Introduction
Helicobacter pylori (H. pylori) is a
spiral-shaped, microaerophilic and Gram-negative bacillus that was
isolated by Marshall and Warrenin 1983(1). H. pylori infection has been
considered as an independent risk factor for gastric cancer for 27
years (2). Both laboratory and
clinical studies have confirmed that H. pylori is a key
factor in the occurrence of gastric cancer. The Kyoto consensus
published in 2015 reported that H. pylori infection is an
infectious disease. The earlier H. pylori is eradicated, the
lower the incidence of gastric cancer will be. The eradication
program should contain the most effective antibiotics available
(3). It has been reported that
eradication is more successful when antibiotics are selected based
on individual susceptibility, community antibiotic sensitivity
testing or antibiotic use and clinical prognosis data (3). However, the increased antibiotic
resistance rates have become a severe challenge for the clinical
eradication of H. pylori. The resistance rate to
tetracycline (TET), amoxicillin (AMX) and furazolidone (FUR) in the
majority of countries and regions in the world is relatively low
(<10%) (4-10).
However, it has been reported that the abuse of antibiotics in Iran
has led to a high antibiotic resistance rate of H. pylori to
TET (38.5%), AMX (27.1%) and FUR (23.9%) (11). As of June 2017, in all World Health
Organization regions, the primary and secondary resistance rates of
clarithromycin (CLA), metronidazole (MNZ) and levofloxacin (LEFX)
were ≥15%, except for the primary and secondary resistance rates of
CLA in America (10%; 95% CI, 4-16%) and Southeast Asia (10%; 95%
CI, 5-16%) and LEFX in Europe (11%; 95% CI, 9-13%), while the
global double resistance rate of CLA and MNZ was 19% (12). It is generally accepted that the
main detection method of the antibiotic resistance of H.
pylori is a drug sensitivity test based on the in vitro
culture of H. pylori (13).
Since H. pylori is a microaerophilic bacterium, its in
vitro culture is challenging and time-consuming. In addition,
numerous hospitals do not dispose the facilities and equipment to
carry out this test. A previous study has indicated that there are
several resistance mechanisms of H. pylori to antibiotics,
including enhanced oxidative free radical scavenging activity,
increased DNA repair enzyme activity, drug efflux pump expression
and activation, as well as resistant gene expression, which is the
focus of the present study (8).
The Vaculationg cytotoxin gene A(VacA) gene exists in almost
all H. pylori strains. However, only ~50% of the strains can
express VacA, an important virulence factor in H.
pylori production that causes vacuolation (13). The VacA gene is divided into
middle region types m1 and m2, intermediate region i1, i2 and i3
and signal sequence types s1a, s1b, s1c and s2(14). The association between VacA
subtypes and H. pylori resistance remains controversial
(15,16). The detection technology of
drug-resistant genes is simpler and faster than the drug
sensitivity in vitro culture-based test, and it can detect
the H. pylori genotype at the same time (17). Therefore, the present study
explored the value of antibiotic-resistant gene detection in H.
pylori and the association between genotype and antibiotic
resistance.
Patients and methods
Patients
Patients with H. pylori infection who were
subjected to gastroscopy were enrolled between December 2017 and
January 2020 in The First Affiliated Hospital of Zhejiang Chinese
Medical University (Zhejiang, China). A total of 133 H.
pylori-positive patients were enrolled in the present study,
with a mean age of 45.1 years (age range, 18-71 years). The
inclusion criteria were as follows: i) Age, 18-70 years old; ii)
pathological findings of H. pylori infection; and iii)
positive in [13C]-urea and [14C]-urea breath
tests. The exclusion criteria were as follows: i) Gastroscopy
contraindications; ii) diagnosis of severe cardiovascular diseases,
immune disorders, poorly controlled diabetes, hypertension or
recently planned subtotal gastrectomy; iii) inability to obtain
biopsies due to poor coagulation function or anticoagulant drugs;
iv) inability to complete gastroscopy due to lack of cooperation;
and v) refusal to participate in a clinical trial.
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Zhejiang Traditional
Chinese Medical University (approval no. 2017-KL-054-02; Hangzhou,
China). All patients provided written informed consent and general
information, such as sex and age. Gastric mucosa samples were
obtained from all patients (three samples per patient; each sample
was 0.4-0.6 mm2). One sample was sent for routine
pathological examination, one was cultured in vitro and
subjected to a drug sensitivity test (agar plate dilution method),
and one was sent for antibiotic resistance gene detection (PCR
reverse dot blot; a chip with mutation sites presented in Table I). Finally, the sensitivity of
H. pylori detection was compared between the two methods.
The mutation sites tested in the present study were as follows:
C556G, A562T and A562G in PBP1 for AMX; T87A, A91G, G91T and
G91A in gyrA for LEFX; 926-928 (AGA to TTC) and 926-927 (AG
to GT) in 16S rRNA for TET; A2142G and A2143G in 23S
rRNA for CLA; and G616A in rdxA for MNZ (Table I). The mutation rate of the related
gene sites was calculated in different antibiotic-resistant strains
and the antibiotic resistance rate was calculated in gene mutant
strains. Finally, the drug resistance rate was compared among
different VacA genotypes.
 | Table IA total of 12 gene mutation types
associated with antibiotic resistance. |
Table I
A total of 12 gene mutation types
associated with antibiotic resistance.
| Mutation type
Gene | Amoxicillin | Levofloxacin | Tetracycline | Clarithromycin | Metron idazole
rdxA |
|---|
| PBP1 | gyrA | 16S
rRNA | 23S
rRNA |
|---|
| Mutant site | 556 | 562 | | 87 | 91 | | | 926-928 | 926-927 | 2142 | 2143 | 616 |
| Mutation | C556G | A562T | A562G | T87A | A91G | G91T | G91A | AGA to TTC | AG to GT | A2142G | A2143G | G616A |
H. pylori culture and drug sensitivity
test H. pylori culture
Gastric mucosa tissue immersed in Bacterial
Preservation Fluid (Hangzhou ZhiYuan Medical Laboratory) was
aspirated using a sterile pipette. A total of 0.5 ml RIPA Lysis
Buffer (Hangzhou ZhiYuan Medical Laboratory) was to form tissue
homogenate. The tissue homogenate was then added to the culture
medium of Columbia blood agar (500 g; Thermo Fisher Scientific
Inc.). The medium was incubated in a microaerobic environment (5%
O2; 10% CO2; 85% N2; 37˚C) for 2-3
days. The suspected colonies were selected for hematoxylin-eosin
staining (6.0% hematoxylin for 5 min and 0.5% eosin 2 min at room
temperature) and were subjected to three enzyme identification
tests [urease (Suzhou Haibo Biotechnology Co., Ltd., 37˚C, 180
min), oxidase (Beijing Luqiao Technology Co., Ltd., room
temperature 5 min) and catalase (Suzhou Haibo Biotechnology Co.,
Ltd. room temperature 5 min; data not shown]. If the cultured
bacteria were identified as H. pylori, the result was
positive; otherwise, it was negative.
Drug sensitivity test of H.
pylori
The H. pylori colonies were selected in
sterile normal saline, shaken and mixed well. Next, the bacterial
suspension containing 0.5 MCF (150 million bacteria/milliliter) was
prepared. A total of 3 µl bacterial suspension was collected using
a pipette gun and added to the blood agar medium, with and without
the following antibiotics: CLA (100 mg x 10 tablets; National
Medical Products Administration), LEFX (100 mg x 10 capsules; China
Academy of Food and Drug Control), MNZ (100 mg x 10 tablets; China
Academy of Food and Drug control), AMX (0.25 g x 30 capsules; Dr.
Ehrenstorfer GmbH). Three gas incubators were purchased from Thermo
Fisher Scientific, Inc. (cat. no. 3427). The antibiotic
concentration was the drug-sensitive minimum inhibitory
concentration of the antibiotic: 1 clarithromycin, 2 levofloxacin,
2 amoxicillin, 2 furazolidone, 2 tetracycline and 8 µg/ml
metronidazole. The medium was incubated in a microaerobic
environment (5% O2; 10% CO2; 85%
N2; 37˚C) for 2 days. The growth state of the strain was
first observed in the culture medium without antibiotics. If it
grew normally, the drug sensitivity test result could be
interpreted; otherwise, the test needed to be repeated. Next, the
colony growth state was observed in the culture medium containing
antibiotics. If the colony growth was inhibited, H. pylori
was considered to be sensitive to the antibiotic; otherwise, H.
pylori was considered to be resistant to the antibiotic.
PCR reverse dot blot Preservation of
samples and sample processing
Gastric mucosa samples were preserved at -80˚C. For
genotyping and drug resistance gene mutation testing of H.
pylori, a nucleic acid extraction reagent was used [Nucleic
acid extraction kit, Yaneng Biosciences (Shenzhen) Co., Ltd., cat.
no. Yueshen Xiebei no. 20150098; model no. pathogen DNA
(centrifugal column type); specification, 25 copies/box]. The
concentration and purity of the template DNA prior to PCR was
determined using a nucleic acid quantitative analyzer (Multi-mode
Readers BioTek).
Preparation of amplification reaction
solution
The sequences of the gene primers used are listed in
Table II. The number of reaction
solutions required for each test was n+2, where n=number of samples
to be tested, and 2 equals one H. pylori positive quality
control sample and one H. pylori negative quality control
material. Reaction solutions I, II and III were removed from the
reagent box, melted and mixed. The amplification reaction solution
was prepared according to the manufacturer's instructions. After
the mixture was prepared, it was evenly mixed and centrifuged at
447.2 x g for 10 sec at 4˚C.
| Table IISequences of the forward primers. |
Table II
Sequences of the forward primers.
| A, Wild-type |
|---|
| Primer (site) | Number | Sequence,
5'→3' |
|---|
| 16S rRNA
(926~928) | 16S W |
CGAAGATACACGAAGAAC |
| 23S rRNA
(2142/2143) | 23S W |
ACGGAAAGACCCCGTG |
| gyrA (87) | 87 W |
GGCGATAATGCGGTTT |
| gyrA (91) | 91 W |
TTTATGATGCACTAGTGAG |
| PBP1(556) | 556 W |
AACCGGGACTTCCAATA |
| PBP1(562) | 562W |
AATGTGGATGCTTGGTTCA |
| rdxA (565) | 565W |
ATTGGGTAAGAGGGTG |
| rdxA (616) | 616W |
AAGTTGATGCAATTACTTG |
| B, Mutant |
| Primer (site) | Number | Sequence,
5'→3' |
| 16S rRNA
(926~928) | 16S TTC |
TCGATTCTACACGAAGAA |
| | 16S GTA |
ATTCGAGTATACACGAAG |
| | 16S TGA |
TCGATGATACACGAAGA |
| 23S rRNA
(2142) | A2142C | GACGGCAAGACCC |
| | A2142G | GACGGGAAGACCC |
| 23S rRNA
(2143) | A2143G |
AGACGGAGAGACCCC |
| gyrA (87) | 87K |
GCGATAARGCGGTTT |
| gyrA (91) | 91G |
ATGGTGCGYTAGTGAGA |
| | 91Y |
TTATTATGCGYTAGTGAG |
| | 91N |
TTATAATGCGCTAGTGAG |
| PBP1(556) | 556S |
GTAAAAGCGGRACTTCT |
| PBP1(562) | 562Y |
AACAACTATATTGATGCTTG |
| | 562D |
ACTTCTAACAACGATATTG |
| rdxA (565) | 565T |
GCTTTGTGTAAGAGGGT |
| rdxA (616) | 616A |
CAAAAGTTGATACAATTACTT |
| β-globin | IC |
CCTCTTATCTTCCTCCCAC |
Sample addition
The sample DNA was tested (three times per sample; 4
µl each) and added into three different amplification reaction
solutions. Quality control samples (DNA of known dose) were also
included (three times per sample; 4 µl each) and added into three
different amplification reaction solutions for PCR
amplification.
PCR amplification
PCR was performed using the H. pylori
Genotypes and drug resistance mutations detecting kit (PCR-reverse
dot blot; Hangzhou Qianji Biosciences Co., Ltd. China). The PCR
parameters are presented in Table
III. The temperature rise and fall rate was 3.0˚C/sec.
| Table IIIParameters for PCR amplification. |
Table III
Parameters for PCR amplification.
| Step no. | Step name | Temperature,
˚C | Time | Cycle number |
|---|
| 1 | Uracil-DNA
glycosylase enzyme reaction | 50 | 10 min | 1 |
| 2 | Initial
denaturation | 95 | 10 min | 1 |
| 3 | Denaturation | 95 | 30 sec | 45 |
| 4 | Annealing | 56 | 30 sec | |
| 5 | Extension | 72 | 30 sec | |
| 6 | Final
extension | 72 | 5 min | 1 |
Hybridization
Film strips marked with the sample number were
placed in a plastic centrifuge tube (15 ml), and 7 ml liquid A was
added. Next, three PCR products corresponding to the sample number
were added, and the tube cover was tightened. Next, the centrifuge
tube was heated in a boiling water bath for 10 min. It was then
removed, the cover was tightened, and the tube was placed into the
hybridization instrument (Combi-H12 FINEPCR) at 48˚C for 1.5 h. A
total of 45 ml liquid B was added to a 50 ml plastic centrifuge
tube, which was then preheated at 48˚C in the hybridization
instrument.
Membrane washing
The membrane strips were removed and transferred to
a 50 ml tube containing preheated liquid B, followed by gentle
washing at 48˚C for 15 min.
Color rendering
The formula of the incubation solution was as
follows: Solution A:Peroxidase=2,000:1. The membrane was washed
twice with liquid A at room temperature for 5 min each time. It was
then washed with liquid C at room temperature for 1 min, and the
color developing solution was prepared. The membrane strip was then
immersed in the color developing solution at room temperature for
10 min and rinsed with pure water twice to observe the results.
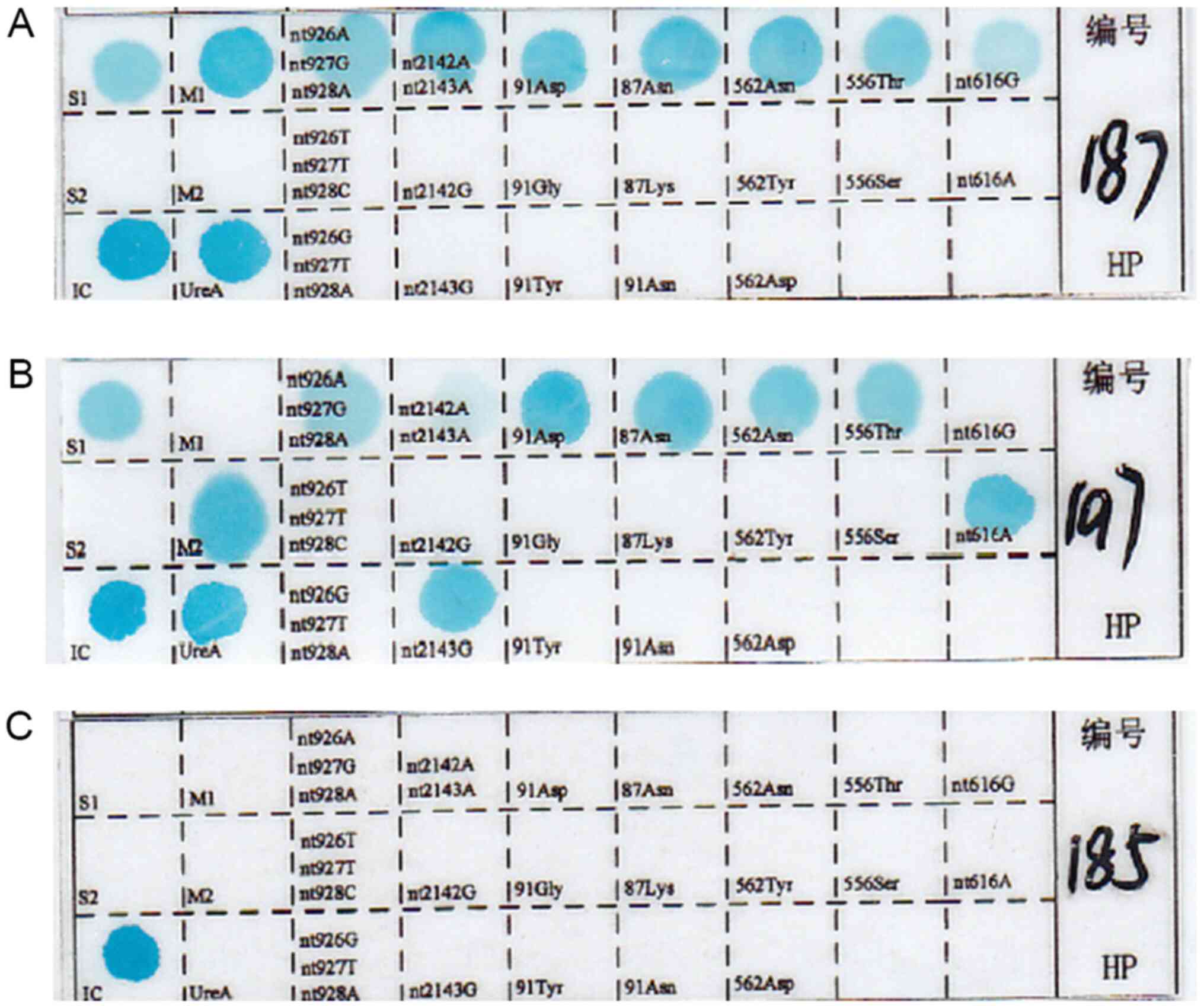
Subsequently, mutation points turned blue (Fig. 1).
Statistical analysis
Fisher's exact test was used to compare two groups
of classification data. The χ2 test was used for
comparison of multiple sample rates. SPSS v17.0 (SPSS, Inc.)
statistical software package was used for statistical analysis.
P<0.05 was considered to indicate a statistically significant
difference.
Results
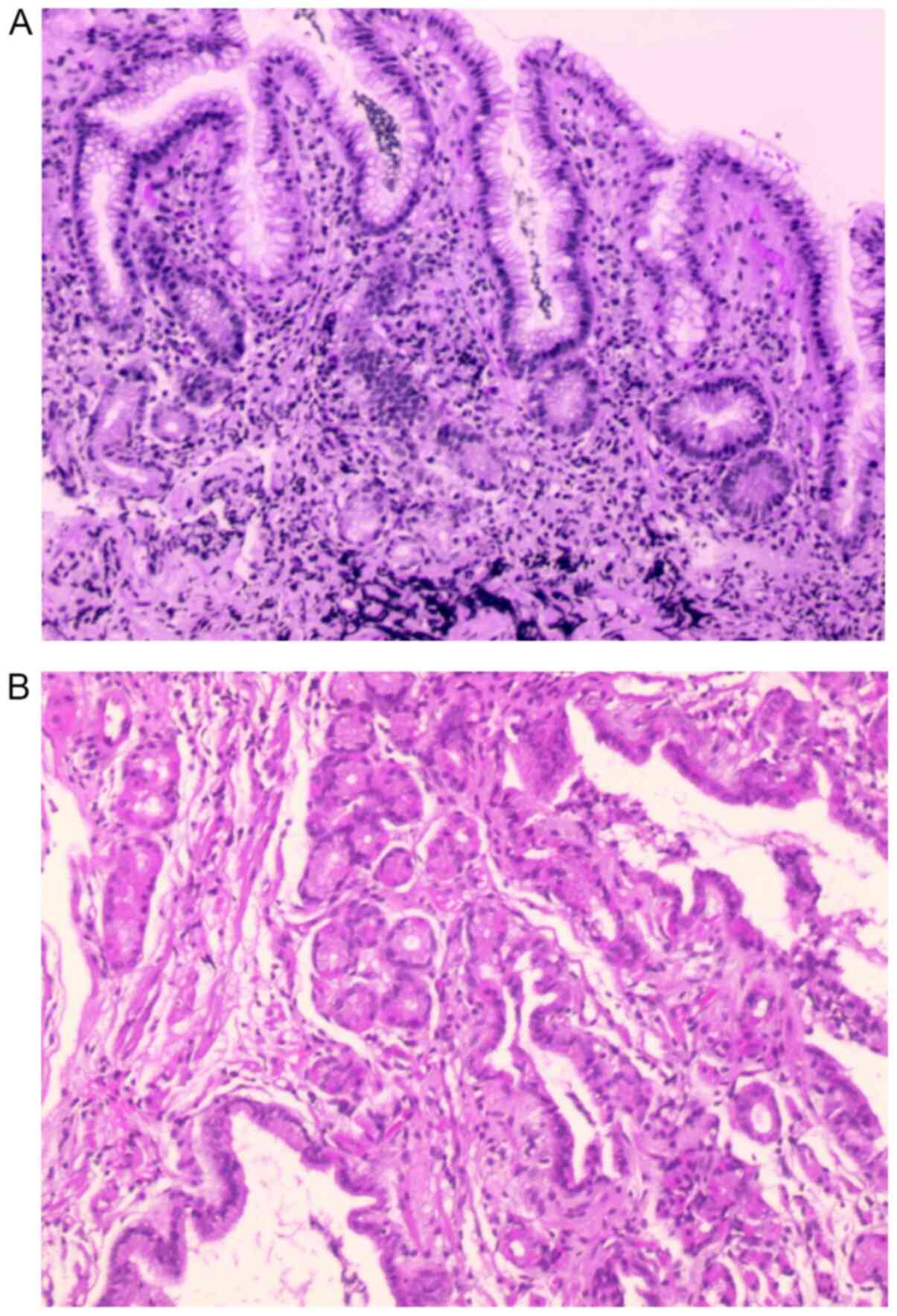
General patient information
The male:female ratio of the enrolled patients was
1.17:1 (P>0.05). A total of 111 strains were successfully
cultured in vitro, 109 strains were identified by
pathologists (Fig. 2), while 108
strains were detected using PCR reverse dot blot. No significant
difference in the sensitivity of H. pylori detection was
identified between the three methods (agar plate dilution method,
83.5%; PCR, 81.2%; pathology, 82.0%; P=0.89). A total of 96
antibiotic-resistant strains were confirmed by both the agar plate
dilution method and PCR reverse dot blot. According to genotyping,
all strains were VacA-positive and type s1, including 44
cases that were type s1m1 and 69 that were type s1m2. The agar
plate dilution method revealed that the resistance rate of MNZ was
90.1% (100/111), followed by 35.1% (39/111) for LEFX and 32.4%
(36/111) for CLA. No strains were resistant to AMX, FUR and TET. A
total of 34 strains were resistant to both CLA and MNZ, 37 strains
were resistant to both LEFX and MNZ, and 23 strains were resistant
to CLA, LEFX and MNZ.
Association analysis between mutations
of different gene loci and drug resistance
The results of the agar plate dilution method and
PCR reverse dot blot were not entirely consistent; 32.1% of strains
that, according to the agar plate dilution method, were
MNZ-resistant exhibited an rdxA G616A mutation. In addition,
numerous LEFX-related resistance gene sites were identified in
gyrA. Among them, the T87A mutation accounted for 32.3%, the
G91A mutation for 22.6%, the A91G mutation rate for 12.9% and the
G91T mutation for 6.5%. The main mutation in CLA-resistant strains
was 23S rRNA A2143G, accounting for 81.8%, with the 23S
rRNA A2142G mutation accounting for 0.0%. Neither AMX-resistant
nor TET-resistant strains were identified in vitro (Table IV).
| Table IVProportion of gene mutations in
drug-resistant heliobacter pylori strains shown by agar plate
dilution. |
Table IV
Proportion of gene mutations in
drug-resistant heliobacter pylori strains shown by agar plate
dilution.
| Antibiotic | Resistant-strain
number | Mutant sites | Mutant strain
number | Mutant rate, % |
|---|
| MNZ | 81 | G616A | 26 | 32.1 |
| LEFX | 31 | T87A | 10 | 32.3 |
| | | A91G | 4 | 12.9 |
| | | G91T | 2 | 6.5 |
| | | G91A | 7 | 22.6 |
| CLA | 33 | A2143G | 27 | 81.8 |
| | | A2142G | 0 | 0.0 |
| AMX | 0 | C556G | 0 | - |
| | | A562T | 0 | - |
| | | A562G | 0 | - |
| TET | 0 | 926~928
(AGA-TTC) | 0 | - |
| | | 926~927
(AG-GT) | 0 | - |
Among all the gene sites studied, rdxA G616A
had the highest accuracy in predicting H. pylori resistance
(81.8%), followed by gyrA G91T (66.7%), gyrA G91A
(54.5%), 23 S rRNA A2143G (49.5%), gyrA T87A (45.5%),
gyrA A91G (33.3%) and 23S rRNA A2142G (0.0%), while
no drug resistance was identified in the strains with
AMX-(PBP1 C556G, PBP1 A562T and PBP1 A562G)
and TET-resistance-related gene mutations [926-927 (AG to GT) and
926-928 (AGA-TTC) in 16S rRNA], according to the in
vitro drug sensitivity test (Table
V).
| Table VProportion of resistant strains in
vitro in strains with single gene loci mutation. |
Table V
Proportion of resistant strains in
vitro in strains with single gene loci mutation.
| Mutant site | Mutant-strain
number | Resistant-stain
number | Resistance rate,
% |
|---|
| G616A | 33 | 27 | 81.8 |
| T87A | 22 | 10 | 45.5 |
| A91G | 12 | 4 | 33.3 |
| G91T | 3 | 2 | 66.7 |
| G91A | 11 | 6 | 54.5 |
| A2143G | 53 | 26 | 49.1 |
| A2142G | 1 | 0 | 0 |
| C556G | 10 | 0 | 0 |
| A562T | 1 | 0 | - |
| A562G | 0 | 0 | - |
| 926~928
(AGA-TTC) | 0 | 0 | 0 |
| 926~927
(AG-GT) | 0 | 0 | 0 |
Among the genes, 81.8% of strains with a rdxA
mutation were revealed to be resistant to MNZ, while 47.8% of
strains with a gyrA mutation and 49.1% strains with a 23S
rRNA mutation were indicated to be resistant to LEFX and CLA,
respectively (Table VI).
| Table VIDrug resistance ratio in different
resistance gene mutant strains. |
Table VI
Drug resistance ratio in different
resistance gene mutant strains.
| Mutant gene | Mutant-strain
number | Resistant-stain
number | Positive rate,
% |
|---|
| rdxA | 33 | 27 | 81.8 |
| gyrA | 46 | 22 | 47.8 |
| 23S
rRNA | 53 | 26 | 49.1 |
| PBP1 | 13 | 0 | 0 |
| 16S
rRNA | 0 | 0 | - |
Drug resistance analysis of different
VacA genotypes
All 108 patients with positive H. pylori
genes presented the VacA s1 type; there were no VacA
s2 type patients. Out of those, 39 (36.1%) had VacA s1m1
strains and 69 (63.9%) VacA s1m2 strains. As indicated using
Fisher's exact test and χ2 analysis, there was no
significant association between genotypes and overall drug
resistance rate, multidrug resistance rate, CLA, LEFX or MNZ
resistance rate (P>0.05; Table
VII, Table VIII, Table IX, Table X and Table XI).
| Table VIIAssociation between Vac A
genotype and antibiotic resistance. |
Table VII
Association between Vac A
genotype and antibiotic resistance.
| Genotype | Resistant
strains | Sensitive
strains | Total |
|---|
| s1m1 | 32 | 2 | 34 |
| s1m2 | 53 | 5 | 58 |
| Total | 85 | 7 | 92 |
| Table VIIIAssociation between Vac A
genotype and multi-antibiotic resistance. |
Table VIII
Association between Vac A
genotype and multi-antibiotic resistance.
| Genotype | Single-antibiotic
resistant strainsa | Double-antibiotic
resistant strains | Triple-antibiotic
resistant strains | Multi-antibiotic
resistant strainsb | Total |
|---|
| s1m1 | 16 | 7 | 9 | 16 | 32 |
| s1m2 | 29 | 13 | 11 | 24 | 53 |
| Total | 45 | 20 | 20 | 40 | 85 |
| Table IXAssociation between Vac A
genotype and CLA resistance. |
Table IX
Association between Vac A
genotype and CLA resistance.
| Genotype | CLA-resistant
strains | CLA-sensitive
strains | Total |
|---|
| s1m1 | 14 | 20 | 34 |
| s1m2 | 19 | 39 | 58 |
| total | 33 | 59 | 92 |
| Table XAssociation between Vac A
genotype and MNZ resistance. |
Table X
Association between Vac A
genotype and MNZ resistance.
| Genotype | MNZ-resistant
strains | MNZ-sensitive
strains | Total |
|---|
| s1m1 | 30 | 4 | 34 |
| s1m2 | 51 | 7 | 58 |
| total | 81 | 11 | 92 |
| Table XIAssociation between Vac A
genotype and LEFX resistance. |
Table XI
Association between Vac A
genotype and LEFX resistance.
| Genotype | LEFX-resistant
strains | LEFX-sensitive
strains | Total |
|---|
| s1m1 | 13 | 21 | 34 |
| s1m2 | 18 | 40 | 58 |
| total | 31 | 61 | 92 |
Discussion
There are several mechanisms of H. pylori
antibiotic resistance, and they vary among different antibiotics
(8). The mechanism underlying CLA
resistance involves reverse binding to the domain V of the 23S
rRNA gene in the ribosomal subunit 50S in bacteria,
thereby interfering with protein synthesis and promoting the early
release of peptide tRNA from the recipient site, thus preventing
the synthesis of the H. pylori peptide chain extension
protein (18). Therefore, a
mutation in domain V of the 23S rRNA gene may decrease the
affinity between the ribosome and drug, leading to the lack of
binding between CLA and 50S ribosome subunit. If the
mutation cannot affect protein synthesis, H. pylori
resistance occurs. In addition, if the mutation cannot entirely
block the affinity between ribosome and antibiotics, sensitivity to
antibiotics can still occur in vitro. Among the point
mutations, certain are clinically more important than others. Seo
et al (19) reported that
19.5, 0.9, 0.2 and 0.7% patients presented A2143G, A2142G, A2142C
and both A2143G and A2142G mutations, respectively. The
CLA-resistant group was treated with proton-pump inhibitor (PPI)
twice a day, AMX (1,000 mg) twice a day and MNZ (500 mg) three
times a day for 7 days (PAM). Groups with other mutations were
treated with PPI twice a day, AMX (1,000 mg) twice a day and CLA
(500 mg) twice a day for 7 days (PAC). As revealed by ITT
(Intention-To-Treat (ITT) and PP(Per-Protocol)analysis, there was a
significant difference between the two groups [PAM vs. PAC, 55.4
vs. 74.3%, P=0.001 (ITT); 66.2 vs. 88.4%, P=0.0001) (PP)] (16). Similarly, Ong et al
(20) treated patients infected
with H. pylori with no CLA mutation points in 23S
rRNA with concomitant (1,000 mg AMX; 500 mg CLA; 500 mg MNZ and
30 mg lansoprazole twice a day for 14 days) or tailored (1,000 mg
AMX; 500 mg CLA and 30 mg lansoprazole twice a day for 14 days)
therapy. There was no significant difference in the eradication
rates between the groups in both point mutation-negative subjects
(91.7 vs. 87.3%; P=0.154). These studies demonstrated that the
detection of CLA mutation points in 23S rRNA could be
efficient for screening of CLA resistant strains in regions where
the CLA resistance rate is >15%. In the present study, the CLA
resistance rate was as high as 32.4%, and the mutation rate of
A2143G and A2142G in 23S rRNA was 81.8 and 0% (Table IV), respectively. The sensitivity
of A2142G and A2143G mutation in 23S rRNA was 81.8%,
rendering them ideal mutations for the detection of CLA resistance
in H. pylori in China. Adding bismuth to standard triple
therapy with CLA may eradicate 77.6% resistant strains with A2143G
or A2142G mutation (21).
LEFX interferes with bacterial DNA synthesis by
inhibiting DNA gyrase and topoisomerase IV (8). The gyrA gene encodes DNA
gyrase, so its mutations can lead to the resistance of H.
pylori to LEFX (22). Liou
et al (23) used a modified
sequential therapy containing LEFX and high-dose esomeprazole (40
mg esomeprazole and 1,000 mg AMX for the first 5 days, followed by
40 mg esomeprazole, 250 mg LEFX and 500 mg MNZ for another 5 days;
all administered twice daily) in patients who did not respond to
first-line treatment. The eradication rates were 84.6% (11/13) and
95.1% (58/61) in patients with and without a gyrA mutation,
respectively (P=0.210), which indicated little value of gyrA
mutation testing. However, Papastergiou et al (24) used a 7-day, genotypic
resistance-guided triple H. pylori eradication therapy
comprising esomeprazole, AMX and either CLA (wild-type 23S
rRNA), LEFX (23S rRNA mutated/wild-type gyrA) or
rifabutin (mutated 23S rRNA/gyrA). The mutation sites
of CLA in Papastergiou et al (24) were A2146G, A2146C and A2147G in the
23S rRNA gene, while the mutation sites of LEFX were N87K,
D91N, D91G and D91Y in the gyrA gene. For patients who
received CLA-, LEFX- and rifabutin-containing triple therapy, the
respective eradication rates were 24/27, 20/20 and 2/4 by ITT and
24/24, 19/19 and 2/3 by PP analysis. The overall eradication rates
exhibited no significant difference between treatment-naïve and
-experienced patients (ITT, 87.5 vs. 94.7%, P=0.64; PP, 96.4 vs.
100%, P=1.00). Therefore, the unsatisfactory results of the
previous trial may be linked to the high resistance rate of MNZ.
The present results indicated that 74.2% of LEFX-resistant strains
had gene mutations. The mutation rates of T87A, G91A, A91G and G91T
in gyrA were 32.3, 22.6, 12.9 and 6.5%, respectively. The
resistance rate of loci 91 (36.1%) was similar to that in the study
by Palmitessa et al (25).
The coincidence rates of mutation sites, as determined by in
vitro drug sensitivity test, were as follows: rdxA G616A
(81.8%), gyrA G91T (66.7%), gyrA G91A (54.5%), 23 S rRNA A2143G
(49.5%), gyrA T87A (45.5%), gyrA A91G (33.3%), and 23 S rRNA A2142G
(0.0%). Since the resistance rate of gyrB is low (26), it was not detected in the present
study, based on the economic benefit ratio. Meta analysis showed
that second-line anti-H. pylori
levofloxacin/amoxicillin-based triple (10-day regimen) therapy was
significantly more effective than standard quadruple therapy
[PPI+bismuth+two antibiotics; odds ratio, 5.05; 95% CI, 2.74-9.31;
P<0.001; heterogeneity index (I2)=75%] (27). Simple molecular susceptibility
testing for CLA and LEFX may achieve a high H. pylori
eradication rate. If CLA or LEFX resistance does exist,
administration of probiotic microorganisms along with triple
therapy would be another treatment possibility (28).
In the present study, no case showed 926-928 and
926-927 loci mutations in 16S rRNA, a TET-related resistance
gene and 556 and 562 loci mutations in an AMX-related
drug-resistant gene, PBP1, occurred in 10 cases (C556G) and
3 case (A562T), respectively. However, TET- and AMX-resistant
strains were not revealed to be among these mutant strains via the
in vitro drug sensitivity test. At the same time,
AMX-resistant strains detected in the in vitro drug
sensitivity test identified no gene mutation. No FUR-resistant
strains were cultured in the present study. In a review, Zamani
et al (29) analyzed
studies on the resistance of H. pylori to FUR published
worldwide, suggesting that the resistance rate of H. pylori
to FUR in most regions was <5%, which may be associated with
A041G, A122G, C349A or G A78G, A112G, A335G, C156T and C165T
mutations in the oorD gene and G353A, A356G, C357T, C347T,
C347G and C346A mutations in the porD gene. The specificity
and sensitivity of several mutation sites mentioned in the review
by Zamani et al (29)
requires further investigation, as they were not detected in the
present study. Generally, the drug resistance rate of H.
pylori to AMX, TET and FUR is low, and thus no drug resistance
test is required prior to eradication if any of these three
antibiotics are used (30)
However, it should be noted that TET has been out of production for
many years in China.
In a study by Lee et al (31), which enrolled 53 H.
pylori-positive patients treated with MNZ-containing sequential
or quadruple therapy between 2011 and 2015, rdxA and
frxA mutations in patients with H. pylori were
analyzed via DNA sequencing. The results indicated that rdxA
mutations served a critical role in MNZ resistance and the outcomes
of eradication therapy. Apart from rdxA, frxA and
frxB are among the most frequently studied mutations;
however, previous studies have suggested that mutations in these
two genes have a synergistic effect with rdxA mutations in
causing MNZ resistance, but cannot cause it alone (32-34).
In theory, if the mutation rate of rdxA in resistant strains
is high enough, tailored therapy could achieve a high eradication
rate of H. pylori, according to the rdxA gene
mutation detection results. However, only 32.1% of the
MNZ-resistant strains had a rdxA mutation in G616A in the
present study, which was slightly higher than the rate observed in
a study in Pakistan (32.1 vs. 23.5%) (35). In addition, all strains (20/20)
that were resistant to both CLA and LEFX were also resistant to MNZ
in the present study. According to a study by Zhang et al
(36), the rdxA mutation
was identified in 90.7% (49/54) of Chinese children. Therefore,
rdxA mutation detection in G616A may not be a good strategy
to improve the eradication rate of H. pylori among adults in
high MNZ-resistant areas, while it may benefit Chinese
children.
As aforementioned, VacA is an important
virulence factor in H. pylori. The association between
VacA subtypes and H. pylori resistance remains
controversial. Karabiber et al (15) demonstrated that VacA
s1c-positive H. pylori was more likely to lead to CLA
resistance, while in MNZ-resistant strains VacA s1 and s1c
were more likely to lead to resistance. Wang et al (37) reported that the resistance rate of
MNZ in the VacA s1m1/m2 strains was lower than that in the
VacA s1m2 strain. However, Bachir et al (38) and Xu and Xie (39) demonstrated that the VacA
genotype was not linked to H. pylori resistance. The present
results indicated that there was no significant difference in H.
pylori multidrug CLA, MNZ and LEFX resistance between
VacA m1 and VacA m2 strains. The present results were
consistent with studies by Bachir et al (38) and Xu and Xie (39). However, the s2 strain was not
detected and the s1 subtype was not classified into s1a,s1b and s1c
in the current study. Intermediate region i1, i2 and i3 were not
detected.
Antibiotic resistance is divided into primary and
secondary resistance (40). The
antibiotic resistance rate increases in association with the number
of therapy failures (38). The
majority of patients included in the present study has received
treatment before; however, due to the unknown antibiotic history of
the patient, it is not possible to accurately calculate the primary
and secondary drug resistance rates of H. pylori in
Zhejiang, China, thus further research is required. The average
time required to culture H. pylori and perform an in
vitro drug sensitivity test is 1 week. However, it only takes 1
day to detect drug-resistant genes, which can save valuable time.
In addition, Giorgio et al (41) demonstrated that reverse
transcription-quantitative PCR on feces, which is non-invasive
compared with the use of gastric mucosa, can be a useful tool for
the detection of DNA sequence and antibiotic resistance-related
gene mutations in H. pylori. A previous study used gene chip
technology to detect the drug resistance of H. pylori in
children, and the main mutation sites were similar to those
identified in the present study (17). Studies focused on adults have only
been used for the detection of single antibiotic resistance
(42,43). The chip used in the present study
covers five commonly used antibiotics. The results of gene
detection were not identical to drug sensitivity test in
vitro. At present, there is a lack of relevant clinical data to
compare the accuracy of the two methods. However, a relevant
clinical trial is in progress. There are numerous non-conventional
rapid methods for antimicrobial resistance diagnostics, including
whole genome sequencing (WGS), matrix-assisted laser
desorption/ionization time-of-flight spectrometry, Fourier
transform infrared spectroscopy and microfluidics technology. WGS
is being expanded in China, while other methods require high
purchase and maintenance costs and large space for equipment.
Therefore, none of them can be widely carried out in the clinic
(44).
In conclusion, VacA m subtypes were not
linked to H. pylori antibiotic resistance. The detection of
associated mutant genes C556G, A562T and A562G in PBP1 for
AMX; T87A, A91G, G91T and G91A in gyrA for LEFX; 926-928
(AGA to TTC) and 926-927 (AG to GT) in 16S rRNA for TET;
A2142G and A2143G in 23S rRNA for CLA; and G616A in
rdxA for MNZ has a clinical application value in predicting
the antibiotic resistance of H. pylori, particularly the
resistance to CLA and LEFX.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by Zhejiang Natural
Science Foundation Committee (grant no. Q20H030019), Zhejiang
Administration of Traditional Chinese Medicine (grant no.
2017ZKL010) and Zhejiang Province Science and Technology Hall
(grant no. 2016C37033).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
BL designed the study. JZ analyzed and interpreted
data. JD analyzed data and wrote the manuscript. YH and LM
performed experiments and revised the manuscript. All authors read
and approved the final manuscript. JD and JZ confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The First Affiliated Hospital of Zhejiang Traditional
Chinese Medical University (approval no. 2017-KL-054-02; Hangzhou,
China). All patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Marshall BJ and Warren JR: Unidentified
curved bacilli in the stomach of patients with gastritis and peptic
ulceration. Lancet. 1:1311–1315. 1984.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Schistosomes, liver flukes and
Helicobacter pylori. IARC Working Group on the evaluation of
carcinogenic risks to humans. Lyon, 7-14 June 1994. IARC Monogr
Eval Carcinog Risks Hum 61: 1-241, 1994.
|
|
3
|
Sugano K, Tack J, Kuipers EJ, Graham DY,
El-Omar EM, Miura S, Haruma K, Asaka M, Uemura N and Malfertheiner
P: faculty members of Kyoto Global Consensus Conference. Kyoto
global consensus report on Helicobacter pylori gastritis.
Gut. 64:1353–1367. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Cheng H, Hu F, Xie Y, Hu P, Wang J, Lv N,
Zhang J, Zhang G, Zhou Z, Wu K, et al: Prevalence of
Helicobacter pylori resistance to antibiotics and its
influence on the treatment outcome in china: A mulficenter clinical
study. Chin J Gastroenterol. 12:525–530. 2007.
|
|
5
|
Song Z, Zhang J, He L, Chen M, Hou X, Li Z
and Zhou L: Prospective multi-region study on primary antibiotic
resistance of Helicobacter pylori strains isolated from
Chinese patients. Dig Liver Dis. 46:1077–1081. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sun QJ, Liang X, Zheng Q, Gu WQ, Liu WZ,
Xiao SD and Lu H: Resistance of Helicobacter pylori to
antibiotics from 2000 to 2009 in Shanghai. World J Gastroenterol.
16:5118–5121. 2010.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Liu DS, Wang YH, Zeng ZR, Zhang ZY, Lu H,
Xu JM, Du YQ, Li Y, Wang JB, Xu SP, et al: Primary antibiotic
resistance of Helicobacter pylori in Chinese patients: A
multiregion prospective 7-year study. Clin Microbiol Infect.
24:780.e5–780.e8. 2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Hu Y, Zhang M, Lu B and Dai J:
Helicobacter pylori and antibiotic resistance, a continuing
and intractable problem. Helicobacter. 21:349–363. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Shao Y, Lu R, Yang Y, Xu Q, Wang B and Ye
G: Antibiotic resistance of Helicobacter pylori to 16
antibiotics in clinical patients. J Clin Lab Anal.
32(e22339)2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Khoury J, Geffen Y, Shaul R, Sholy H,
Chowers Y and Saadi T: Secondary antibiotic resistance of
Helicobacter pylori isolates in Israeli children and adults.
J Glob Antimicrob Resist. 10:182–185. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Saniee P, Hosseini F, Kadkhodaei S,
Siavoshi F and Khalili-Samani S: Helicobacter pylori
multidrug resistance due to misuse of antibiotics in Iran. Arch
Iran Med. 21:283–288. 2018.PubMed/NCBI
|
|
12
|
Savoldi A, Carrara E, Graham DY, Conti M
and Tacconelli E: Prevalence of antibiotic resistance in
Helicobacter pylori: A systematic review and meta-analysis
in world health organization regions. Gastroenterology.
155:1372–1382.e17. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Pohl D, Keller PM, Bordier V and Wagner K:
Review of current diagnostic methods and advances in
Helicobacter pylori diagnostics in the era of next
generation sequencing. World J Gastroenterol. 25:4629–4660.
2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Thi Huyen Trang T, Thanh Binh T and
Yamaoka Y: Relationship between vacA types and development of
gastroduodenal diseases. Toxins (Basel). 8(182)2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Karabiber H, Selimoglu MA, Otlu B,
Yildirim O and Ozer A: Virulence factors and antibiotic resistance
in children with Helicobacter pylori gastritis. J Pediatr
Gastroenterol Nutr. 58:608–612. 2014.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Liou JM, Chang CY, Chen MJ L, et al: The
Primary Resistance of Helicobacter pyloriin Taiwan after the
National Policy to Restrict Antibiotic Consumption and Its Relation
to Virulence Factors-A Nationwide Study. PLoS One. 10(5,
e0124199)2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Yin G, Bie S, Gu H, Shu X, Zheng W, Peng
K, Zhao H, Li F, Chen B, Botchway BOA, et al: Application of gene
chip technology in the diagnostic and drug resistance detection of
Helicobacter pylori in children. J Gastroenterol Hepatol.
35:1331–1339. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Brisson-Noël A, Trieu-Cuot P and Courvalin
P: Mechanism of action of spiramycin and other macrolides. J
Antimicrob Chemother. 22 (Suppl B):S13–S23. 1988.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Seo SI, Do BJ, Kang JG, Kim HS, Jang MK,
Kim HY and Shin WG: Helicobacter pylori eradication
according to sequencing-based 23S ribosomal RNA point mutation
associated with clarithromycin resistance. J Clin Med.
9(54)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ong S, Kim SE, Kim JH, Yi NH, Kim TY, Jung
K, Park MI and Jung HY: Helicobacter pylori eradication
rates with concomitant and tailored therapy based on 23S rRNA point
mutation: A multicenter randomized controlled trial. Helicobacter.
24(e12654)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Kim YJ, Chung WC and Kim DB: Efficacy of
bismuth added to standard triple therapy as the first-line
eradication regimen for Helicobacter pylori infection.
Helicobacter. 26(e12792)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Hooper DC: Emerging mechanisms of
fluoroquinolone resistance. Emerg Infect Dis. 7:337–341.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Liou JM, Chen CC, Chen MJ, Chang CY, Fang
YJ, Lee JY, Sheng WH, Wang HP, Wu MS and Lin JT: Empirical modified
sequential therapy containing levofloxacin and high-dose
esomeprazole in second-line therapy for Helicobacter pylori
infection: A multicentre clinical trial. J Antimicrob Chemother.
66:1847–1852. 2011.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Papastergiou V, Mathou N, Licousi S,
Evgenidi A, Paraskeva KD, Giannakopoulos A, Stavrou PZ, Platsouka E
and Karagiannis JA: Seven-day genotypic resistance-guided triple
Helicobacter pylori eradication therapy can be highly
effective. Ann Gastroenterol. 31:198–204. 2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Palmitessa V, Monno R, Panarese A, Cuppone
R, Burattini O, Marangi S, Curlo M, Fumarola L, Petrosillo A,
Parisi A, et al: Evaluation of antibiotic resistance of
Helicobacter pylori strains isolated in Bari, Southern
Italy, in 2017-2018 by phenotypic and genotyping methods. Microb
Drug Resist. 26:909–917. 2020.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Miyachi H, Miki I, Aoyama N, Shirasaka D,
Matsumoto Y, Toyoda M, Mitani T, Morita Y, Tamura T, Kinoshita S,
et al: Primary levofloxacin resistance and gyrA/B mutations among
Helicobacter pylori in Japan. Helicobacter. 11:243–249.
2006.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Di Caro S, Fini L, Daoud Y, Grizzi F,
Gasbarrini A, De Lorenzo A, Di Renzo L, McCartney S and Bloom S:
Levofloxacin/amoxicillin-based schemes vs quadruple therapy for
Helicobacter pylori eradication in second-line. World J
Gastroenterol. 18:5669–5678. 2012.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Debnath N and Yadav A: Indigenous
probiotic lactobacillus strains to combat gastric pathogen
Helicobacter pylori: Microbial interference therapy. Indu
pal Kaur, Sandip V. Pawar and Praveen rishi. Chandigarh, India. In:
Probiotic Research in Therapeutics, pp215-230, 2021.
|
|
29
|
Zamani M, Rahbar A and Shokri-Shirvani J:
Resistance of Helicobacter pylori to furazolidone and
levofloxacin: A viewpoint. World J Gastroenterol. 23:6920–6922.
2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Xie C and Lv N: Helicobacter pylori
infection in China. Dis Surveill. 33:272–275. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lee SM, Kim N, Kwon YH, Nam RH, Kim JM,
Park JY, Lee YS and Lee DH: rdxA, frxA, and efflux pump in
metronidazole-resistant Helicobacter pylori: Their relation
to clinical outcomes. J Gastroenterol Hepatol. 33:681–688.
2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Jeong JY, Mukhopadhyay AK, Dailidiene D,
Wang Y, Velapatiño B, Gilman RH, Parkinson AJ, Nair GB, Wong BC,
Lam SK, et al: Sequential inactivation of rdxA (HP0954) and frxA
(HP0642) nitroreductase genes causes moderate and high-level
metronidazole resistance in Helicobacter pylori. J
Bacteriol. 182:5082–5090. 2000.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Jeong JY and Berg DE: Mouse-colonizing
Helicobacter pylori SS1 is unusually susceptible to
metronidazole due to two complementary reductase activities.
Antimicrob Agents Chemother. 44:3127–3132. 2000.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Kwon DH, El-Zaatari FA, Kato M, Osato MS,
Reddy R, Yamaoka Y and Graham DY: Analysis of rdxA and involvement
of additional genes encoding NAD(P)H flavin oxidoreductase (FrxA)
and ferredoxin-like protein (FdxB) in metronidazole Helicobacter
pylori. Antimicrob Agents Chemother. 44:2133–2142.
2000.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Rasheed F, Campbell BJ, Alfizah H, Varro
A, Zahra R, Yamaoka Y and Pritchard DM: Analysis of Clinical
Isolates of Helicobacter pylori in Pakistan reveals high
degrees of pathogenicity and high frequencies of antibiotic
resistance. Helicobacter. 19:387–399. 2014.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Zhang Y, Wen Y, Xiao Q, Zheng W, Long G,
Chen B, Shu X and Jiang M: Mutations in the antibiotic target genes
related to clarithromycin, metronidazole and levofloxacin
resistance in Helicobacter pylori strains from children in
China. Infect Drug Resist. 13:311–322. 2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Wang D, Guo Q, Yuan Y and Gong Y: The
antibiotic resistance of Helicobacter pylori to five
antibiotics and influencing factors in an area of China with a high
risk of gastric cancer. BMC Microbiol. 19(152)2019.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Bachir M, Allem R, Tifrit A, Medjekane M,
Drici AE, Diaf M and Douidi KT: Primary antibiotic resistance and
its relationship with cagA and vacA genes in Helicobacter
pylori isolates from Algerian patients. Braz J Microbiol.
49:544–551. 2018.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Xu J and Xie GY: Relationship between cagA
and VacA genotypes of Helicobacter pylori and drug
resistance. Med J West China. 5:818–819, 821. 2010.(In
Chinese).
|
|
40
|
Schubert JP, Gehlert J, Rayner CK,
Roberts-Thomson IC, Costello S, Mangoni AA and Bryant RV:
Antibiotic resistance of Helicobacter pylori in Australia
and New Zealand: A systematic review and meta-analysis. J
Gastroenterol Hepatol. 36:1450–1456. 2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Giorgio F, Ierardi E, Sorrentino C,
Principi M, Barone M, Losurdo G, Iannone A, Giangaspero A, Monno R
and Di Leo A: Helicobacter pylori DNA isolation in the
stool: An essential pre-requisite for bacterial noninvasive
molecular analysis. Scand J Gastroenterol. 15:1429–1432.
2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Chen S, Li Y and Yu C: Oligonucleotide
microarray: A new rapid method for screening the 23S rRNA gene of
Helicobacter pylori for single nucleotide polymorphisms
associated with clarithromycin resistance. J Gastroenterol Hepatol.
23:126–131. 2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Xuan SH, Zhou YG, Shao B, Cui YL, Li J,
Yin HB, Song XP, Cong H, Jing FX, Jin QH, et al: Enzymic
colorimetry-based DNA chip: A rapid and accurate assay for
detecting mutations for clarithromycin resistance in the 23S rRNA
gene of Helicobacter pylori. J Med Microbiol. 58:1443–1448.
2009.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Kaprou GD, Bergšpica I, Alexa EA,
Alvarez-Ordóñez A and Prieto M: Rapid methods for antimicrobial
resistance diagnostics. Antibiotics (Basel). 10:209–239.
2021.PubMed/NCBI View Article : Google Scholar
|