Introduction
Lung cancer has brought a heavy burden to human
society and families, with high morbidity and mortality rates among
all types of malignant tumors (1).
The statistical report on cancer incidence and mortality published
by the International Cancer Center (GLOBOCAN 2018) suggests that
lung cancer is a leading cause of death of patients with cancer in
China and worldwide (2). Globally,
it is estimated that there are 2.1 million new cases of lung cancer
and 1.8 million lung cancer-related deaths each year (2). In China, the incidence of lung cancer
is reported to be 5.7 per million and the total number of new cases
is ~787,000, accounting for 20% of the total malignant tumor cases
(3).
The occurrence and development of lung cancer are
closely related to the tumor micro-environment, personal living
habits and genetic differences (4,5).
Smoking, history of lung-related diseases, air pollutants, family
history of tumors, ethnic differences, obesity, occupational
exposure and poor eating habits may increase the risk of lung
cancer (6). Although smoking is a
well-recognized risk factor for lung cancer, lung cancer also
occurs in 10-25% of non-smokers. The influence of personal genetic
factors on the pathogenesis of lung cancer should not be
underestimated (7,8).
With the rise of molecular epidemiology, the ENCODE
project indicated that up to 75% of human genome nucleotides may be
transcribed into the original transcripts, producing a series of
common RNAs including non-coding RNAs (ncRNA) (9). A large number of studies have
indicated that most ncRNAs do not encode proteins but have specific
functions (10). At the level of
apparent modification, long ncRNA (lncRNA) affects numerous aspects
of chromosome biology by remodeling chromatin structure and
regulating a large number of genes. Based on its various
characteristics and regulatory mechanisms, the abnormal expression
of lncRNA may also be involved in the occurrence, development and
metastasis of tumors, thus affecting tumor staging and the
occurrence of various cancers (11,12).
The potential impact of lncRNA on the occurrence, development,
treatment and prognosis of lung cancer is worthy of further
investigation (13).
Single nucleotide polymorphism (SNP) refers to the
polymorphism in the DNA sequence caused by the variation of a
single nucleotide in the genome. It is a heritable variation that
exists widely in the human genome (14). It is estimated that there is one
SNP per 1,000 bases on average, which is recognized as the
third-generation genetic marker after restriction fragment length
polymorphism and microsatellites. SNP is involved in the
transcription of coding genes by influencing the binding of
transcription factors, thereby affecting gene expression.
Researchers have discovered a large number of SNPs associated with
diseases through genome-wide association studies and about
one-third of the SNPs are located in non-protein coding regions,
indicating the SNPs of lncRNA may also have an important role in
disease (15). Emerging evidence
suggested that the SNPs of lncRNAs are related to the
susceptibility to and prognosis of various types of malignant
tumor, such as breast cancer, gastric cancer and colorectal cancer
(16).
In recent years, researchers have discovered that
the homeobox (HOX) gene family participates in embryonic
development (17). In their study
from 2007, Rinn et al (18)
used high-resolution microarray technology to identify and analyze
hundreds of ncRNAs transcribed from the HOX gene cluster from 11
types of fibroblasts and discovered HOX transcript antisense
intergenic RNA (HOTAIR) at the HOXC gene locus located on
chromosome 12. HOTAIR was recently considered a novel oncogene, but
it is controversial whether the SNP of lncRNA HOTAIR has a risk
effect to influence lung cancer susceptibility (19). LncRNA HOTAIR is located at 12q13.13
and the length is 2.1 kb. There are four gene clusters in the
genome: HOXA, HOXB, HOXC and HOXD (20). The transcription of these four gene
clusters produces several long-chain ncRNAs and their expression
has a temporal and spatial specificity (21). Although HOTAIR is produced from the
HOXC sequence, it has the function of inhibiting the expression of
the HOXD gene. The secondary structure of HOTAIR contains four
independent folding units, two of which are evolutionarily
conserved protein-binding domains (22). HOTAIR is produced and enriched in
the nucleus and expressed in the cytoplasm. The HOTAIR-mediated
epigenetic silencing function works by acting as a scaffold for the
polycomb repressive complex 2 (PRC2) and lysine specific
demethylase 1/RE1-silencing transcription factor (REST)
co-repressor 1/REST complex. PRC2 binds to the 5' end of HOTAIR and
regulates chromosome occupancy through the subunit EZH2 of the PRC2
complex, leading to the methylation of histone H3 lysine in the
chromosome sequence region of the HOXD gene (23). Studies have indicated that at least
16 types of malignant tumor have increased expression of HOTAIR
(24-26).
Furthermore, it was reported that overexpressed HOTAIR combines
with PRC2 to regulate gene expression patterns by changing the
methylation status of H3K27, thus increasing the ability of cancer
cells to metastasize and invade. Knockdown of HOTAIR or PRC2 is
able to inhibit the invasion and metastasis ability of cancer cells
(27). A number of studies
focusing on lung cancer have reported that HOTAIR has a
proto-oncogene function in lung cancer. For instance, compared with
adjacent tissues, the expression of HOTAIR in lung cancer tissue is
significantly higher (28). The
increased HOTAIR expression in non-small cell lung cancer is
similar to that of lung adenocarcinoma and lung squamous cells
(29). Another study determined
that elevated HOTAIR expression was associated with the brain
metastasis rate of lung cancer (30). The association between the SNP of
lncRNA HOTAIR and lung cancer susceptibility remains to be further
investigated.
Thus, the present systematic review and
meta-analysis was performed to investigate the association between
lncRNA HOTAIR SNP and lung cancer susceptibility.
Materials and methods
Literature search and search
strategy
This systematic review and meta-analysis was
performed following the preferred reporting items for systematic
review and meta-analysis (PRISMA) guidelines (31). A computerized literature search was
performed in the PubMed (https://pubmed.ncbi.nlm.nih.gov/), Ovid Medline
(https://www.wolterskluwer.com/en/solutions/ovid/ovid-medline-901),
Embase (https://www.embase.com) and Cochrane
Library databases (https://www.cochranelibrary.com/) up to May 2nd 2021.
Relevant studies were also searched in Chinese databases, including
Chinese National Knowledge Infrastructure (CNKI; https://www.cnki.net/) and Wanfang (https://www.wanfangdata.com.cn/index.html). Google
scholar (https://scholar.google.com/) was
searched for gray literature that is not formally published in
sources such as books or journal articles.
Regarding the search strategy, terms included the
following items: (‘lncRNA’ or ‘long non-coding RNA’) and (‘SNP’ or
‘polymorphism’ or ‘variants’) and (‘lung cancer’ or ‘lung tumor’ or
‘lung neoplasm’ or ‘adenocarcinoma’ or ‘lung carcinoma’) and
(‘HOTAIR’ or ‘HOX transcript antisense intergenic RNA’). The items
were searched for keywords and medical sub-headings. All results
were downloaded and imported into Endnote X6 (Thomson Reuters) for
further literature screening.
Inclusion and exclusion criteria
All of the studies evaluating and assessing the
association between lncRNA HOTAIR polymorphism and the
susceptibility to lung cancer were included in the present study.
The inclusion criteria were as follows: i) Patients with lung
cancer; ii) case-control studies on the polymorphism of lncRNA
HOTAIR; iii) number of patients >50. The exclusion criteria were
as follows: i) Reviews, observational studies, comments, case
series and case reports; ii) lack of data on lncRNA HOTAIR; iii)
lack of information on lung cancer risk; and iv) lack of available
data that were able to be extracted from the text/table/graphs or
obtained from the authors on request. There was no limitation in
terms of the language of the studies included and the reference
lists of reviews were also screened for the retrieval and further
inclusion of studies.
Literature screening, data extraction
and quality assessment
A total of two investigators (CK and XF)
independently screened titles and abstracts according to the
inclusion and exclusion criteria. The full-text was further
screened for data extraction or assessed for the inclusion
criteria. In the case of any disagreements, a third investigator
(XH) was consulted to resolve the divergence.
The data were extracted by two reviewers
independently (CK and XF). All the data were imported into a
standard form, including the name of the first author of the study,
publication year, recruitment years and country, genotyping methods
and the common polymorphism of HOTAIR. In addition, the sample
size, as well as the age, sex, smoking history and alcohol use of
healthy participants (control group) and patients with lung cancer
(case group) were recorded. The raw data are provided as
supplemental material (Table
SI).
The study quality was evaluated according to
Newcastle-Ottawa Scale (NOS), with a score of 7-9 considered to
indicate high quality, whereas a score of 0-6 was indicative of low
quality (32).
Statistical analysis
The meta-analysis was performed with Stata 15.0
software (StataCorp LP). The genotype frequencies of lncRNA HOTAIR
polymorphisms for the Hardy-Weinberg equilibrium (HWE) were
calculated for the controls using the χ2 test and
P<0.05 was considered to indicate a significant disequilibrium.
The risk was pooled by the odds ratio (OR) and 95% confidence
intervals (CI). The result of the pooled OR was visualized through
the forest plot by using the metan module of Stata. The
heterogeneity existing among studies was evaluated by the
I2 statistic and χ2 test, with I2
≥50% indicating the presence of heterogeneity. The random-effects
model was used if heterogeneity was present among studies, while
otherwise, the fixed-effects model was used. The risks of the
heterozygote and variant homozygote compared with the wild-type
homozygote were estimated. Furthermore, the dominant and recessive
mode of the variant allele were evaluated. The sensitivity analysis
was performed by reperforming the OR calculation while omitting one
individual study at a time. Sensitivity analysis was performed for
rs12826786, rs1899663, rs4759314 and rs920778 for all modes,
including the dominant mode, recessive mode, additive mode,
heterozygote mode, homozygote mode and allele mode. Egger's linear
regression method was used to statistically assess the publication
bias by Stata 15.0. The funnel plot for exploring the
missed-reported articles was also analysed with the
metafunnel module of Stata to visually display the outcomes
of publication bias assessment. Subgroup analysis was performed
using the metan module of Stata. The further heterogeneity
analysis was performed based on the results of the subgroup
analysis. P<0.05 was considered to indicate statistical
significance for all analyses.
Results
Study selection and characteristics of
included studies
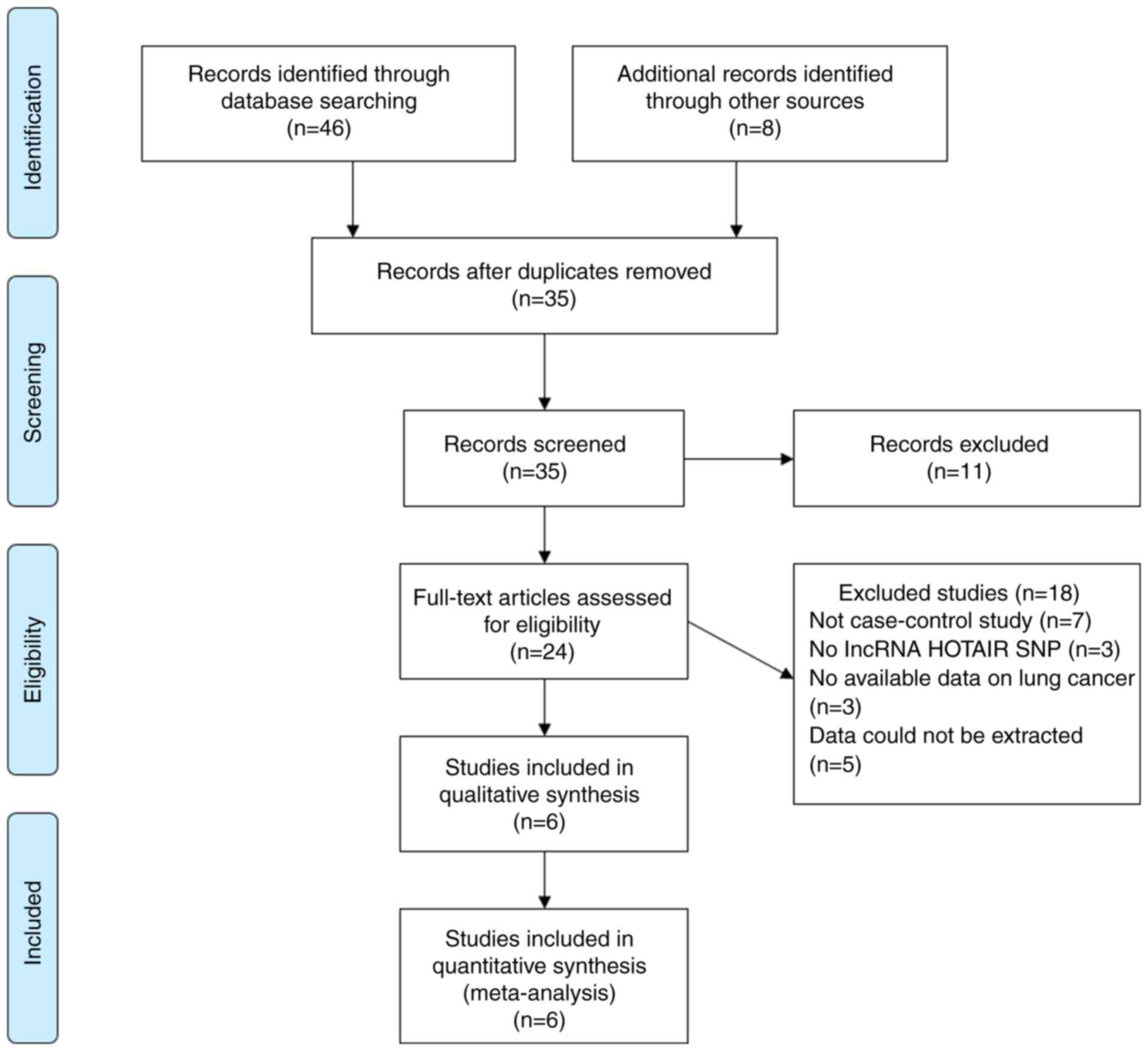
A total of 46 studies were retrieved by searching
the databases as described above, as well as 8 studies via other
methods. Thus, a total of 54 studies focusing on the association of
lncRNA HOTAIR with lung cancer were identified based on the search
strategy. After excluding duplicates and other studies based on the
inclusion and exclusion criteria, six studies were finally included
in the present study (33-38).
The literature screening process is presented in Fig. 1.
The characteristics of the included studies are
described in Table I. The year of
publication ranged from 2016 to 2020 and the recruitment period
ranged from 1995 to 2017. Of the six studies, four were from China
and the other two studies were from Turkey and Japan, respectively.
Furthermore, two studies used MassArray to detect the SNPs of
lncRNA, while the other four studies used the TaqMan assay instead.
A total of 1,715 patients with lung cancer and 2,745 healthy
participants were included in the analysis of the present study.
The average age of the lung cancer group was 57.9-64.3 years, while
it was 56.6-64.8 years for the healthy participants (control
group). The proportion of male subjects ranged from 29.9 to 96.9%
in the control group, as compared to 35.4-81.6% in the lung cancer
group. A history of smoking was present in 20.1-96.6% of the
healthy participants and in 29.9-99.4% of the patients with lung
cancer. The results of the quality assessment are listed in
Table I. Of the studies, four
scored as >6 and were considered to be of high quality.
 | Table ICharacteristics of the included
studies. |
Table I
Characteristics of the included
studies.
| | Control group | Case group | |
|---|
| Author (year) | Country | Recruitment
year | Cancer type | Ethnicity | Genotyping
method | Quality score | Sample, n | Age, years | Male sex, n
(%) | Smoking, n (%) | Alcohol
consumption, n (%) | Sample, n | Age, years | Male sex, n
(%) | Smoking, n (%) | Alcohol
consumption, n (%) | (Refs.) |
|---|
| Ren (2020) | China | 2015-2017 | All lung
cancer | Asian | MassArray
system | 8 | 196 | 64.82±10.00 | 139 (70.9) | 190 (96.9) | 188 (95.9) | 196 | 64.32±10.26 | 136 (69.4) | 195 (99.4) | 196(100) | (33) |
| Minn (2020) | Japan | 1995-2006 | All lung
cancer | Asian | TaqMan
Assay-PCR | 8 | 1,241 | NG | NG | NG | NG | 129 | NG | NG | NG | NG | (34) |
| Wang (2018) | China | 2006-2010 | All lung
cancer | Asian | TaqMan
Assay-PCR | 7 | 451 | NG | 280 (62.1) | 159 (35.2) | NG | 262 | NG | 188 (71.8) | 169 (64.5) | NG | (35) |
| Li (2018) | China | NG | All lung
cancer | Asian | TaqMan
Assay-PCR | 7 | 551 | 56.69±15.65 | 165 (29.9) | 111 (20.1) | NG | 543 | 57.99±11.51 | 192 (35.4) | 165 (30.4) | NG | (36) |
| Dadaş (2018) | Turkey | NG | NSCLC | Caucasian | TaqMan
Assay-PCR | 6 | 93 | 61.77±12.00 | 67 (72.0) | 61 (65.6) | NG | 87 | 59.27±10.55 | 71 (81.6) | 26 (29.9) | NG | (37) |
| Gong (2016) | China | 2011-2013 | All lung
cancer | Asian | MassARRAY | 5 | 213 | NG | 80 | NG | NG | 498 | NG | 394 (79.1) | NG | NG | (38) |
Genotype frequency distributions
Table II displays
the genotype frequency distributions of the five SNPs of lncRNA
HOTAIR. Regarding HOTAIR rs128261786 and HOTAIR rs1899663,
PHWE<0.05 was calculated for the study by Wang et
al (35), indicating
unbalanced data in terms of the two SNPs.
| Table IICommon gene polymorphisms of homeobox
transcript antisense intergenic RNA in the included studies. |
Table II
Common gene polymorphisms of homeobox
transcript antisense intergenic RNA in the included studies.
| A, rs920778 |
|---|
| | Controls | Cases | |
|---|
| Author (year) | Total | AA | AG | GG | Total | AA | AG | GG |
PHWE | (Refs.) |
|---|
| Ren (2020) | 183 | 104 | 69 | 10 | 184 | 114 | 60 | 10 | 0.7416 | (33) |
| Minn (2020) | 1,241 | 698 | 473 | 70 | 129 | 75 | 53 | 1 | 0.3844 | (34) |
| Wang (2018) | 451 | 233 | 192 | 26 | 262 | 110 | 132 | 20 | 0.0947 | (35) |
| Li (2018) | 551 | 326 | 203 | 22 | 543 | 324 | 197 | 22 | 0.1640 | (36) |
| B, rs12826786 |
| | Controls | Cases | |
| Author (year) | Total | AA | AG | GG | Total | AA | AG | GG |
PHWE | (Refs.) |
| Wang (2018) | 451 | 355 | 85 | 11 | 262 | 200 | 49 | 13 | 0.0047 | (35) |
| Li (2018) | 551 | 381 | 156 | 14 | 543 | 340 | 185 | 18 | 0.6767 | (36) |
| Dadaş (2018) | 93 | 38 | 41 | 14 | 87 | 33 | 41 | 13 | 0.5934 | (37) |
| C, rs1899663 |
| | Controls | Cases | |
| Author (year) | Total | AA | AG | GG | Total | AA | AG | GG |
PHWE | (Refs.) |
| Ren (2020) | 188 | 123 | 60 | 5 | 187 | 131 | 52 | 4 | 0.4656 | (33) |
| Wang (2018) | 452 | 254 | 189 | 9 | 262 | 178 | 66 | 18 | 0.0001 | (35) |
| Dadaş (2018) | 93 | 41 | 40 | 12 | 87 | 39 | 37 | 11 | 0.6495 | (37) |
| D, rs4759314 |
| | Controls | Cases | |
| Author (year) | Total | AA | AG | GG | Total | AA | AG | GG |
PHWE | (Refs.) |
| Wang (2018) | 451 | 412 | 37 | 2 | 262 | 238 | 21 | 3 | 0.2463 | (35) |
| Ren (2020) | 184 | 168 | 16 | 0 | 175 | 158 | 17 | 0 | 0.5375 | (33) |
| Li (2018) | 551 | 489 | 61 | 1 | 543 | 526 | 16 | 1 | 0.5268 | (36) |
Main findings
The association between different HOTAIR SNPs and
lung cancer susceptibility is presented in Table III. No significant association
with the susceptibility for lung cancer was obtained for SNPs
rs12826786, rs4359714 and rs920778 (P>0.05 for all modes).
However, the lncRNA HOTAIR rs1899663 C>A polymorphism was a risk
factor for lung cancer (dominant mode, AA+CA vs. CC: OR=0.816, 95%
CI=0.707-0.942, P=0.005). However, for the recessive mode, additive
mode, heterozygote mode, homozygote mode and allele mode, no
significant association with lung cancer was observed.
| Table IIIAssociation of homeobox transcript
antisense intergenic RNA gene polymorphisms and lung cancer
risk. |
Table III
Association of homeobox transcript
antisense intergenic RNA gene polymorphisms and lung cancer
risk.
| SNP/mode | Comparison | N | OR (95% CI) | P-value | I2
(%) | Model for
meta-analysis |
|---|
| rs12826786 | | | | | | |
|
Dominant | TT+CT vs. CC | 3 | 0.964
(0.793-1.172) | 0.716 | 56.30 | Random-effects |
|
Recessive | TT vs. CC+CT | 3 | 1.101
(0.732-1.657) | 0.643 | 42.50 | Fixed-effects |
|
Additive | TT+CC vs. TC | 3 | 1.043
(0.987-1.102) | 0.134 | 26.40 | Fixed-effects |
|
Heterozygote | CT vs. CC | 3 | 0.899
(0.783-1.032) | 0.131 | 29.60 | Fixed-effects |
|
Homozygote | TT vs. CC | 3 | 1.085
(0.730-1.613) | 0.686 | 49.90 | Fixed-effects |
|
Allele | T vs. C | 3 | 0.990
(0.782-1.254) | 0.934 | 66.50 | Random-effects |
| rs1899663 | | | | | | |
|
Dominant | AA+CA vs. CC | 3 | 0.816
(0.707-0.942) | 0.005 | 37.70 | Fixed-effects |
|
Recessive | AA vs. CC+CA | 3 | 1.527
(0.552-4.225) | 0.415 | 72.10 | Random-effects |
|
Additive | AA+CC vs. AC | 3 | 1.135
(0.969-1.331) | 0.117 | 69.90 | Random-effects |
|
Heterozygote | CA vs. CC | 3 | 0.801
(0.607-1.056) | 0.116 | 64.50 | Random-effects |
|
Homozygote | AA vs. CC | 3 | 1.382
(0.588-3.247) | 0.457 | 62.30 | Random-effects |
|
Allele | A vs. C | 3 | 0.886
(0.762-1.030) | 0.115 | 0.00 | Fixed-effects |
| rs4759314 | | | | | | |
|
Dominant | GG+AG vs. AA | 3 | 1.630
(0.720-3.690) | 0.241 | 85.00 | Random-effects |
|
Recessive | GG vs. AA+AG | 2 | 1.933
(0.447-8.350) | 0.377 | 0.00 | Fixed-effects |
|
Additive | AA+GG vs. AG | 3 | 0.966
(0.906-1.030) | 0.285 | 83.60 | Random-effects |
|
Heterozygote | AG vs. AA | 3 | 1.612
(0.670-3.878) | 0.286 | 86.20 | Random-effects |
|
Homozygote | GG vs. AA | 2 | 1.983
(0.456-8.617) | 0.361 | 0.00 | Fixed-effects |
|
Allele | G vs. A | 3 | 1.649
(0.769-3.537) | 0.198 | 82.90 | Random-effects |
| rs920778 | | | | | | |
|
Dominant | GG+AG vs. AA | 4 | 1.029
(0.902-1.173) | 0.670 | 53.60 | Random-effects |
|
Recessive | GG vs. AA+AG | 4 | 0.914
(0.640-1.304) | 0.620 | 43.80 | Fixed-effects |
|
Additive | AA+GG vs. AG | 4 | 0.967
(0.908-1.029) | 0.289 | 36.90 | Fixed-effects |
|
Heterozygote | AG vs. AA | 4 | 1.050
(0.957-1.151) | 0.305 | 42.90 | Fixed-effects |
|
Homozygote | GG vs. AA | 4 | 0.984
(0.555-1.747) | 0.957 | 53.60 | Random-effects |
|
Allele | G vs. A | 4 | 1.008
(0.865-1.176) | 0.917 | 57.70 | Random-effects |
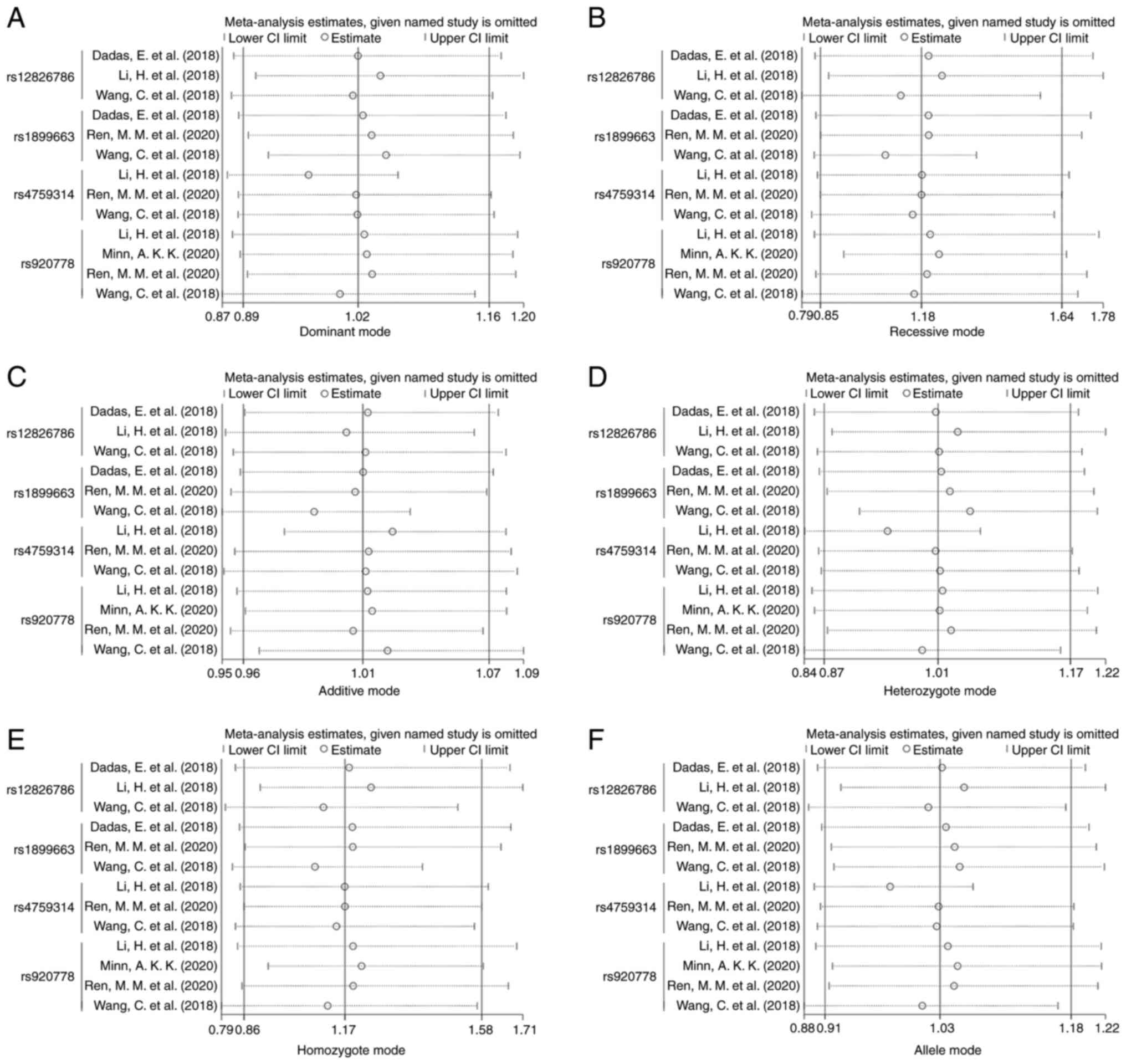
The sensitivity analysis suggested that the omission
of any single study did not significantly alter the overall pooled
ORs in dominant mode, recessive mode, additive mode, heterozygote
mode, homozygote mode and allele mode (Fig. 2).
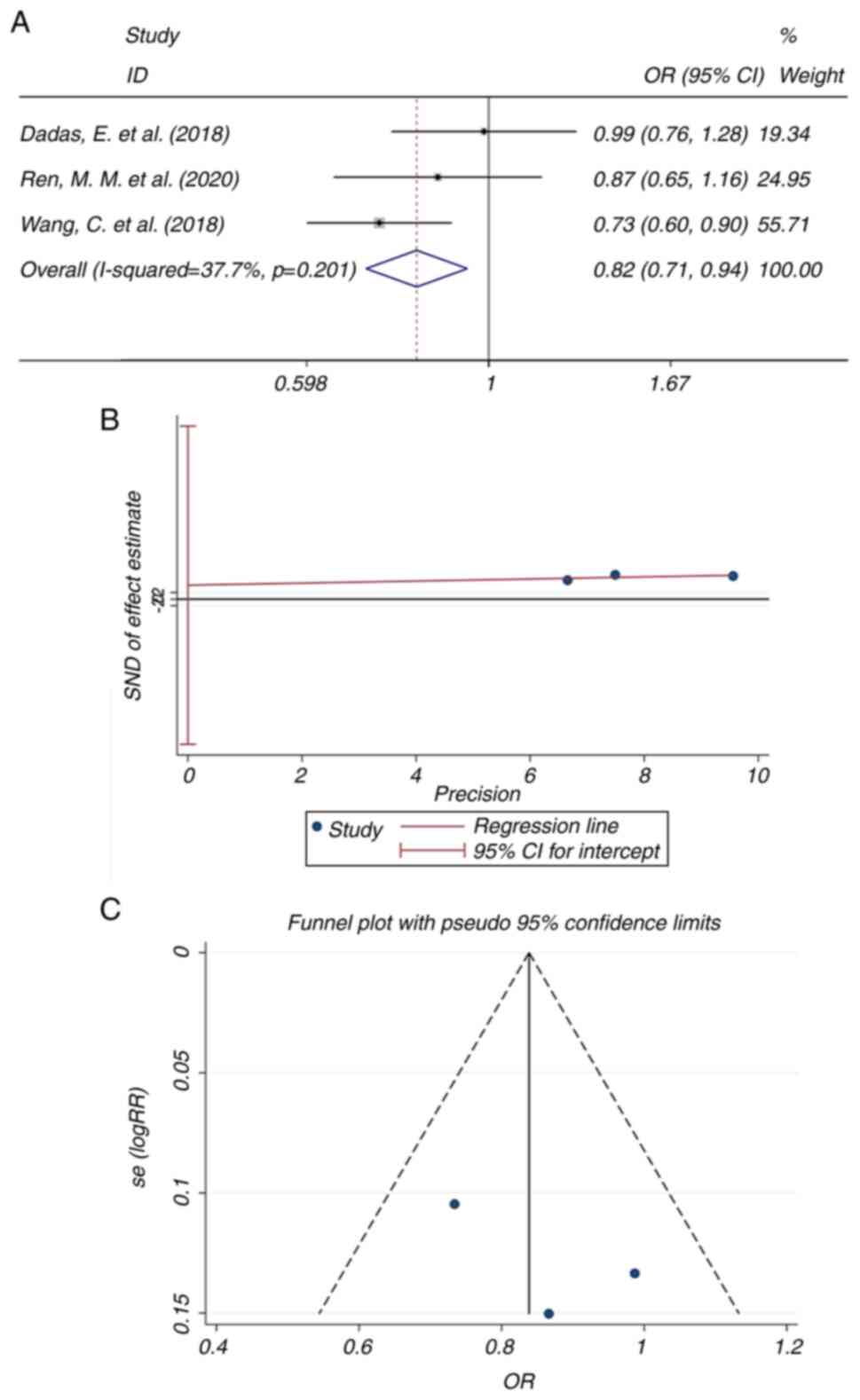
Since the lncRNA HOTAIR rs1899663 C>A
polymorphism was identified as a risk factor for lung cancer
(OR=0.816; Fig. 3A), a further
analysis was performed, including a publication bias analysis and
subgroup analysis in terms of rs1899663 (dominant mode, AA+CA vs.
CC). The P-value as calculated using Egger's test (Fig. 3B) was 0.466 and the funnel plot
indicated a good symmetry (Fig.
3C). These results suggested that no significant publication
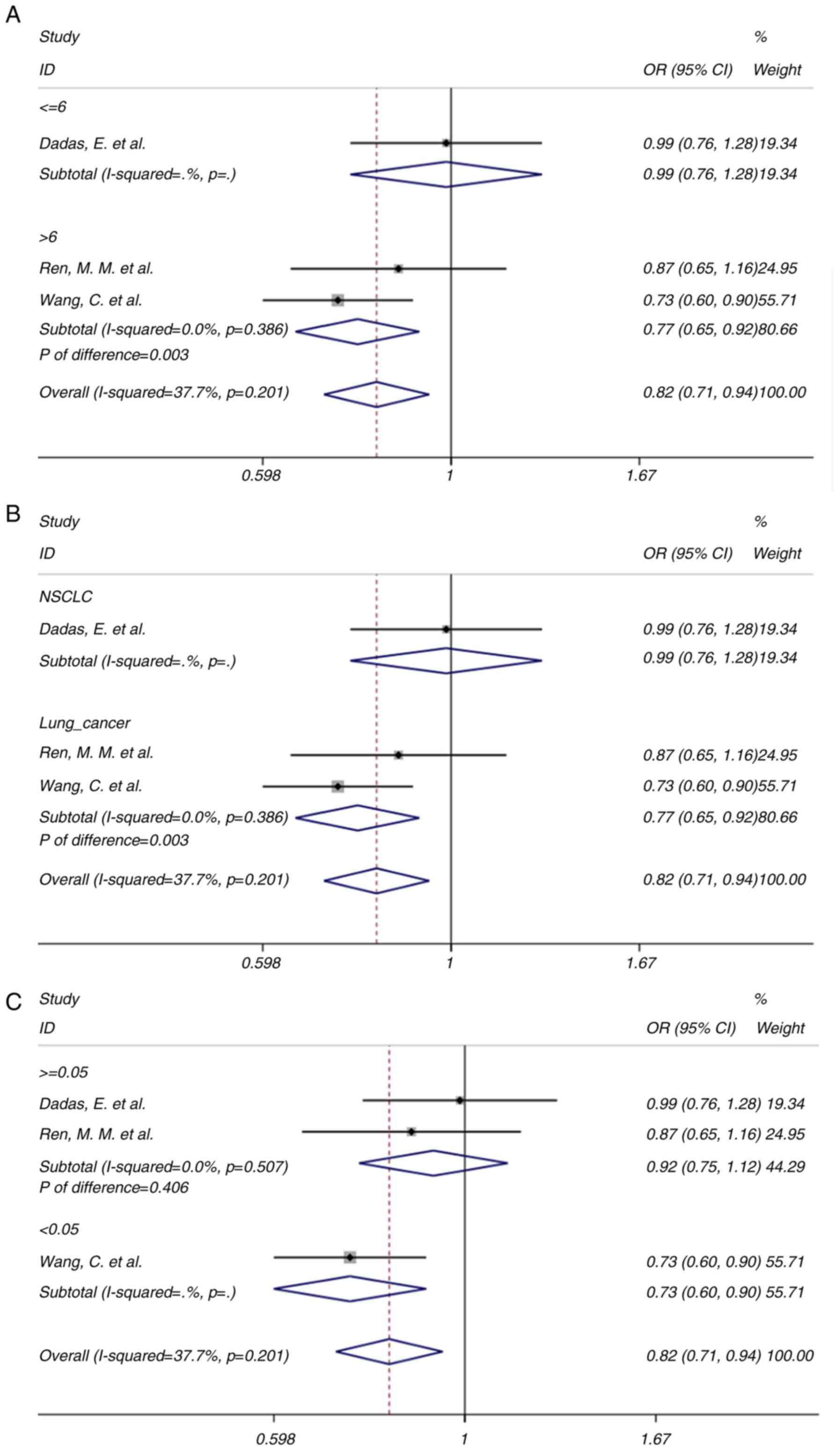
bias was present. In the further subgroup analysis, the studies
with higher quality (NOS score >6) also suggested a significant
association of the rs1899663 C>A polymorphism with the risk of
lung cancer (OR=0.77, 95% CI=0.65-0.92, P=0.003; Fig. 4A). Similar results were obtained in
the subgroup of cancer type (lung cancer) (Fig. 4B). As for the studies with
PHWE≥0.05 (Fig. 4C),
the trend for the risk of lung cancer was similar (OR=0.92, 95%
CI=0.75-1.12, P=0.406). As for the study by Wang et al
(35) with
PHWE<0.05, a significant association was observed
(OR=0.73, 95% CI=0.60-0.90, P=0.003). Of note, in each of the
subgroups, no heterogeneity was detected (I2=0 for all
subgroups with the number of studies >1), indicating that the
NOS quality, type of cancer and gene frequency disequilibrium may
be potential sources of heterogeneity. However, the overall
heterogeneity for the rs1899663 C>A polymorphism was minor
(I2=37.7%, P=0.201).
Discussion
The present study indicated that the lncRNA HOTAIR
rs1899663 C>A polymorphism is a risk factor for lung cancer. In
previous studies on the topic of HOTAIR-associated lung cancer
susceptibility, two or three SNPs were usually investigated in an
individual study. However, at least four SNPs (rs128261786,
rs1899663, rs920778 and rs4759314) of HOTAIR have been reported in
the lung cancer population. The present meta-analysis included all
of these SNPs into the analysis and therefore arrived at a more
comprehensive conclusion. The previous results remain controversial
in that each SNP has been reported to be related to lung cancer in
one or two of these previous studies. However, in the present
meta-analysis, these data were integrated and according to the
results, only rs1899663 was a significant risk factor. To the best
of our knowledge, the present study was the first systematic review
and meta-analysis focusing on the association between lncRNA HOTAIR
and lung cancer susceptibility that has integrated all these four
SNPs into its analysis, which is the novelty of this work.
Genetic differences in DNA sequences lead to
phenotypic variation, which affects changes in phenotypic
characteristics, disease risks and responses to the environment
(39). At the same time, human
genetic diversity is not limited to individual polymorphisms, but
also to the specific combinations of tightly linked alleles. The
most common form of gene polymorphism is SNP, which refers to the
polymorphism in the DNA sequence caused by the variation of a
single nucleotide base in the genome. The types of SNP include
conversion, transversion, insertion and deletion (40). SNP is associated with the function
and expression level of the downstream protein by changing the
protein-coding sequence and influencing the splicing changes of
exons and introns, thereby affecting the individual's
susceptibility to disease, prognosis and responsiveness to drugs
(41).
To date, only a small number of studies have
reported on the relationship between the SNP of the HOTAIR sequence
and the susceptibility to malignancies. A study reported an
enhancer-like sequence on the intron 1,719-2,353 bp downstream of
the transcription start position of HOTAIR (42). There was a risk site for esophageal
squamous cell carcinoma, rs920778, carrying the rs920778T allele.
The expression level of the reported gene was higher than that of
the rs920778C allele. This study also performed SNP typing and
quantitative PCR using esophageal squamous cell carcinoma tissues
and determined that individuals with the genotype rs920778TT had
higher expression of HOTAIR than patients carrying rs920778CC
(42). A study on papillary
thyroid carcinoma indicated that in females, the rs920778TT and
rs920778CT genotype increased the risk of developing papillary
thyroid carcinoma by 0.75- and 0.46-fold, respectively (43). The study also verified the effect
of the rs920778 locus polymorphism on HOTAIR expression in cells
and tissues. It was indicated that cells carrying rs920778T alleles
had higher HOTAIR expression than those with rs920778C alleles. In
thyroid squamous cell carcinoma tissues and normal tissues, the
expression of HOTAIR in cancer tissues and adjacent normal tissues
of rs920778TT and rs920778CT genotype carriers was higher than that
in rs920778CC genotype carriers (43). Previous results suggested that the
expression of HOTAIR in individuals carrying the rs920778T allele
was higher than that in individuals with rs920778C (44). Except for the proto-oncogene role
in lung cancer, HOTAIR has also been indicated to be related to
resistance to chemotherapy. Liu et al (45) reported that HOTAIR expression was
significantly upregulated in cisplatin-resistant lung cancer cells
and that knockdown of HOTAIR by RNA interference was able to
resensitize the responses to cisplatin both in vitro and
in vivo. On the other hand, overexpression of HOTAIR
decreased the sensitivity of lung cancer cells to cisplatin. The
HOTAIR-associated chemosensitivity regulation was indicated to
involve the inhibition of cell proliferation, induction of G0/G1
cell-cycle arrest and apoptosis enhancement through regulation of
cyclin-dependent kinase inhibitor p21WAF1/CIP1 (p21)
expression (45). Accordingly,
another clinical investigation by the same group (28) suggested that high levels of HOTAIR
expression were correlated with a more advanced pathological stage
of non-small cell lung cancer (NSCLC), lymph-node metastasis, as
well as a relatively poor prognosis. The upregulated HOTAIR may
regulate NSCLC cell invasion and metastasis, partially via
downregulation of HOXA5(28). In
the present study, a critical literature review was performed to
further understand the mechanisms of HOTAIR in cancer development.
Apart from the p21WAF1/CIP1 and HOXA5 pathways, the most
commonly acknowledged mechanism of HOTAIR is through the axis of
microRNA (miR) and downstream signaling molecules. These axes
include but are not limited to the following:
HOTAIR/miR-222-3p/Cyclin Dependent Kinase 19 axis (46), HOTAIR/specificity protein
1/miR-199a axis (47),
HOTAIR/miR-29b/phosphate and tension homology deleted on chromosome
10/PI3K axis (48),
HOTAIR/miR-203/zinc finger E-box binding homeobox 1 axis (49), HOTAIR/estrogen receptor
1/miR-130b-3p axis (50),
HOTAIR/miR-34a/Janus kinase 2/STAT3 axis (51), HOTAIR/miR-129-5p/ribosomal protein
L14 axis (52),
HOTAIR/miR-129-5p/frizzled class receptor 7 axis (53) and
HOTAIR/miR-149-5p/doublecortin-like kinase 1 axis (54). Furthermore, HOTAIR also interacts
with several classic signaling pathways, which are commonly
involved in oncogenesis, such as CCL22 signaling (55), the Wnt/β-catenin signaling pathway
(56), EZH2 and H3K27 methylation
signaling (57). All of this
evidence, as well as that provided by the present meta-analysis,
suggests the importance of lncRNA HOTAIR SNP in the development of
lung cancer.
Of note, there are certain limitations to the
present study. First, due to the small sample of the included
studies, it was not possible to analyze certain SNPs because only
one study reported the respective relation. Furthermore, lung
cancer susceptibility was indicated to be associated with the
individual characteristics, such as lifestyle and family history,
which was not possible to be extracted. In addition, the majority
of the populations were from Asia and additional data are required
for other populations. This work was not entered in registries such
as the Cochrane Library or PROSPERO, yet the present meta-analysis
was performed based on the PRISMA guidelines.
In conclusion, the present study was the first to
systematically explore the association between lncRNA HOTAIR SNP
and lung cancer susceptibility. It was indicated that populations
carrying the lncRNA HOTAIR rs1899663 C>A polymorphism may have a
high risk of developing lung cancer. LncRNA HOTAIR may be an
important novel target in lung cancer prevention and prognosis
prediction and future investigations are required to clarify the
intrinsic mechanism of HOTAIR in lung cancer.
Supplementary Material
Raw dataset.
Acknowledgements
We thank our collaborator, Professor Jianjun Qiao
(Department of Dermatology, the First Affiliated Hospital of
Zhejiang University Medicine School, Hangzhou, China), for language
and scientific editing support.
Funding
Funding: No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
Design of the meta-analysis: CK, XF and XH.
Literature screening: CK and XF. Quality assessment: SC and XH.
Check and approving the authenticity of the raw data: CK and XF.
Statistical analysis: CK and XF and JL. Manuscript writing and
revision: CK, XF, JL, SC and XH. All authors have read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Barta JA, Powell CA and Wisnivesky JP:
Global epidemiology of lung cancer. Ann Glob Health.
85(8)2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Cao M and Chen W: Epidemiology of lung
cancer in China. Thorac Cancer. 10:3–7. 2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Taghvaee S, Sowlat MH, Hassanvand MS,
Yunesian M, Naddafi K and Sioutas C: Source-specific lung cancer
risk assessment of ambient PM2.5-bound polycyclic
aromatic hydrocarbons (PAHs) in central Tehran. Environ Int.
120:321–332. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Brasky TM, White E and Chen CL: Long-term,
supplemental, one-carbon metabolism-related vitamin B use in
relation to lung cancer risk in the vitamins and lifestyle (VITAL)
cohort. J Clin Oncol. 35:3440–3448. 2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Hicks BM, Filion KB, Yin H, Sakr L, Udell
JA and Azoulay L: Angiotensin converting enzyme inhibitors and risk
of lung cancer: Population based cohort study. BMJ.
363(k4209)2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Lin KF, Wu HF, Huang WC, Tang PL, Wu MT
and Wu FZ: Propensity score analysis of lung cancer risk in a
population with high prevalence of non-smoking related lung cancer.
BMC Pulm Med. 17(120)2017.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Fortunato O, Borzi C, Milione M, Centonze
G, Conte D, Boeri M, Verri C, Moro M, Facchinetti F, Andriani F, et
al: Circulating mir-320a promotes immunosuppressive macrophages M2
phenotype associated with lung cancer risk. Int J Cancer.
144:2746–2761. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
ENCODE Project Consortium. The ENCODE
(ENCyclopedia of DNA elements) project. Science. 306:636–340.
2004.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015.PubMed/NCBI View
Article : Google Scholar
|
|
11
|
Wu Y, Shao A, Wang L, Hu K, Yu C, Pan C
and Zhang S: The role of lncRNAs in the distant metastasis of
breast cancer. Front Oncol. 9(407)2019.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang Y and Tang L: The application of
lncRNAs in cancer treatment and diagnosis. Recent Pat Anticancer
Drug Discov. 13:292–301. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Lu T, Wang Y, Di Chen JL and Jiao W:
Potential clinical application of lncRNAs in non-small cell lung
cancer. Onco Targets Ther. 11:8045–8052. 2018.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Engle L, Simpson C and Landers J: Using
high-throughput SNP technologies to study cancer. Oncogene.
25:1594–1601. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Syvänen AC: Toward genome-wide SNP
genotyping. Nat Genet. 37 (Suppl):S5–S10. 2005.PubMed/NCBI View
Article : Google Scholar
|
|
16
|
Andrew AS, Gui J, Sanderson AC, Mason RA,
Morlock EV, Schned AR, Kelsey KT, Marsit CJ, Moore JH and Karagas
MR: Bladder cancer SNP panel predicts susceptibility and survival.
Hum Genet. 125:527–539. 2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Duboule D: The rise and fall of Hox gene
clusters. Development. 134:2549–2560. 2007.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Rinn JL, Kertesz M, Wang JK, Squazzo SL,
Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E and
Chang HY: Functional demarcation of active and silent chromatin
domains in human HOX loci by noncoding RNAs. Cell. 129:1311–1323.
2007.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hajjari M and Salavaty A: HOTAIR: an
oncogenic long non-coding RNA in different cancers. Cancer Biol
Med. 12:1–9. 2015.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Loewen G, Jayawickramarajah J, Zhuo Y and
Shan B: Functions of lncRNA HOTAIR in lung cancer. J Hematol Oncol.
7(90)2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
22
|
Somarowthu S, Legiewicz M, Chillón I,
Marcia M, Liu F and Pyle AM: HOTAIR forms an intricate and modular
secondary structure. Mol Cell. 58:353–361. 2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Bhan A and Mandal SS: LncRNA HOTAIR: A
master regulator of chromatin dynamics and cancer. Biochim Biophys
Acta. 1856:151–164. 2015.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhuang Y, Wang X, Nguyen HT, Zhuo Y, Cui
X, Fewell C, Flemington EK and Shan B: Induction of long intergenic
non-coding RNA HOTAIR in lung cancer cells by type I collagen. J
Hematol Oncol. 6(35)2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Gupta RA, Shah N, Wang KC, Kim J, Horlings
HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, et al: Long
non-coding RNA HOTAIR reprograms chromatin state to promote cancer
metastasis. Nature. 464:1071–1076. 2010.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Wu Y, Zhang L, Wang Y, Li H, Ren X, Wei F,
Yu W, Wang X, Zhang L, Yu J and Hao X: Long noncoding RNA HOTAIR
involvement in cancer. Tumour Biol. 35:9531–9538. 2014.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Xu ZY, Yu QM, Du YA, Yang LT, Dong RZ,
Huang L, Yu PF and Cheng XD: Knockdown of long non-coding RNA
HOTAIR suppresses tumor invasion and reverses
epithelial-mesenchymal transition in gastric cancer. Int J Biol
Sci. 9:587–597. 2013.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Liu XH, Liu ZL, Sun M, Liu J, Wang ZX and
De W: The long non-coding RNA HOTAIR indicates a poor prognosis and
promotes metastasis in non-small cell lung cancer. BMC Cancer.
13(464)2013.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Zhao W, An Y, Liang Y and Xie XW: Role of
HOTAIR long noncoding RNA in metastatic progression of lung cancer.
Eur Rev Med Pharmacol Sci. 18:1930–1936. 2014.PubMed/NCBI
|
|
30
|
Nakagawa T, Endo H, Yokoyama M, Abe J,
Tamai K, Tanaka N, Sato I, Takahashi S, Kondo T and Satoh K: Large
noncoding RNA HOTAIR enhances aggressive biological behavior and is
associated with short disease-free survival in human non-small cell
lung cancer. Biochem Biophys Res Commun. 436:319–324.
2013.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Moher D, Liberati A, Tetzlaff J and Altman
DG: PRISMA Group. Preferred reporting items for systematic reviews
and meta-analyses: The PRISMA statement. PLoS Med.
6(e1000097)2009.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Stang A: Critical evaluation of the
Newcastle-Ottawa scale for the assessment of the quality of
nonrandomized studies in meta-analyses. Eur J Epidemiol.
25:603–605. 2010.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ren MM, Xu S, Wei YB, Yang JJ, Yang YN,
Sun SS, Li YJ, Wang PY and Xie SY: Roles of HOTAIR in lung cancer
susceptibility and prognosis. Mol Genet Genomic Med.
8(e1299)2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Minn AKK, Sato N, Mieno MN, Arai T and
Muramatsu M: Association study of long non-coding RNA HOTAIR
rs920778 polymorphism with the risk of cancer in an elderly
Japanese population. Gene. 729(144263)2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wang C, Li Y, Li YW, Zhang HB, Gong H,
Yuan Y, Li WT, Liu HY and Chen J: HOTAIR lncRNA SNPs rs920778 and
rs1899663 are associated with smoking, male gender, and squamous
cell carcinoma in a Chinese lung cancer population. Acta Pharmacol
Sin. 39:1797–1803. 2018.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Li H, Yang Z, Li J, Lv X, Gao M, Bi Y,
Zhang Z, Wang S, Li S, Li N, et al: Genetic variants in lncRNA
HOTAIR are associated with lung cancer susceptibility in a Chinese
Han population in China: A case-control study. Cancer Manag Res.
10:5209–5218. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Dadaş E and Aydin M: Effect of HOTAIR
rs12826786 and rs1899663 polymorphisms on lung cancer
susceptibility and clinicopathological characteristics in a Turkish
population: A hospital-based case-control study. Cell Mol Biol
(Noisy-le-grand). 64:97–102. 2018.
|
|
38
|
Gong WJ, Yin JY, Li XP, Fang C, Xiao D,
Zhang W, Zhou HH, Li X and Liu ZQ: Association of
well-characterized lung cancer lncRNA polymorphisms with lung
cancer susceptibility and platinum-based chemotherapy response.
Tumour Biol. 37:8349–8358. 2016.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Dor Y and Cedar H: Principles of DNA
methylation and their implications for biology and medicine.
Lancet. 392:777–786. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Shimizu Y, Sato S, Noguchi Y, Koyamatsu J,
Yamanashi H, Higashi M, Nagayoshi M, Kadota K, Kawashiri SY, Nagata
Y, et al: Impact of single nucleotide polymorphism on short stature
and reduced tongue pressure among community-dwelling elderly
Japanese participants: A cross-sectional study. Environ Health Prev
Med. 22(62)2017.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Leaché AD and Oaks JR: The utility of
single nucleotide polymorphism (SNP) data in phylogenetics. Annu
Rev Ecol Evol Syst. 48:69–84. 2017.
|
|
42
|
Zhang X, Zhou L, Fu G, Sun F, Shi J, Wei
J, Lu C, Zhou C, Yuan Q and Yang M: The identification of an ESCC
susceptibility SNP rs920778 that regulates the expression of lncRNA
HOTAIR via a novel intronic enhancer. Carcinogenesis. 35:2062–2067.
2014.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zhu H, Lv Z, An C, Shi M, Pan W, Zhou L,
Yang W and Yang M: Onco-lncRNA HOTAIR and its functional genetic
variants in papillary thyroid carcinoma. Sci Rep.
6(31969)2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Pan W, Liu L, Wei J, Ge Y, Zhang J, Chen
H, Zhou L, Yuan Q, Zhou C and Yang M: A functional lncRNA HOTAIR
genetic variant contributes to gastric cancer susceptibility. Mol
Carcinog. 55:90–96. 2016.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Liu Z, Sun M, Lu K, Liu J, Zhang M, Wu W,
De W, Wang Z and Wang R: The long noncoding RNA HOTAIR contributes
to cisplatin resistance of human lung adenocarcinoma cells via
downregualtion of p21(WAF1/CIP1) expression. PLoS One.
8(e77293)2013.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Fan L, Lei H, Lin Y, Zhou Z, Li J, Wu A,
Shu G, Ruger S and Yin G: Hotair promotes the migration and
proliferation in ovarian cancer by miR-222-3p/CDK19 axis. Cell Mol
Life Sci. 79(254)2022.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Chen J, Hou SF, Tang FJ, Liu DS, Chen ZZ,
Zhang HL and Wang SH: HOTAIR/Sp1/miR-199a critically regulates
cancer stemness and malignant progression of cutaneous squamous
cell carcinoma. Oncogene. 41:99–111. 2022.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Zhang W, Wu Q, Liu Y, Wang X, Ma C and Zhu
W: LncRNA HOTAIR promotes chemoresistance by facilitating
epithelial to mesenchymal transition through miR-29b/PTEN/PI3K
signaling in cervical cancer. Cells Tissues Organs. 211:16–29.
2022.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Zhang W, Liu J, Wu Q, Liu Y and Ma C:
HOTAIR contributes to stemness acquisition of cervical cancer
through regulating miR-203 interaction with ZEB1 on
epithelial-mesenchymal transition. J Oncol.
2021(4190764)2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Zhang M, Wu K, Zhang P, Qiu Y, Bai F and
Chen H: HOTAIR facilitates endocrine resistance in breast cancer
through ESR1/miR-130b-3p axis: Comprehensive analysis of
mRNA-miRNA-lncRNA network. Int J Gen Med. 14:4653–4663.
2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Deng S, Wang J, Zhang L, Li J and Jin Y:
LncRNA HOTAIR promotes cancer stem-like cells properties by
sponging miR-34a to activate the JAK2/STAT3 pathway in pancreatic
ductal adenocarcinoma. Onco Targets Ther. 14:1883–1893.
2021.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Sun N, Zhang W, Liu J, Yang X and Chu Q:
Propofol inhibits the progression of cervical cancer by regulating
HOTAIR/miR-129-5p/RPL14 axis. Onco Targets Ther. 14:551–564.
2021.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Wu D, Zhu J, Fu Y, Li C and Wu B: LncRNA
HOTAIR promotes breast cancer progression through regulating the
miR-129-5p/FZD7 axis. Cancer Biomark. 30:203–212. 2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Zhan Y, Abuduwaili K, Wang X, Shen Y,
Nuerlan S and Liu C: Knockdown of long non-coding RNA HOTAIR
suppresses cisplatin resistance, cell proliferation, migration and
invasion of DDP-resistant NSCLC cells by targeting
miR-149-5p/doublecortin-like kinase 1 axis. Cancer Manag Res.
12:7725–7737. 2020.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Liang H and Peng J: LncRNA HOTAIR promotes
proliferation, invasion and migration in NSCLC cells via the CCL22
signaling pathway. PLoS One. 17(e0263997)2022.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Tang Y, Song G, Liu H, Yang S, Yu X and
Shi L: Silencing of long non-coding RNA HOTAIR alleviates
epithelial-mesenchymal transition in pancreatic cancer via the
Wnt/β-catenin signaling pathway. Cancer Manag Res. 13:3247–3257.
2021.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Dai ZY, Jin SM, Luo HQ, Leng HL and Fang
JD: LncRNA HOTAIR regulates anoikis-resistance capacity and
spheroid formation of ovarian cancer cells by recruiting EZH2 and
influencing H3K27 methylation. Neoplasma. 68:509–518.
2021.PubMed/NCBI View Article : Google Scholar
|