC or
agomiR-98-5p, Kcnq1ot1-MUT + agomiR-NC or agomiR-98-5p, detected by
dual-luciferase reporter assay. Relative mRNA expression of (C)
Kcnq1ot1 and (D) miR-98-5p in MC3T3-E1 cells transfected with
control, sh-NC or sh-Kcnq1ot1, detected by RT-qPCR. Data are
expressed as mean ± SD. ***P<0.001 vs. control;
###P<0.001 vs. sh-NC. Kcnq1ot1, long non-coding RNA
KCNQ1 opposite strand/antisense transcript 1; miR, microRNA; WT,
wild-type; NC, negative control; MUT, mutant; sh, short
hairpin. |
Antagonizing miR-98-5p reverses the
inhibitory effect of Kcnq1ot1 knockdown on osteogenic
differentiation
To determine the role of the interaction between
miR-98-5p and Kcnq1ot1 in osteogenic differentiation, MC3T3-E1
cells were transfected with sh-Kcnq1ot1 or co-transfected with
sh-Kcnq1ot1 and antagomiR-98-5p. The knockdown effect of
antagomiR-98-5p was detected by RT-qPCR (Fig. 3A). miR-98-5p expression was
increased by sh-Kcnq1ot1; this was rescued following
co-transfection of sh-Kcnq1ot1 and antagomiR-98-5p (Fig. 3B). Kcnq1ot1 knockdown also
decreased ALP activity, whereas inhibition of miR-98-5p restored
ALP activity (Fig. 3C). Kcnq1ot1
knockdown significantly inhibited the formation of mineralized
nodules, which was rescued following co-transfection of sh-Kcnq1ot1
and antagomiR-98-5p in MC3T3-E1 cells (Fig. 3D). Kcnq1ot1 knockdown also
significantly inhibited the expression of RUNX2, COL1A1,
osteopontin and osteocalcin, whereas antagonizing miR-98-5p in the
presence of Kcnq1ot1 knockdown significantly upregulated expression
of these proteins (Fig. 3E). These
results indicated that antagonizing miR-98-5p may reverse the
inhibitory effect of Kcnq1ot1 knockdown on osteogenic
differentiation.
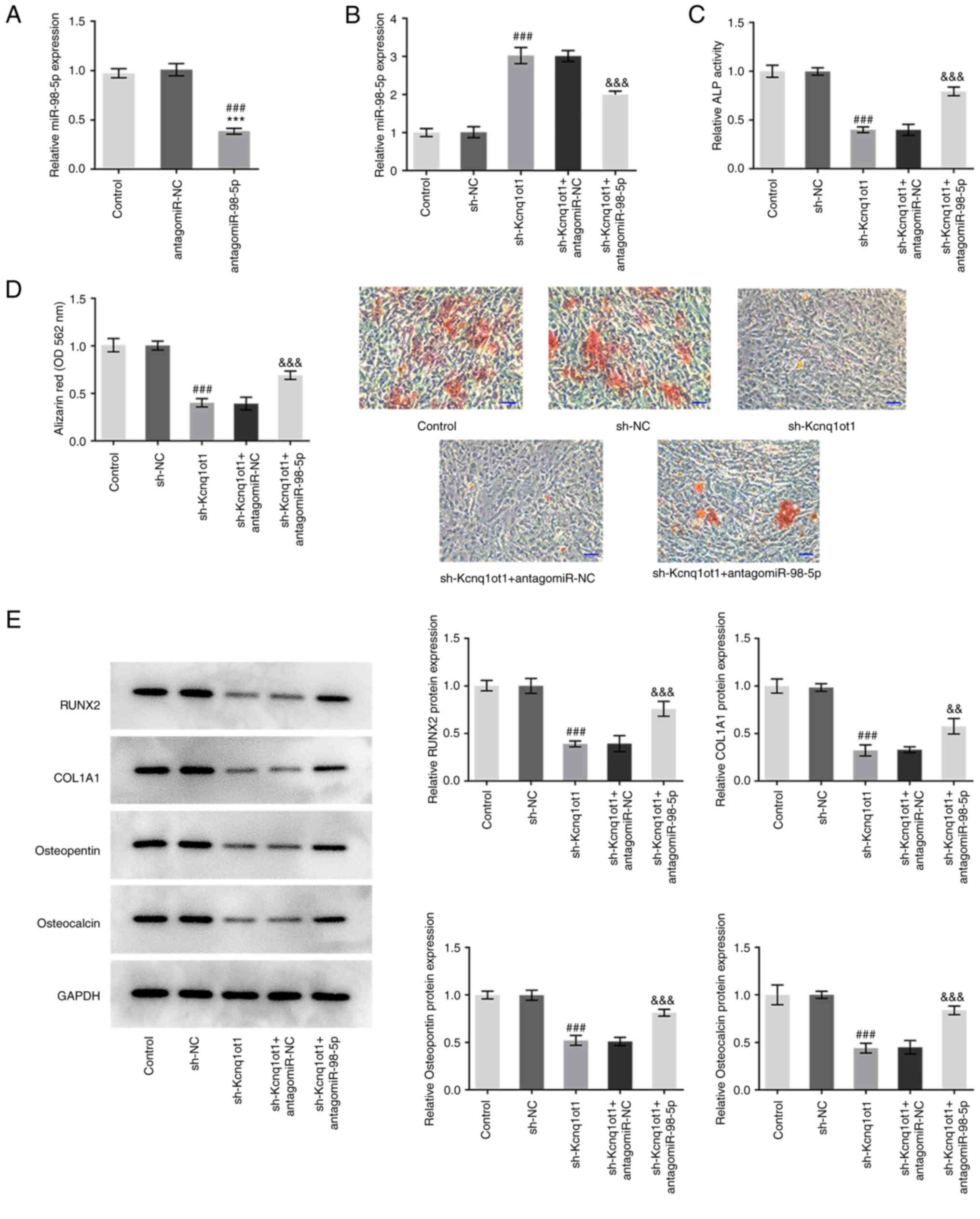 | Figure 3Antagonizing miR-98-5p reverses the
inhibitory effect of Kcnq1ot1 knockdown on osteogenic
differentiation. (A) Relative mRNA expression of miR-98-5p in
MC3T3-E1 cells was detected by reverse transcription-quantitative
PCR following transfection with (A) antagomiR-98-5p or (B) control,
sh-NC, sh-Kcnq1ot1, sh-Kcnq1ot1 + antagomiR-NC or sh-Kcnq1ot1 +
antagomiR-98-5p. (C) Relative ALP activity in MC3T3-E1 cells
transfected with control, sh-NC, sh-Kcnq1ot1, sh-Kcnq1ot1 +
antagomiR-NC or sh-Kcnq1ot1 + antagomiR-98-5p detected by ALP assay
kit. (D) Formation of mineralized nodules in MC3T3-E1 cells
transfected with control, sh-NC, sh-Kcnq1ot1, sh-Kcnq1ot1 +
antagomiR-NC or sh-Kcnq1ot1 + antagomiR-98-5p, detected by alizarin
red S staining assay. Scale bar, 50 µm. (E) Relative protein
expression of osteogenic differentiation-associated RUNX, COL1A1,
osteopontin and osteocalcin in MC3T3-E1 cells transfected with
control, sh-NC, sh-Kcnq1ot1, sh-Kcnq1ot1 + antagomiR-NC or
sh-Kcnq1ot1 + antagomiR-98-5p detected by western blotting. Data
are expressed as mean ± SD. ***P<0.001 vs. control;
###P<0.001 vs. antagomiR-NC or sh-NC;
&&P<0.01,
&&&P<0.001 vs. sh-Kcnq1ot1 +
antagomiR-NC. Kcnq1ot1, long non-coding RNA KCNQ1 opposite
strand/antisense transcript 1; miR, microRNA; ALP, alkaline
phosphatase; RUNX, Runt-related transcription factor 2; COL1A1,
collagen type I α 1; sh, short hairpin; NC, negative control; OD,
optical density. |
Kcnq1ot1 regulates Tbx5 expression via
miR-98-5p
TargetScan software was used to predict the binding
sites between Tbx5 and miR-98-5p (Fig.
4A). The transfection efficiency of pc-Kcnq1ot1 and
agomiR-98-5p was detected by RT-qPCR (Fig. 4B and C). A dual luciferase reporter assay
confirmed the results of the TargetScan prediction as significantly
decreased relative luciferase activity was observed in MC3T3-E1
cells co-transfected with Tbx5-WT and agomiR-98-5p. Furthermore,
MC3T3-E1 cells transfected with Tbx5-WT, agomiR-98-5p and
pc-Kcnq1ot1 exhibited increased luciferase activity compared with
the Tbx5-WT + agomiR-98-5p group (Fig.
4D). The results of RT-qPCR and western blotting both showed
that Tbx5 expression was significantly downregulated by
agomiR-98-5p but upregulated following Kcnq1ot1 overexpression
(Fig. 4E and F). These results indicated that Kcnq1ot1
may exert a regulatory effect on Tbx5 expression via modulating
miR-98-5p.
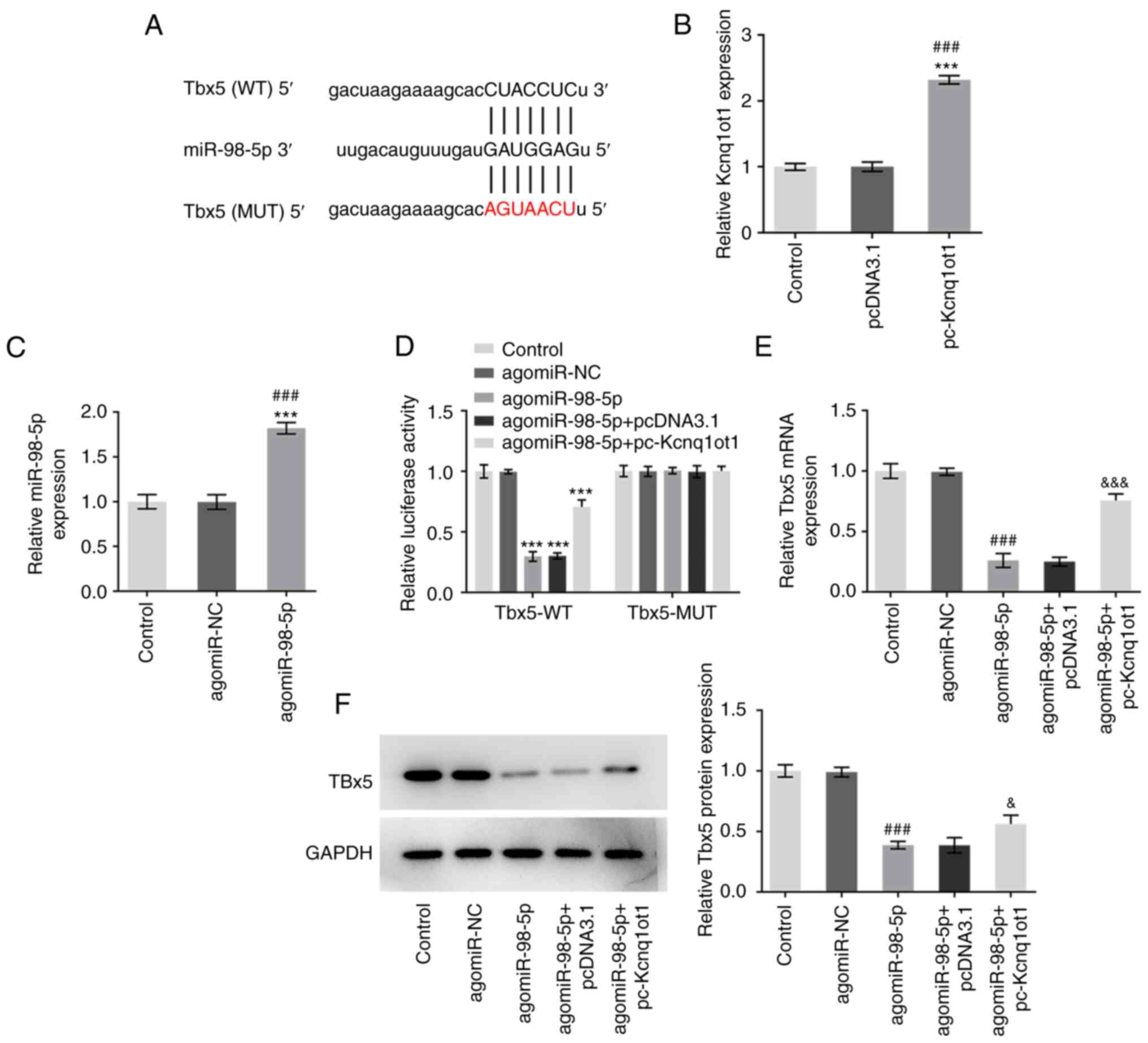 | Figure 4Kcnq1ot1 regulates Tbx5 expression via
miR-98-5p. (A) TargetScan prediction of the shared binding sites
between miR-98-5p and Tbx5. Expression of (B) Kcnq1ot1 and (C)
miR-98-5p was measured by RT-qPCR following transfection with
pc-Kcnq1ot1 and agomiR-98-5p. (D) Relative luciferase activity in
MC3T3-E1 cells transfected with control, Tbx5-WT + agomiR-NC,
Tbx5-WT + agomiR-98-5p, Tbx5-WT + agomiR-98-5p + pcDNA3.1, Tbx5-WT
+ agomiR-98-5p + pc-Kcnq1ot1, Tbx5-MUT + agomiR-NC, Tbx5-MUT +
agomiR-98-5p, Tbx5-MUT + agomiR-98-5p + pcDNA3.1, Tbx5-MUT +
agomiR-98-5p + pc-Kcnq1ot1 detected by dual-luciferase reporter
assay. Relative expression of Tbx5 in MC3T3-E1 cells transfected
with control, agomiR-NC, agomiR-98-5p, agomiR-98-5p + pcDNA3.1 or
agomiR-98-5p + pc-Kcnq1ot1, detected by (E) RT-qPCR and (F) western
blot analysis. Data are expressed as mean ± SD.
***P<0.001 vs. control; ###P<0.001 vs.
pcDNA3.1 or agomiR-NC; &P<0.05,
&&&P<0.001 vs. agomiR-98-5p + pcDNA3.1.
Kcnq1ot1, long non-coding RNA KCNQ1 opposite strand/antisense
transcript 1; miR, microRNA; sh, short hairpin; NC, negative
control; Tbx5, T-box transcription factor 5; RT-q, reverse
transcription-quantitative; WT, wild-type; MUT, mutant. |
miR-98-5p overexpression and Tbx5
knockdown reverse the promotive effect of Kcnq1ot1 overexpression
on osteogenic differentiation and mineralization
To determine the role of the interaction between
miR-98-5p and Kcnq1ot1, as well as that between Tbx5 and Kcnq1ot1,
in osteogenic differentiation and mineralization, agomiR-98-5p or
sh-Tbx5 were transfected into MC3T3-E1 cells overexpressing
Kcnq1ot1. The transfection efficiency of sh-Tbx5 was detected by
RT-qPCR (Fig. 5A). Tbx5 expression
was decreased in both the pc-Kcnq1ot1 + agomiR-98-5p and
pc-Kcnq1ot1 + sh-Tbx5 groups (Fig.
5B). Meanwhile, the relative increase in ALP activity following
Kcnq1ot1 overexpression was decreased following miR-98-5p
overexpression or Tbx5 knockdown (Fig.
5C). Kcnq1ot1 overexpression significantly increased
mineralization of MC3T3-E1 cells, and this was decreased following
transfection with agomiR-98-5p or sh-Tbx5 (Fig. 5D and E). These results suggested that
overexpressing miR-98-5p or knocking down Tbx5 may reverse the
promotive effect of Kcnq1ot1 overexpression on osteogenic
differentiation and mineralization.
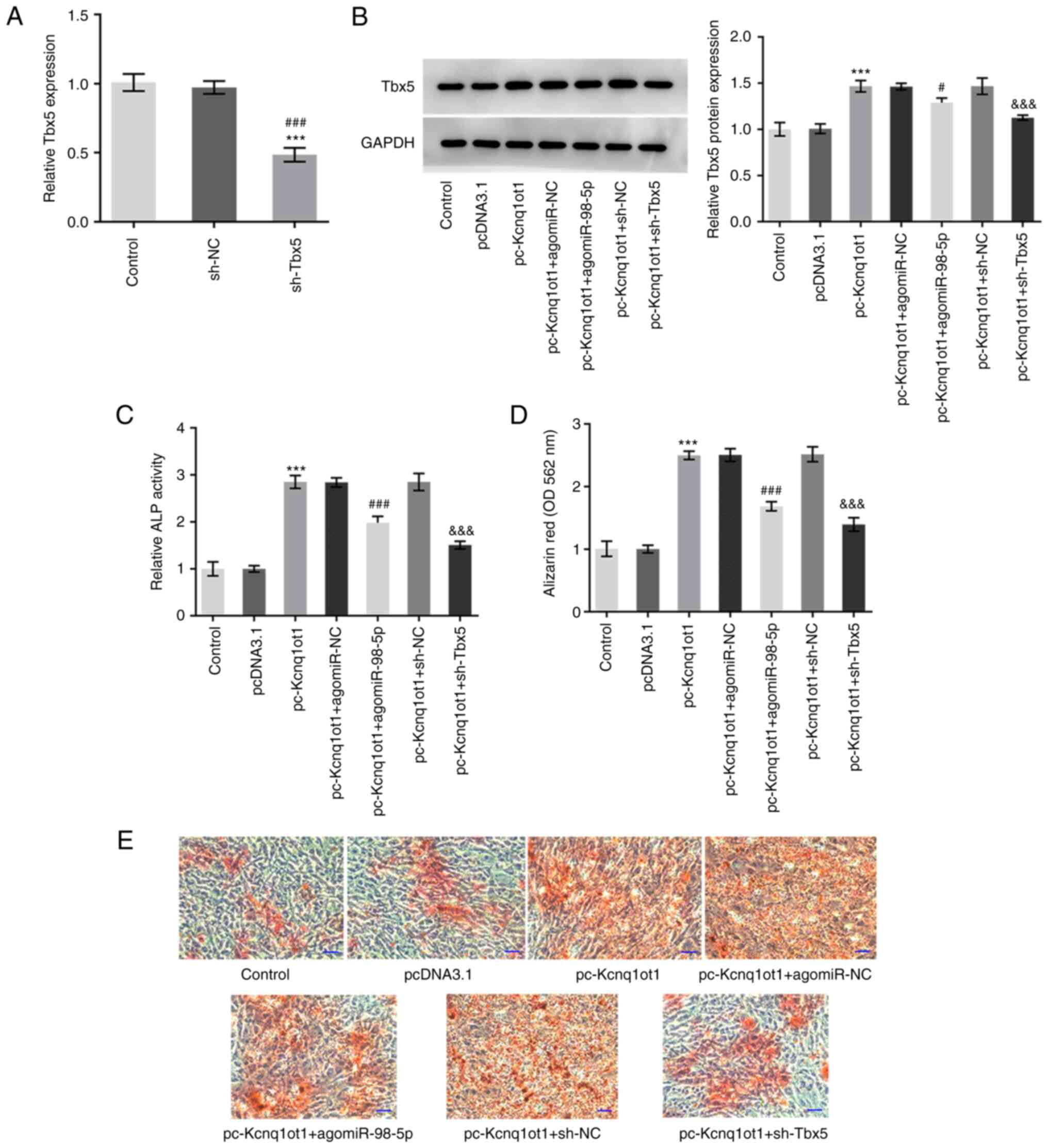 | Figure 5miR-98-5p overexpression and Tbx5
knockdown reverse the promotive effect of Kcnq1ot1 overexpression
on osteogenic differentiation and mineralization. (A) Relative mRNA
expression of Tbx5 in MC3T3-E1 cells was detected by reverse
transcription-quantitative PCR following transfection with sh-Tbx5.
(B) Relative protein expression of Tbx5 in MC3T3-E1 cells
transfected with control, pcDNA3.1, pc-Kcnq1ot1, pc-Kcnq1ot1 +
agomiR-NC, pc-Kcnq1ot1 + agomiR-98-5p, pc-Kcnq1ot1 + sh-NC or
pc-Kcnq1ot1 + sh-Tbx5 detected by western blotting. (C) Relative
ALP activity in MC3T3-E1 cells transfected with control, pcDNA3.1,
pc-Kcnq1ot1, pc-Kcnq1ot1 + agomiR-NC, pc-Kcnq1ot1 + agomiR-98-5p,
pc-Kcnq1ot1 + sh-NC or pc-Kcnq1ot1 + sh-Tbx5 detected by ALP assay
kit. (D) Formation of mineralized nodules in MC3T3-E1 cells
transfected with control, pcDNA3.1, pc-Kcnq1ot1, pc-Kcnq1ot1 +
agomiR-NC, pc-Kcnq1ot1 + agomiR-98-5p, pc-Kcnq1ot1 + sh-NC or
pc-Kcnq1ot1 + sh-Tbx5 (E) detected by alizarin red S staining
assay. Scale bar, 50 µm. Data are expressed as mean ± SD.
***P<0.001 vs. pcDNA3.1; #P<0.05,
###P<0.001 vs. sh-NC or pc-Kcnq1ot1 + agomiR-NC;
&&&P<0.001 vs. pc-Kcnq1ot1 + sh-NC.
Kcnq1ot1, long non-coding RNA KCNQ1 opposite strand/antisense
transcript 1; miR, microRNA; sh, short hairpin; NC, negative
control; Tbx5, T-box transcription factor 5; ALP, alkaline
phosphatase; OD, optical density. |
miR-98-5p overexpression and Tbx5
knockdown reverse the promotive effect of Kcnq1ot1 overexpression
on osteogenic differentiation-associated protein expression
The expression levels of osteogenic
differentiation-associated proteins, RUNX2, COL1A1, osteopontin and
osteocalcin, were detected following co-transfection of pc-Kcnq1ot1
with agomiR-98-5p or sh-Tbx5. The expression levels of
differentiation-associated proteins were increased by Kcnq1ot1
knockdown and inhibited by co-transfection of pc-Kcnq1ot1 and
agomiR-98-5p or pc-Kcnq1ot1 and sh-Tbx5 (Fig. 6). These findings suggested that
miR-98-5p overexpression and Tbx5 knockdown may reverse the
promotive effect of Kcnq1ot1 overexpression on osteogenic
differentiation-associated protein expression levels.
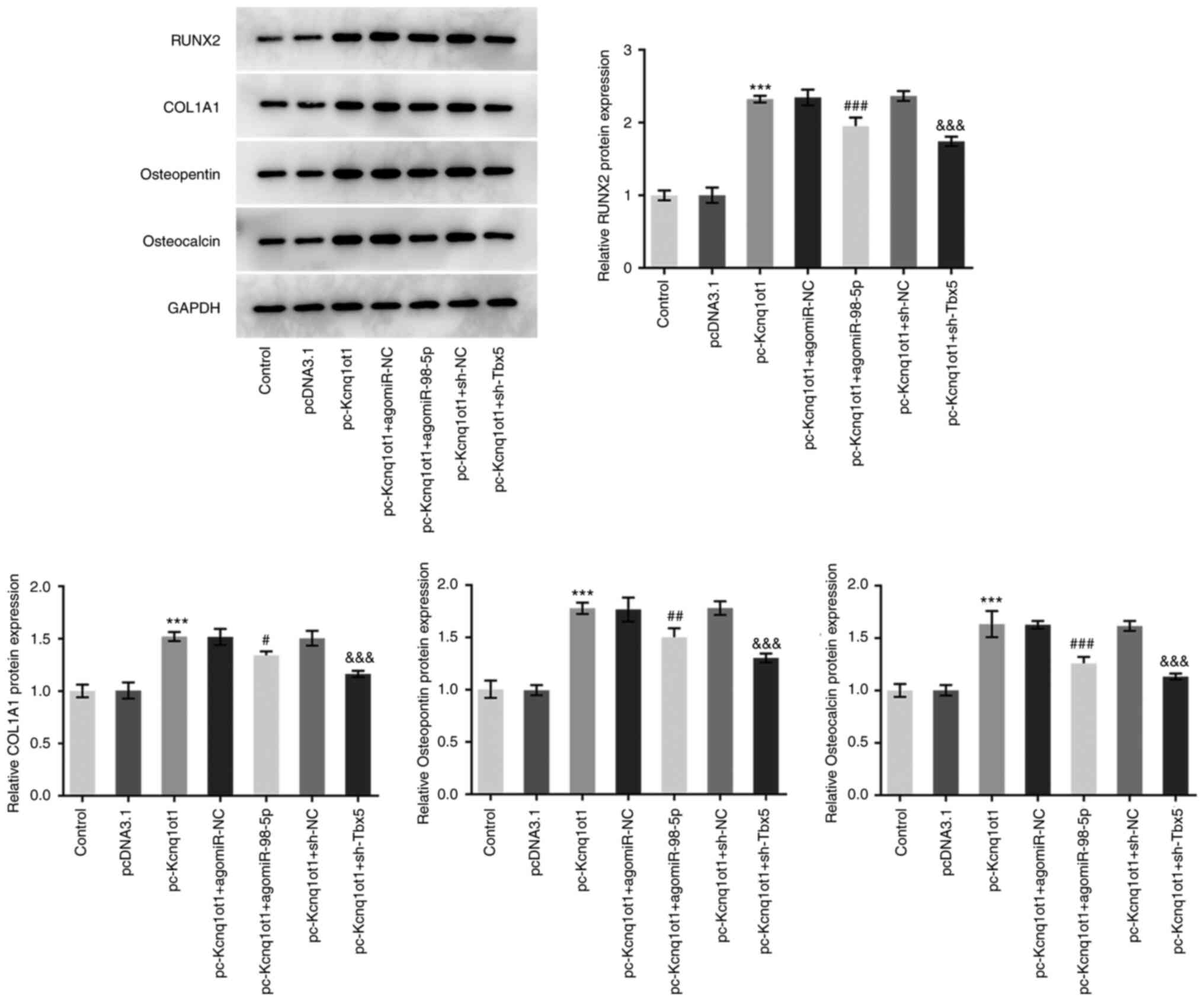 | Figure 6miR-98-5p overexpression and Tbx5
knockdown reverse the promotive effect of Kcnq1ot1 overexpression
on osteogenic differentiation-associated protein expression.
Relative protein expression of osteogenic
differentiation-associated RUNX2, COL1A1, osteopontin and
osteocalcin in MC3T3-E1 cells transfected with control, pcDNA3.1,
pc-Kcnq1ot1, pc-Kcnq1ot1 + agomiR-NC, pc-Kcnq1ot1 + agomiR-98-5p,
pc-Kcnq1ot1 + sh-NC or pc-Kcnq1ot1 + sh-Tbx5 detected by western
blotting. Data are expressed as mean ± SD. ***P<0.001
vs. pcDNA3.1; #P<0.05, ##P<0.01,
###P<0.001 vs. pc-Kcnq1ot1 + agomiR-NC;
&&&P<0.001 vs. pc-Kcnq1ot1 + sh-NC.
Kcnq1ot1, long non-coding RNA KCNQ1 opposite strand/antisense
transcript 1; miR, microRNA; sh, short hairpin; NC, negative
control; Tbx5, T-box transcription factor 5; RUNX, Runt-related
transcription factor 2; COL1A1, collagen type I α 1. |
Discussion
Osteoporosis is a metabolic bone disease that is
characterized by a severe decrease in bone density and mass
(22). Dysfunction of either
osteoblasts or osteoclasts can affect bone formation and
resorption, ultimately leading to metabolic bone disease (22,23).
In osteoporosis, back pain or body aches decrease quality of life,
whereas spinal deformities and fractures can be disabling, limiting
mobility and the ability to function independently (24). In addition, osteoporosis increases
the risk of lung infection and bedsores when patients are bedridden
for long periods of time. These not only seriously threaten quality
of life and survival rate of patients, but also pose a heavy
economic burden to individuals, families and society (25).
The involvement of miRNAs, lncRNAs and circular RNAs
in various types of disease, including osteoporosis, has been
reported by an increasing number of studies (26-28).
A recent study showed that adipogenesis and osteogenesis of tendon
stem cells are inhibited by Kcnq1ot1 knockdown, which exerts its
effect via indirect inhibition of the miR-138 target genes
peroxisome proliferator activator receptor γ and RUNX2(29). Kcnq1ot1 has also been shown to
positively regulate osteogenic differentiation of bone marrow
mesenchymal stem cells by sponging miR-214(30). The proliferative capacity of human
osteoblastic cell line is less than that of mouse MC3T3-E1 cells,
so human cell lines were not suitable for the present study. In
addition, induction of osteogenic differentiation in human cells is
harder than in mouse cell lines (31). Human and mouse genes are highly
homologous, thus MC3T3-E1 cell line was investigated in the present
study rather than human osteoblastic cells (32). In the present study, increased
expression of Kcnq1ot1 was observed in MC3T3-E1 cells. Kcnq1ot1
silencing significantly inhibited osteogenic differentiation and
mineralization in MC3T3-E1 cells, which was consistent with a
previous report (33). According
to starBase, Kcnq1ot1 shares binding sites with miR-98-5p. A dual
luciferase reporter assay was then performed, which verified the
binding between Kcnq1ot1 and miR-98-5p. Furthermore, the expression
of miR-98-5p was significantly upregulated in MC3T3-E1 cells
following Kcnq1ot1 knockdown. miR-98-5p overexpression inhibits
osteogenic differentiation and proliferation of MC3T3-E1
osteoblasts by targeting high mobility group AT-Hook2, thereby
obstructing bone regeneration (20). In agreement with the aforementioned
studies, miR-98-5p silencing rescued the inhibitory effect of
Kcnq1ot1 knockdown on osteogenic differentiation and
mineralization, suggesting that Kcnq1ot1 silencing may serve a
suppressive role in bone formation by upregulating miR-98-5p
expression.
The present study investigated the interaction
between Kcnq1ot1 and miR-98-5p and the mechanism of miR-98-5p in
osteogenic differentiation. According to a previous study,
miR-98-5p targets the transcription factor Tbx5 and obstructs the
transition of rat mesenchymal stem cells into cardiomyocytes
(19). Furthermore, TargetScan
predicted binding of miR-98-5p to Tbx5. Tbx5 has been reported to
promote the differentiation of 5-azacytidine-treated cardiac
fibroblasts into cardiomyocytes (34). Another study showed that miR-10-5p
impedes myocardial differentiation of bone marrow mesenchymal stem
cells via inhibition of Tbx5(35).
Therefore, it was hypothesized that Kcnq1ot1 may target miR-98-5p
and upregulate expression of Tbx5, thereby promoting
differentiation. In the present study, the dual luciferase reporter
assay confirmed the binding between miR-98-5p and Tbx5. The data
revealed that knockdown of miR-98-5p significantly decreased Tbx5
expression, which was rescued by Kcnq1ot1 overexpression. This
suggested that Kcnq1ot1 regulated Tbx5 expression via miR-98-5p. In
addition, less osteogenic differentiation and mineralization, as
well as decreased expression levels of RUNX2, COL1A1, osteopontin
and osteocalcin in MC3T3-E1 cells, were observed following Kcnq1ot1
overexpression or transfection with agomiR-98-5p or sh-Tbx5. These
results suggested that miR-98-5p overexpression or Tbx5 knockdown
may reverse the promotive effect of Kcnq1ot1 overexpression on the
osteogenic differentiation of MC3T3-E1 cells. The present results
also revealed that the increase in Tbx5 expression levels induced
by Kcnq1ot1 overexpression was significantly suppressed by sh-Tbx5.
However, the suppressive effect of agomiR-98-5p on the increased
Tbx5 level was marginal. It was hypothesized that other
unidentified specific pathways regulate the Kcnq1ot1/miR-98-5p/Tbx5
axis. This should be confirmed in vivo and in clinical
practice. The MC3T3-E1 cell line is a classical and common cell
model used to study osteogenic differentiation. Thus, MC3T3-E1 was
selected as a representative to investigate the role of Kcnq1ot1 in
bone formation and the underlying mechanism (36,37).
Human osteoblast cell lines should be used to verify the results.
Utilization of multiple cell lines may better reveal the mechanism
underlying the effect of Kcnq1ot1 on osteogenic differentiation.
However, the present study focused on the target and mechanism by
which osteogenic differentiation regulates osteoporosis in the
present study. Another limitation of the study was that the
expression of Kcnq1ot1 and miR-98-5p was not assessed at early
timepoints (such as day 1-3). The biological effects of
Kcnq1ot1/miR-98-5p in other pre-osteoblast cell lines and other
potential mechanisms should be investigated in future.
In conclusion, the present study showed that
Kcnq1ot1 serves a role in regulating osteogenic differentiation and
mineralization of MC3T3-E1 by modulating expression of
miR-98-5p/Tbx5. Kcnq1ot1 may be a potential effective therapeutic
molecular biomarker for treatment of osteoporosis to improve
patient quality of life.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by Basic Research
Project of Qinghai Provincial Science and Technology Department,
China (grant no. 2020-ZJ-755).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FW and FZhe designed the experiments and wrote the
manuscript. FW, FZha and FZhe performed the experiments and
analyzed the data. FZhe revised the manuscript and supervised the
experiments. FW and FZha confirm the authenticity of all the raw
data. All authors have read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Coughlan T and Dockery F: Osteoporosis and
fracture risk in older people. Clin Med (Lond). 14:187–191.
2014.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Lamichhane AP: Osteoporosis-an update.
JNMA J Nepal Med Assoc. 44:60–66. 2005.PubMed/NCBI
|
|
3
|
Compston J: Glucocorticoid-induced
osteoporosis: An update. Endocrine. 61:7–16. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Lane NE: Glucocorticoid-induced
osteoporosis: New insights into the pathophysiology and treatments.
Curr Osteoporos Rep. 17:1–7. 2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Stein E and Shane E: Secondary
osteoporosis. Endocrinol Metab Clin North Am. 32:115–134.
2003.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Gass M and Dawson-Hughes B: Preventing
osteoporosis-related fractures: An overview. Am J Med. 119 (Suppl
1):S3–S11. 2006.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Delaney MF: Strategies for the prevention
and treatment of osteoporosis during early postmenopause. Am J
Obstet Gynecol. 194 (2 Suppl):S12–S23. 2006.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Carmona RJ and Adachi JD: Calcium and
vitamin D for osteoporotic fracture prevention. Pol Arch Med Wewn.
117:441–442. 2007.PubMed/NCBI
|
|
9
|
Aspray TJ and Hill TR: Osteoporosis and
the ageing skeleton. Subcell Biochem. 91:453–476. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wang Y, Tao Y, Hyman ME, Li J and Chen Y:
Osteoporosis in china. Osteoporos Int. 20:1651–1662.
2009.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chen P, Li Z and Hu Y: Prevalence of
osteoporosis in China: A meta-analysis and systematic review. BMC
Public Health. 16(1039)2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zeng Q, Li N, Wang Q, Feng J, Sun D, Zhang
Q, Huang J, Wen Q, Hu R, Wang L, et al: The prevalence of
osteoporosis in China, a nationwide, multicenter DXA survey. J Bone
Miner Res. 34:1789–1797. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Shoback D, Rosen CJ, Black DM, Cheung AM,
Murad MH and Eastell R: Pharmacological management of osteoporosis
in postmenopausal women: An endocrine society guideline update. J
Clin Endocrinol Metab. 105(dgaa048)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Johnston CB and Dagar M: Osteoporosis in
older adults. Med Clin North Am. 104:873–884. 2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chen X and Yan GY: Novel human
lncRNA-disease association inference based on lncRNA expression
profiles. Bioinformatics. 29:2617–2624. 2013.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Khorkova O, Hsiao J and Wahlestedt C:
Basic biology and therapeutic implications of lncRNA. Adv Drug
Deliv Rev. 87:15–24. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Wang SZ, Jia J and Chen CH:
LncRNA-KCNQ1OT1: A potential target in exosomes derived From ADSCs
for the treatment of osteoporosis. Stem Cells Int.
19(7690006)2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wang JL, Wei X, Wang AG, Bai Y and Wu XJ:
Kcnq1ot1 regulates osteogenic differentiation of hBMSC by
miR-320a/Smad5 axis. Eur Rev Med Pharmacol Sci. 24:2843–2854.
2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Sun HH, Sun PF and Liu WY: MiR-98-5p
regulates myocardial differentiation of mesenchymal stem cells by
targeting Tbx5. Eur Rev Med Pharmacol Sci. 22:7841–7848.
2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zheng F, Wang F and Xu Z: MicroRNA-98-5p
prevents bone regeneration by targeting high mobility group AT-Hook
2. Exp Ther Med. 18:2660–2666. 2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang GL, Xia XL, Li XL, He FH and Li JL:
Identification and expression analysis of the MSP130-related-2 gene
from Hyriopsis cumingii. Genet Mol Res. 14:4903–4913.
2015.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Chen X, Wang Z, Duan N, Zhu G, Schwarz EM
and Xie C: Osteoblast-osteoclast interactions. Connect Tissue Res.
59:99–107. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Mazess RB: Fracture risk: A role for
compact bone. Calcif Tissue Int. 47:191–193. 1990.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Miller PD: Management of severe
osteoporosis. Expert Opin Pharmacother. 17:473–488. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Srivastava M and Deal C: Osteoporosis in
elderly: Prevention and treatment. Clin Geriatr Med. 18:529–555.
2002.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang Y, Yujiao W, Fang W, Linhui Y, Ziqi
G, Zhichen W, Zirui W and Shengwang W: The roles of miRNA, lncRNA
and circRNA in the development of osteoporosis. Biol Res.
53(40)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Jin D, Wu X, Yu H, Jiang L, Zhou P, Yao X,
Meng J, Wang L, Zhang M and Zhang Y: Systematic analysis of
lncRNAs, mRNAs, circRNAs and miRNAs in patients with postmenopausal
osteoporosis. Am J Transl Res. 10:1498–1510. 2018.PubMed/NCBI
|
|
28
|
Li Y, Li J, Chen L and Xu L: The roles of
long non-coding RNA in osteoporosis. Curr Stem Cell Res Ther.
15:639–645. 2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yu Y, Chen Y, Zhang X, Lu X, Hong J, Guo X
and Zhou D: Knockdown of lncRNA Kcnq1ot1 suppresses the adipogenic
and osteogenic differentiation of tendon stem cell via
downregulating miR-138 target genes PPARgamma and RUNX2. Cell
Cycle. 17:2374–2385. 2018.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Wang CG, Liao Z, Xiao H, Liu H, Hu YH,
Liao QD and Zhong D: LncRNA Kcnq1ot1 promoted BMP2 expression to
regulate osteogenic differentiation by sponging miRNA-214. Exp Mol
Pathol. 107:77–84. 2019.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Czekanska EM, Stoddart MJ, Richards RG and
Hayes JS: In search of an osteoblast cell model for in vitro
research. Eur Cell Mater. 24:1–17. 2012.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Prasad A, Kumar SS, Dessimoz C, Bleuler S,
Laule O, Hruz T, Gruissem W and Zimmermann P: Global regulatory
architecture of human, mouse and rat tissue transcriptomes. BMC
Genomics. 14(716)2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Gao X, Ge J, Li W, Zhou W and Xu L: LncRNA
Kcnq1ot1 promotes osteogenic differentiation to relieve osteolysis
via wnt/beta-catenin activation. Cell Biosci. 8(19)2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Jia Y, Chang Y, Guo Z and Li H:
Transcription factor Tbx5 promotes cardiomyogenic differentiation
of cardiac fibroblasts treated with 5-azacytidine. J Cell Biochem.
120:16503–16515. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Li M, Zhang YL, Huang H and Xiong Y:
MicroRNA-10-5p regulates differentiation of bone marrow mesenchymal
stem cells into cardiomyocytes by targeting Tbx5. Eur Rev Med
Pharmacol Sci. 23:479–485. 2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Li W, Zhang S, Liu J, Liu Y and Liang Q:
Vitamin K2 stimulates MC3T3-E1 osteoblast differentiation and
mineralization through autophagy induction. Mol Med Rep.
19:3676–3684. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Lu X, Lu J, Zhang L and Xu Y: Effect of
ANGPTL7 on proliferation and differentiation of MC3T3-E1 cells. Med
Sci Monit. 25:9524–9530. 2019.PubMed/NCBI View Article : Google Scholar
|