Introduction
Diabetic nephropathy (DN), clinically manifested as
persistent albuminuria and glomerular filtration rate decrement, is
a common complication of diabetes; in detail, DN prevalence in
diabetes patients ranges from 18.7-24.0% in China (1-3).
At present, the management of DN mainly contains blood
glucose/pressure control and lipid control (4,5).
Nevertheless, DN remains the leading reason for end-stage kidney
disease; among which irreversible fibrosis, excessive proliferation
and inflammation flare in renal glomerular basement membrane cells
and mesangial cells are the main pathological manifestations during
the progression of DN (6-8).
Hence, exploring the underlying mechanism of these pathological
alterations may help improve DN management more effectively.
Dual specificity phosphatase (DUSP) 22, also known
as Jun N-terminal kinase pathway-associated phosphatase, is a
tyrosine-specific protein participating in several cellular
processes (including cell proliferation and apoptosis) due to its
unique function of dephosphorylating serine/threonine (9-11).
For instance, a previous study revealed that DUSP22-knockdown T
cells accelerate dysregulation of inflammatory cytokines (11). Another study found that DUSP22
regulates the transcription of interleukin (IL)-6 and inflammation
response via dephosphorylating signal transducer and activator of
transcription 3 (STAT3) (10).
Meanwhile, DUSP22 is an important regulator of the
mitogen-activated protein kinases (MAPKs), while the activation of
MAPKs mediates mesangial cell apoptosis and tubulointerstitial
fibrosis (12,13). Combining that inflammation and
glomerular fibrosis are implicated in DN pathogenesis, it was
hypothesized that DUSP22 may serve as a protective factor of DN,
while it has not been studied yet.
Hence, the current study aimed to assess the effect
of DUSP22 on cell proliferation, apoptosis, fibrosis, inflammation
and its potential mediated signaling pathway in mouse mesangial
cell line (SV40-MES13) under both high glucose (HG) and low glucose
(LG) conditions.
Materials and methods
Cell culture
Considering that the SV40-MES13 cells were commonly
used to establish the cellular DN model according to previous
studies, the same cell line was chosen in the present study
(14-17).
SV40-MES13 was obtained from National Collection of Authenticated
Cell Cultures (Shanghai, China). Cells were maintained in DMEM/F12
medium (HyClone; Cytiva) supplemented with 5% fetal bovine serum
(Merck KGaA) and 1% penicillin/streptomycin (Beyotime Institute of
Biotechnology) at 37˚C and 5% CO2.
Cell transfection
DUSP22 overexpression and control plasmids were
purchased from Sangon Biotech Co., Ltd. DUSP22 and control small
interfering (si) RNAs were synthesized by Shanghai GenePharma Co.,
Ltd. Briefly, SV40-MES13 cells were seeded in six-well
(2x105 cells/well) or 96-well plate (5x103
cells/well) and cultured into 80% confluence. Cells were then
transfected (100 or 5 pM) into cells using HilyMAX Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) at 37˚C for 6 h
according to the manufacturers' protocol. The sense sequences of
siRNAs were as follows: DUSP22, 5'-CGGGCCTGTACATTGGCAACTTCAA-3';
and control, 5'-CGGGTCCATTTACGGCAATTGCCAA-3'.
Glucose treatment
The SV40-MES13 cells were seeded in six-well plates
(2x105 cells/well) and divided into HG and LG groups. In
the HG group, SV40-MES13 cells were stimulated with 25 mM D-glucose
(Shanghai Aladdin Biochemical Technology Co., Ltd.) for 48 h to
establish cellular DN model (14-18),
and transfected with DUSP22 overexpression plasmid (HG-oe-DUSP22)
or control plasmid (HG-oe-NC) as aforementioned. In the LG group,
SV40-MES13 cells were stimulated with 5.5 mM D-glucose supplemented
with 19.5 mM D-mannitol (Sigma-Aldrich; Merck KGaA) for 48 h, and
transfected with DUSP22 siRNA (LG-si-DUSP22) or siRNA control
(LG-si-NC) as aforementioned. Untransfected cells cultured in HG or
LG medium were used as control group (HG-Control or LG-Control).
Cells were then incubated for 48 h, and harvested for reverse
transcription-quantitative PCR (RT-qPCR), cell apoptosis and
western blotting assays. The cell supernatant was used for
inflammatory cytokines assessment using ELISA.
Cell proliferation assay
Cell proliferation detection of SV40-MES13 cells was
performed using Cell Counting Kit-8 (CCK-8; Dojindo Laboratories,
Inc.). In brief, cells in the HG or LG group were plated on a
96-well plate (5x103 cells/well) and transfected as
indicated. At 0, 24, 48 and 72 h after transfection, 10 µl CCK-8
detection buffer was added and cells were incubated for 2 h at
37˚C. A microplate reader (Tosoh Corporation) was adopted to assess
cell proliferation with an optical density (OD) value of 450 nm
being measured.
Cell apoptosis assay
The TUNEL detection kit (Elabscience Biotechnology,
Inc.) was used for assessing cell apoptotic rate after treatment.
In brief, SV40-MES13 cells were fixed with 4% paraformaldehyde
(Wuhan Servicebio Technology Co., Ltd.) for 15 min and incubated
with Triton X-100 (Wuhan Servicebio Technology Co., Ltd.) for 10
min at 37˚C. Afterwards, cells were incubated with apoptosis
detection buffer for 0.5 h at 37˚C. After being stained with DAPI
(5 mg/l; Sangon Biotech Co., Ltd.) for 10 min and sealed by
Antifade Mounting Medium (Beyotime Institute of Biotechnology),
cell apoptotic rate was evaluated using fluorescence microscope
(Olympus Corporation) with five random fields being selected.
ELISA
The supernatant of SV40-MES13 cells was collected at
48 h after treatment. The content of inflammatory cytokines in cell
supernatant was detected using mouse tumor necrosis factor-alpha
(TNF-α; cat. no. D721150), IL-1β (cat. no. D721017), IL-6 (cat. no.
D721022), and IL-12 (cat. no. D721174) ELISA kits (Sangon Biotech
Co., Ltd.), respectively. The experiment was performed in
accordance with the manufacturer's protocol. The OD value at 450 nm
was measured using a microplate reader (Tosoh Corporation).
RNA extraction and RT-qPCR
Total RNA of SV40-MES13 cells was extracted using
RNAzol® RT (Sigma-Aldrich; Merck KGaA). The concentration of RNA
was analyzed using Qubit-4 Flurometer (Invitrogen; Thermo fisher
Scientific, Inc.). Reverse Transcription kit (Qiagen GmbH) was used
for cDNA synthesis in accordance with the kit's protocol. The
quantification of DUSP22, fibronectin 1 (FN1), collagen I (COL1A1)
and transforming growth factor beta 1 (TGF-β1) was performed using
the SYBR Green PCR kit (Qiagen GmbH) and normalized to the level of
β-actin. The thermocycling conditions of qPCR were as follows: 95˚C
for 5 min, 1 cycle; 95˚C for 10 sec, 61˚C for 30 sec, 40 cycles.
The results were calculated using the
2-ΔΔCq method (19). The sequences of primers used for
RT-qPCR were as follows: DUSP22 forward,
5'-GCCAGGCCTATGTTGGAGGGAGTT-3' and reverse,
5'-TGTATGCGATCACCAGTGTCAC-3'; FN1 forward, 5'-ATGTGGACCCCTCCTGATAGT
and reverse, 5'-GCCCAGTGATTTCAGCAAAGG-3'; COL1A1 forward,
5'-CTCGTGGATTGCCTGGAACA-3' and reverse, 5'-GCACCAACAGCACCATCGT-3';
TGF-β1 forward, 5'-TGACGTCACTGGAGTTGTACGG-3' and reverse,
5'-GGTTCATGTCATGGATGGTGC-3'; and β-actin forward,
5'-AAGACCTCTATGCCAACACAGTG-3' and reverse,
5'-CATCGTACTCCTGCTTGCTGAT-3'.
Western blot analysis
SV40-MES13 cells were lysed in RIPA containing 1%
protease and phosphatase inhibitor cocktail (cat. No. P1048;
Beyotime Institute of Biotechnology) for protein extraction. The
protein quantification was performed using the BCA quantification
kit (Beyotime Institute of Biotechnology). A total of 30 µg protein
of each group were separated by 4-20% SDS-PAGE precast gels and
transferred into polyvinylidene difluoride membrane (both from
Beyotime Institute of Biotechnology). The membrane was then blocked
using 5% BSA solution for 1.5 h at 37˚C and incubated overnight at
4˚C with the following primary antibodies: DUSP22 (1:2,000; cat.
no. ab70124), caspase 3 (1:5,000; cat. no. ab184787), cleaved
caspase 3 (1:1,000; cat. no. ab214430; all from Abcam), Bcl2
(1:2,000; cat. no. AF6139), FN1 (1:1,000; cat. no. AF5335), COL1A1
(1:1,000; cat. no. AF7001; all from Affinity Biosciences, Ltd.),
TGF-β1 (1:2,000; cat. no. ab215715; Abcam), p-extracellular
signal-regulated kinase (ERK) (Thr202/Tyr204) (1:1,000; cat. no.
AF1015), ERK (1:1,000; cat. no. AF0155), p-c-Jun N-terminal kinase
(JNK) (Tyr185) (1:2,000; cat. no. AF3318; all from Affinity
Biosciences, Ltd.), JNK (1:1,000; cat. no. GB114321; Wuhan
Servicebio Technology Co., Ltd.), p-P38 (Thr180/Tyr182) (1:1,000;
cat. no. AF4001), P38 (1:1,000; cat. no. AF6456), Ki-67 (1:1,500;
cat. no. AF0198; all from Affinity Biosciences, Ltd.) and β-actin
(1:5,000; cat. no. GB15003; Wuhan Servicebio Technology Co., Ltd.).
Afterwards, the membranes were incubated with HRP-conjugated goat
anti-rabbit secondary antibodies (1:10,000; cat. no. ab205718;
Abcam) for 1 h at 37˚C. Finally, ECL Plus kit (Shanghai Yeasen
Biotechnology Co., Ltd.) was used for chemiluminescence.
Considering the similar molecular weights of caspase 3, cleaved
caspase 3, Bcl2, and β-actin, the membrane was stripped and
re-probed for the second antibody once the first protein band was
visualized (20). The
densitometric analysis was performed using Image J 1.8 (National
Institutes of Health).
Statistical analysis
GraphPad 7.0 software (GraphPad Software, Inc.) was
used for data analysis and graph plotting. Multigroup comparison
was analyzed by one-way ANOVA followed by Tukey's post hoc test.
The data are presented as the mean value ± standard deviation, and
each experiment was replicated for three times. P<0.05 was
considered to indicate a statistically significant difference.
Results
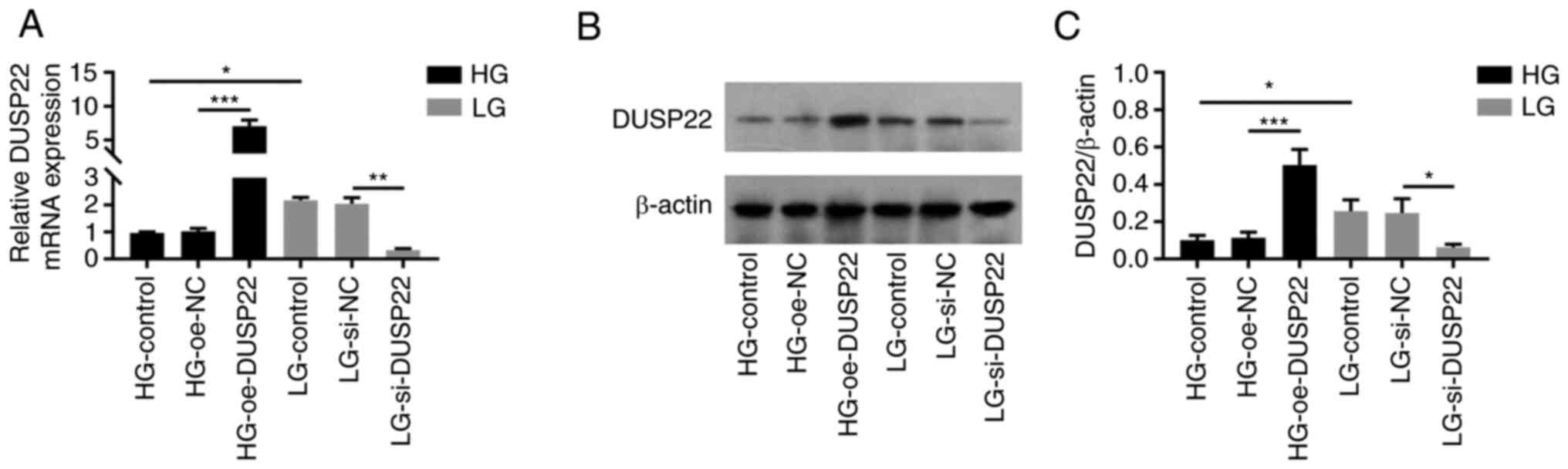
DUSP22 expression in SV40-MES13
cells
DUSP22 mRNA expression was decreased in HG-Control
group compared with LG-Control group (P<0.05); moreover, DUSP22
expression was increased in HG-oe-DUSP22 group compared with that
in HG-oe-NC group (P<0.001), but it was decreased in the
LG-si-DUSP22 group compared with the LG-si-NC group (P<0.01;
Fig. 1A), which indicated that the
transfection was successful. Furthermore, DUSP22 protein expression
showed a similar trend to DUSP22 mRNA expression among groups (all
P<0.05; Fig. 1B and C).
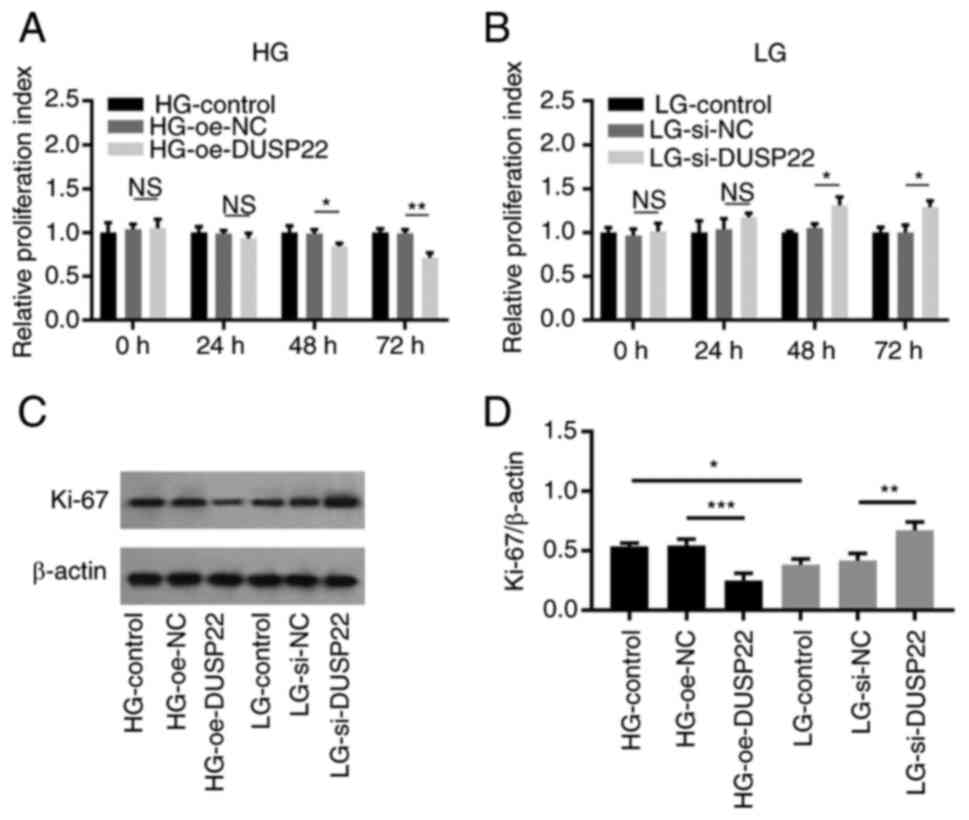
Effect of DUSP22 on SV40-MES13 cell
proliferation
Cell proliferation at 48 (P<0.05) and 72 h
(P<0.01) was reduced in the HG-oe-DUSP22 group compared with
that in the HG-oe-NC group (Fig.
2A). On the contrary, cell proliferation at 48 (P<0.05) and
72 h (P<0.05) was elevated in the LG-si-DUSP22 group compared
with the LG-si-NC group (Fig. 2B).
Additionally, the proliferation biomarker Ki-67 was determined to
further validate the results, which showed that Ki-67 was decreased
in the HG-oe-DUSP22 group compared with the HG-oe-NC group
(P<0.001); while it was increased in the LG-si-DUSP22 group
compared with that in the LG-si-NC group (P<0.01; Fig. 2C and D).
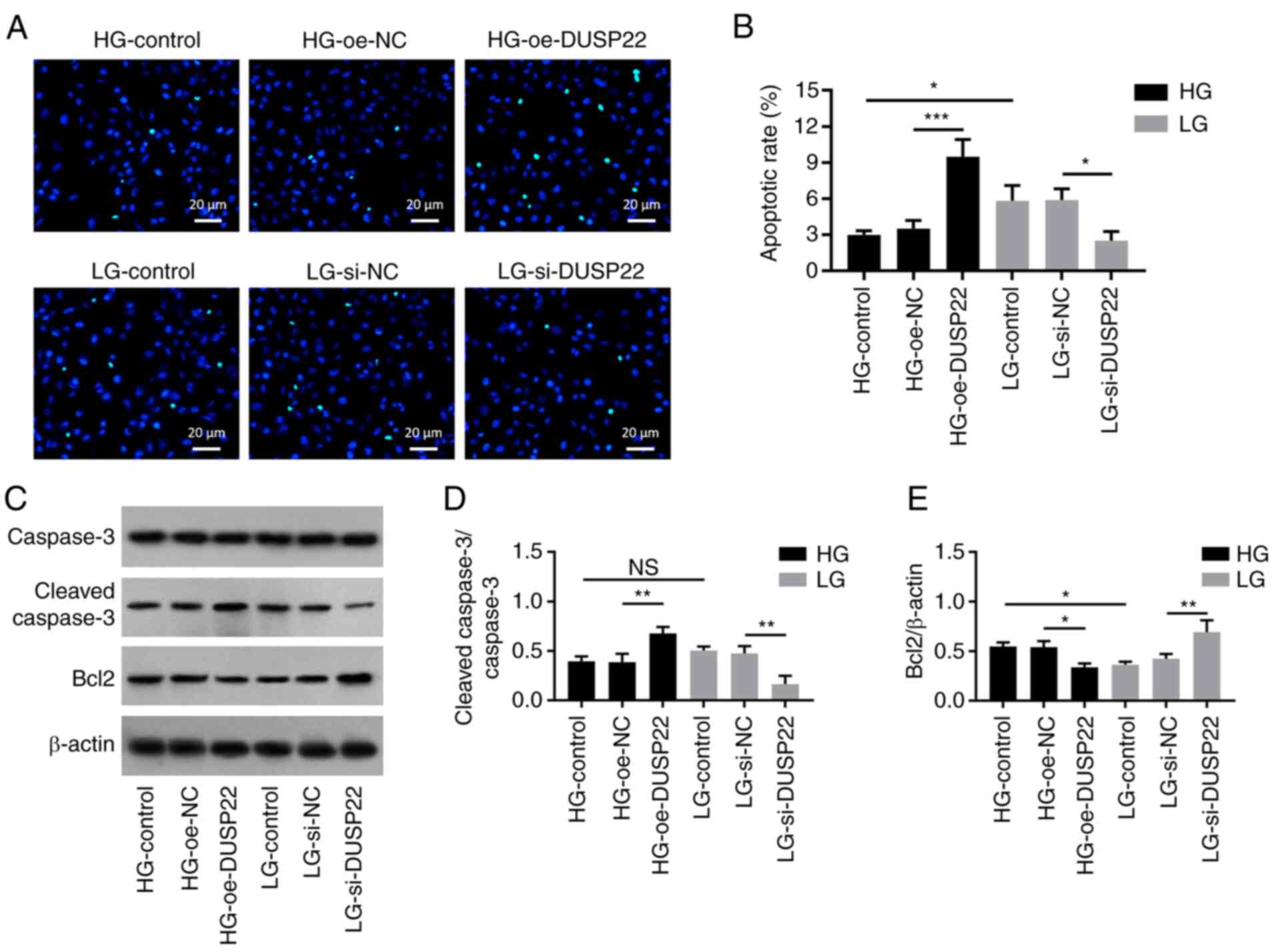
Effect of DUSP22 on SV40-MES13 cell
apoptosis
TUNEL-reflected apoptotic rate (P<0.001; Fig. 3A and B) and the expression of cleaved caspase 3
(P<0.01; Fig. 3C and D) were elevated, while the expression of
Bcl2 (P<0.05; Fig. 3C and
E) was reduced in the HG-oe-DUSP22
group compared with those in the HG-oe-NC group. By contrast,
TUNEL-reflected apoptotic rate (P<0.05) and expression of
cleaved caspase 3 (P<0.01) were decreased, but expression of
Bcl2 (P<0.01) was increased in the LG-si-DUSP22 group compared
with the LG-si-NC group.
Effect of DUSP22 on SV40-MES13 cell
fibrosis
FN1 mRNA (P<0.05; Fig. 4A), COL1A1 mRNA (P<0.01; Fig. 4B) and TGF-β1 mRNA (P<0.001;
Fig. 4C) levels were reduced in
the HG-oe-DUSP22 group compared with the HG-oe-NC group; while they
were upregulated in the LG-si-DUSP22 group compared with the
LG-si-NC group (all P<0.01). In terms of fibrosis-related
proteins (Fig. 4D), FN1 protein
(Fig. 4E), COL1A1 I protein
(Fig. 4F) and TGF-β1 protein
(Fig. 4G) displayed a similar
trend among groups with their corresponding mRNAs (all
P<0.05).
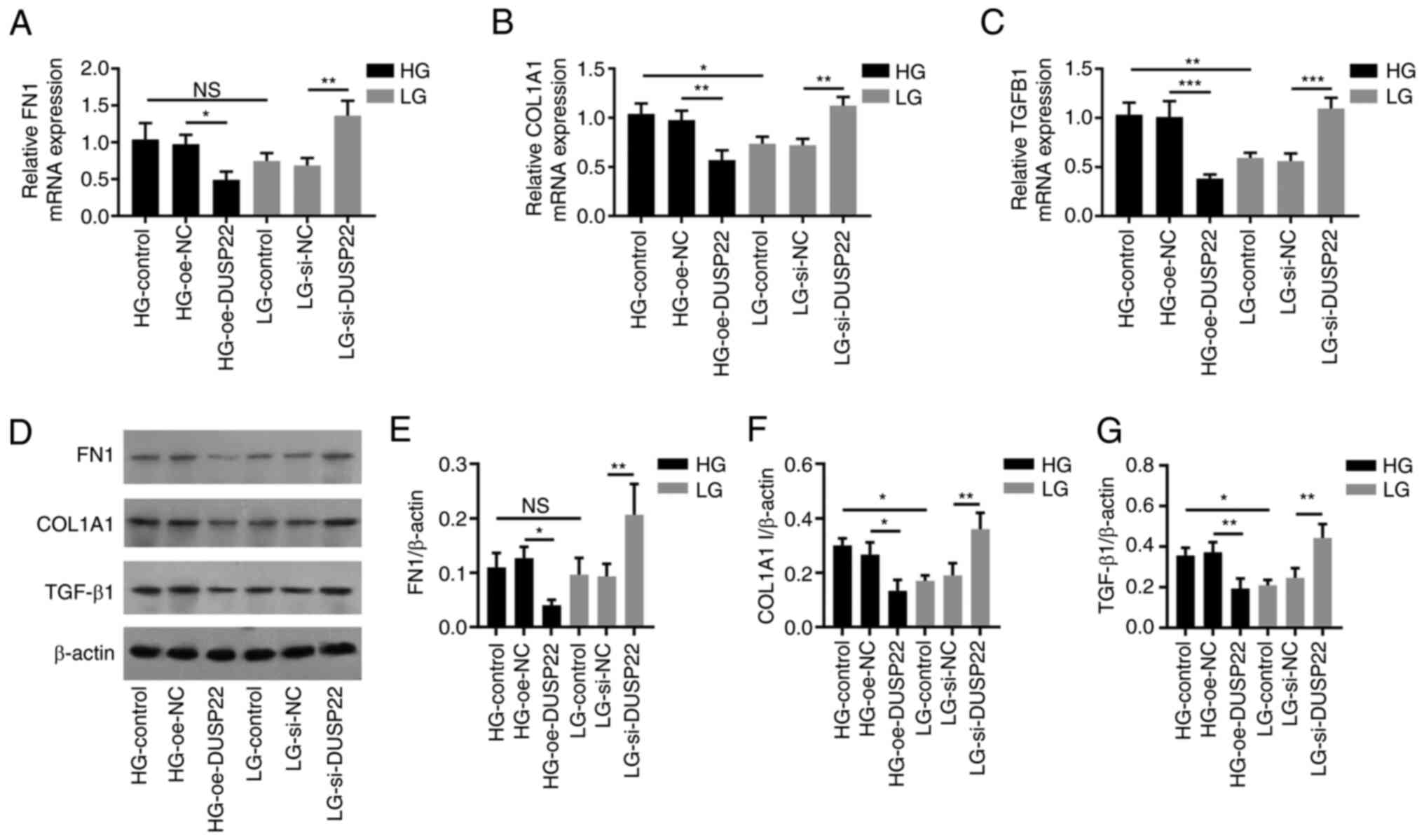 | Figure 4DUSP22 suppresses SV40-MES13 cell
fibrosis. (A-C) Effect of DUSP22 on mRNA levels of (A) FN1, (B)
COL1A1, and (C) TGF-β1 in SV40-MES13 cells. (D) Western blot
analysis revealing the effect of DUSP22 on (D and E) FN1, (D and F)
COL1A1 and (D and G) TGF-β1 in SV40-MES13 cells. (E-G)
Densitometric analysis of western blotting at panel D.
*P<0.05, **P<0.01 and
***P<0.001. DUSP22, dual specificity phosphatase 22;
FN1, fibronectin 1; COL1A1, collagen I; TGF-β1, transforming growth
factor beta 1; HG, high glucose; LG, low glucose; oe,
overexpression; si-, small interfering; NC, negative control; ns,
no significance. |
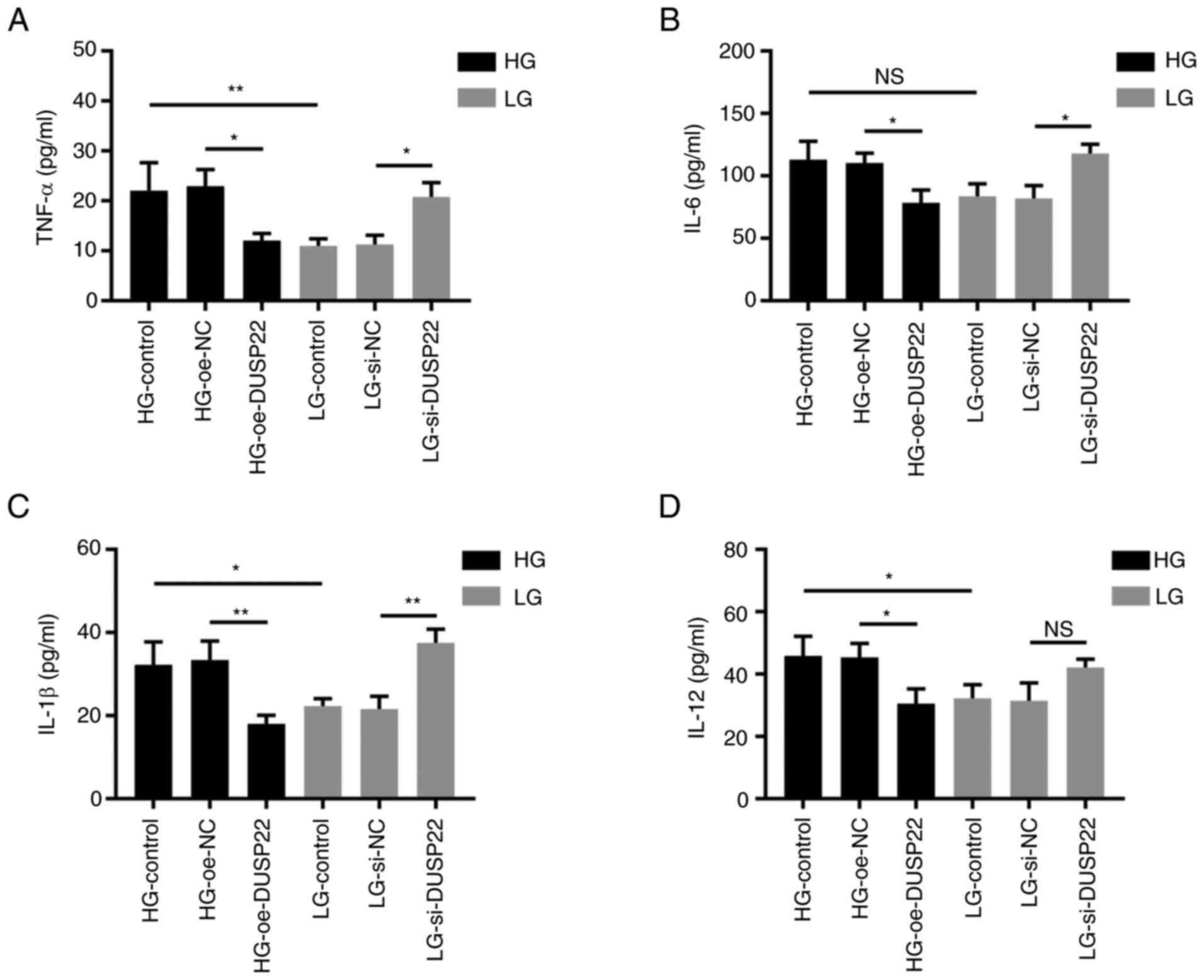
Effect of DUSP22 on SV40-MES13 cell
inflammation
TNF-α (P<0.05; Fig.
5A), IL-6 (P<0.05; Fig.
5B), IL-1β (P<0.01; Fig.
5C) and IL-12 (P<0.05; Fig.
5D) were downregulated in the HG-oe-DUSP22 group compared with
those in the HG-oe-NC group; while TNF-α (P<0.05), IL-6
(P<0.05), and IL-1β (P<0.01) were significantly increased in
the LG-si-DUSP22 group compared with the LG-si-NC, except for IL-12
(P>0.05).
Determination of DUSP22 mediated
pathways
As revealed using western blot analysis (Fig. 6A), p-ERK (P<0.001), p-JNK
(P<0.05) and p-P38 (P<0.01) expression levels were decreased
in the HG-oe-DUSP22 group compared with the HG-oe-NC group; by
contrast, they were increased in the LG-si-DUSP22 group compared
with the LG-si-NC group (all P<0.05; Fig. 6B).
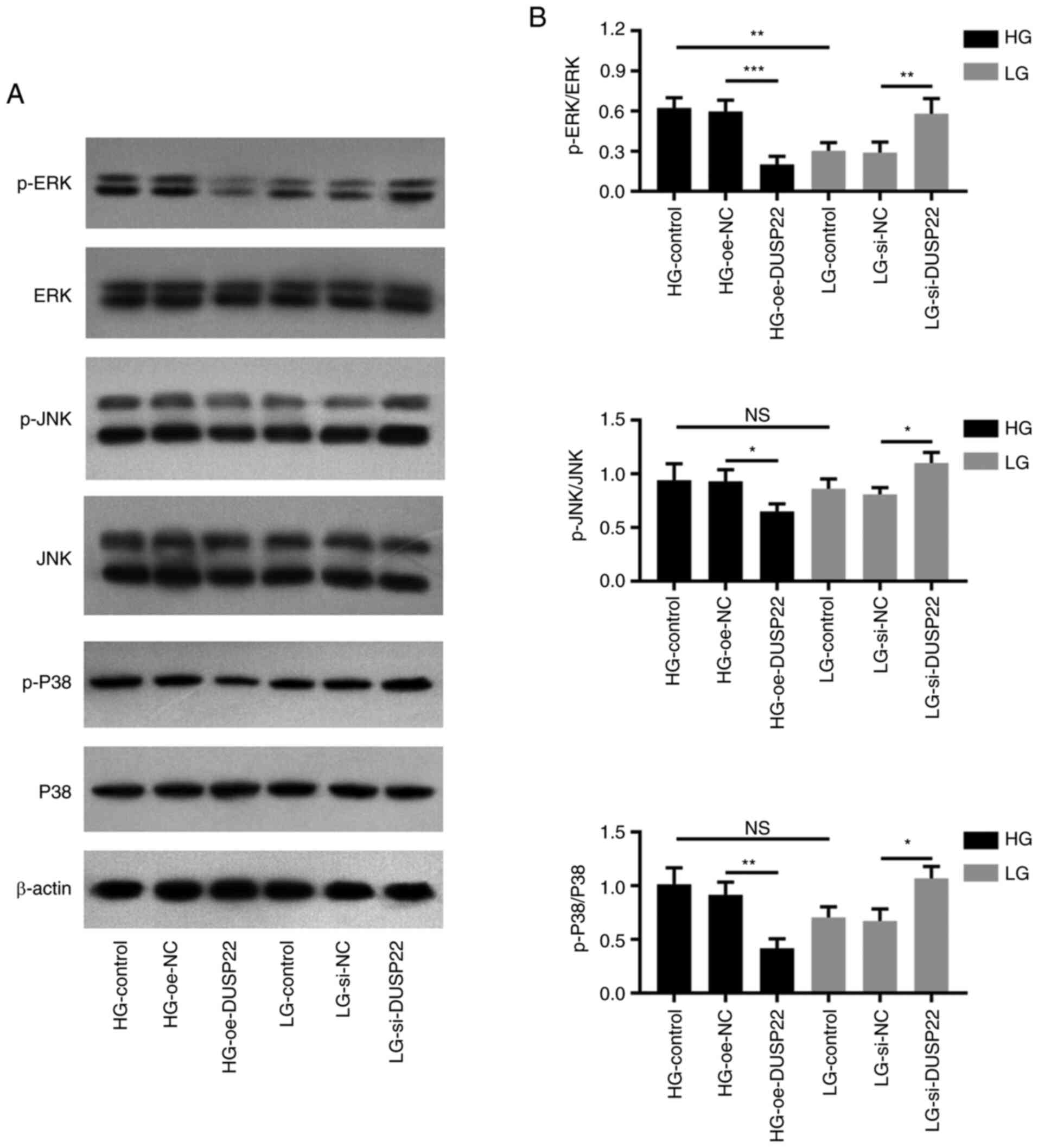 | Figure 6DUSP22 suppresses the MAPK signaling
pathway in diabetic nephropathy. (A) Western blot analysis
demonstrating the effect of DUSP22 on the MAPK signaling pathway in
SV40-MES13 cells. Densitometric analysis of western blotting at
panel A revealing the effect of DUSP22 on p-ERK, p-JNK and p-P38 in
SV40-MES13 cells. (B) Impact of DUSP22 on p-ERK, p-JNK, and p-P38
in SV40-MES13 cells. *P<0.05, **P<0.01
and ***P<0.001. DUSP22, dual specificity phosphatase
22; p-, phosphorylated; HG, high glucose; LG, low glucose; oe,
overexpression; si-, small interfering; NC, negative control; ns,
no significance. |
Discussion
DUSPs are widely known as dephosphorylated proteins
which serve as protective factors in inflammation-related injuries
via inactivating the MAPK signaling pathway (21-23).
For example, a recent study revealed that DUSP26 knockdown promotes
fibrosis in kidney glomeruli via enhancing TGF-β1 expression, then
renal injury and dysfunction of DN are greatly accelerated
(22). As a member of the DUSP
family, DUSP22 shares a similar function of regulating MAPK
signaling, which is a crucial pathway in the development of renal
diseases (24,25). For instance, a recent study
identified the dysregulation of DUSP22 in IgA nephropathy (26). In addition, it was demonstrated
that the aberrant DUSP22 expression is linked with increased
disease activity and poor renal outcomes in patients with systemic
lupus erythematosus nephritis (27). The involvement of DUSP22 in DN
pathogenesis remains unclear; thus, the present study was conducted
and it was found that DUSP22 overexpression promoted mesangial cell
apoptosis, but it suppressed mesangial cell proliferation and
fibrosis under the HG condition; while DUSP22 siRNA caused the
opposite effect in LG-treated mesangial cells. These findings
suggested that DUSP22 weakened DN hyperproliferation and fibrosis,
which may be due to the following possible reasons: i) DUSP22
overexpression restrained MAPKs (including ERK, JNK, and p38),
while both JNK and p38 may enhance renal cell proliferation but
inhibit renal cell apoptosis via reducing anti-apoptotic proteins
(such as Bcl2) (28,29). Therefore, DUSP22 in SV40-MES13
cells inhibited cell proliferation but promoted cell apoptosis; ii)
DUSP22 suppressed epithelial-to-mesenchymal transition (EMT) via
restraining the p38/MAPK pathway; meanwhile, EMT could accelerate
HG-induced renal fibrosis (23,30,31).
Thus, DUSP22 reduced renal fibrosis in SV40-MES13 cells.
Furthermore, the results of the TUNEL assay and cleaved-caspase-3
shared a similar trend. The only discrepancy was that the apoptotic
rate was decreased, while cleaved caspase 3 only demonstrated a
decreasing trend (lacked statistical significance) in the
HG-Control group compared with the LG-Control group. A possible
explanation may be that cleaved caspase 3 protein expression and
TUNEL-reflected cell apoptosis themselves presented a mild
difference (32,33).
In line with other chronic renal diseases,
inflammation dominates in the development of DN; in turn, the HG
environment can induce inflammatory injury as well (34). Accordingly, it was observed in the
current study that TNF-α, IL-1β and IL-12 were elevated in
mesangial cells under the HG condition compared with those under
the LG condition. With regard to the effect of DUSP22 on
inflammation, it has been recognized that DUSP22 attenuates
inflammatory cytokine recruitment via inhibiting several signaling
pathways [such as the T cell receptor (TCR)] signaling pathway and
the nuclear factor-kappa B pathway] (11,27,35).
For instance, a previous study revealed that IL-17, IL-6 and
interferon-γ are increased in DUSP22-knockout CD4+ T
cells through the modulation of TCR signaling (11). Similarly, it was identified in the
present study that DUSP22 overexpression downregulated TNF-α, IL-6,
IL-1β and IL-12 in HG-treated SV40-MES13 cells; while DUSP22 siRNA
showed the opposite effect on them under the LG condition, which
indicated that DUSP22 inhibited inflammation in DN. A probable
explanation may be the following: DUSP22 could inactive the MAPK
pathway while the latter triggered several inflammatory pathways;
subsequently, the excessive secretion of pro-inflammatory cytokines
(including TNF-α, IL-6, IL-1β and IL-12) was suppressed (35).
Additionally, SV40-MES13 cells in the HG group were
stimulated using 25 mM D-glucose for 48 h to establish the cellular
DN model and it was found that HG protected from apoptosis.
Meanwhile, certain previous studies also identified that HG
promotes mesangial cell proliferation and fibrosis at the same
concentration (18,36). Whereas one study revealed that HG
may increase apoptosis in human renal proximal tubular epithelial
cells under 30 mmol/l D-glucose (37), which may be possibly due to that
different glucose concentrations and different cells may cause
different trends. TGF-β1 was strongly activated in the murine renal
mesangial cell line, which induced cell proliferation and fibrosis
in high-glucose media (38). On
the contrary, the HG condition activated p38 mitogen-activated
protein kinase (p38 MAPK) in human renal proximal tubular
epithelial cells, which further promoted cell apoptosis (37,39).
Hence, different phenomena are observed in different cell
types.
The activation of the MAPK signaling pathway
(contains ERK, JNK and p38) promotes inflammation response and cell
death in renal tubular and membrane (40,41).
As to the detailed role of the MAPK signaling pathway in DN
pathogenesis, a previous study revealed that the MAPK pathway
modulates cell apoptosis, over-production of inflammatory cytokines
and extracellular matrix dysregulation of DN (25). In the present study, it was found
that DUSP22 overexpression restrained p-JNK, p-ERK and p-p38 under
HG-treated SV40-MES13 cells, whereas DUSP22 siRNA exhibited the
opposite effect under the LG condition, which suggested that DUSP22
blocked the MAPK signaling pathway in SV40-MES13 cells. The
limitations to the present study were non-negligible: Firstly,
HG-group cells were only transfected with DUSP22 overexpression
plasmid, and LG-group cells were only transfected with DUSP22
siRNA. Therefore, further study transfecting HG-group and LG-group
cells with both DUSP22 overexpression and DUSP22 siRNA was
necessary. Secondly, the flow cytometry experiments were warranted
in further studies to validate the apoptotic results. Thirdly,
since no human cells were used in the present study, there exists a
potential limitation in translating results into clinical
applications. Fourthly, renal mesangial and endothelial cells did
not downregulate glucose transporters under HG conditions;
subsequently, the Krebs cycle resulted in more nicotinamide adenine
dinucleotide (42). In this case,
the same cell number was expected to provide a more intense signal
in the CCK-8 assay when cultured under HG conditions. Thus, further
study should conduct the BrdU assay to verify the cell
proliferation results. In conclusion, it was revealed that DUSP22
suppresses HG-induced mesangial cell hyperproliferation, fibrosis,
inflammation and the MAPK pathway, indicating its potency in DN
treatment.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NR and SS contributed equally to the conception and
design and drafted the manuscript. LZ and NZ contributed to
analyzing the data and revised the manuscript critically for
important intellectual content. NR and SS confirm the authenticity
of all the raw data. All authors read and approved the final
manuscript, and agreed to be accountable for all aspects of the
work in ensuring that questions related to the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Samsu N: Diabetic nephropathy: Challenges
in pathogenesis, diagnosis, and treatment. Biomed Res Int.
2021(1497449)2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Sagoo MK and Gnudi L: Diabetic
nephropathy: An overview. Methods Mol Biol. 2067:3–7.
2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhang XX, Kong J and Yun K: Prevalence of
diabetic nephropathy among patients with type 2 diabetes Mellitus
in China: A meta-analysis of observational studies. J Diabetes Res.
2020(2315607)2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Selby NM and Taal MW: An updated overview
of diabetic nephropathy: Diagnosis, prognosis, treatment goals and
latest guidelines. Diabetes Obes Metab. 22 (Suppl 1):S3–S15.
2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Yamazaki T, Mimura I, Tanaka T and Nangaku
M: Treatment of diabetic kidney disease: Current and future.
Diabetes Metab J. 45:11–26. 2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Agarwal R: Pathogenesis of diabetic
nephropathy. Chronic kidney disease and type 2 diabetes. Arlington
(VA) American Diabetes Association 2021.
|
|
7
|
Cao Z and Cooper ME: Pathogenesis of
diabetic nephropathy. J Diabetes Investig. 2:243–247.
2011.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yu SM and Bonventre JV: Acute kidney
injury and progression of diabetic kidney disease. Adv Chronic
Kidney Dis. 25:166–180. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chen AJ, Zhou G, Juan T, Colicos SM,
Cannon JP, Cabriera-Hansen M, Meyer CF, Jurecic R, Copeland NG,
Gilbert DJ, et al: The dual specificity JKAP specifically activates
the c-Jun N-terminal kinase pathway. J Biol Chem. 277:36592–36601.
2002.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Sekine Y, Ikeda O, Hayakawa Y, Tsuji S,
Imoto S, Aoki N, Sugiyama K and Matsuda T: DUSP22/LMW-DSP2
regulates estrogen receptor-alpha-mediated signaling through
dephosphorylation of Ser-118. Oncogene. 26:6038–6049.
2007.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Li JP, Yang CY, Chuang HC, Lan JL, Chen
DY, Chen YM, Wang X, Chen AJ, Belmont JW and Tan TH: The
phosphatase JKAP/DUSP22 inhibits T-cell receptor signalling and
autoimmunity by inactivating Lck. Nat Commun.
5(3618)2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Li J, Jin S, Barati MT, Rane S, Lin Q, Tan
Y, Cai L and Rane MJ: ERK and p38 MAPK inhibition controls NF-E2
degradation and profibrotic signaling in renal proximal tubule
cells. Life Sci. 287(120092)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Grynberg K, Ma FY and Nikolic-Paterson DJ:
The JNK signaling pathway in renal fibrosis. Front Physiol.
8(829)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Xiao L, Chen A, Gao Q, Xu B, Guo X and
Guan T: Pentosan polysulfate ameliorates fibrosis and inflammation
markers in SV40 MES13 cells by suppressing activation of PI3K/AKT
pathway via miR-446a-3p. BMC Nephrol. 23(105)2022.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wu R, Niu Z, Ren G, Ruan L and Sun L:
CircSMAD4 alleviates high glucose-induced inflammation,
extracellular matrix deposition and apoptosis in mouse glomerulus
mesangial cells by relieving miR-377-3p-mediated BMP7 inhibition.
Diabetol Metab Syndr. 13(137)2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chen Z, Gao H, Wang L, Ma X, Tian L, Zhao
W, Li K, Zhang Y, Ma F, Lu J, et al: Farrerol alleviates high
glucose-induced renal mesangial cell injury through the
ROS/Nox4/ERK1/2 pathway. Chem Biol Interact.
316(108921)2020.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Zhao L, Chen H, Zeng Y, Yang K, Zhang R,
Li Z, Yang T and Ruan H: Circular RNA circ_0000712 regulates high
glucose-induced apoptosis, inflammation, oxidative stress, and
fibrosis in (DN) by targeting the miR-879-5p/SOX6 axis. Endocr J.
68:1155–1164. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Zhang P, Sun Y, Peng R, Chen W, Fu X,
Zhang L, Peng H and Zhang Z: Long non-coding RNA Rpph1 promotes
inflammation and proliferation of mesangial cells in diabetic
nephropathy via an interaction with Gal-3. Cell Death Dis.
10(526)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Aggarwal M, Saxena R, Asif N, Sinclair E,
Tan J, Cruz I, Berry D, Kallakury B, Pham Q, Wang TTY and Chung FL:
p53 mutant-type in human prostate cancer cells determines the
sensitivity to phenethyl isothiocyanate induced growth inhibition.
J Exp Clin Cancer Res. 38(307)2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Huang CY and Tan TH: DUSPs, to MAP kinases
and beyond. Cell Biosci. 2(24)2012.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Huang F, Sheng XX and Zhang HJ: DUSP26
regulates podocyte oxidative stress and fibrosis in a mouse model
with diabetic nephropathy through the mediation of ROS. Biochem
Biophys Res Commun. 515:410–416. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Guo H, Jian Z, Liu H, Cui H, Deng H, Fang
J, Zuo Z, Wang X, Zhao L, Geng Y, et al: TGF-β1-induced EMT
activation via both Smad-dependent and MAPK signaling pathways in
Cu-induced pulmonary fibrosis. Toxicol Appl Pharmacol.
418(115500)2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Chuang HC and Tan TH: MAP4K family kinases
and DUSP family phosphatases in T-Cell signaling and systemic lupus
erythematosus. Cells. 8(1433)2019.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Fang Y, Tian X, Bai S, Fan J, Hou W, Tong
H and Li D: Autologous transplantation of adipose-derived
mesenchymal stem cells ameliorates streptozotocin-induced diabetic
nephropathy in rats by inhibiting oxidative stress,
pro-inflammatory cytokines and the p38 MAPK signaling pathway. Int
J Mol Med. 30:85–92. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Li M, Wang L, Shi DC, Foo JN, Zhong Z,
Khor CC, Lanzani C, Citterio L, Salvi E, Yin PR, et al: Genome-Wide
meta-analysis identifies three novel susceptibility loci and
reveals ethnic heterogeneity of genetic susceptibility for IgA
nephropathy. J Am Soc Nephrol. 31:2949–2963. 2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Chuang HC, Chen YM, Hung WT, Li JP, Chen
DY, Lan JL and Tan TH: Downregulation of the phosphatase
JKAP/DUSP22 in T cells as a potential new biomarker of systemic
lupus erythematosus nephritis. Oncotarget. 7:57593–57605.
2016.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Yue J and Lopez JM: Understanding MAPK
signaling pathways in apoptosis. Int J Mol Sci.
21(2346)2020.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Shen Y, Teng L, Qu Y, Liu J, Zhu X, Chen
S, Yang L, Huang Y, Song Q and Fu Q: Anti-proliferation and
anti-inflammation effects of corilagin in rheumatoid arthritis by
downregulating NF-κB and MAPK signaling pathways. J Ethnopharmacol.
284(114791)2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Loeffler I and Wolf G:
Epithelial-to-mesenchymal transition in diabetic nephropathy: Fact
or fiction? Cells. 4:631–652. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Xu M, Wang S, Wang Y, Wu H, Frank JA,
Zhang Z and Luo J: Role of p38ү MAPK in regulation of EMT and
cancer stem cells. Biochim Biophys Acta Mol Basis Dis.
1864:3605–3617. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wu C, Zhou XX, Li JZ, Qiang HF, Wang Y and
Li G: Pretreatment of cardiac progenitor cells with bradykinin
attenuates H2O2-induced cell apoptosis and
improves cardiac function in rats by regulating autophagy. Stem
Cell Res Ther. 12(437)2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Du X, Wang X, Cui K and Chen Y, Zhang C,
Yao K, Hao Y and Chen Y: Tanshinone IIA and Astragaloside IV
Inhibit miR-223/JAK2/STAT1 signalling pathway to alleviate
lipopolysaccharide-induced damage in nucleus pulposus cells. Dis
Markers. 2021(6554480)2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wada J and Makino H: Inflammation and the
pathogenesis of diabetic nephropathy. Clin Sci (Lond). 124:139–152.
2013.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Lim AK, Nikolic-Paterson DJ, Ma FY, Ozols
E, Thomas MC, Flavell RA, Davis RJ and Tesch GH: Role of MKK3-p38
MAPK signalling in the development of type 2 diabetes and renal
injury in obese db/db mice. Diabetologia. 52:347–358.
2009.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Li A, Peng R, Sun Y, Liu H, Peng H and
Zhang Z: LincRNA 1700020I14Rik alleviates cell proliferation and
fibrosis in diabetic nephropathy via miR-34a-5p/Sirt1/HIF-1α
signaling. Cell Death Dis. 9(461)2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Chen P, Yuan Y, Zhang T, Xu B, Gao Q and
Guan T: Pentosan polysulfate ameliorates apoptosis and inflammation
by suppressing activation of the p38 MAPK pathway in high
glucosetreated HK2 cells. Int J Mol Med. 41:908–914.
2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Yoon JJ, Lee HK, Kim HY, Han BH, Lee HS,
Lee YJ and Kang DG: Sauchinone protects renal mesangial cell
dysfunction against angiotensin II by improving renal fibrosis and
inflammation. Int J Mol Sci. 21(7003)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Lv ZM, Wang Q, Wan Q, Lin JG, Hu MS, Liu
YX and Wang R: The role of the p38 MAPK signaling pathway in high
glucose-induced epithelial-mesenchymal transition of cultured human
renal tubular epithelial cells. PLoS One. 6(e22806)2011.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Dong Q, Jie Y, Ma J, Li C, Xin T and Yang
D: Renal tubular cell death and inflammation response are regulated
by the MAPK-ERK-CREB signaling pathway under hypoxia-reoxygenation
injury. J Recept Signal Transduct Res. 39:383–391. 2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Ju A, Cho YC, Kim BR, Park SG, Kim JH, Kim
K, Lee J, Park BC and Cho S: Scaffold Role of DUSP22 in
ASK1-MKK7-JNK signaling pathway. PLoS One.
11(e0164259)2016.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Kosanam H, Thai K, Zhang Y, Advani A,
Connelly KA, Diamandis EP and Gilbert RE: Diabetes induces lysine
acetylation of intermediary metabolism enzymes in the kidney.
Diabetes. 63:2432–2439. 2014.PubMed/NCBI View Article : Google Scholar
|