Introduction
As tubulointerstitial fibrosis is a common endpoint
in renal disease with no effective treatment other than dialysis,
the need to understand the molecules and mechanisms involved is
increasingly urgent. Histologically, tubulointerstitial fibrosis is
an accumulation of the extracellular matrix (ECM) in the
interstitium. ECM-producing cells are primarily activated
fibroblasts (1). Various cells
such as pericytes, endothelial cells, residual fibroblasts and
tubular epithelial cells are known to be the origin of fibroblasts
(2).
The epithelial-mesenchymal transformation (EMT) has
been studied in cancer and benign fibrotic diseases (3). Once acute injury is imposed on the
kidney, various chemokines and growth factors cause inflammation,
which in turn leads to the secretion of transforming growth
factor-β (TGF-β) via release of active TGF-β from latent
TGF-β-binding protein via protease cleavage (4). TGF-β is the primary molecule
responsible for EMT (5,6), and the canonical and non-canonical
pathways are the downstream pathways of TGF-β (4,7). The
hallmark of EMT is loss of epithelial phenotypes and acquisition of
mesenchymal phenotypes with activation of profibrotic genes to
produce the ECM, including fibronectin and collagen types I and III
(3,5,8).
Lysyl oxidase-like 2 (LOXL2) is a member of the
lysyl oxidase family, originally known as a copper-dependent amine
oxidase, that is involved in cross-linking collagen and elastin of
the ECM (9). Studies have also
demonstrated additional functions for LOXL2 independent of its
catalytic activity, such as organ development (10), tumour invasion (11) and EMT (12,13).
In mice lung fibroblast cells, LOXL2 has been
revealed to play prominent roles for fibrogenesis via regulation of
the TGF-β/Smad signaling pathway (14). LOXL2 has been also identified to be
important in promoting both glomerular and interstitial
pathogenesis associated with Alport syndrome in mice (15). In a previous study conducted by the
authors, it was found that LOXL2 is expressed in kidney podocytes
and tubular epithelial cells, and its expression is increased in
the folic acid-induced murine fibrosis model (16). In the present study, the effect and
therapeutic role of LOXL2 inhibitor AB0023 on the progression of
tubulointerstitial fibrosis in mice was evaluated. In order to
evaluate the contribution of LOXL2 in EMT, an in vitro study
using immortalized human proximal tubular epithelial cells (HK-2
cells) was also performed.
Materials and methods
Animal model of tubulointerstitial
fibrosis and LOXL2 inhibition
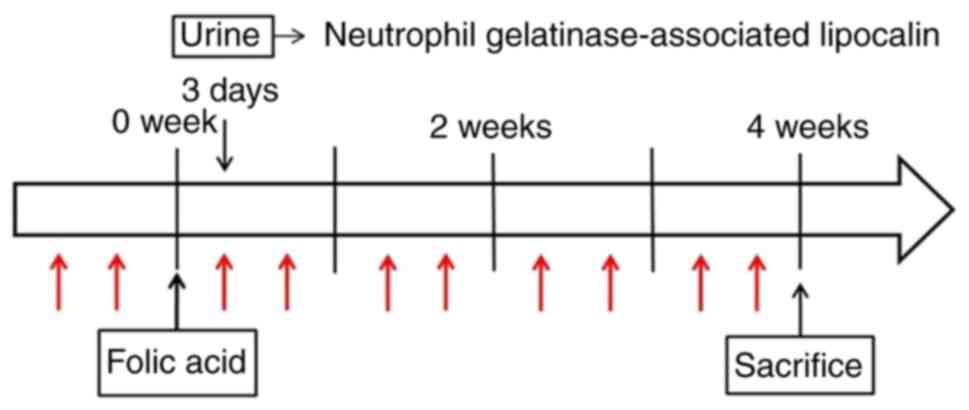
Male CD1 mice at 8 weeks of age (Orient Bio, Inc.)
were used in the present study. The animals were housed in a
facility maintained at 20˚C and 12-h alternating light/dark cycles
with free access to rodent chow and water. Tubulointerstitial
fibrosis was induced by intraperitoneal injection of folic acid
(240 µg/g body weight) (17,18).
The folic acid solution was prepared by dissolving folic acid
powder (Sigma-Aldrich; Merck KGaA) in 0.3 M NaHCO3.
Control CD1 mice were injected intraperitoneally with the same
volume of vehicle (NaHCO3). Urinary excretion of
neutrophil gelatinase-associated lipocalin (NGAL) (19) was measured immediately before
injection and at 3 days after injection using a Mouse
Lipocalin-2/NGAL Quantikine ELISA kit (cat. no. MLCN20; R&D
Systems, Inc.) to ensure successful injection of folic acid, as
manifested by a log scale increase in NGAL. The concentration of
urinary NGAL was normalized by the concentration of urinary
creatinine as measured by the QuantiChrom Creatinine Assay kit
(BioAssay Systems). Mice without an increase in NGAL at 3 days
post-folic acid injection, indicating that folic acid was not
successfully injected, were omitted from the study. Ultimately, 16
mice injected with folic acid and six mice injected with vehicle
were examined in the present study. The present study was approved
(approval no. 2015-0247) by the Institutional Animal Care and Use
Committee of the Yonsei University Health System (Seoul, Republic
of Korea). All experiments involving animals were carried out in
accordance with the standards set forth by the Institutional Animal
Care and Use Committee of Yonsei University Health System.
To inhibit LOXL2, AB0023 (cat. no. GS-607601; Gilead
Sciences, Inc.), an inhibitory monoclonal antibody against LOXL2,
was used. A total of 9 of the 16 folic acid-injected mice were
treated with a dosage of 15 mg/kg body weight of AB0023 at 1 week
before folic acid injection and twice a week for 4 weeks
afterwards. The dosage was chosen according to the manufacturer's
guideline, supported by a reference study using the same
intraperitoneal dosage (20).
Adverse effect was not observed during the experiment. The
remaining seven mice were injected with immunoglobulin G (IgG)
(cat. no. GS-645864; Gilead Sciences, Inc.) at the same dosage and
on the same time schedule as the AB0023 treatment group. Mice were
sacrificed via cervical dislocation 4 weeks after folic acid or
vehicle injection, and the right kidneys were harvested (Fig. 1). Fresh frozen tissues were stored
at -70˚C for western blot analysis and collagen measurement.
Additional kidney tissues were fixed in 4% formaldehyde for 24 h at
room temperature and embedded in paraffin overnight at 55-65˚C
using an automatic tissue processer (EFTP-FAST 360; Intelsint
S.R.L.).
Evaluation of tubulointerstitial
fibrosis Semiquantitative analysis via histologic examination
Paraffin-embedded samples of the AB0023-treated,
IgG-injected, and vehicle-injected groups were cut into 4-µm
sections. Sections were deparaffinized by submersing slides in
xylene and dehydrated in 2 changes of absolute alcohol for 2 min, 1
change of 95% alcohol for 2 min, and 1 change of 70% alcohol for 2
min. The sections were stained with Masson trichrome and
picro-sirius red. For Masson trichrome, sections were treated with
Weigert's iron hematoxylin for 8 min to stain nuclei, Biebrich
scarlet-acid fuchsin for 15 min, phosphomolybdic-phosphotungstic
acid for 15 min and aniline blue for 15 min at room temperature.
For picro-sirius red staining, sections were treated with Weigert's
Iron Hematoxylin for 8 min to stain nuclei and Direct Red 80
(Sigma-Aldrich; Merck KGaA) for 1 h at room temperature to
visualize collagen before washing with 0.5% glacial acid. Slides
were examined by light microscopy with or without polarization for
picro-sirius red or trichrome staining, respectively. Images were
captured serially along the cortex at x200 magnification and the
area of interstitial fibrosis was measured using ImageJ software
(version 1.50i; National Institutes of Health).
Quantitative analysis via total
collagen analysis
The content of collagen in fresh frozen cortex was
evaluated by measuring hydroxyproline using the Total Collagen
Assay kit (QuickZyme Biosciences) according to the manufacturer's
guide. Briefly, samples were hydrolysed at 95˚C in 6 M HCl for 20 h
and then centrifuged at 13,000 x g for 10 min at room temperature.
The supernatant was collected and assayed by ELISA according to the
manufacturer's protocol. Total protein in the hydrolysed sample was
also measured using the Total Protein Assay kit (QuickZyme
Biosciences) and the relative amount of collagen per protein was
analysed.
Renal cell culture and
transfection
HK-2 cells were purchased from the American Type
Culture Collection and cultured in Dulbecco's modified Eagle's
medium/Nutrient Mixture F-12 (Gibco; Thermo Fisher Scientific,
Inc.) supplemented with 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.). To silence LOXL2 expression at the
cellular level, LOXL2 small hairpin (sh)RNA lentiviral particles
(cat. no. sc-45222-v; Santa Cruz Biotechnology, Inc.) were
transduced into HK-2 cells at room temperature cultured on collagen
I (2 mg/ml; cat. no. 354236; Corning Inc.,)-coated dishes (21). HK-2 cells were treated with media
containing 5 µg/ml of polybrene (cat. no. sc-134220; Santa Cruz
Biotechnology, Inc.), and then LOXL2 shRNA lentiviral particles and
control shRNA lentiviral particles (cat. no. sc-108080; Santa Cruz
Biotechnology, Inc.) of 1 and 2 multiplicity of infection (MOI)
were added and incubated overnight at 37˚C. Transfected cells were
selected by selection media containing 2 µg/ml puromycin
dihydrochloride (cat. no. sc-108071; Santa Cruz Biotechnology,
Inc.). After confirming the decrease in LOXL2 expression by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analysis, cells of 2 MOI were treated with serum-free
media for 24 h and then media containing TGF-β (20 ng/ml; R&D
Systems, Inc.) for 72 h. Other LOXL2-deficient cells (2 MOI)
were treated with serum-free media for 24 h and then media
containing vehicle (0.1% 4 mM HCl/BSA). Control shRNA lentiviral
particles were transduced into another line of HK-2 cells in the
same manner as that of LOXL2 shRNA particles and were further
incubated with media containing either TGF-β (20 ng/ml; R&D
Systems, Inc.) or vehicle (0.1% 4 mM HCl/BSA) after 24 h of serum
starvation.
RT-qPCR
Total RNA of transfected cells was extracted using
RNeasy kit (cat. no. 74104; Qiagen, Inc.). RNA was reverse
transcribed to cDNA using the Qiagen Quantitect Reverse
Transcription kit (cat. no. 205311; Qiagen, Inc.) in 20 µl reaction
volumes containing 1 µg of RNA. The reaction was carried out at
42˚C for 2 min, 42˚C for 15 min and held at 95˚C for 3 min. The
cDNA products were diluted at 1:5 to contain 30 ng of cDNA in 3 µl
for use in qPCR. qPCR was performed using a TaqMan Gene Expression
Master Mix assay (cat. no. 4369016; Applied Biosystems; Thermo
Fisher Scientific, Inc.), with 18S ribosomal RNA gene (cat. no.
Mm03928990_g1; Applied Biosystems; Thermo Fisher Scientific, Inc.)
as reference gene and human LOXL2 (cat. no. Hs00158757_m1; Applied
Biosystems; Thermo Fisher Scientific, Inc.) probe according to the
manufacturer's protocol. As the manufacturer does not provide the
primer sequences of human LOXL2 and 18S ribosomal RNA gene, it has
not been possible to provide the information. The qPCR reactions
were carried out according to the product instruction (50˚C for 2
min, 95˚C for 10 min, and then 40 cycles of 95˚C for 10 sec, 60˚C 1
min). All data were analyzed using the 2-ΔΔCq method
(22).
Western blot analysis for LOXL2,
Smad-related molecules and EMT-related molecules
Fresh frozen kidney tissues from mice, injected with
AB0023 or control IgG followed by folic acid or injected only with
vehicle, were homogenised and western blotting was performed
following the same protocol as previously described by the authors
(16). Radioimmunoprecipitation
assay buffer (Biosesang) with a protease inhibitor cocktail (Roche
Diagnostics) was prepared. The samples were centrifuged at 13,000 x
g for 30 min at 4˚C. The protein concentration was measured through
bicinchoninic acid protein assay kit (Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. When the protein
samples (50 µg) were separated by 10% sodium dodecyl
sulphate-polyacrylamide gel electrophoresis for 2 h at 100 V, they
were transferred onto a polyvinylidene fluoride membrane and
blocked with 3% skim milk for 1 h at room temperature. Primary
antibodies were incubated with the membrane overnight at 4˚C. The
anti-mouse-specific primary antibodies purchased from Cell
Signaling Technology, Inc. included anti-Smad2 (1:1,000; cat. no.
5339), anti-phosphorylated (p)-Smad2 (Ser465/467; 1:500; cat. no.
3108), anti-Smad3 (1:1,000; cat. no. 9523), anti-p-Smad3
(Ser423/425; 1:1,000; cat. no. 9520) and anti-Smad4 (1:1,000; cat.
no. 38454). The blocking solution used for the anti-p-Smad2 and
anti-p-Smad3 antibodies contained 5% bovine serum albumin
(Sigma-Aldrich; Merck KGaA). The membrane was washed with
Tris-buffered saline containing 0.1% Tween-20. It was then
incubated with horseradish peroxidase-labelled secondary antibodies
(1:5,000; cat. no. sc-2020; Santa Cruz Biotechnology, Inc. and
1:5,000; cat. no. K4003; Dako; Agilent Technologies, Inc.) for 1 h
at room temperature. Pierce Enhanced Chemiluminescence Western
Blotting Substrate (Thermo Fisher Scientific, Inc.) was used to
visualize protein bands. The membrane was stripped with Restore
Western Blot Stripping Buffer (Thermo Fisher Scientific, Inc.) for
15 min at room temperature, and then it was incubated with an
anti-β-actin antibody (1:2,000; cat. no. sc-47778; Santa Cruz
Biotechnology, Inc.). The bands were semi-quantified by
densitometry using ImageJ software (version 1.50i; National
Institutes of Health).
In addition, HK-2 cells with LOXL2 shRNA or control
transfection after TGF-β challenge or incubated with vehicle were
lysed in buffer and western blotting was performed in similar
manner. Due to their high molecular weight, 6%-acrylamide gel was
used for fibronectin, ZO-1 and E-cadherin. Except for acrylamide
percentage, all the membranes were electrophoresed under identical
experimental conditions. The primary antibodies applied to HK-2
cells were anti-vimentin (1:5,000; cat. no. ab92547; Abcam),
anti-E-cadherin (1:500; cat. no. 610181; BD Biosciences),
anti-zonula occludens (ZO)-1 (1:500; cat. no. ab2272;
Sigma-Aldrich; Merck KGaA), anti-fibronectin (1:1,000; cat. no.
sc8422; Santa Cruz Biotechnology, Inc.) and anti-LOXL2 (1:500; cat.
no. ab96233; Abcam).
Statistical analysis
Quantitative analysis was performed for the western
blotting and RT-qPCR results. Vehicle-injected mice, folic
acid-injected mice treated with AB0023, and folic acid-injected
mice treated with control IgG were compared. For comparing
fibrosis, Kruskal-Wallis test followed by Dunn's post hoc tests
with Bonferroni corrections was performed. When comparing the folic
acid injected groups treated with either AB0023 or control IgG,
Mann-Whitney U test was performed. For comparing the Smads levels,
Mann-Whitney U test was used. In addition, HK-2 cells with or
without LOXL2 inhibition were analyzed by Kruskal-Wallis test
followed by Dunn's post hoc tests with Bonferroni corrections and
Mann-Whitney U test. Data are expressed as the mean ± standard
deviation. The analyses were performed using SPSS version 25
software (IBM Corp.). P<0.05 was considered to indicate a
statistically significant difference.
Results
LOXL2 inhibition prevents the
progression of tubulointerstitial fibrosis in the mouse model
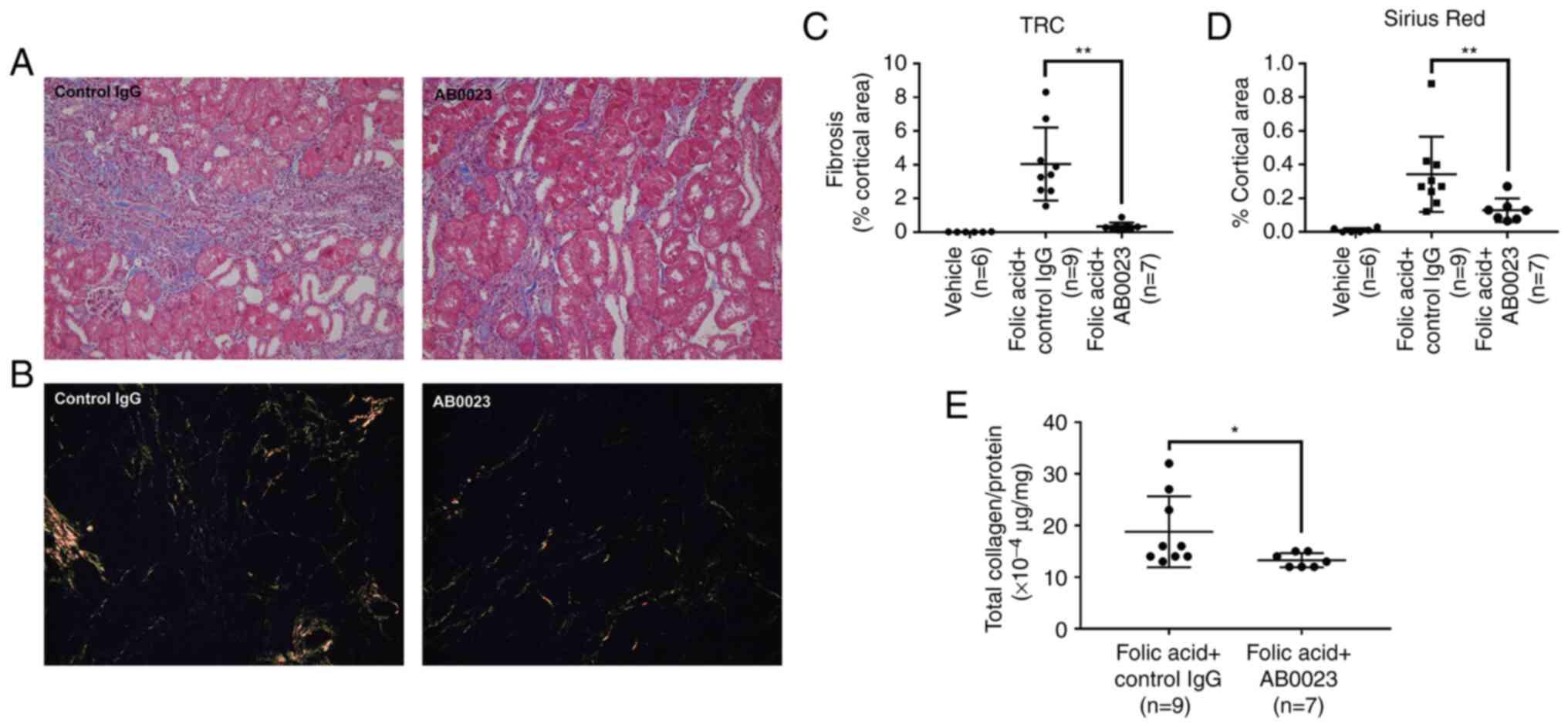
The amount of fibrosis measured by trichrome
(Fig. 2A) and picro-sirius red
staining (Fig. 2B) decreased in
mice treated with AB0023, compared with the control IgG-treated
group (Fig. 2C and D). Quantitative measurement of fibrosis
by total collagen analysis also showed that fibrosis decreased in
mice treated with AB0023 (Fig.
2E).
LOXL2 inhibition may influence the
canonical TGF-β/Smad signalling pathway
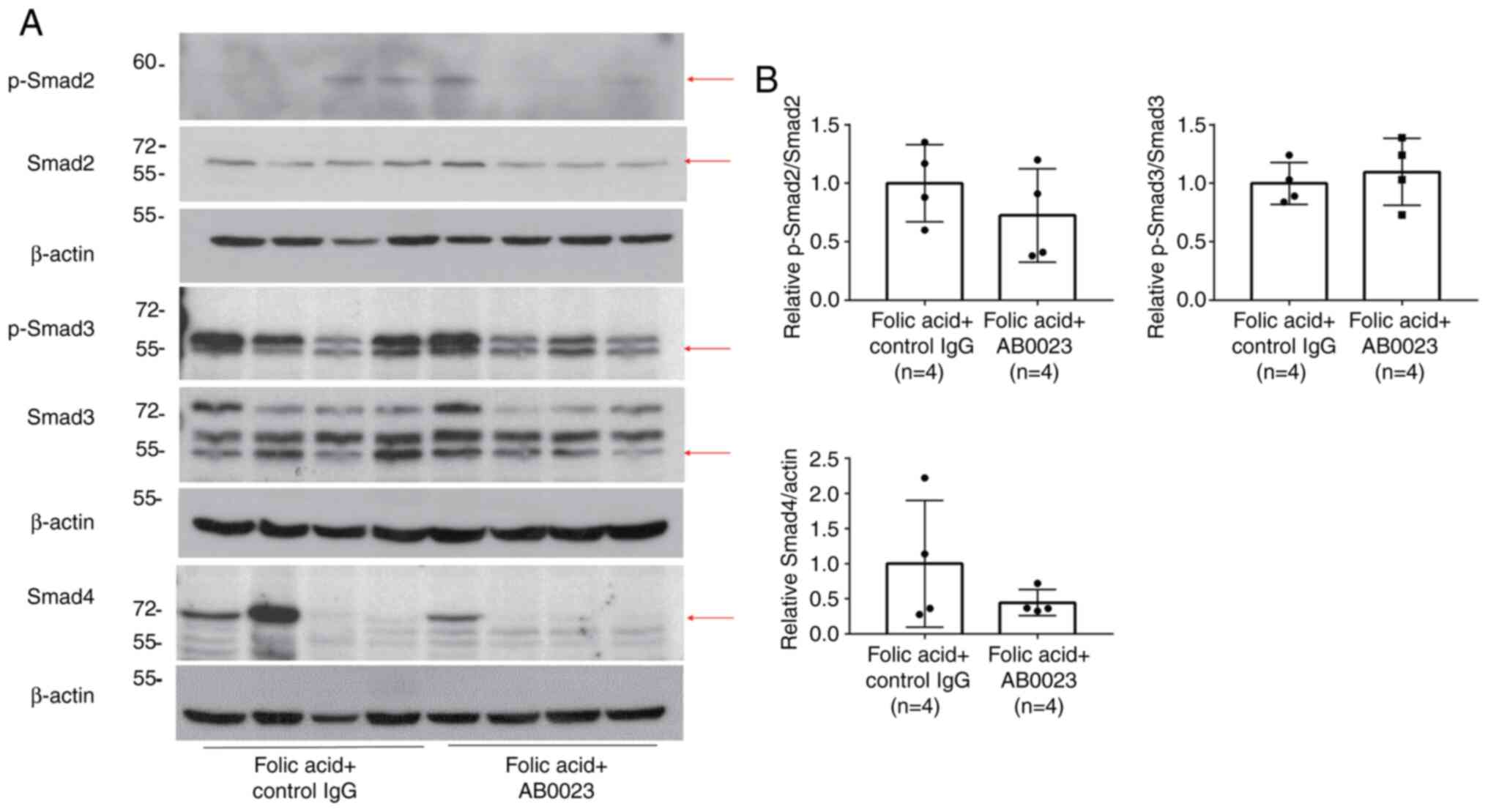
Smad signaling pathway molecules, including p-Smad3,
p-Smad2, and Smad4 exhibited no significant difference with LOXL2
inhibition (Fig. 3). However, the
amounts of p-Smad2 and Smad4 tended to decrease in the
AB0023-treated group compared with the control group.
LOXL2 knockdown in HK-2 cells reduces
the expression of some EMT-associated molecules
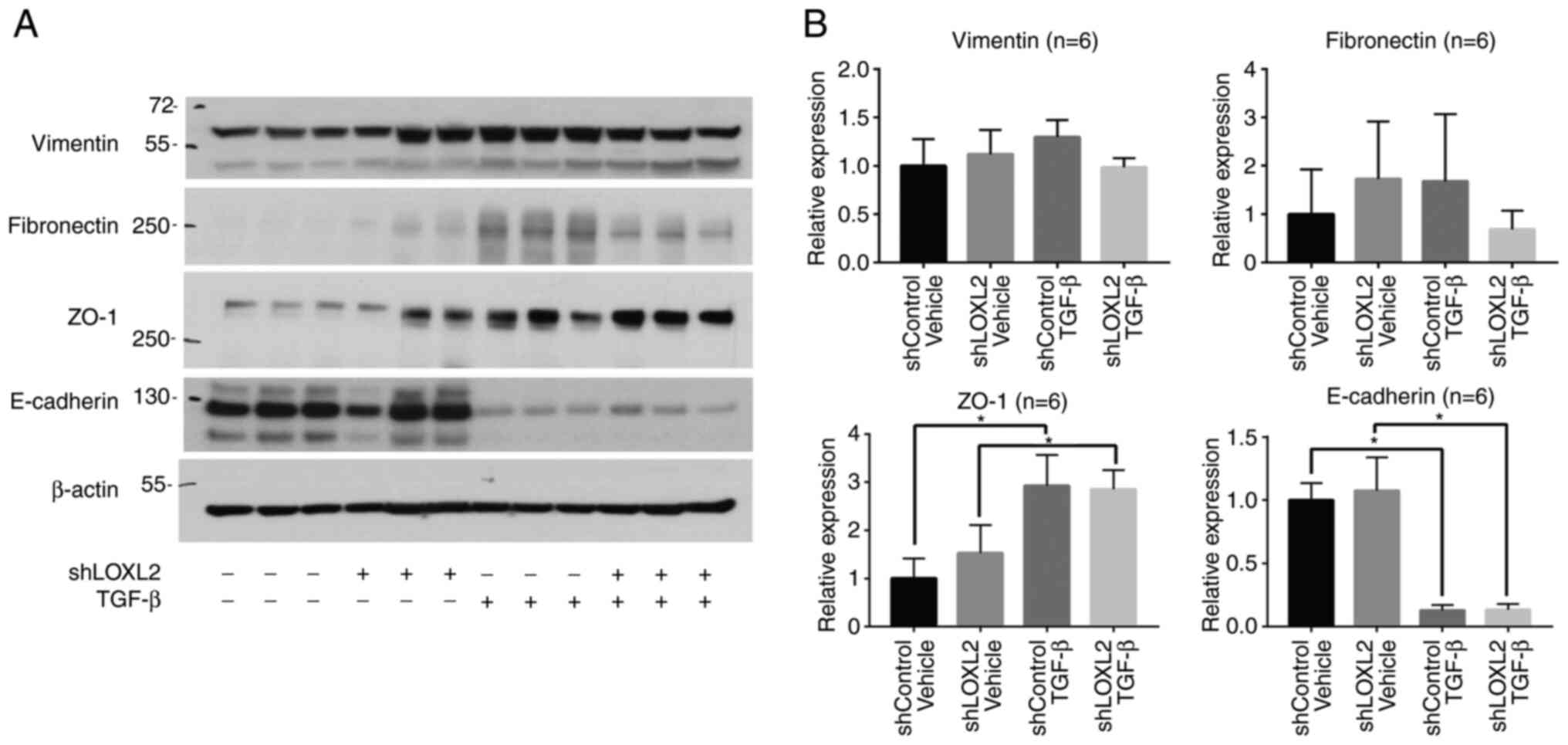
Transfection of HK-2 cells with LOXL2 shRNA resulted
in LOXL2 knockdown (Fig. S1). In
control HK-2 cells transfected with control shRNA, TGF-β treatment
(72 h) reduced the levels of epithelial marker E-cadherin, and
increased the levels of mesenchymal markers vimentin and
fibronectin. Multiple comparison analysis revealed the decreasing
trends of vimentin and fibronectin level in LOXL2 knockdown cells
compared with the control shRNA-transduced cells after TGF-β
treatment (Fig. 4). Considering
that the level of vimentin and fibronectin increases after TGF-β
challenge in control cells, the decreasing trend of those in LOXL2
knockdown cells after TGF-β challenge is more meaningful. The
epithelial markers ZO-1 and E-cadherin did not show a significant
difference after TGF-β treatment in LOXL2 knockdown and control
cells, while the level of E-cadherin was markedly decreased by
TGF-β challenge compared with the shControl Vehicle.
Discussion
Inhibition of LOXL2 via AB0023 has been revealed to
reduce fibrosis in various organs. For instance, AB0023 attenuated
postoperative fibrosis in a rabbit model of glaucoma surgery
(23). AB0023 also attenuated
tetrachloride-induced hepatic fibrosis in BALB/c mice, and
high-dose bleomycin-induced pulmonary fibrosis in C57BL/6 mice
(20). Although a clinical trial
of simtuzumab, a humanized form of AB0023, resulted in no
significant changes in fibrosis in human immunodeficiency virus-
and hepatitis C virus-infected adults, serum samples suggested
upregulation of TGF-β3 and interleukin-10 pathways with treatment,
suggesting the future evaluation for clinical trials with
simtuzumab after the modulation of TGF-β3(24). It was previously observed that
LOXL2 is expressed in tubular epithelial cells, and a role for
LOXL2 in TGF-β-mediated tubulointerstitial fibrosis was presumed.
This hypothesis is supported by the increased LOXL2 mRNA and
protein levels detected in the kidneys of mice with folic
acid-induced tubulointerstitial fibrosis (11). Intraperitoneal folic acid injection
was used to CD1 mice model in both the previous (16) and the present study. CD1 mice were
selected since it has been identified that this strain is
susceptible to folic acid-induced kidney injury (25,26),
and this strain showed the least mortality after folic acid
injection according to our experience. The positive regulatory role
of LOXL2 in tubulointerstitial fibrosis was confirmed by the
reduction of fibrosis in this folic acid-induced fibrosis mouse
model with the inhibition of LOXL2 by the LOXL2-specific antibody
AB0023. Recently, Nguyen et al (27) revealed that LOXL2 inhibition leads
to reduction of renal fibrosis in murine kidney injury induced by
cyclosporine-A. The present study was different in two points.
First, folic acid administration induces both acute kidney injury
and then chronic kidney disease, a course more natural and similar
to how human renal fibrosis occurs. Second, the effects of LOXL2
inhibition were examined in not only in murine models but also in
HK-2 cells, a proximal tubular cell line driven from human
kidney.
In addition to fibrosis, TGF-β is involved in
various biological activities, such as cell proliferation,
apoptosis, differentiation, autophagy and the immune response
(28). Thus, it is critical to
investigate therapeutic strategies related to the downstream
pathways of TGF-β due to the possible adverse effects of directly
targeting this cytokine (29).
EMT is a major mechanism that contributes to renal
fibrosis in response to multiple molecules, including
TGF-β1(1), connective tissue
growth factor (CTGF) (30), and
angiotensin II (11), in tubular
epithelial cells. Fibroblasts arising from tubular epithelial cells
(27,31) through EMT express CTGF, a
fibrogenic cytokine and a downstream mediator of the TGF-β1 pathway
in renal fibrosis, although research nowadays has put more
importance on resident interstitial fibroblasts (32,33).
CTGF itself is also known to induce EMT in the kidney. In the
kidney proximal tubule cells, TGF-β1 induces ECM proteins such as
fibronectin and collagen IV via CTGF-dependent and -independent
pathways (34). Finally, the ECM
turnover is imbalanced, resulting in ECM accumulation and renal
fibrosis. Although the in vivo role of EMT has previously
been questioned (35), its
relationship with renal fibrosis remains to be valuable.
Among multiple molecules related to EMT, TGF-β1 is
the most potent inducer of EMT (5,8). As
aforementioned, EMT is a well-described process characterized by a
loss of epithelial cell adhesion molecules, such as E-cadherin and
ZO-1, de novo α-SMA expression and actin filament
reorganization, transformation of myofibroblastic morphology,
tubular basement membrane disruption and cell
migration/infiltration into the interstitium (5,8).
However, conflicting results have been reported from in vivo
studies as tubular cells that have undergone partial EMT relay
proinflammatory and profibrogenic signals to the interstitium
without directly contributing to the myofibroblast population
(36). Accordingly, the
relationship between LOXL2 and the TGF-β pathway in vivo,
and the TGF-β-mediated relationship between LOXL2 and EMT in
vitro, were investigated in the present study. Thus, canonical
pathway-related molecules were studied in vivo and markers
expressed by tubular cells during EMT were studied in
vitro.
The lack of significant differences in the level of
Smad molecules after LOXL2 inhibition in the present study may be
due to the lapse of time between folic acid injection and analysis.
Murine kidneys were harvested at 4 weeks after this injury, by
which time fibrogenesis could have already been completed. Stallons
et al (18) reported that
TGF-β1 and α-SMA levels increased until 6 days after folic acid
injection, and gradually decreased afterwards in a similar
experiment where a 250 mg/kg dose of folic acid was injected
intraperitoneally. Tang et al (37) reported that after the
administration of high glucose doses, the expression of p-Smad2 and
p-Smad3 increased in HK-2 cells for 30 to 60 min and 30 to 120 min,
respectively, before decreasing gradually. The present study
differed from this experiment; in particular, the time between
intervention and injury was substantially longer than that in
previous studies. A more rapid analysis of Smad molecules after
folic acid injection may have revealed a more pronounced change in
their expression in the present study.
Although statistically less significant, the
decreasing trends of the effect of AB0023 on the expression levels
of pSmad2 and Smad4 are similar to those published by Wen et
al (14), demonstrating the
possible association of LOXL2 and canonical pathway. It was
hypothesized that LOXL2 may act on downstream of the TGF-β pathway
or LOXL2 and TGF-β may interact indirectly (14). In line with the aforementioned
study, the decreasing tendency of pSmad2 and Smad4 expression after
LOXL2 inhibition, reinforcing the regulatory role of LOXL2 on
TGF-β/Smad signaling pathway, was also revealed in the present
study. EMT is located more downstream of the TGF-β pathway than
Smad molecules, and LOXL2 inhibition did not show significant
difference in EMT markers other than vimentin. This possibly
indicates more indirect relationship between LOXL2 and EMT compared
with Smad molecules. Therefore, it was suggested that LOXL2 may be
located upstream from the EMT but downstream of TGF-β, or LOXL2 may
regulate the TGF-β pathway indirectly. It was found that pSmad3
level tended to increase after AB0023 treatment, whereas Wen et
al (14) found pSmad2/3 level
decreased after LOXL2 RNA inhibition. The discrepancy may be
attributed to the difference in methods of LOXL2 inhibition: RNA
inhibition and inhibitory antibody often has different off-targets
and the power of inhibitory effect differs (38). Furthermore, the discrepancy may be
attributed to the different nature of the experiment. In
vivo experiment is complicated by various extracellular signals
including cell-to-cell interaction and cell-to-ECM interaction;
such unwanted effects are reduced in in vitro
experiment.
Experiments on HK-2 cells in vitro after
TGF-β challenge revealed no significant difference in the
myofibroblast marker nor epithelial marker in LOXL2 knockdown
cells. However, there was a decreasing trend in vimentin and
fibronectin. These data indicated that LOXL2 may play a regulatory
role in EMT. Other studies have shown no reduction in epithelial
markers, such as E-cadherin, with LOXL2 inhibition after TGF-β
challenge, while a significant reduction of E-cadherin was observed
in the present study. However, cell types and experimental methods
used in the present study differ from those previously reported
(12,13,39).
Although EMT markers level remained insignificant, it can be
inferred that LOXL2 may be related to EMT pathway at least
partially based on decreasing trends of vimentin and fibronectin.
Further studies are warranted to elucidate the mechanisms
underlying the LOXL2-EMT-related pathway, particularly
investigating the EMT markers after blocking the TGF-β/Smad
signaling pathway.
In conclusion, inhibition of LOXL2 ameliorates renal
fibrosis. LOXL2 may be associated with TGF-β-mediated
tubulointerstitial fibrosis and EMT. Improved understanding of the
role of LOXL2 in the kidney may illuminate the pathophysiology of
tubulointerstitial fibrosis and glomerulosclerosis, and potentially
lead to the discovery of novel therapeutic targets for treating
these conditions.
Supplementary Material
Transfection of HK-2 cells with LOXL2
shRNA results in LOXL2 knockdown. HK-2 cells were transfected with
either LOXL2 shRNA lentivirus (1 and 2 MOI) to knockdown LOXL2
expression, or control shRNA virus (1 and 2 MOI). Both the (A)
LOXL2 RNA and (B) protein were semi-quantified. Transfection of
LOXL2 shRNA resulted in LOXL2 knockdown, whereas transfection of
control shRNA did not. LOXL2, lysyl oxidase-like 2; sh-, short
hairpin; MOI, multiplicity of infection; shControl, HK-2 cells
transfected with control shRNA; shLOXL2, LOXL2-deficient HK-2 cells
by LOXL2 shRNA transfection.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Science, ICT and Future Planning
(grant no. NRF-2015R1C1A1A02036671).
Availability of data and materials
All data generated or analyzed during this study are
available from the corresponding author on reasonable request.
Authors' contributions
SEC performed data analysis and interpretation,
drafted and revised the manuscript. NJ performed experiments and
retrieved data. HYC and HJJ conceptualized the present study. BJL
contributed to the conceptualization of the present study, data
analysis, interpretation of data and revising the manuscript. All
authors read and approved the final version of the manuscript. SEC
and BJ Lim confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was approved (approval no.
2015-0247) by the Institutional Animal Care and Use Committee of
the Yonsei University Health System (Seoul, Republic of Korea). All
experiments involving animals were carried out in accordance with
the standards set forth by the Institutional Animal Care and Use
Committee of Yonsei University Health System.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Meran S and Steadman R: Fibroblasts and
myofibroblasts in renal fibrosis. Int J Exp Pathol. 92:158–167.
2011.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Liu Y: Cellular and molecular mechanisms
of renal fibrosis. Nat Rev Nephrol. 7:684–696. 2011.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014.PubMed/NCBI View
Article : Google Scholar
|
|
4
|
Meng XM, Nikolic-Paterson DJ and Lan HY:
TGF-β: The master regulator of fibrosis. Nat Rev Nephrol.
12:325–338. 2016.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yang J and Liu Y: Dissection of key events
in tubular epithelial to myofibroblast transition and its
implications in renal interstitial fibrosis. Am J Pathol.
159:1465–1475. 2001.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Friedman SL, Sheppard D, Duffield JS and
Violette S: Therapy for fibrotic diseases: Nearing the starting
line. Sci Transl Med. 5(167sr161)2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Sutariya B, Jhonsa D and Saraf MN: TGF-β:
The connecting link between nephropathy and fibrosis.
Immunopharmacol Immunotoxicol. 38:39–49. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Hills CE and Squires PE: The role of TGF-β
and epithelial-to mesenchymal transition in diabetic nephropathy.
Cytokine Growth Factor Rev. 22:131–139. 2011.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Nishioka T, Eustace A and West C: Lysyl
oxidase: From basic science to future cancer treatment. Cell Struct
Funct. 37:75–80. 2012.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Maki JM, Sormunen R, Lippo S,
Kaarteenaho-Wiik R, Soininen R and Myllyharju J: Lysyl oxidase is
essential for normal development and function of the respiratory
system and for the integrity of elastic and collagen fibers in
various tissues. Am J Pathol. 167:927–936. 2005.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Ahn SG, Dong SM, Oshima A, Kim WH, Lee HM,
Lee SA, Kwon SH, Lee JH, Lee JM, Jeong J, et al: LOXL2 expression
is associated with invasiveness and negatively influences survival
in breast cancer patients. Breast Cancer Res Treat. 141:89–99.
2013.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Cuevas EP, Moreno-Bueno G, Canesin G,
Santos V, Portillo F and Cano A: LOXL2 catalytically inactive
mutants mediate epithelial-to-mesenchymal transition. Biol Open.
3:129–137. 2014.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Cuevas EP, Eraso P, Mazon MJ, Santos V,
Moreno-Bueno G, Cano A and Portillo F: LOXL2 drives
epithelial-mesenchymal transition via activation of IRE1-XBP1
signalling pathway. Sci Rep. 7(44988)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wen X, Liu Y, Bai Y, Li M, Fu Q and Zheng
Y: LOXL2, a copper-dependent monoamine oxidase, activates lung
fibroblasts through the TGF-β/Smad pathway. Int J Mol Med.
42:3530–3541. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Cosgrove D, Dufek B, Meehan DT, Delimont
D, Hartnett M, Samuelson G, Gratton MA, Phillips G, MacKenna DA and
Bain G: Lysyl oxidase like-2 contributes to renal fibrosis in
Col4alpha3/Alport mice. Kidney Int. 94:303–314. 2018.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Choi SE, Jeon N, Choi HY, Shin JI, Jeong
HJ and Lim BJ: Lysyl oxidaselike 2 is expressed in kidney tissue
and is associated with the progression of tubulointerstitial
fibrosis. Mol Med Rep. 16:2477–2482. 2017.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Long DA, Woolf AS, Suda T and Yuan HT:
Increased renal angiopoietin-1 expression in folic acid-induced
nephrotoxicity in mice. J Am Soc Nephrol. 12:2721–2731.
2001.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Stallons LJ, Whitaker RM and Schnellmann
RG: Suppressed mitochondrial biogenesis in folic acid-induced acute
kidney injury and early fibrosis. Toxicol Lett. 224:326–332.
2014.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang W, Li Z, Chen Y, Wu H, Zhang S and
Chen X: Prediction value of serum NGAL in the diagnosis and
prognosis of experimental acute and chronic kidney injuries.
Biomolecules. 10(981)2020.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Barry-Hamilton V, Spangler R, Marshall D,
McCauley S, Rodriguez HM, Oyasu M, Mikels A, Vaysberg M, Ghermazien
H, Wai C, et al: Allosteric inhibition of lysyl oxidase-like-2
impedes the development of a pathologic microenvironment. Nat Med.
16:1009–1017. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Yen CL, Li YJ, Wu HH, Weng CH, Lee CC,
Chen YC, Chang MY, Yen TH, Hsu HH, Hung CC, et al: Stimulation of
transforming growth factor-beta-1 and contact with type I collagen
cooperatively facilitate irreversible transdifferentiation in
proximal tubular cells. Biomed J. 39:39–49. 2016.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Van Bergen T, Marshall D, Van de Veire S,
Vandewalle E, Moons L, Herman J, Smith V and Stalmans I: The role
of LOX and LOXL2 in scar formation after glaucoma surgery. Invest
Ophthalmol Vis Sci. 54:5788–5796. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Meissner EG, McLaughlin M, Matthews L,
Gharib AM, Wood BJ, Levy E, Sinkus R, Virtaneva K, Sturdevant D,
Martens C, et al: Simtuzumab treatment of advanced liver fibrosis
in HIV and HCV-infected adults: Results of a 6-month open-label
safety trial. Liver Int. 36:1783–1792. 2016.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Fu Y, Tang C, Cai J, Chen G, Zhang D and
Dong Z: Rodent models of AKI-CKD transition. Am J Physiol Renal
Physiol. 315:F1098–F1106. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Walkin L, Herrick SE, Summers A, Brenchley
PE, Hoff CM, Korstanje R and Margetts PJ: The role of mouse strain
differences in the susceptibility to fibrosis: A systematic review.
Fibrogenesis Tissue Repair. 6(18)2013.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Nguyen LT, Saad S, Shi Y, Wang R, Chou
ASY, Gill A, Yao Y, Jarolimek W and Pollock CA: Lysyl oxidase
inhibitors attenuate cyclosporin A-induced nephropathy in mouse.
Sci Rep. 11(12437)2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Bottinger EP and Bitzer M: TGF-beta
signaling in renal disease. J Am Soc Nephrol. 13:2600–2610.
2002.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pohlers D, Brenmoehl J, Loffler I, Müller
CK, Leipner C, Schultze-Mosgau S, Stallmach A, Kinne RW and Wolf G:
TGF-beta and fibrosis in different organs-molecular pathway
imprints. Biochim Biophys Acta. 1792:746–756. 2009.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Cheng M, Liu F, Peng Y, Chen J, Chen G,
Xiao L and Liu H: Construction of a CTGF and RFP-coexpressed renal
tubular epithelial cell and its application on evaluation of
CTGF-specific siRNAs on epithelial-mesenchymal transition. Urology.
83:1443.e1–e8. 2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Vadasz Z, Kessler O, Akiri G,
Gengrinovitch S, Kagan HM, Baruch Y, Izhak OB and Neufeld G:
Abnormal deposition of collagen around hepatocytes in Wilson's
disease is associated with hepatocyte specific expression of lysyl
oxidase and lysyl oxidase like protein-2. J Hepatol. 43:499–507.
2005.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Burns WC, Twigg SM, Forbes JM, Pete J,
Tikellis C, Thallas-Bonke V, Thomas MC, Cooper ME and Kantharidis
P: Connective tissue growth factor plays an important role in
advanced glycation end product-induced tubular
epithelial-to-mesenchymal transition: Implications for diabetic
renal disease. J Am Soc Nephrol. 17:2484–2494. 2006.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Mack M and Yanagita M: Origin of
myofibroblasts and cellular events triggering fibrosis. Kidney Int.
87:297–307. 2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Qi W, Chen X, Poronnik P and Pollock CA:
Transforming growth factor-beta/connective tissue growth factor
axis in the kidney. Int J Biochem Cell Biol. 40:9–13.
2008.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kriz W, Kaissling B and Le Hir M:
Epithelial-mesenchymal transition (EMT) in kidney fibrosis: fact or
fantasy? J Clin Invest. 121:468–474. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
36
|
Grande MT, Sanchez-Laorden B, Lopez-Blau
C, De Frutos CA, Boutet A, Arévalo M, Rowe RG, Weiss SJ,
López-Novoa JM and Nieto MA: Snail1-induced partial
epithelial-to-mesenchymal transition drives renal fibrosis in mice
and can be targeted to reverse established disease. Nat Med.
21:989–997. 2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Tang WB, Ling GH, Sun L, Zhang K, Zhu X,
Zhou X and Liu FU: Smad anchor for receptor activation regulates
high glucose-induced EMT via modulation of Smad2 and Smad3
activities in renal tubular epithelial cells. Nephron. 130:213–220.
2015.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Weiss WA, Taylor SS and Shokat KM:
Recognizing and exploiting differences between RNAi and
small-molecule inhibitors. Nat Chem Biol. 3:739–744.
2007.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Peinado H, Cruz MDC, Olmeda D, Csiszar K,
Fong KSK, Vega S, Nieto MA, Cano A and Portillo F: A molecular role
for lysyl oxidase-like 2 enzyme in snail regulation and tumor
progression. EMBO J. 24:3446–3458. 2005.PubMed/NCBI View Article : Google Scholar
|