The number of patients diagnosed with bladder cancer
(BLCA) is the tenth highest globally and men are ~3-4 times more
likely to develop BLCA than women (1,2).
BLCA can be divided into muscle-invasive BLCA (MIBC) and non-muscle
invasive BLCA (NMIBC). However, given the high mortality and
progression of MIBC and the high recurrence rate of NMIBC, bladder
cancer remains a difficult problem worldwide (3,4). The
value of bacillus Calmette Guerin (BCG) against immunotherapy in
NMIBC is widely recognized. Nonetheless, with the use of BCG, a
number of problems have appeared, such as BCG intolerance, poor
effectiveness and tumor recurrence (5). Radical cystectomy and peripheral
lymph node dissection is the gold standard treatment for advanced
MIBC. However, in some cases, patients cannot tolerate surgery or
want to retain urinary bladder function because of other disease
conditions (6). Accordingly, a new
therapeutic approach for BLCA is warranted.
Over the years, next-generation sequencing has
revealed a number of therapeutic targets for BLCA and the use of
immune checkpoints inhibitor (ICI) has offered hope for BLCA
patients (7). Nevertheless, since
ICI is expensive and patient response rates are low, significant
emphasis has been placed on combining drugs, hoping they will
complement each other (8). Natural
plant compounds are considered a good source of combination
chemotherapy due to their availability (9).
Phytochemicals are bioactive compounds extracted
from natural plants, which have been widely studied to treat
diseases, especially cancer, in vivo and in vitro,
since they are easily obtained, highly safe and non-toxic (10). Polyphenolic compounds are widely
recognized because of their wide distribution and variety. More
than 8,000 polyphenol compounds have been identified in nature.
They represent essential plant products that can be used against
cardiovascular diseases and for cancer prevention and treatment in
humans (11). Flavonoids are a
subgroup of polyphenols which represent secondary metabolites.
Flavonoids are widely regarded as the most common polyphenols in
fruits, chocolate, flowers, vegetables and tea. Their
pharmacological effects have attracted much interest, including
antioxidant, antibacterial anti-inflammatory, cardiac and liver
protective and anticancer properties (12,13).
In addition, they have been documented to prevent breast,
colorectal, thyroid, prostate, lung and ovarian cancers (13). However, flavonoids are rarely used
clinically, possibly because of their low solubility, poor
absorption and lack of accurate epidemiological data (13).
DNA damage is usually caused by damage to
single-base or double strands of DNA in tumor cells by external and
internal stimuli such as chemotherapy drugs. Double strand breaks
have the most lethal effects on cells (14). Cells can activate several
biological signals and processes in response to DNA damage,
including cell cycle arrest, apoptosis and checkpoint activation,
collectively called DNA damage response (15). The cell cycle is roughly divided
into four phases: G1 (proteins preparation), S (DNA
replication), G2 (checking the integrity of replication)
and M (Mitosis). The daughter cells then go into a resting state,
known as the G0 phase (16). It is well-established that cell
cycle progression is largely regulated by cyclin-dependent kinases
(CDKs), which phosphorylate key substrates to maintain the normal
course of the cell cycle (17).
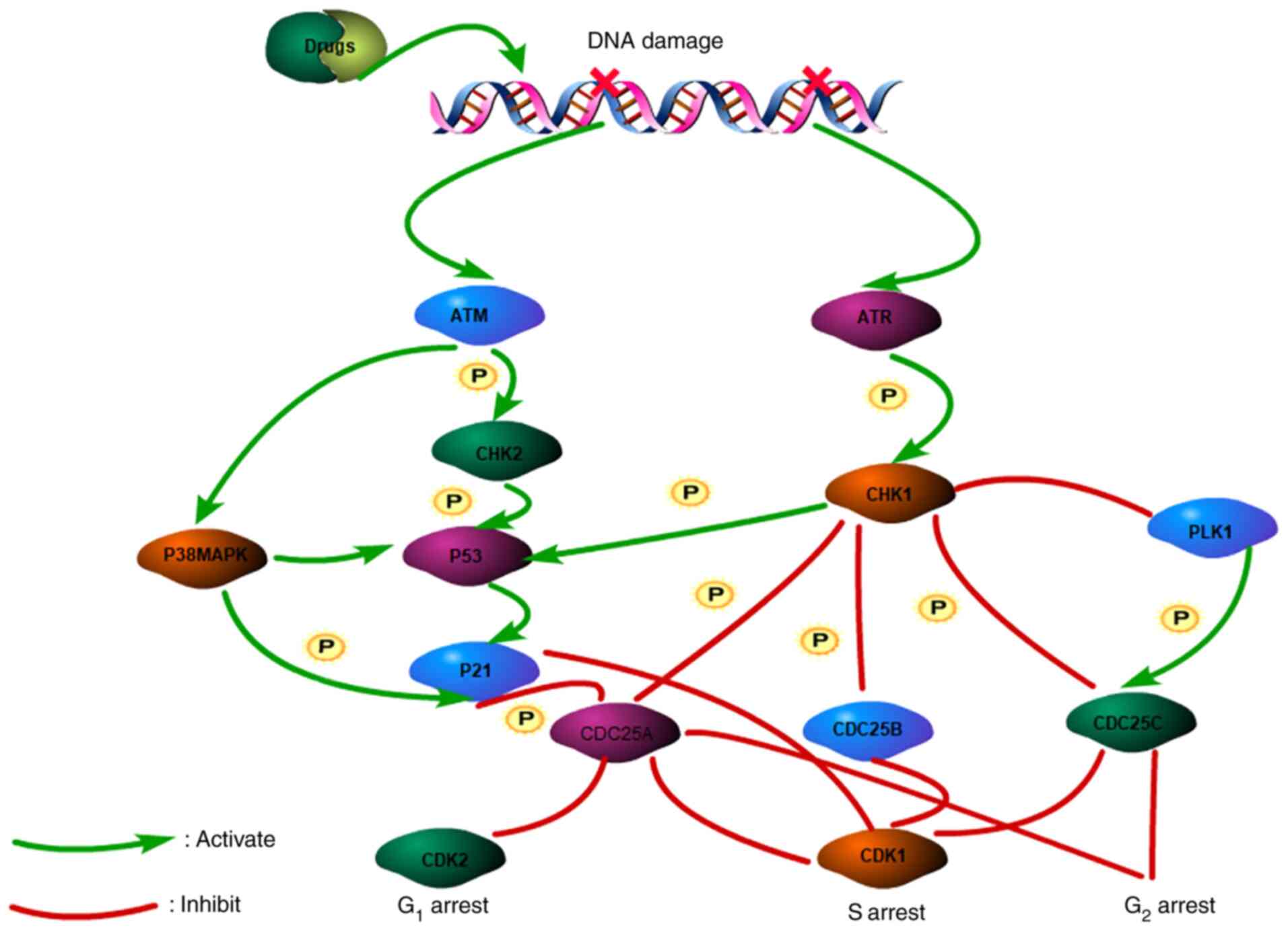
Cell cycle arrest in the G1/S phase mainly depends on
Ataxia telangiectasia mutated (ATM) activation. Notably, ATM
directly activates P38MAPK, checkpoint kinases 2 (CHK-2) and P53
leading to the accumulation of P21. The activation of ATM- and
Rad3-related (ATR) and checkpoint kinases 1 (CHK1) lead to
phosphorylated CDC25 and S or G2/M phases arrest
(18,19). DNA damage in cancer cells provides
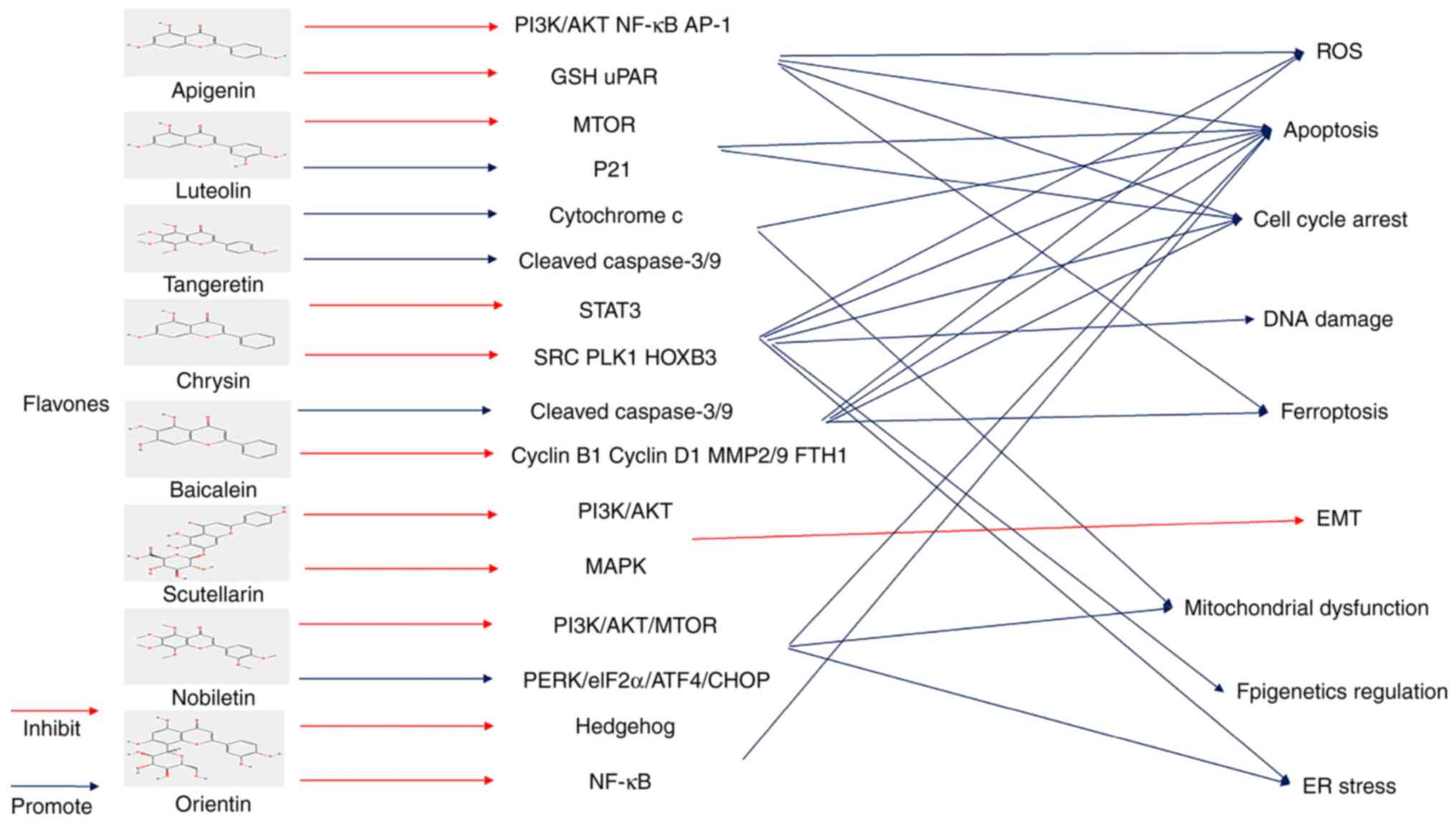
an opportunity for DNA repair by blocking the cell cycle. However,
if cancer cells are not repaired properly, they will die (20) (Fig.
1). Treatment with flavonoids can damage DNA in BLCA cells,
leading to cell cycle arrest. The sustained action of the drug can
eventually lead to cell death, such as apoptosis and other
programmed cell death.
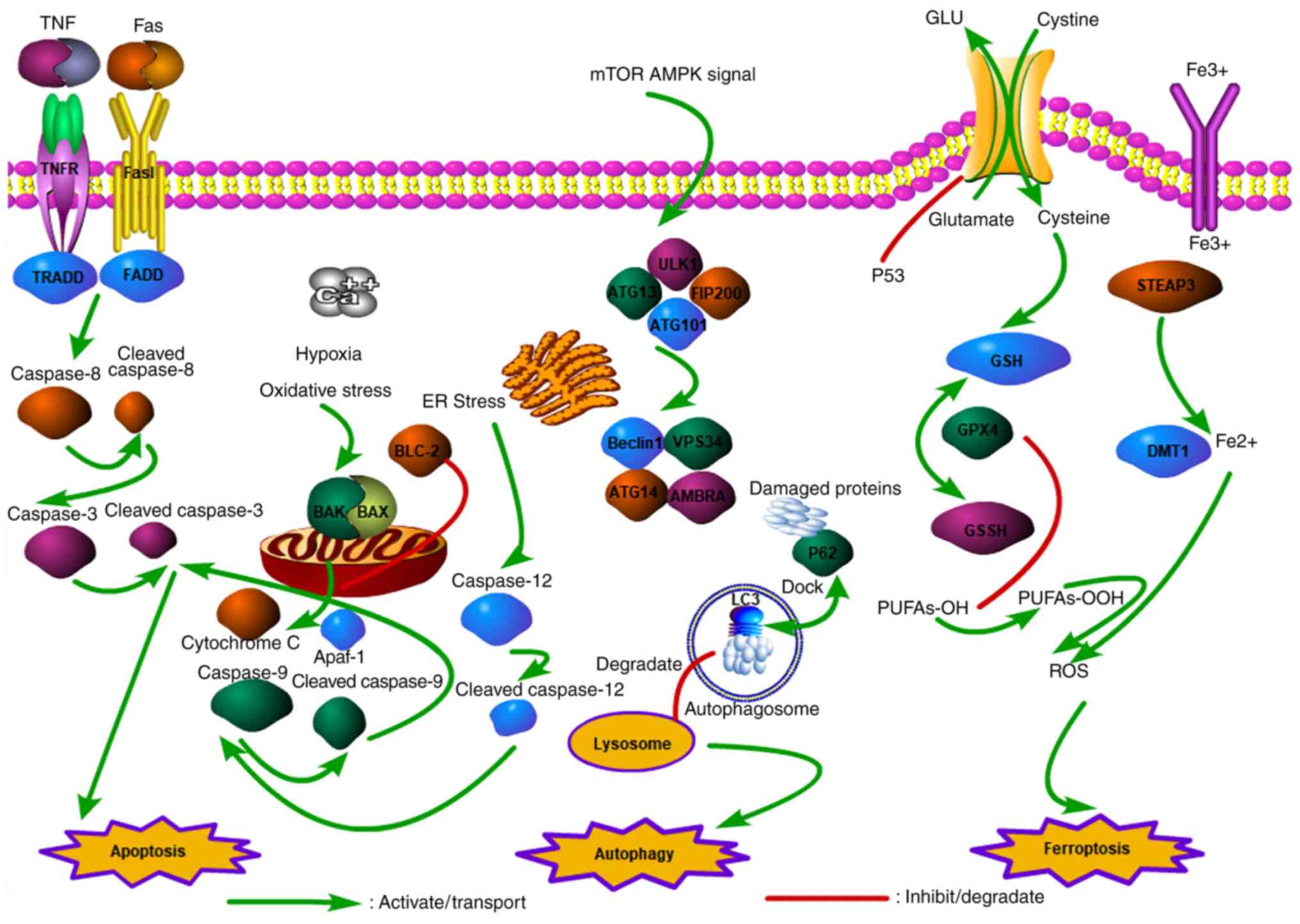
ROS are free radicals or molecules with one or more
unpaired electrons. The production of ROS in the cell depends
mainly on the oxidative stress signal stimulation by the electron
transport chain of mitochondria (21). In addition, inflammatory cells and
several enzymatic cell complexes are involved in ROS production.
The extrinsic sources of ROS mainly include radiation or drugs
(22). ROS exhibit a two-way
regulatory effect on cancer cells. Cancer cells exhibit a mild to
moderate increase in ROS due to genetic mutations or metabolic
changes, which help activate ROS-sensitive signaling pathways and
promote proliferation, invasion and differentiation of cancer
cells. Nevertheless, as a result of chemotherapy and other drugs,
the level of ROS is significantly elevated, which can cause cancer
cells to exceed existing redox limits, leading to apoptosis,
autophagy, or DNA damage (23). A
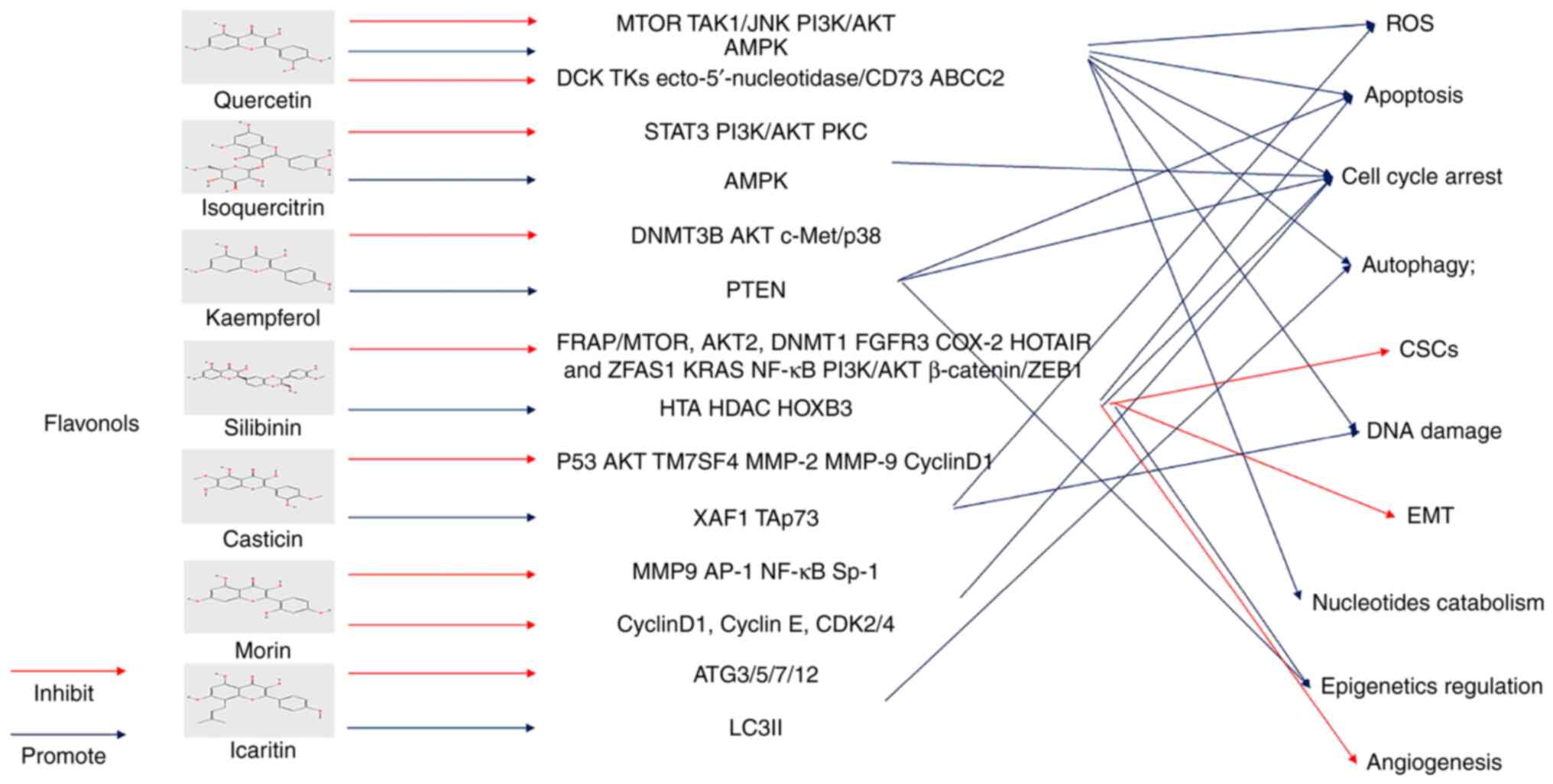
number of flavonoids can reportedly activate ROS levels and induce
BLCA cell death.
Epigenetics refers to the indirect regulation of
genes in the DNA sequence, which causes gene silencing or
overexpression and affects cell phenotype and biological function
(39). Epigenetic regulation and
modification can be divided into DNA methylation, histone
methylation, acetylation, ubiquitination and ncRNA (noncoding RNAs)
(40). DNA methylation is one of
the earliest and most widely studied modifications, involving
methylation of the 5-carbon of the Cytosine-phosphate-Guanine
islands cytosine residue, called 5-methylcytosine (41). Aberrant DNA methylation is common
in cancer genomes. Natural plant compounds are thought to influence
DNA methylation patterns by altering the global hypomethylation of
oncogenes and the hypermethylation of suppressor genes, affecting
the progression of cancer (42).
The methylation and acetylation of histone modifications are the
most widely studied. Histone methylation changes the structure and
function of chromatin, mainly through histone methyltransferases
and histone demethylases, associated with prognosis in a variety of
cancers and regulated by the active ingredients of Chinese herbs
(43). The acetylation of histones
is mainly achieved by histone acetyltransferases and histone
deacetylases (HDACs). The acetyl group of acetyl coenzyme A can be
transferred to the terminal of histone amino acids by histone
acetyltransferase to enhance DNA expression and transcription.
However, HDAC removes the acetyl group, resulting in chromatin
densification and gene transcription suppression (44). Proto-oncogenes may be activated by
hyperacetylation, while hypoacetylation of tumor suppressor genes
is usually limited to the promoter and induces gene silencing,
closely related to cancer phenotypes and traits (45). These epigenetic regulatory enzymes
may be used as therapeutic targets for BLCA.
microRNAs (miRNAs) are noncoding RNAs of ~17-25
nucleotides involved in almost all biological functions of cancer,
including proliferation, invasion, metastasis, angiogenesis and
apoptosis (46). miRNAs have been
found to act on the 3' UTR site of mRNA to suppress its expression.
Large numbers of miRNAs are reportedly upregulated or downregulated
in cancer, suggesting that they can act as biomarkers in cancer
(47). Researchers have
investigated the relationship between competitive endogenous RNAs
(ceRNAs) and cancer. Long noncoding RNAs (lncRNAs) and circular
RNAs (circRNAs) can directly target mRNAs and sponge miRNA to
regulate mRNAs expression. Notably, lncRNA/miRNA/mRNA and circRNA
miRNA/mRNA interact to form ceRNAs networks that serve regulatory
roles in cancer progression or suppression (48).
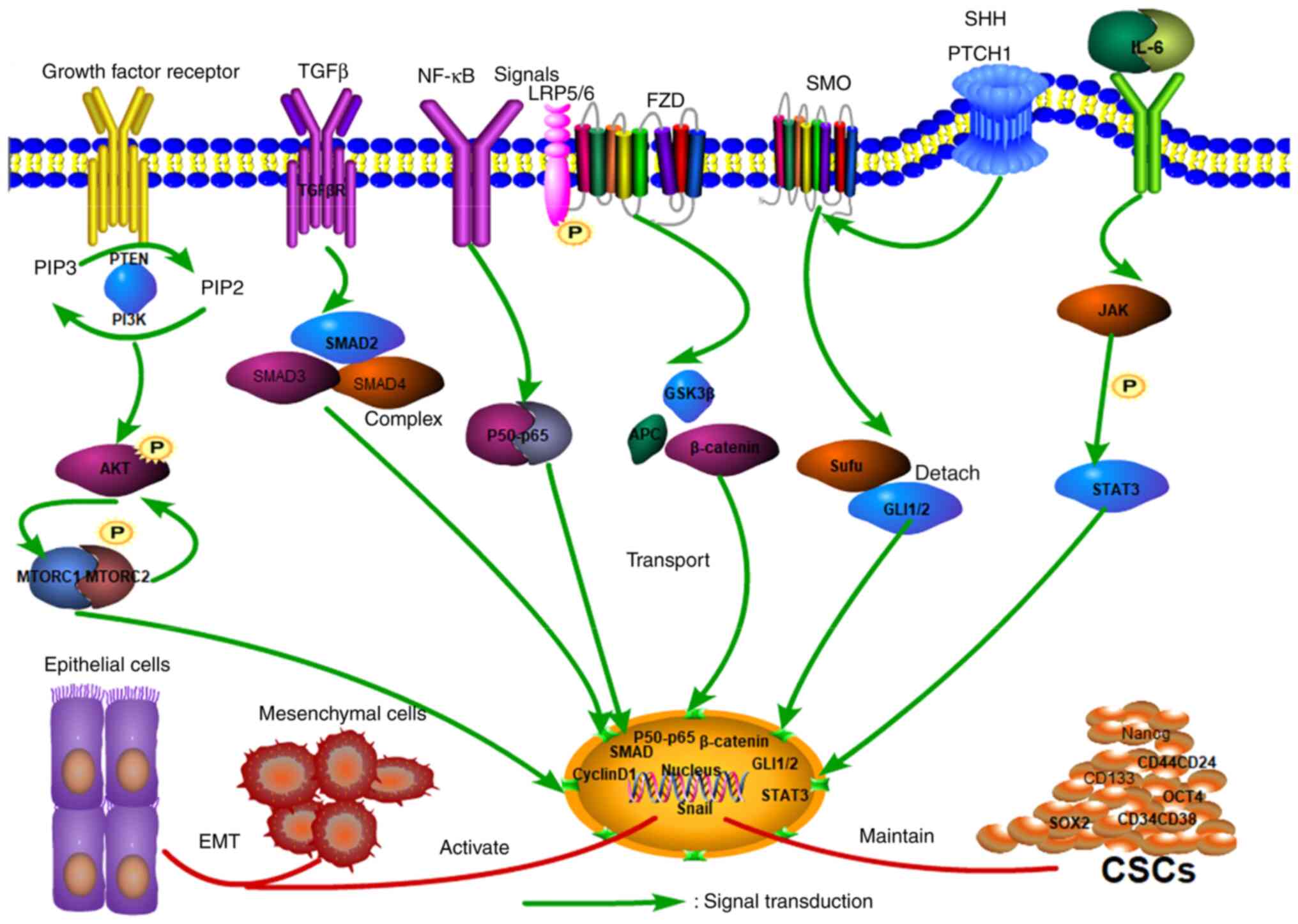
Angiogenesis primarily involves the growth of new
capillary blood vessels from the existing vascular system complex
process (49), usually due to the
proliferation and migration of endothelial cells following
stimulation to form primary sprouts. The new vascular structures
are formed by forming the basement membrane (50). Cancer cells require nutrients and
oxygen to maintain their growth through pathological angiogenesis,
which depends mainly on the overactivation of angiogenic factors.
The most important of these is the VEGF family, which serves a role
in tumor progression (51).
Nevertheless, a single angiogenesis inhibitor can only block tumor
progression to some extent. Angiogenesis inhibitors interfere with
other normal physiological functions in humans, including blood
pressure maintenance, kidney function and wound healing. It should
be borne in mind that inhibiting VEGF signaling to block tumor
angiogenesis is associated with a risk of hypertension (52).
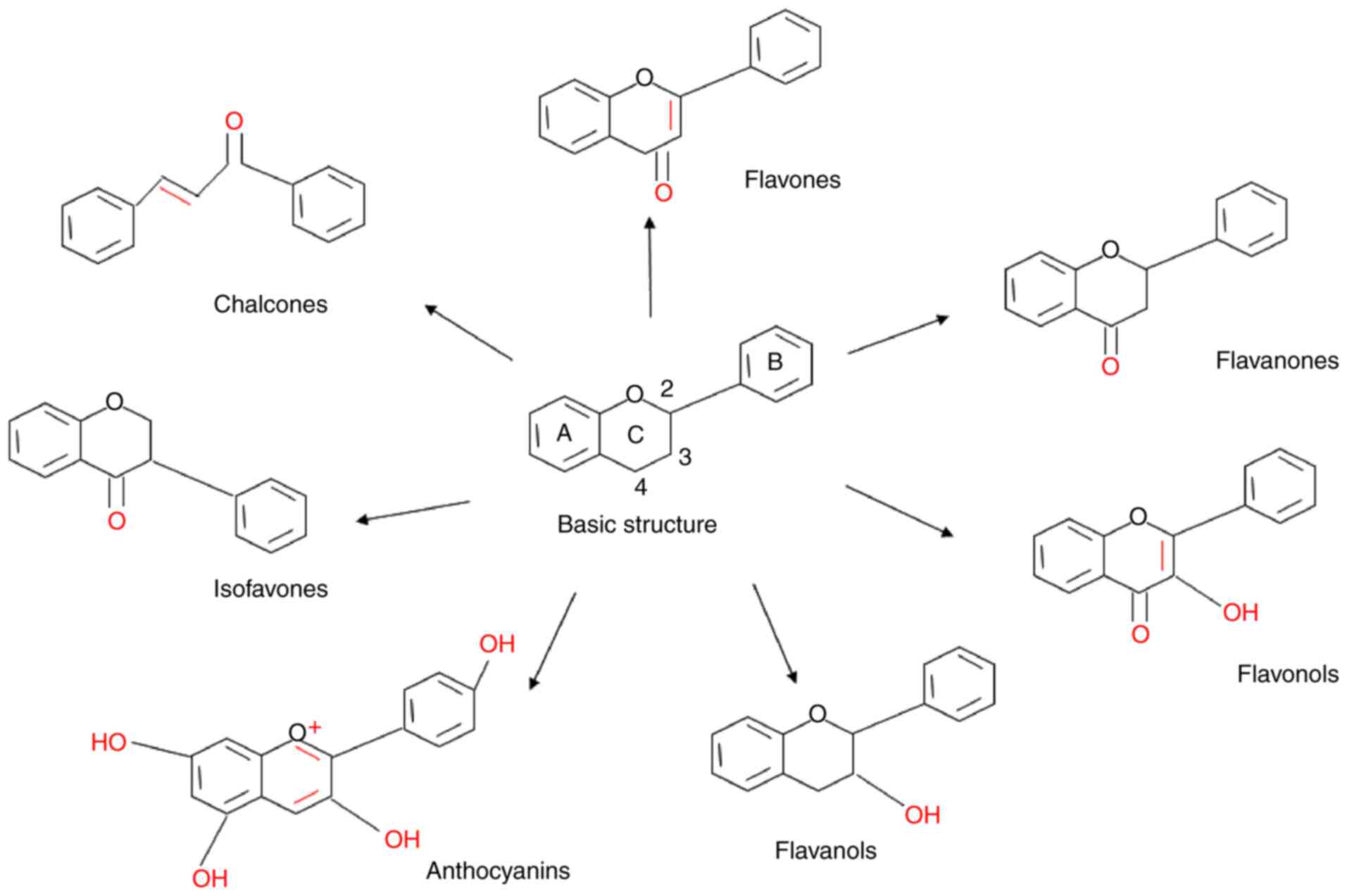
Flavonoids have a basic skeleton consisting of a
15-carbon (C6-C3-C6) phenylpropanoid chain, with two aromatic rings
(A and B) and a C heterocyclic pyran ring in the middle connected
to A and B (61). Compounds that
are connected to the 3C position of the C ring to the B ring are
termed isoflavones. However, in other types of flavonoids, B rings
are linked to the 2C, including flavones, flavanones, flavonols,
flavanols and anthocyanins. Flavones have only one keto group at
the 4C position and have a double bond between 2C and 3C, while
flavanones (dihydroflavones) have no double bond structure.
Flavanols have no keto group but have one hydroxyl group at the 3C
position and no double bonds between positions 2 and 3. The
anthocyanins are replaced by multiple hydroxyl groups, including
the 3C position and the C ring has double bonds. Flavonols have a
hydroxyl group at 3C and a keto group at 4C. Finally, chalcones
lack the ring C of the basic flavonoid structure (62,63).
(Fig. 4). The flavonoids inhibit
the development of BLCA through different mechanisms, which will be
discussed in detail later (Table
I).
Flavones are characterized by being unmodified at 3C
and can be oxidized at 4C. They can coexist with anthocyanins and
flavonols in flowers and act as plant protectors. They are usually
found in tea, parsley and citrus fruits (64).
Flavonols are also abundantly found in fruits and
vegetables. Compared with flavones, Flavonols have a hydroxyl group
which can be glycosylated on the C ring. Flavonols such as
quercetin and kaempferol have been extensively studied. Their
intake is strongly associated with health, reducing the risk of
vascular disease (63).
The development of new compounds based on quercetin
has led to improved and more significant effects. Q-ZnCPX, a novel
compound consisting of quercetin and zinc, has a stronger
inhibition and anti-metastasis effect, which may ameliorate the
disadvantages of quercetin, including low absorption and rapid
metabolism (98). Recently,
researchers have synthesized 8-trifluoromethyl-3, 5, 7, 3',
4'-O-pentamethyl-quercetin (TFQ) based on the chemical modification
of quercetin by fluorination. TFQ is believed to affect BLCA growth
through the AMPK/mTOR pathway (99). The usefulness and biocompatibility
of nanostructures have attracted much attention. Research has shown
that they could interfere with the proliferation and enhance the
radiosensitization of BLCA cells by loading quercetin into titanate
nanotubes (TNT) (100).
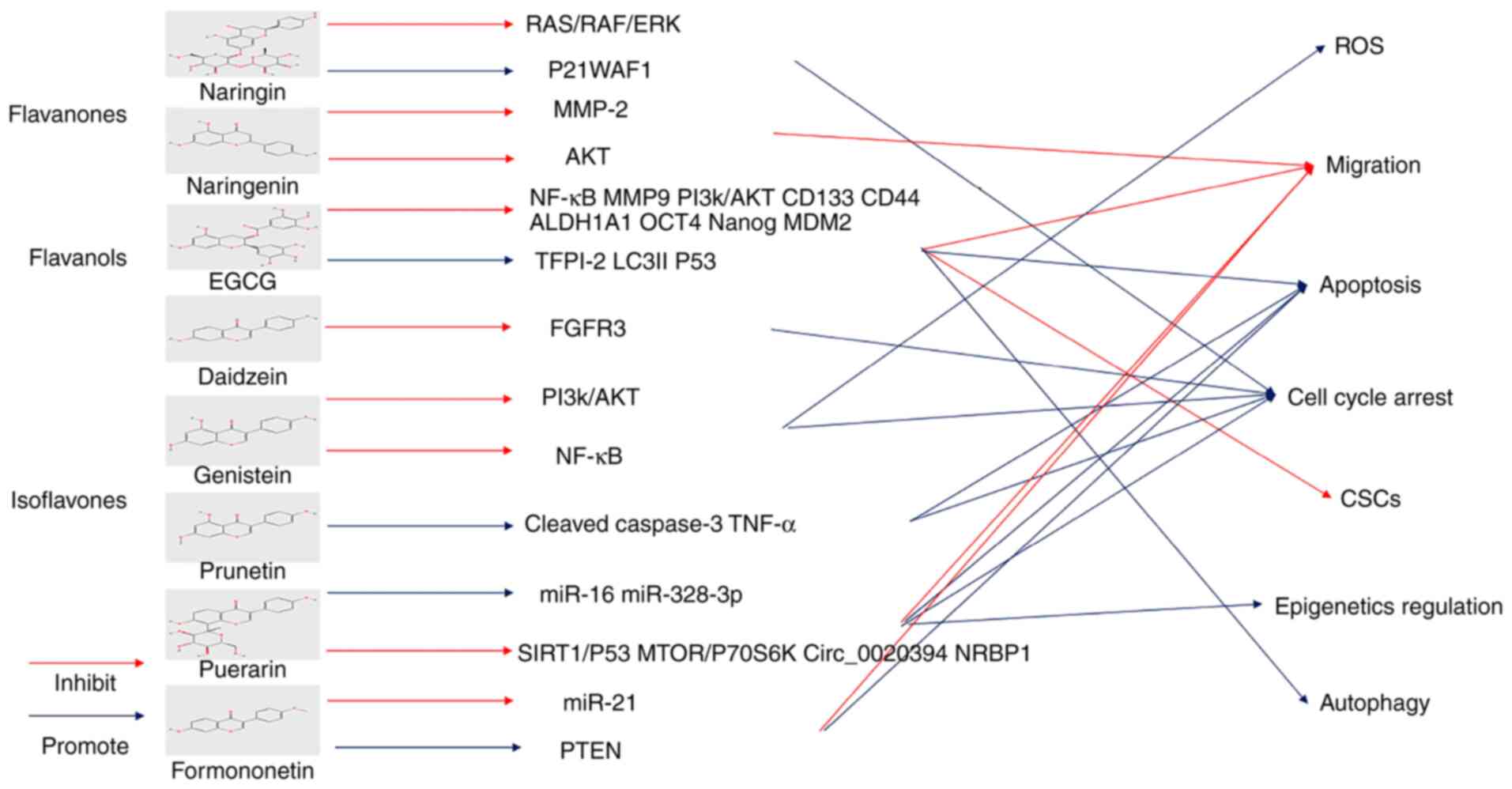
It is well-established that flavanones, also known
as dihydroflavones, have a saturated c-ring. Flavanones are found
mainly in citrus fruits such as oranges and lemons (63). Among them, hesperidin and naringin
are the most abundant ingredients with anti-oxidation and
anti-inflammatory properties and even maintain intestinal health
(125).
Green tea has attracted much interest worldwide for
its effects on cancer prevention (129). Current evidence suggests that
polyphenols, the main active compounds in tea, serve an important
anticancer role (130). Catechins
belong to the flavanol class of the flavonoid family and are the
main component of tea polyphenols (130,131). Of these, epigallocatechin gallate
(EGCG) is the most abundant and biologically active member of the
catechin family, accounting for >50% of the family (132). High consumption of green tea
could reduce the recurrence and progression of urothelial carcinoma
(133). Notably, it has been
shown that green tea polyphenols can inhibit cytoplasmic human
antigen R expression in a BLCA model. In addition, it can suppress
BLCA cell proliferation and angiogenesis and the expression of
related proteins, including VEGF-A, heme oxygenase (HO)-1 and
COX-2(134). Mg (II)-catechin
nanoparticles (Mg (II)-Cat NPs) display a significant inhibitory
effect on BLCA, given their improved biocompatibility and stronger
cellular uptake. In addition, eukaryotic translation initiation
factor 5A2 (EIF5A2) small interfering RNA (siRNA) can be loaded
into the tumor site to further enhance the anti-BLCA effect via the
PI3K/AKT pathway (135).
In animal models, EGCG prevents bladder tumor
implantation and development by reducing proteolytic activity, with
a slightly higher therapeutic effect compared with mitomycin C
(136). Next-generation
sequencing reveals the related mRNAs, miRNAs and mechanisms of EGCG
on BFTC-905 cells (137). EGCG
can inhibit the proliferation and migration of BLCA cells (SW780,
5637 and T24) and promote cell apoptosis by suppressing NF-KB and
MMP9 and PI3k/AKT pathways (138-140).
As well as apoptosis, tissue factor pathway inhibitor 2 is reported
to be upregulated by EGCG to inhibit the growth of BLCA cells via
decreasing promoter hypermethylation (141). Notably, low-dose EGCG promotes
LC3I to LC3II, suggesting the occurrence of autophagy. The
autophagy effect is blocked by a PI3K/AKT inhibitor (LY294002)
(142). The effect of EGCG on
bladder CSCs has also been studied. In this respect, EGCG has been
shown to inhibit the expression of CD133, CD44, ALDH1A1, OCT4 and
Nanog and sonic hedgehog signaling pathways to inhibit bladder CSCs
(143). It has been suggested
that EGCG can be combined with docetaxel to enhance the induction
of apoptosis in BLCA cells by modulating the NF-κB/MDM2/p53 pathway
(144).
Anthocyanins and anthocyanidins are plant pigments
that account for various colors in plants and fruits. Anthocyanins
are anthocyanidins structurally modified by sugar and acyl acids
found mainly in dark fruits with excellent potential to inhibit
tumor progression (145,146). The combination of anthocyanins, a
bladder cancer preventive agent and mitomycin C has been reported
to increase BLCA cell death (147).
Isoflavones are mainly derived from soybean and
soybean products foods. A high content of daidzein and genistein is
present in isoflavones. Isoflavones are also thought to be
protective agents against hormonal disorders and suppress a wide
range of cancers, including prostate and breast cancer (153).
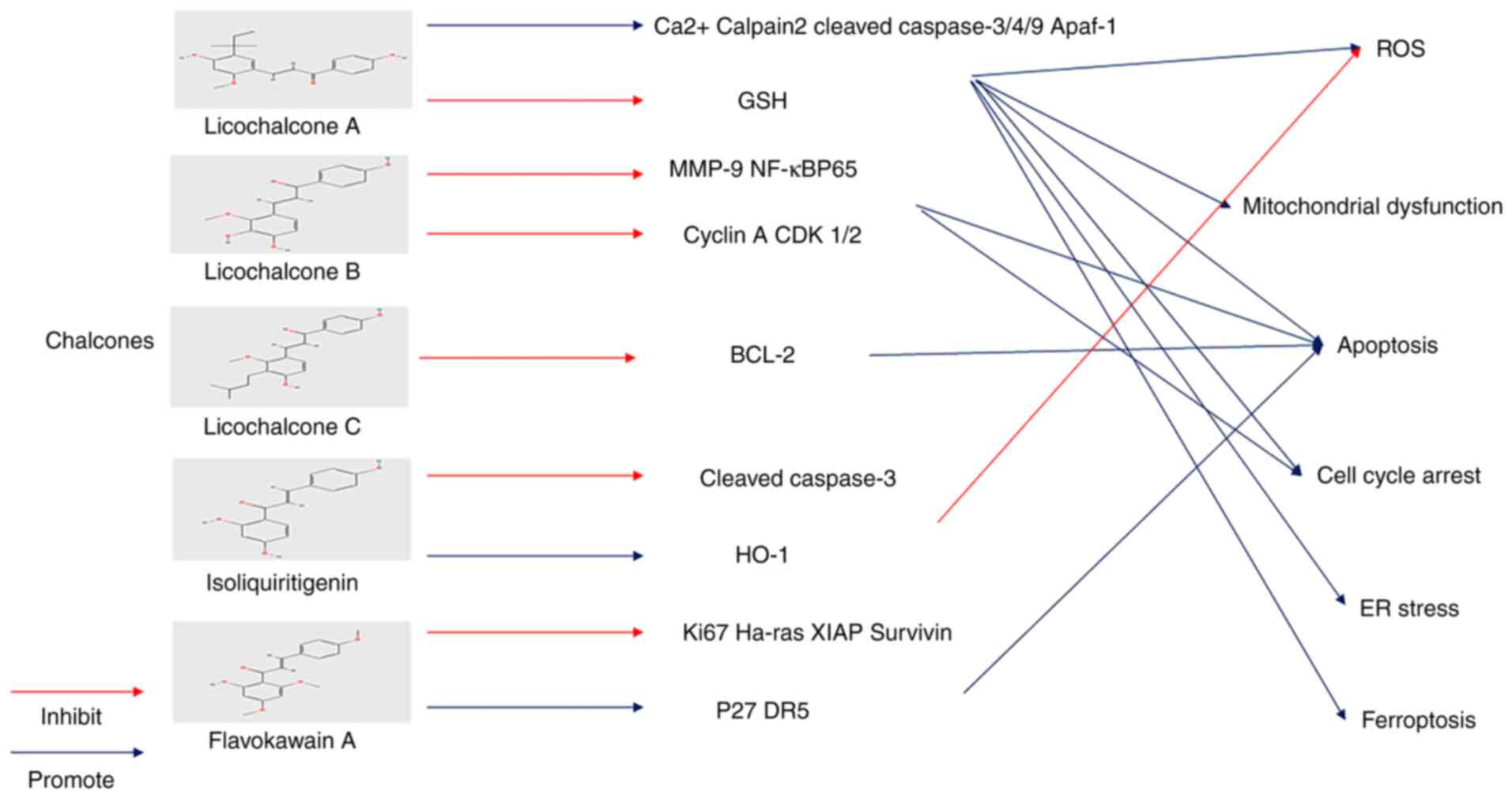
Chalcones are widely found in fruits and vegetables
and are important components and biological precursors of
flavonoids. They have a basic 1, 3-diaryl-2-propen-1-one chemical
scaffold and two aromatic rings connected by an unsaturated α,
β-carbonyl system (166). The
effect of chalcones on BLCA has been extensively studied in recent
years.
In addition, Licochalcone B (LCB) can reduce the
expression of MMP-9 mRNA and protein, but MMP-2 does not. LCB can
promote nuclear translocation of NFκB and suppress NF-кBP65 protein
expression. This indicates that LCB exerts a potential therapeutic
effect on the invasion and metastasis of BLCA (171). In addition, LCB can regulate the
cell cycle by inhibiting cyclin A and CDK 1/2 mRNA. LCB inhibits
colony formation and promoted apoptosis of BLCA cells (172).
Licochalcone C (LCC) has also been shown to induce
T24 cell apoptosis by regulating the biological function of the
Bcl-2 family (173).
IPP51 (1-(2,4-dimethoxyphenyl)-3-(1-methylindolyl)
propenone) is a novel derivative for chalcone that can promote
apoptosis and G2+M accumulation in BLCA cells and
inhibit mitosis and destroy microtubules by promoting the
production of soluble tubulin and inhibiting tubulin
polymerization. In addition, IPP51 exerts an anti-angiogenesis
effect (180,181).
Flavonoids have been found to serve a powerful role
in sensitizing patients to chemotherapy. Cisplatin is one of the
most common chemotherapy drugs in clinical practice. It has been
used for a number of years and is still the cornerstone of
chemotherapy for advanced BLCA and metastasis. Reducing its side
effects and making it more sensitive to patients has become a
research hotspot (182,183). Current evidence suggests that
isoliquiritigenin can improve the nephrotoxicity of cisplatin and
increase the damage to BLCA cells (175). In addition, silibinin has been
shown to alleviate chemodrug-induced chemoresistance through the
NF-κB pathway (115).
Chemotherapy remains an important means to treat cancer;
chemotherapy drugs combined with other drugs, including immune
checkpoint inhibitors, have been used to treat BLCA. However, due
to the high selectivity of patients to checkpoint inhibitors, the
effect is not ideal. Flavonoids represent a promising candidate for
a new class of drugs that can be combined with chemotherapy to
suppress the recurrence and progression of BLCA. Given that they
are harmless and widely available, they bring less financial burden
and psychological stress to patients.
The modification of nanoparticles offsets some of
the drawbacks of flavonoids. Flavonoids are widely acknowledged for
their poor targeting ability and faster metabolism, which are major
concerns affecting their efficacy (184). Nanoparticles can be encapsulated
and target tumors to increase their half-life and reduce
immunogenicity. In addition, nanoparticles can be loaded with
various drugs to improve drug resistance and with diagnostic agents
for integrated treatment (185).
Notably, the Mg (II)-Cat/siEIF5A2 nanoparticle combined with
flavonoid and siRNA yields a stronger BLCA inhibitory effect
(135). The combination of
flavonoids and nanoparticles remains rare in the treatment of BLCA
and deserves further study.
The mechanisms underlying the therapeutic effect of
flavonoids are quite extensive and the generation of ROS seems to
act as a switch in a variety of mechanisms (22). Further work on ROS is warranted.
The majority of studies have primarily investigated the mechanism
of cell cycle arrest and apoptosis, with more emphasis needed on
autophagy and ferroptosis. Indeed, autophagy has both positive and
negative effects on cancer (31).
Different concentrations of drugs may have different effects on
autophagy. In addition, inhibition of autophagy appears to promote
cell apoptosis. The autophagy changes can be accurately assessed by
detecting the transformation from LC3I to LC3II (31). Accordingly, there is still much
room for research on ferroptosis in flavonoids. Notably, the change
in the GSH/GSSG ratio and the expression of GPX4 can reflect the
occurrence of ferroptosis (36).
ROS activation is also key to the occurrence of ferroptosis. P53 is
not only a tumor suppressor gene but also a regulator of
ferroptosis. Its upregulation can promote ferroptosis in cells by
inhibiting the system Xc-transporter. CSCs play an important role
in the progression of BLCA and multiple marker genes are
overafctivated in CSCs. Targeting these genes, including OCT4,
KLF4, c-MYC and Nanog, can inhibit the transformation of BLCA stem
cells. In addition, CSCs and EMT have been documented in the
abnormal activation of multiple common pathways, including the WNT,
STAT3 and NF-KB pathways, which can be investigated in future
studies.
Funding: No funding was received.
Data sharing is not applicable to this article, as
no data sets were generated or analyzed during the current
study.
YL, ZhL and HJ wrote the first draft and drew the
figures and tables. ZaL and LD designed this article and modified
it. YX revised the draft and the figures. Data authentication is
not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Dobruch J, Daneshmand S, Fisch M, Lotan Y,
Noon AP, Resnick MJ, Shariat SF, Zlotta AR and Boorjian SA: Gender
and bladder cancer: A collaborative review of etiology, biology,
and outcomes. Eur Urol. 69:300–310. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Richters A, Aben KKH and Kiemeney LALM:
The global burden of urinary bladder cancer: An update. World J
Urol. 38:1895–1904. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Xia Y, Chen R, Lu G, Li C, Lian S, Kang TW
and Jung YD: Natural phytochemicals in bladder cancer prevention
and therapy. Front Oncol. 11(652033)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Han J, Gu X, Li Y and Wu Q: Mechanisms of
BCG in the treatment of bladder cancer-current understanding and
the prospect. Biomed Pharmacother. 129(110393)2020.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Kimura T, Ishikawa H, Kojima T, Kandori S,
Kawahara T, Sekino Y, Sakurai H and Nishiyama H: Bladder
preservation therapy for muscle invasive bladder cancer: The past,
present and future. Jpn J Clin Oncol. 50:1097–1107. 2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Tran L, Xiao JF, Agarwal N, Duex JE and
Theodorescu D: Advances in bladder cancer biology and therapy. Nat
Rev Cancer. 21:104–121. 2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Bednova O and Leyton JV: Targeted
molecular therapeutics for bladder cancer-A new option beyond the
mixed fortunes of immune checkpoint inhibitors? Int J Mol Sci.
21(7268)2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Rutz J, Janicova A, Woidacki K, Chun FK,
Blaheta RA and Relja B: Curcumin-A viable agent for better bladder
cancer treatment. Int J Mol Sci. 21(3761)2020.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zanoaga O, Braicu C, Jurj A, Rusu A, Buiga
R and Berindan-Neagoe I: Progress in research on the role of
flavonoids in lung cancer. Int J Mol Sci. 20(4291)2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Niedzwiecki A, Roomi MW, Kalinovsky T and
Rath M: Anticancer efficacy of polyphenols and their combinations.
Nutrients. 8(552)2016.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kumar S and Pandey AK: Chemistry and
biological activities of flavonoids: An overview.
ScientificWorldJournal. 2013(162750)2013.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Amawi H, Ashby CR Jr and Tiwari AK: Cancer
chemoprevention through dietary flavonoids: What's limiting? Chin J
Cancer. 36(50)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lama-Sherpa TD and Shevde LA: An emerging
regulatory role for the tumor microenvironment in the DNA damage
response to double-strand breaks. Mol Cancer Res. 18:185–193.
2020.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Srinivas US, Tan BWQ, Vellayappan BA and
Jeyasekharan AD: ROS and the DNA damage response in cancer. Redox
Biol. 25(101084)2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Harashima H, Dissmeyer N and Schnittger A:
Cell cycle control across the eukaryotic kingdom. Trends Cell Biol.
23:345–356. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lim S and Kaldis P: Cdks, cyclins and
CKIs: Roles beyond cell cycle regulation. Development.
140:3079–3093. 2013.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Carusillo A and Mussolino C: DNA Damage:
From threat to treatment. Cells. 9(1665)2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Solier S, Zhang YW, Ballestrero A, Pommier
Y and Zoppoli G: DNA damage response pathways and cell cycle
checkpoints in colorectal cancer: Current concepts and future
perspectives for targeted treatment. Curr Cancer Drug Targets.
12:356–371. 2012.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Kastan MB and Bartek J: Cell-cycle
checkpoints and cancer. Nature. 432:316–323. 2004.PubMed/NCBI View Article : Google Scholar
|
|
21
|
de Sá Junior PL, Câmara DAD, Porcacchia
AS, Fonseca PMM, Jorge SD, Araldi RP and Ferreira AK: The roles of
ROS in cancer heterogeneity and therapy. Oxid Med Cell Longev.
2017(2467940)2017.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Aggarwal V, Tuli HS, Varol A, Thakral F,
Yerer MB, Sak K, Varol M, Jain A, Khan MA and Sethi G: Role of
reactive oxygen species in cancer progression: Molecular mechanisms
and recent advancements. Biomolecules. 9(735)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Perillo B, Di Donato M, Pezone A, Di Zazzo
E, Giovannelli P, Galasso G, Castoria G and Migliaccio A: ROS in
cancer therapy: the bright side of the moon. Exp Mol Med.
52:192–203. 2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Xu X, Lai Y and Hua ZC: Apoptosis and
apoptotic body: Disease message and therapeutic target potentials.
Biosci Rep. 39(BSR20180992)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hengartner MO: Apoptosis: Corralling the
corpses. Cell. 104:325–328. 2001.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Schneider P and Tschopp J: Apoptosis
induced by death receptors. Pharm Acta Helv. 74:281–286.
2000.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Indran IR, Tufo G, Pervaiz S and Brenner
C: Recent advances in apoptosis, mitochondria and drug resistance
in cancer cells. Biochim Biophys Acta. 1807:735–745.
2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Bertheloot D, Latz E and Franklin BS:
Necroptosis, pyroptosis and apoptosis: An intricate game of cell
death. Cell Mol Immunol. 18:1106–1121. 2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res.
30(87)2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Szegezdi E, Fitzgerald U and Samali A:
Caspase-12 and ER-stress-mediated apoptosis: The story so far. Ann
N Y Acad Sci. 1010:186–194. 2003.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Levy JMM, Towers CG and Thorburn A:
Targeting autophagy in cancer. Nat Rev Cancer. 17:528–542.
2017.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Amaravadi RK, Kimmelman AC and Debnath J:
Targeting autophagy in cancer: Recent advances and future
directions. Cancer Discov. 9:1167–1181. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
White E, Mehnert JM and Chan CS:
Autophagy, metabolism, and cancer. Clin Cancer Res. 21:5037–5046.
2015.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Amaravadi R, Kimmelman AC and White E:
Recent insights into the function of autophagy in cancer. Genes
Dev. 30:1913–1930. 2016.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Mou Y, Wang J, Wu J, He D, Zhang C, Duan C
and Li B: Ferroptosis, a new form of cell death: Opportunities and
challenges in cancer. J Hematol Oncol. 12(34)2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Xu T, Ding W, Ji X, Ao X, Liu Y, Yu W and
Wang J: Molecular mechanisms of ferroptosis and its role in cancer
therapy. J Cell Mol Med. 23:4900–4912. 2019.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Bebber CM, Müller F, Prieto Clemente L,
Weber J and von Karstedt S: Ferroptosis in cancer cell biology.
Cancers (Basel). 12(164)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Li J, Cao F, Yin HL, Huang ZJ, Lin ZT, Mao
N, Sun B and Wang G: Ferroptosis: past, present and future. Cell
Death Dis. 11(88)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Tiffon C: The impact of nutrition and
environmental epigenetics on human health and disease. Int J Mol
Sci. 19(3425)2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Margueron R and Reinberg D: Chromatin
structure and the inheritance of epigenetic information. Nat Rev
Genet. 11:285–296. 2010.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Mahmoud AM and Ali MM: Methyl donor
micronutrients that modify DNA methylation and cancer outcome.
Nutrients. 11(608)2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Jasek K, Kubatka P, Samec M, Liskova A,
Smejkal K, Vybohova D, Bugos O, Biskupska-Bodova K, Bielik T, Zubor
P, et al: DNA methylation status in cancer disease: Modulations by
plant-derived natural compounds and dietary interventions.
Biomolecules. 9(289)2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Huang Z, Huang Q, Ji L, Wang Y, Qi X, Liu
L, Liu Z and Lu L: Epigenetic regulation of active Chinese herbal
components for cancer prevention and treatment: A follow-up review.
Pharmacol Res. 114:1–12. 2016.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Qin J, Wen B, Liang Y, Yu W and Li H:
Histone modifications and their role in colorectal cancer (Review).
Pathol Oncol Res. 26:2023–2033. 2020.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Audia JE and Campbell RM: Histone
modifications and cancer. Cold Spring Harb Perspect Biol.
8(a019521)2016.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Ali Syeda Z, Langden SSS, Munkhzul C, Lee
M and Song SJ: Regulatory mechanism of MicroRNA expression in
cancer. Int J Mol Sci. 21(1723)2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Hanahan D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Koch AE and Distler O: Vasculopathy and
disordered angiogenesis in selected rheumatic diseases: Rheumatoid
arthritis and systemic sclerosis. Arthritis Res Ther. 9 (Suppl
2)(S3)2007.PubMed/NCBI View
Article : Google Scholar
|
|
51
|
Ramjiawan RR, Griffioen AW and Duda DG:
Anti-angiogenesis for cancer revisited: Is there a role for
combinations with immunotherapy? Angiogenesis. 20:185–204.
2017.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Rajabi M and Mousa SA: The role of
angiogenesis in cancer treatment. Biomedicines.
5(34)2017.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Pan G, Liu Y, Shang L, Zhou F and Yang S:
EMT-associated microRNAs and their roles in cancer stemness and
drug resistance. Cancer Commun (Lond). 41:199–217. 2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Eun K, Ham SW and Kim H: Cancer stem cell
heterogeneity: Origin and new perspectives on CSC targeting. BMB
Rep. 50:117–125. 2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Barzegar Behrooz A, Syahir A and Ahmad S:
CD133: Beyond a cancer stem cell biomarker. J Drug Target.
27:257–269. 2019.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Huang T, Song X, Xu D, Tiek D, Goenka A,
Wu B, Sastry N, Hu B and Cheng SY: Stem cell programs in cancer
initiation, progression, and therapy resistance. Theranostics.
10:8721–8743. 2020.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Du B and Shim JS: Targeting
epithelial-mesenchymal transition (EMT) to overcome drug resistance
in cancer. Molecules. 21(965)2016.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Lehman HL, Kidacki M and Stairs DB: Twist2
is NFkB-responsive when p120-catenin is inactivated and EGFR is
overexpressed in esophageal keratinocytes. Sci Rep.
10(18829)2020.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Luongo F, Colonna F, Calapà F, Vitale S,
Fiori ME and De Maria R: PTEN tumor-suppressor: The dam of stemness
in cancer. Cancers (Basel). 11(1076)2019.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Kopustinskiene DM, Jakstas V, Savickas A
and Bernatoniene J: Flavonoids as anticancer agents. Nutrients.
12(457)2020.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Abotaleb M, Samuel SM, Varghese E,
Varghese S, Kubatka P, Liskova A and Büsselberg D: Flavonoids in
cancer and apoptosis. Cancers (Basel). 11(28)2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Panche AN, Diwan AD and Chandra SR:
Flavonoids: An overview. J Nutr Sci. 5(e47)2016.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Hostetler GL, Ralston RA and Schwartz SJ:
Flavones: Food sources, bioavailability, metabolism, and
bioactivity. Adv Nutr. 8:423–435. 2017.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Shi MD, Shiao CK, Lee YC and Shih YW:
Apigenin, a dietary flavonoid, inhibits proliferation of human
bladder cancer T-24 cells via blocking cell cycle progression and
inducing apoptosis. Cancer Cell Int. 15(33)2015.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Zhu Y, Mao Y, Chen H, Lin Y, Hu Z, Wu J,
Xu X, Xu X, Qin J and Xie L: Apigenin promotes apoptosis, inhibits
invasion and induces cell cycle arrest of T24 human bladder cancer
cells. Cancer Cell Int. 13(54)2013.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Xia Y, Yuan M, Li S, Thuan UT, Nguyen TT,
Kang TW, Liao W, Lian S and Jung YD: Apigenin Suppresses the
IL-1β-induced expression of the urokinase-type plasminogen
activator receptor by inhibiting MAPK-Mediated AP-1 and NF-κB
signaling in human bladder cancer T24 cells. J Agric Food Chem.
66:7663–7673. 2018.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Lin Y, Shi R, Wang X and Shen HM:
Luteolin, a flavonoid with potential for cancer prevention and
therapy. Curr Cancer Drug Targets. 8:634–646. 2008.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Kilani-Jaziri S, Frachet V, Bhouri W,
Ghedira K, Chekir-Ghedira L and Ronot X: Flavones inhibit the
proliferation of human tumor cancer cell lines by inducing
apoptosis. Drug Chem Toxicol. 35:1–10. 2012.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Iida K, Naiki T, Naiki-Ito A, Suzuki S,
Kato H, Nozaki S, Nagai T, Etani T, Nagayasu Y, Ando R, et al:
Luteolin suppresses bladder cancer growth via regulation of
mechanistic target of rapamycin pathway. Cancer Sci. 111:1165–1179.
2020.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Yang G, Wang Z, Wang W, Zhou X, Hu X and
Yang J: Anticancer activity of Luteolin and its synergism effect
with BCG on human bladder cancer cell line BIU-87. Zhong Nan Da Xue
Xue Bao Yi Xue Ban. 39:371–378. 2014.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
72
|
Lin JJ, Huang CC, Su YL, Luo HL, Lee NL,
Sung MT and Wu YJ: Proteomics analysis of tangeretin-induced
apoptosis through mitochondrial dysfunction in bladder cancer
cells. Int J Mol Sci. 20(1017)2019.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Mani R and Natesan V: Chrysin: Sources,
beneficial pharmacological activities, and molecular mechanism of
action. Phytochemistry. 145:187–196. 2018.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Xu Y, Tong Y, Ying J, Lei Z, Wan L, Zhu X,
Ye F, Mao P, Wu X, Pan R, et al: Chrysin induces cell growth
arrest, apoptosis, and ER stress and inhibits the activation of
STAT3 through the generation of ROS in bladder cancer cells. Oncol
Lett. 15:9117–9125. 2018.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Lima APB, Almeida TC, Barros TMB, Rocha
LCM, Garcia CCM and da Silva GN: Toxicogenetic and
antiproliferative effects of chrysin in urinary bladder cancer
cells. Mutagenesis: Aug 13, 2020 (Epub ahead of print).
|
|
76
|
Yang Y, Liu K, Yang L and Zhang G: Bladder
cancer cell viability inhibition and apoptosis induction by
baicalein through targeting the expression of anti-apoptotic genes.
Saudi J Biol Sci. 25:1478–1482. 2018.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Choi EO, Park C, Hwang HJ, Hong SH, Kim
GY, Cho EJ, Kim WJ and Choi YH: Baicalein induces apoptosis via
ROS-dependent activation of caspases in human bladder cancer 5637
cells. Int J Oncol. 49:1009–1018. 2016.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Li HL, Zhang S, Wang Y, Liang RR, Li J, An
P, Wang ZM, Yang J and Li ZF: Baicalein induces apoptosis via a
mitochondrial-dependent caspase activation pathway in T24 bladder
cancer cells. Mol Med Rep. 7:266–270. 2013.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Kong N, Chen X, Feng J, Duan T, Liu S, Sun
X, Chen P, Pan T, Yan L, Jin T, et al: Baicalin induces ferroptosis
in bladder cancer cells by downregulating FTH1. Acta Pharm Sin B.
11:4045–4054. 2021.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Wu JY, Tsai KW, Li YZ, Chang YS, Lai YC,
Laio YH, Wu JD and Liu YW: Anti-bladder-tumor effect of baicalein
from scutellaria baicalensis georgi and its application in vivo.
Evid Based Complement Alternat Med. 2013(579751)2013.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Peng L, Wen L, Shi QF, Gao F, Huang B,
Meng J, Hu CP and Wang CM: Scutellarin ameliorates pulmonary
fibrosis through inhibiting NF-κB/NLRP3-mediated
epithelial-mesenchymal transition and inflammation. Cell Death Dis.
11(978)2020.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Lv WL, Liu Q, An JH and Song XY:
Scutellarin inhibits hypoxia-induced epithelial-mesenchymal
transition in bladder cancer cells. J Cell Physiol.
234:23169–23175. 2019.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Ashrafizadeh M, Zarrabi A, Saberifar S,
Hashemi F, Hushmandi K, Hashemi F, Moghadam ER, Mohammadinejad R,
Najafi M and Garg M: Nobiletin in cancer therapy: How this plant
derived-natural compound targets various oncogene and
onco-suppressor pathways. Biomedicines. 8(110)2020.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Goan YG, Wu WT, Liu CI, Neoh CA and Wu YJ:
Involvement of mitochondrial dysfunction, endoplasmic reticulum
stress, and the PI3K/AKT/mTOR pathway in nobiletin-induced
apoptosis of human bladder cancer cells. Molecules.
24(2881)2019.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Tian F, Tong M, Li Z, Huang W, Jin Y, Cao
Q, Zhou X and Tong G: The effects of orientin on proliferation and
apoptosis of T24 human bladder carcinoma cells occurs through the
inhibition of nuclear factor-kappaB and the hedgehog signaling
pathway. Med Sci Monit. 25:9547–9554. 2019.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Stavric B: Quercetin in our diet: From
potent mutagen to probable anticarcinogen. Clin Biochem.
27:245–248. 1994.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Rauf A, Imran M, Khan IA, Ur-Rehman M,
Gilani SA, Mehmood Z and Mubarak MS: Anticancer potential of
quercetin: A comprehensive review. Phytother Res. 32:2109–2130.
2018.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Adami BS, Diz FM, Oliveira Gonçalves GP,
Reghelin CK, Scherer M, Dutra AP, Papaléo RM, de Oliveira JR,
Morrone FB, Wieck A and Xavier LL: Morphological and mechanical
changes induced by quercetin in human T24 bladder cancer cells.
Micron. 151(103152)2021.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Oršolić N, Karač I, Sirovina D, Kukolj M,
Kunštić M, Gajski G, Garaj-Vrhovac V and Štajcar D:
Chemotherapeutic potential of quercetin on human bladder cancer
cells. J Environ Sci Health A Tox Hazard Subst Environ Eng.
51:776–781. 2016.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Su Q, Peng M, Zhang Y, Xu W, Darko KO, Tao
T, Huang Y, Tao X and Yang X: Quercetin induces bladder cancer
cells apoptosis by activation of AMPK signaling pathway. Am J
Cancer Res. 6:498–508. 2016.PubMed/NCBI
|
|
91
|
Wei L, Liu JJ, Cao J, Du NC, Ji LN and
Yang XL: Role of autophagy in quercetin-induced apoptosis in human
bladder carcinoma BIU-87 cells. Zhonghua Zhong Liu Za Zhi.
34:414–418. 2012.PubMed/NCBI(In Chinese).
|
|
92
|
Tan DQ and Liu XH: Mechanism in growth
inhibition of quercetin on human bladder cancer cell line. Zhongguo
Zhong Yao Za Zhi. 42:1742–1746. 2017.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
93
|
Rockenbach L, Bavaresco L, Fernandes
Farias P, Cappellari AR, Barrios CH, Bueno Morrone F and Oliveira
Battastini AM: Alterations in the extracellular catabolism of
nucleotides are involved in the antiproliferative effect of
quercetin in human bladder cancer T24 cells. Urol Oncol.
31:1204–1211. 2013.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Berger SI and Iyengar R: Network analyses
in systems pharmacology. Bioinformatics. 25:2466–2472.
2009.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Dong Y, Hao L, Fang K, Han XX, Yu H, Zhang
JJ, Cai LJ, Fan T, Zhang WD, Pang K, et al: A network pharmacology
perspective for deciphering potential mechanisms of action of
Solanum nigrum L. in bladder cancer. BMC Complement Med Ther.
21(45)2021.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Cho CJ, Yu CP, Wu CL, Ho JY, Yang CW and
Yu DS: Decreased drug resistance of bladder cancer using
phytochemicals treatment. Kaohsiung J Med Sci. 37:128–135.
2021.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Oršolić N, Odeh D, Jembrek MJ, Knežević J
and Kučan D: Interactions between cisplatin and quercetin at
physiological and hyperthermic conditions on cancer cells in vitro
and in vivo. Molecules. 25(3271)2020.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Lee YH and Tuyet PT: Synthesis and
biological evaluation of quercetin-zinc (II) complex for
anti-cancer and anti-metastasis of human bladder cancer cells. In
Vitro Cell Dev Biol Anim. 55:395–404. 2019.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Tao T, He C, Deng J, Huang Y, Su Q, Peng
M, Yi M, Darko KO, Zou H and Yang X: A novel synthetic derivative
of quercetin,
8-trifluoromethyl-3,5,7,3',4'-O-pentamethyl-quercetin, inhibits
bladder cancer growth by targeting the AMPK/mTOR signaling pathway.
Oncotarget. 8:71657–71671. 2017.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Alban L, Monteiro WF, Diz FM, Miranda GM,
Scheid CM, Zotti ER, Morrone FB and Ligabue R: New quercetin-coated
titanate nanotubes and their radiosensitization effect on human
bladder cancer. Mater Sci Eng C Mater Biol Appl.
110(110662)2020.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Shui L, Wang W, Xie M, Ye B, Li X, Liu Y
and Zheng M: Isoquercitrin induces apoptosis and autophagy in
hepatocellular carcinoma cells via AMPK/mTOR/p70S6K signaling
pathway. Aging (Albany NY). 12:24318–24332. 2020.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Chen F, Chen X, Yang D, Che X, Wang J, Li
X, Zhang Z, Wang Q, Zheng W, Wang L, et al: Isoquercitrin inhibits
bladder cancer progression in vivo and in vitro by regulating the
PI3K/Akt and PKC signaling pathways. Oncol Rep. 36:165–172.
2016.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Wu P, Liu S, Su J, Chen J, Li L, Zhang R
and Chen T: Apoptosis triggered by isoquercitrin in bladder cancer
cells by activating the AMPK-activated protein kinase pathway. Food
Funct. 8:3707–3722. 2017.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Ran J, Wang Y, Zhang W, Ma M and Zhang H:
Research on the bioactivity of isoquercetin extracted from
marestail on bladder cancer EJ cell and the mechanism of its
occurrence. Artif Cells Nanomed Biotechnol. 44:859–864.
2016.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Imran M, Salehi B, Sharifi-Rad J, Aslam
Gondal T, Saeed F, Imran A, Shahbaz M, Tsouh Fokou PV, Umair Arshad
M, Khan H, et al: Kaempferol: A key emphasis to its anticancer
potential. Molecules. 24(2277)2019.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Qiu W, Lin J, Zhu Y, Zhang J, Zeng L, Su M
and Tian Y: Kaempferol modulates DNA methylation and downregulates
DNMT3B in bladder cancer. Cell Physiol Biochem. 41:1325–1335.
2017.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Wu P, Meng X, Zheng H, Zeng Q, Chen T,
Wang W, Zhang X and Su J: Kaempferol attenuates ROS-Induced
hemolysis and the molecular mechanism of its induction of apoptosis
on bladder cancer. Molecules. 23(2592)2018.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Dang Q, Song W, Xu D, Ma Y, Li F, Zeng J,
Zhu G, Wang X, Chang LS, He D and Li L: Kaempferol suppresses
bladder cancer tumor growth by inhibiting cell proliferation and
inducing apoptosis. Mol Carcinog. 54:831–840. 2015.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Xie F, Su M, Qiu W, Zhang M, Guo Z, Su B,
Liu J, Li X and Zhou L: Kaempferol promotes apoptosis in human
bladder cancer cells by inducing the tumor suppressor, PTEN. Int J
Mol Sci. 14:21215–21226. 2013.PubMed/NCBI View Article : Google Scholar
|
|
110
|
DE Oliveira DT, Savio AL, Marcondes JP,
Barros TM, Barbosa LC, Salvadori DM and DA Silva GN: Cytotoxic and
toxicogenomic effects of silibinin in bladder cancer cells with
different TP53 status. J Biosci. 42:91–101. 2017.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Barros TMB, Lima APB, Almeida TC and da
Silva GN: Inhibition of urinary bladder cancer cell proliferation
by silibinin. Environ Mol Mutagen. 61:445–455. 2020.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Li F, Sun Y, Jia J, Yang C, Tang X, Jin B,
Wang K, Guo P, Ma Z, Chen Y, et al: Silibinin attenuates
TGF-β1-induced migration and invasion via EMT suppression and is
associated with COX-2 downregulation in bladder transitional cell
carcinoma. Oncol Rep. 40:3543–3550. 2018.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Wu K, Ning Z, Zeng J, Fan J, Zhou J, Zhang
T, Zhang L, Chen Y, Gao Y, Wang B, et al: Silibinin inhibits
β-catenin/ZEB1 signaling and suppresses bladder cancer metastasis
via dual-blocking epithelial-mesenchymal transition and stemness.
Cell Signal. 25:2625–2633. 2013.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Imai-Sumida M, Chiyomaru T, Majid S, Saini
S, Nip H, Dahiya R, Tanaka Y and Yamamura S: Silibinin suppresses
bladder cancer through down-regulation of actin cytoskeleton and
PI3K/Akt signaling pathways. Oncotarget. 8:92032–92042.
2017.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Sun Y, Guan Z, Zhao W, Jiang Y, Li Q,
Cheng Y and Xu Y: Silibinin suppresses bladder cancer cell
malignancy and chemoresistance in an NF-κB signal-dependent and
signal-independent manner. Int J Oncol. 51:1219–1226.
2017.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Prack Mc Cormick B, Langle Y, Belgorosky
D, Vanzulli S, Balarino N, Sandes E and Eiján AM: Flavonoid silybin
improves the response to radiotherapy in invasive bladder cancer. J
Cell Biochem. 119:5402–5412. 2018.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Gándara L, Sandes E, Di Venosa G, Prack Mc
Cormick B, Rodriguez L, Mamone L, Batlle A, Eiján AM and Casas A:
The natural flavonoid silybin improves the response to Photodynamic
Therapy of bladder cancer cells. J Photochem Photobiol B.
133:55–64. 2014.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Ramchandani S, Naz I, Lee JH, Khan MR and
Ahn KS: An Overview of the potential antineoplastic effects of
casticin. Molecules. 25(1287)2020.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Xu H, Shi HL, Hao JW, Shu KP, Zhang YT and
Hou TQ: Casticin inhibits the proliferation, migration and invasion
of bladder cancer cells by inhibition of TM7SF4 expression.
Zhonghua Zhong Liu Za Zhi. 44:334–340. 2022.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
120
|
Huang AC, Cheng YD, Huang LH, Hsiao YT,
Peng SF, Lu KW, Lien JC, Yang JL, Lin TS and Chung JG: Casticin
induces DNA damage and impairs DNA repair in human bladder cancer
TSGH-8301 cells. Anticancer Res. 39:1839–1847. 2019.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Chung YH and Kim D: RIP kinase-mediated
ROS production triggers XAF1 expression through activation of TAp73
in casticin-treated bladder cancer cells. Oncol Rep. 36:1135–1142.
2016.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Gao X, Xu J, Jiang L, Liu W, Hong H, Qian
Y, Li S, Huang W, Zhao H, Yang Z, et al: Morin alleviates aflatoxin
B1-induced liver and kidney injury by inhibiting heterophil
extracellular traps release, oxidative stress and inflammatory
responses in chicks. Poult Sci. 100(101513)2021.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Shin SS, Won SY, Noh DH, Hwang B, Kim WJ
and Moon SK: Morin inhibits proliferation, migration, and invasion
of bladder cancer EJ cells via modulation of signaling pathways,
cell cycle regulators, and transcription factor-mediated MMP-9
expression. Drug Dev Res. 78:81–90. 2017.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Pan XW, Li L, Huang Y, Huang H, Xu DF, Gao
Y, Chen L, Ren JZ, Cao JW, Hong Y and Cui XG: Icaritin acts
synergistically with epirubicin to suppress bladder cancer growth
through inhibition of autophagy. Oncol Rep. 35:334–342.
2016.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Stevens Y, Rymenant EV, Grootaert C, Camp
JV, Possemiers S, Masclee A and Jonkers D: The intestinal fate of
citrus flavanones and their effects on gastrointestinal health.
Nutrients. 11(1464)2019.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Kim DI, Lee SJ, Lee SB, Park K, Kim WJ and
Moon SK: Requirement for Ras/Raf/ERK pathway in naringin-induced
G1-cell-cycle arrest via p21WAF1 expression. Carcinogenesis.
29:1701–1709. 2008.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Liao AC, Kuo CC, Huang YC, Yeh CW, Hseu
YC, Liu JY and Hsu LS: Naringenin inhibits migration of bladder
cancer cells through downregulation of AKT and MMP-2. Mol Med Rep.
10:1531–1536. 2014.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Juhem A, Boumendjel A, Touquet B, Guillot
A, Popov A, Ronot X and Martel-Frachet V: AG11, a novel
dichloroflavanone derivative with anti-mitotic activity towards
human bladder cancer cells. Anticancer Res. 33:4445–4452.
2013.PubMed/NCBI
|
|
129
|
Khan N, Afaq F and Mukhtar H: Cancer
chemoprevention through dietary antioxidants: Progress and promise.
Antioxid Redox Signal. 10:475–510. 2008.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Khan N and Mukhtar H: Tea polyphenols in
promotion of human health. Nutrients. 11(39)2018.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Bernatoniene J and Kopustinskiene DM: The
role of catechins in cellular responses to oxidative stress.
Molecules. 23(965)2018.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Khan N, Afaq F, Saleem M, Ahmad N and
Mukhtar H: Targeting multiple signaling pathways by green tea
polyphenol (-)-epigallocatechin-3-gallate. Cancer Res.
66:2500–2505. 2006.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Yasuda T, Miyata Y, Nakamura Y, Sagara Y,
Matsuo T, Ohba K and Sakai H: High Consumption of Green tea
suppresses urinary tract recurrence of urothelial cancer via
down-regulation of human antigen-R expression in never smokers. In
Vivo. 32:721–729. 2018.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Matsuo T, Miyata Y, Asai A, Sagara Y,
Furusato B, Fukuoka J and Sakai H: Green tea polyphenol induces
changes in cancer-related factors in an animal model of bladder
cancer. PLoS One. 12(e0171091)2017.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Chen Z, Yu T, Zhou B, Wei J, Fang Y, Lu J,
Guo L, Chen W, Liu ZP and Luo J: Mg(II)-Catechin nanoparticles
delivering siRNA targeting EIF5A2 inhibit bladder cancer cell
growth in vitro and in vivo. Biomaterials. 81:125–134.
2016.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Jankun J, Keck RW and Selman SH:
Epigallocatechin-3-gallate prevents tumor cell implantation/growth
in an experimental rat bladder tumor model. Int J Oncol.
44:147–152. 2014.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Lee HY, Chen YJ, Chang WA, Li WM, Ke HL,
Wu WJ and Kuo PL: Effects of epigallocatechin gallate (EGCG) on
urinary bladder urothelial carcinoma-next-generation sequencing and
bioinformatics approaches. Medicina (Kaunas).
55(768)2019.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Luo KW, Wei Chen, Lung WY, Wei XY, Cheng
BH, Cai ZM and Huang WR: EGCG inhibited bladder cancer SW780 cell
proliferation and migration both in vitro and in vivo via
down-regulation of NF-κB and MMP-9. J Nutr Biochem. 41:56–64.
2017.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Luo KW, Lung WY, Chun-Xie Luo XL and Huang
WR: EGCG inhibited bladder cancer T24 and 5637 cell proliferation
and migration via PI3K/AKT pathway. Oncotarget. 9:12261–12272.
2018.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Qin J, Wang Y, Bai Y, Yang K, Mao Q, Lin
Y, Kong D, Zheng X and Xie L: Epigallocatechin-3-gallate inhibits
bladder cancer cell invasion via suppression of NF-κB-mediated
matrix metalloproteinase-9 expression. Mol Med Rep. 6:1040–1044.
2012.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Feng C, Ho Y, Sun C, Xia G, Ding Q and Gu
B: Epigallocatechin gallate inhibits the growth and promotes the
apoptosis of bladder cancer cells. Exp Ther Med. 14:3513–3518.
2017.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Yin Z, Li J, Kang L, Liu X, Luo J, Zhang
L, Li Y and Cai J: Epigallocatechin-3-gallate induces
autophagy-related apoptosis associated with LC3B II and Beclin
expression of bladder cancer cells. J Food Biochem.
45(e13758)2021.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Sun X, Song J, Li E, Geng H, Li Y, Yu D
and Zhong C: (-)-Epigallocatechin-3-gallate inhibits bladder cancer
stem cells via suppression of sonic hedgehog pathway. Oncol Rep.
42:425–435. 2019.PubMed/NCBI View Article : Google Scholar
|
|
144
|
Luo KW, Zhu XH, Zhao T, Zhong J, Gao HC,
Luo XL and Huang WR: EGCG enhanced the anti-tumor effect of
doxorubicine in bladder cancer via NF-κB/MDM2/p53 pathway. Front
Cell Dev Biol. 8(606123)2020.PubMed/NCBI View Article : Google Scholar
|
|
145
|
Mottaghipisheh J, Doustimotlagh AH, Irajie
C, Tanideh N, Barzegar A and Iraji A: The promising therapeutic and
preventive properties of anthocyanidins/anthocyanins on prostate
cancer. Cells. 11(1070)2022.PubMed/NCBI View Article : Google Scholar
|
|
146
|
Alappat B and Alappat J: Anthocyanin
pigments: Beyond aesthetics. Molecules. 25(5500)2020.PubMed/NCBI View Article : Google Scholar
|
|
147
|
Higgins JA, Zainol M, Brown K and Jones
GD: Anthocyans as tertiary chemopreventive agents in bladder
cancer: Anti-oxidant mechanisms and interaction with mitomycin C.
Mutagenesis. 29:227–235. 2014.PubMed/NCBI View Article : Google Scholar
|
|
148
|
Li WL, Ji GH, Zhang XZ and Yu HY: The
influence and mechanisms of purple sweet potato anthocyanins on the
growth of bladder cancer BIU87 cell. Zhonghua Yi Xue Za Zhi.
98:457–459. 2018.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
149
|
Li WL, Yu HY, Zhang XJ, Ke M and Hong T:
Purple sweet potato anthocyanin exerts antitumor effect in bladder
cancer. Oncol Rep. 40:73–82. 2018.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Yang N, Gao J, Hou R, Xu X, Yang N and
Huang S: Grape seed proanthocyanidins inhibit migration and
invasion of bladder cancer cells by reversing EMT through
suppression of TGF-β signaling pathway. Oxid Med Cell Longev.
2021(5564312)2021.PubMed/NCBI View Article : Google Scholar
|
|
151
|
Fishman AI, Johnson B, Alexander B, Won J,
Choudhury M and Konno S: Additively enhanced antiproliferative
effect of interferon combined with proanthocyanidin on bladder
cancer cells. J Cancer. 3:107–112. 2012.PubMed/NCBI View Article : Google Scholar
|
|
152
|
Liu J, Zhang WY, Kong ZH and Ding DG:
Induction of cell cycle arrest and apoptosis by grape seed
procyanidin extract in human bladder cancer BIU87 cells. Eur Rev
Med Pharmacol Sci. 20:3282–3291. 2016.PubMed/NCBI
|
|
153
|
Křížová L, Dadáková K, Kašparovská J and
Kašparovský T: Isoflavones. Molecules. 24(1076)2019.PubMed/NCBI View Article : Google Scholar
|
|
154
|
He Y, Wu X, Cao Y, Hou Y, Chen H, Wu L, Lu
L, Zhu W and Gu Y: Daidzein exerts anti-tumor activity against
bladder cancer cells via inhibition of FGFR3 pathway. Neoplasma.
63:523–531. 2016.PubMed/NCBI View Article : Google Scholar
|
|
155
|
Russo M, Russo GL, Daglia M, Kasi PD, Ravi
S, Nabavi SF and Nabavi SM: Understanding genistein in cancer: The
‘good’ and the ‘bad’ effects: A review. Food Chem. 196:589–600.
2016.PubMed/NCBI View Article : Google Scholar
|
|
156
|
Park C, Cha HJ, Lee H, Hwang-Bo H, Ji SY,
Kim MY, Hong SH, Jeong JW, Han MH, Choi SH, et al: Induction of
G2/M cell cycle arrest and apoptosis by genistein in human bladder
cancer T24 cells through Inhibition of the ROS-Dependent PI3k/Akt
signal transduction pathway. Antioxidants (Basel).
8(327)2019.PubMed/NCBI View Article : Google Scholar
|
|
157
|
Wang Y, Wang H, Zhang W, Shao C, Xu P, Shi
CH, Shi JG, Li YM, Fu Q, Xue W, et al: Genistein sensitizes bladder
cancer cells to HCPT treatment in vitro and in vivo via
ATM/NF-κB/IKK pathway-induced apoptosis. PLoS One.
8(e50175)2013.PubMed/NCBI View Article : Google Scholar
|
|
158
|
Köksal Karayildirim Ç, Nalbantsoy A and
Karabay Yavaşoğlu NU: Prunetin inhibits nitric oxide activity and
induces apoptosis in urinary bladder cancer cells via CASP3 and
TNF-α genes. Mol Biol Rep. 48:7251–7259. 2021.PubMed/NCBI View Article : Google Scholar
|
|
159
|
Zhou YX, Zhang H and Peng C: Puerarin: A
review of pharmacological effects. Phytother Res. 28:961–975.
2014.PubMed/NCBI View Article : Google Scholar
|
|
160
|
Jiang K, Chen H, Tang K, Guan W, Zhou H,
Guo X, Chen Z, Ye Z and Xu H: Puerarin inhibits bladder cancer cell
proliferation through the mTOR/p70S6K signaling pathway. Oncol
Lett. 15:167–174. 2018.PubMed/NCBI View Article : Google Scholar
|
|
161
|
Ye G, Kan S, Chen J and Lu X: Puerarin in
inducing apoptosis of bladder cancer cells through inhibiting
SIRT1/p53 pathway. Oncol Lett. 17:195–200. 2019.PubMed/NCBI View Article : Google Scholar
|
|
162
|
Jiang QQ, Liu B and Yuan T: MicroRNA-16
inhibits bladder cancer proliferation by targeting Cyclin D1. Asian
Pac J Cancer Prev. 14:4127–4130. 2013.PubMed/NCBI View Article : Google Scholar
|
|
163
|
Liu X, Li S, Li Y, Cheng B, Tan B and Wang
G: Puerarin inhibits proliferation and induces apoptosis by
upregulation of miR-16 in bladder cancer cell line T24. Oncol Res.
26:1227–1234. 2018.PubMed/NCBI View Article : Google Scholar
|
|
164
|
Du L, Zhang L and Sun F: Puerarin inhibits
the progression of bladder cancer by regulating
circ_0020394/miR-328-3p/NRBP1 axis. Cancer Biother Radiopharm.
37:435–450. 2020.PubMed/NCBI View Article : Google Scholar
|
|
165
|
Wu Y, Zhang X, Li Z, Yan H, Qin J and Li
T: Formononetin inhibits human bladder cancer cell proliferation
and invasiveness via regulation of miR-21 and PTEN. Food Funct.
8:1061–1066. 2017.PubMed/NCBI View Article : Google Scholar
|
|
166
|
Ouyang Y, Li J, Chen X, Fu X, Sun S and Wu
Q: Chalcone derivatives: Role in anticancer therapy. Biomolecules.
11(894)2021.PubMed/NCBI View Article : Google Scholar
|
|
167
|
Yuan X, Li D, Zhao H, Jiang J, Wang P, Ma
X, Sun X and Zheng Q: Licochalcone A-induced human bladder cancer
T24 cells apoptosis triggered by mitochondria dysfunction and
endoplasmic reticulum stress. Biomed Res Int.
2013(474272)2013.PubMed/NCBI View Article : Google Scholar
|
|
168
|
Yang X, Jiang J, Yang X, Han J and Zheng
Q: Licochalcone A induces T24 bladder cancer cell apoptosis by
increasing intracellular calcium levels. Mol Med Rep. 14:911–919.
2016.PubMed/NCBI View Article : Google Scholar
|
|
169
|
Jiang J, Yuan X, Zhao H, Yan X, Sun X and
Zheng Q: Licochalcone A inhibiting proliferation of bladder cancer
T24 cells by inducing reactive oxygen species production. Biomed
Mater Eng. 24:1019–1025. 2014.PubMed/NCBI View Article : Google Scholar
|
|
170
|
Hong SH, Cha HJ, Hwang-Bo H, Kim MY, Kim
SY, Ji SY, Cheong J, Park C, Lee H, Kim GY, et al:
Anti-proliferative and pro-apoptotic effects of licochalcone A
through ROS-Mediated cell cycle arrest and apoptosis in human
bladder cancer cells. Int J Mol Sci. 20(3820)2019.PubMed/NCBI View Article : Google Scholar
|
|
171
|
Zhao H, Yuan X, Jiang J, Wang P, Sun X,
Wang D and Zheng Q: Antimetastatic effects of licochalcone B on
human bladder carcinoma T24 by inhibition of matrix
metalloproteinases-9 and NF-кB activity. Basic Clin Pharmacol
Toxicol. 115:527–533. 2014.PubMed/NCBI View Article : Google Scholar
|
|
172
|
Yuan X, Li T, Xiao E, Zhao H, Li Y, Fu S,
Gan L and Wang Z, Zheng Q and Wang Z: Licochalcone B inhibits
growth of bladder cancer cells by arresting cell cycle progression
and inducing apoptosis. Food Chem Toxicol. 65:242–251.
2014.PubMed/NCBI View Article : Google Scholar
|
|
173
|
Wang P, Yuan X, Wang Y, Zhao H, Sun X and
Zheng Q: Licochalcone C induces apoptosis via B-cell lymphoma 2
family proteins in T24 cells. Mol Med Rep. 12:7623–7628.
2015.PubMed/NCBI View Article : Google Scholar
|
|
174
|
Wang KL, Yu TC and Hsia SM: Perspectives
on the role of isoliquiritigenin in cancer. Cancers (Basel).
13(115)2021.PubMed/NCBI View Article : Google Scholar
|
|
175
|
Patricia Moreno-Londoño A, Bello-Alvarez C
and Pedraza-Chaverri J: Isoliquiritigenin pretreatment attenuates
cisplatin induced proximal tubular cells (LLC-PK1) death and
enhances the toxicity induced by this drug in bladder cancer T24
cell line. Food Chem Toxicol. 109(Pt 1):143–154. 2017.PubMed/NCBI View Article : Google Scholar
|
|
176
|
Li X, Xu X, Ji T, Liu Z, Gu M, Hoang BH
and Zi X: Dietary feeding of Flavokawain A, a Kava chalcone,
exhibits a satisfactory safety profile and its association with
enhancement of phase II enzymes in mice. Toxicol Rep. 1:2–11.
2014.PubMed/NCBI View Article : Google Scholar
|
|
177
|
Liu Z, Xu X, Li X, Liu S, Simoneau AR, He
F, Wu XR and Zi X: Kava chalcone, flavokawain A, inhibits
urothelial tumorigenesis in the UPII-SV40T transgenic mouse model.
Cancer Prev Res (Phila). 6:1365–1375. 2013.PubMed/NCBI View Article : Google Scholar
|
|
178
|
Liu Z, Song L, Xie J, Simoneau AR, Uchio E
and Zi X: Chemoprevention of urothelial cell carcinoma
tumorigenesis by dietary flavokawain A in UPII-Mutant Ha-ras
transgenic mice. Pharmaceutics. 14(496)2022.PubMed/NCBI View Article : Google Scholar
|
|
179
|
Wu CM, Lin KW, Teng CH, Huang AM, Chen YC,
Yen MH, Wu WB, Pu YS and Lin CN: Chalcone derivatives inhibit human
platelet aggregation and inhibit growth in human bladder cancer
cells. Biol Pharm Bull. 37:1191–1198. 2014.PubMed/NCBI View Article : Google Scholar
|
|
180
|
Martel-Frachet V, Keramidas M, Nurisso A,
DeBonis S, Rome C, Coll JL, Boumendjel A, Skoufias DA and Ronot X:
IPP51, a chalcone acting as a microtubule inhibitor with in vivo
antitumor activity against bladder carcinoma. Oncotarget.
6:14669–14686. 2015.PubMed/NCBI View Article : Google Scholar
|
|
181
|
Martel-Frachet V, Areguian J, Blanc M,
Touquet B, Lamarca A, Ronot X and Boumendjel A: Investigation of a
new 1,3-diarylpropenone as a potential antimitotic agent targeting
bladder carcinoma. Anticancer Drugs. 20:469–476. 2009.PubMed/NCBI View Article : Google Scholar
|
|
182
|
Desilets A, Adam JP and Soulières D:
Management of cisplatin-associated toxicities in bladder cancer
patients. Curr Opin Support Palliat Care. 14:286–292.
2020.PubMed/NCBI View Article : Google Scholar
|
|
183
|
Cai Z, Zhang F, Chen W, Zhang J and Li H:
MiRNAs: A promising target in the chemoresistance of bladder
cancer. Onco Targets Ther. 12:11805–11816. 2019.PubMed/NCBI View Article : Google Scholar
|
|
184
|
Dobrzynska M, Napierala M and Florek E:
Flavonoid nanoparticles: A promising approach for cancer therapy.
Biomolecules. 10(1268)2020.PubMed/NCBI View Article : Google Scholar
|
|
185
|
Sun T, Zhang YS, Pang B, Hyun DC, Yang M
and Xia Y: Engineered nanoparticles for drug delivery in cancer
therapy. Angew Chem Int Ed Engl. 53:12320–12364. 2014.PubMed/NCBI View Article : Google Scholar
|
|
186
|
Kim S, Chen J, Cheng T, Gindulyte A, He J,
He S, Li Q, Shoemaker BA, Thiessen PA, Yu B, et al: PubChem in
2021: New data content and improved web interfaces. Nucleic Acids
Res. 49(D1):D1388–D1395. 2021.PubMed/NCBI View Article : Google Scholar
|
|
187
|
Patil M, Pabla N and Dong Z: Checkpoint
kinase 1 in DNA damage response and cell cycle regulation. Cell Mol
Life Sci. 70:4009–4021. 2013.PubMed/NCBI View Article : Google Scholar
|
|
188
|
Schmitt E, Paquet C, Beauchemin M and
Bertrand R: DNA-damage response network at the crossroads of
cell-cycle checkpoints, cellular senescence and apoptosis. J
Zhejiang Univ Sci B. 8:377–397. 2007.PubMed/NCBI View Article : Google Scholar
|
|
189
|
Yang L, Shi P, Zhao G, Xu J, Peng W, Zhang
J, Zhang G, Wang X, Dong Z, Chen F and Cui H: Targeting cancer stem
cell pathways for cancer therapy. Signal Transduct Target Ther.
5(8)2020.PubMed/NCBI View Article : Google Scholar
|