Introduction
Fibroblasts are tissue-resident stromal cells that
are important for maintaining the structural integrity of tissues
(1). They function to synthesize
and integrate structural proteins, such as collagen and elastin,
into the extracellular matrix (ECM) of connective tissues (1). In environments where this homeostasis
is disturbed, such as during wound healing, chronic inflammation
and cancer, fibroblasts are activated to proliferate and upregulate
ECM production (2,3). Activated fibroblasts acquire various
smooth muscle features, including enhanced formation of contractile
stress fibers and expression of α-smooth muscle actin (α-SMA)
(2,3). Cells with these characteristics are
known as myofibroblasts (2,3).
Although transient acquisition of this myofibroblast phenotype
confers beneficial effects on normal tissue repair processes,
persistence of myofibroblasts is associated with the development of
diseases mediated by tissue stiffening and deformation (4). Stiff scar tissues adversely alters
normal organ function (4). In
addition, fibrosis is characterized by the abnormally excessive
accumulation of ECM proteins, which contributes to organ failure in
various chronic diseases affecting the liver, kidney, skin, lungs
and the heart (5). By contrast,
activated fibroblasts, especially cancer-associated fibroblasts
(CAFs) in the tumor stroma, serve an important role in
tumorigenesis by stimulating angiogenesis, cancer cell
proliferation and invasion (6).
Activated CAFs can also produce a variety of growth factors and
proinflammatory cytokines, such as TGF-β, vascular endothelial
growth factor, IL-6 and CXC-chemokine ligand 12, to promote tumor
progression (6-8).
CAFs have been reported to contribute to ECM remodeling and cancer
cell invasion by secreting connective tissue growth factor (CTGF),
collagen, fibronectin and elastin (9), implicating CAFs to be targets for
anti-cancer therapy (6-8).
CTGF, also known as cellular communication network
factor 2, is a regulatory protein that has been demonstrated to be
involved in several biological processes, such as cell
proliferation, angiogenesis and wound healing (10). In addition, CTGF has been
associated with a number of pathological processes, such as tumor
development, cardiovascular diseases, inflammatory diseases and
tissue fibrosis in major organs (10). CTGF was first discovered as a
protein secreted by endothelial cells during angiogenesis under
normal conditions (11). CTGF
expression is generally higher in blood vessels compared with that
in other organs or tissues (11).
CTGF mRNA is expressed at particularly high levels in developing
blood vessels and in the large blood vessels of the adult heart,
suggesting that CTGF is involved in the development and maintenance
of blood vessels (12). However,
one of the main roles of CTGF is considered to be the promotion of
myofibroblast differentiation and angiogenesis (13-15).
CTGF is typically secreted into the extracellular environment,
where it interacts with cell surface receptors, growth factors and
the ECM (13-15).
Subsequently, CTGF mediates downstream effects by binding to
heterodimeric cell-surface integrin complexes, such as
α6, β1, αV and β3
integrins (13-15).
The present study aimed to investigate whether CTGF
from endothelial cells after hypoxia/reoxygenation (H/R) could
stimulate the differentiation of neighboring fibroblasts into
myofibroblasts in a paracrine manner.
Materials and methods
Cell culture conditions
HUVECs (passages 4-10; Lonza Group, Ltd.) were
cultured in EGM™-2 Endothelial Cell Growth Medium-2 BulletKit™
(Lonza Group, Ltd.) containing all the included supplements at 37˚C
in a humidified atmosphere containing 5% CO2. Normal human dermal
fibroblasts (PromoCell GmbH) were cultured in DMEM (Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (Thermo Fisher
Scientific, Inc.) at 37˚C in a humidified atmosphere containing 5%
CO2. For the H/R conditions, the cells were first incubated at 37˚C
for 16 h in a hypoxic incubator (Thermo Scientific 3131 Forma
Incubator; Thermo Fisher Scientific, Inc.) filled with 1% O2 and 5%
CO2, balanced with N2, before being placed under normoxic
conditions at 37˚C for 2 h for reoxygenation treatment. For the
preparation of conditioned media (CM), the medium of HUVECs was
changed with EBM™-2 Basal Medium (Lonza Group, Ltd.) containing 1%
FBS at 37˚C for 18 h before it was collected, and filtered using a
Minisart® Syringe Filter (0.25 µm; Sartorius AG).
RNA extraction and semi-quantitative
reverse transcription-PCR (sqRT-PCR)
Total RNA was isolated from HUVECs using TRIzol™
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). To remove
genomic DNA, total RNA (4 µg) was treated with RQ1 RNase-Free DNase
(cat. no. M6101; Promega Corporation) prior to sqRT-qPCR, according
to the manufacturer's protocols. A PrimeScript™ First Strand cDNA
Synthesis Kit (Takara Bio, Inc.) was used to synthesize cDNA from 2
µg total RNA in a total volume of 20 µl, according to the
manufacturer's protocols. PCR was performed on 1 µl cDNA in a total
volume of 25 µl using TaKaRa Ex Taq® DNA Polymerase
(cat. no. RR01AM; Takara Bio, Inc.) according to the manufacturer's
instructions. The following primer pair was used for human CTGF:
Forward, 5'-GCATCCGTACTCCCAAAATCTC-3' and reverse,
5'-ATGTCTCTCACTCTCT GGCTTC-3' (melting temperature: 55˚C; 27 cycles
at 94˚C for 30 sec, 55˚C for 30 sec, 72˚C for 30 sec). GAPDH was
used for normalization: Forward, 5'-CGTGGAAGGACTCATGAC-3' and
reverse, 5'-CAAATTCGTTGTCATACCAG-3' (melting temperature: 55˚C; 27
cycles at 94˚C for 30 sec, 55˚C for 30 sec, 72˚C for 30 sec). The
PCR products were mixed with Ezstain DNA loading dye (cat. no.
B006M; Enzynomics, Inc.) and analyzed using a 1.2% agarose gel.
Semiquantification of band intensity was performed using ImageJ
software (V1.8.0; National Institutes of Health) and normalized to
the intensity of GAPDH.
Vector construction
The coding region of human CTGF (NM_001901) was
PCR-amplified from HUVEC cDNA (1 µl) using TaKaRa Ex
Taq® DNA Polymerase (cat. no. RR01AM; Takara Bio, Inc.)
according to the manufacturer's instructions, cloned into the
pGEM®-T Easy Vector Systems (cat. no. A1360; Promega
Corporation), before being subcloned into the pShuttle-CMV vector
in the AdEasy adenoviral vector system (cat. no. 240009; Agilent
Technologies, Inc.) to produce CTGF-expressing adenovirus. The
primers are as follows: cdsCTGF forward,
5'-GAGTCGACAGTGCCAACCATGACCGC-3' (nucleotides plus SalI
adapter) and cdsCTGF reverse, 5'-GACTCGAGCTGGCTTCATGCCATGTC-3'
(nucleotides plus XhoI adapter). PCR was performed for 27
cycles at 94˚C for 1 min, 60˚C for 1 min and 72˚C for 1 min. All
PCR-amplified fragments and cloning junctions were verified by DNA
sequencing performed by SolGent Co., Ltd. Adenoviral CTGF cloning
was performed according to the manufacturer's protocols. Production
and harvesting of adenoviruses were performed as described
(16,17). The pShuttle-CMV vector containing
CTGF (1 µg) was cotransformed with pAdEasy-1 vector (100 ng) into
BJ5183 competent cells (20 µl; supplied in the in the AdEasy
adenoviral vector system), where homologous recombination occurred.
The recombinant adenoviral vector expressing human CTGF was
transfected into 293A cells to obtain viral particles. 293A cells
(cat. no. R70507; Thermo Fisher Scientific, Inc.) were cultured in
DMEM (Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin
(Thermo Fisher Scientific, Inc.) at 37˚C. 293A cells
(5x106 cells/100 mm dish) were plated 24 h before
transfection. Subsequently, 10 µg recombinant adenoviral vector DNA
was used for transfection with MetaFectene PRO (cat. no. T040-1.0;
Biontex Laboratories GmbH) according to the manufacturer's
instruction. Transfected cells were incubated at 37˚C for 7-10
days, the adenovirus-producing 293A cells were collected and the
virus particles were purified. The infection into HUVECs was
performed as previously described (18). The harvested adenoviruses (25 MOI)
were added to cells in endothelial basal medium (Lonza Group, Ltd.)
containing 1% FBS at 37˚C for 4 h, then virus-containing medium was
removed and growth medium was added. After 24 h, medium was removed
and the cells were washed one time with endothelial basal medium,
and then incubated in endothelial basal media containing 1% FBS at
37˚C for 18 h before collecting CM. Control HUVECs were infected
with a control adenovirus generated with control shuttle vector
(pShuttle-CMV-lacZ) and pAdEasy-1 vector.
Reagents
Recombinant human CTGF (rhCTGF; cat. no. 30R-2206;
0.5 or 1 µg/ml) was purchased from Fitzgerald Industries
International, Inc. (19,20), and used to treat fibroblasts at
37˚C for 3 days. Function-blocking Anti-Integrin αVβ3 Antibody
(clone LM609; cat. no. MAB1976; 10 µg/ml) and Anti-Integrin α6
Antibody (clone NKI-GoH3; cat. no. MAB1378; 10 µg/ml) were
purchased from Sigma-Aldrich; Merck KGaA (21,22).
Normal mouse IgG (cat. no. NI03; 10 µg/ml) was purchased from
Sigma-Aldrich; Merck KGaA. Functional-blocking antibodies were used
to treat fibroblasts at 37˚C for 3 days. To analyze the half-life
or synthesis rate of the CTGF protein in HUVECs, the proteasome
inhibitor MG132 (cat. no. M8699; 10 µM) or the translational
blocker cycloheximide (CHX; cat. no. C0934; 10 µg/ml) was used
(both Sigma-Aldrich; Merck KGaA) to treat cells at 37˚C for the
indicated time (2 h for MG132; 30, 60 or 120 min for CHX).
Western blotting
HUVECs were harvested and lysed using RIPA buffer
(cat. no. 9806; Cell Signaling Technology, Inc.) containing Xpert
Protease Inhibitor Cocktail Solution (cat. no. P3100; GenDEPOT) and
Xpert Phosphatase Inhibitor Cocktail Solution (cat. no. P3200;
GenDEPOT). Total protein content was determined by Pierce™ BCA
Protein Assay Kit (cat. no. 23225; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Total protein (20-30
µg) was separated by SDS-PAGE on 12% gels and transferred onto a
nitrocellulose blotting membrane (pore size, 0.45 µm; cat. no.
10600003; Amersham; Cytiva). After transfer, the membrane was
incubated with 5% skimmed milk (cat. no. 232100; BD Biosciences) in
PBS containing 0.1% Tween-20 for 1 h at room temperature. The
membranes were then immunoblotted with a specific antibody against
CTGF (1:500; cat. no. sc-14939; Santa Cruz Biotechnology, Inc.) for
16 h at 4˚C. β-actin (1:2,000; cat. no. sc-47778; Santa Cruz
Biotechnology, Inc.), which was used as an internal control, was
incubated with the membranes for 16 h at 4˚C. For the incubation
with secondary antibodies, HRP-conjugated goat anti-mouse antibody
(1:5,000; cat. no. 1031-05; SouthernBiotech) or HRP-conjugated
rabbit anti-goat antibody (1:5,000; cat. no. 6160-05;
SouthernBiotech) was applied for 1 h at room temperature.
Chemiluminescence signals were obtained with SuperSignal™ West Pico
PLUS Chemiluminescent Substrate (cat. no. 34580; Thermo Fisher
Scientific, Inc.) and chemiluminescence intensity was measured
using the ImageQuant™ LAS 4000 apparatus (Cytiva). The
quantification of band intensity was performed using ImageJ
software.
Immunofluorescence staining
Fibroblasts were grown on cover glasses
(2x104 cells/well in a 12-well plate) in a monolayer and
treated with either CM or rhCTGF (0.5 or 1 µg/ml) at 37˚C for 3
days. The cells were then fixed with 4% paraformaldehyde for 10 min
at 4˚C, before being incubated with blocking buffer comprising of
2% bovine serum albumin (cat. no. 0332; Amresco, LLC) in 1X PBS for
1 h at room temperature. The cells were labeled with anti-α-SMA
antibody (1:800; cat. no. ab7817; Abcam) overnight at 4˚C, followed
by Alexa Fluor™ 488-conjugated Goat anti-Mouse IgG (H + L)
Cross-Adsorbed Secondary Antibody (1:200; cat. no. A-11001; Thermo
Fisher Scientific, Inc.) for 1 h at room temperature. The nuclei
were stained with DAPI (1:5,000; cat. no. D1306; Molecular Probes;
Thermo Fisher Scientific, Inc.) for 1 h at room temperature and
mounted using a fluorescent mounting medium (cat. no. S3023; DAKO;
Agilent Technologies, Inc.). Fluorescent images were obtained using
a Zeiss LSM 700 laser scanning confocal microscope (Carl Zeiss AG;
magnifications, x100, x200 and x400). The quantification of
SMA+ area (n=5-6 fields/group) was performed using
ImageJ software.
Statistical analysis
All data are expressed as the mean ± standard error
of the mean from three or four independent experiments. All of the
significance analysis was performed using the SigmaPlot version
14.0 software (SPSS, Inc.). The statistical differences were
compared using one-way ANOVA followed by the Tukey's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
CTGF treatment causes the
differentiation of fibroblasts into myofibroblasts
It was reported in our previous study that
endothelial cells undergo endothelial-to-mesenchymal transition
(EndMT) when subjected to ischemia/reperfusion, influencing
neighboring fibroblasts to actively participate in cardiac fibrosis
(23). Although it was found in
this previous study that CTGF from EndMT cells has a paracrine
influence on fibroblast activation (23), the direct effects of CTGF on the
activation of fibroblasts into myofibroblasts or its mechanism were
not examined. Therefore, in the present study, the effects of
hypoxia/reoxygenation (H/R) on CTGF expression in HUVECs was first
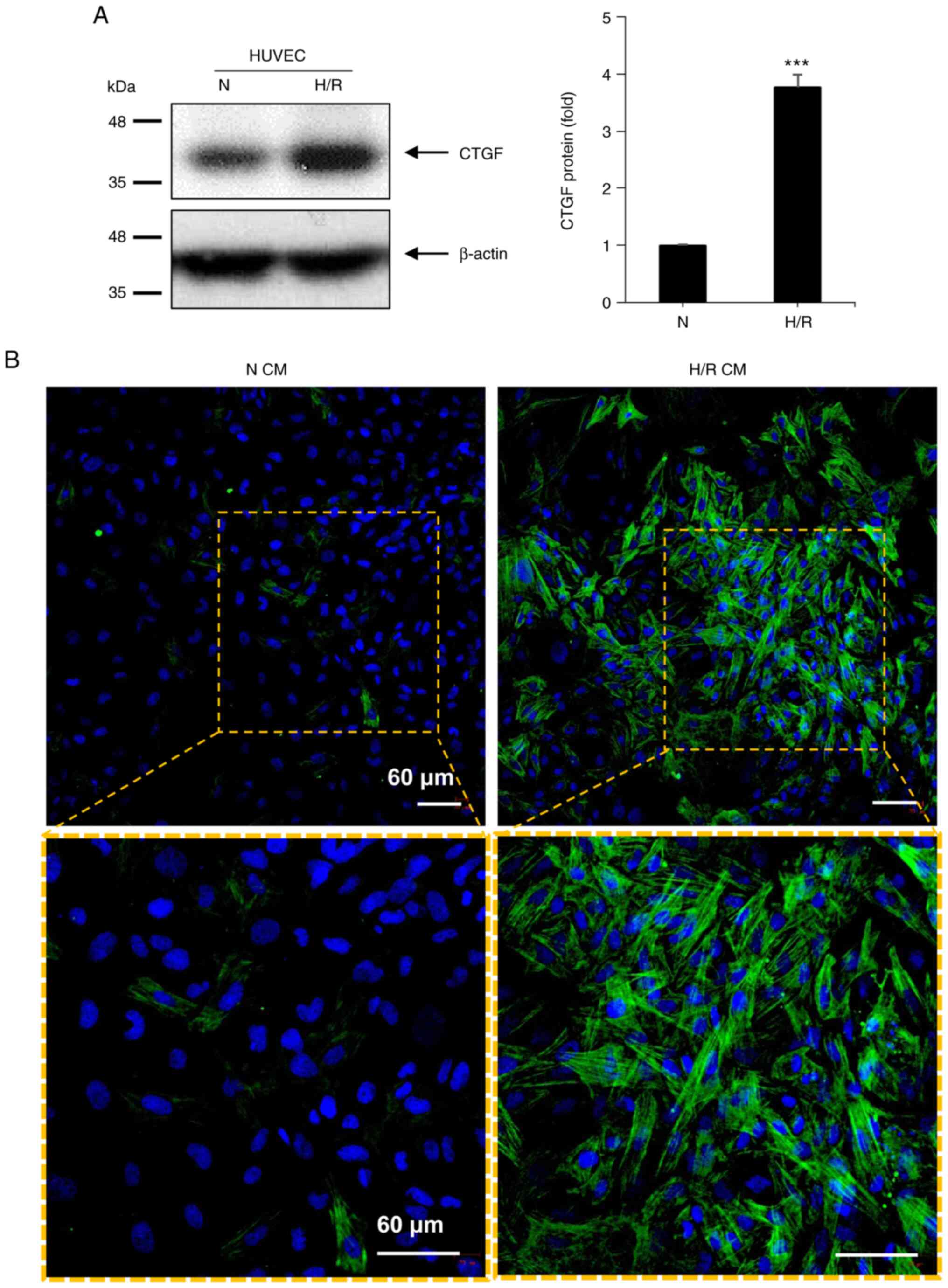
assessed (Fig. 1A). It was found
that H/R significantly increased the expression of the CTGF protein
(Fig. 1A). Culture supernatants
from both normoxic and H/R HUVECs were then obtained and were used
to treat fibroblasts (Fig. 1B).
The immunofluorescence of α-SMA stress fibers was examined, which
indicates the generation of myofibroblasts (2). Fibroblasts treated with CM from
normoxic HUVECs (N CM) showed punctate or patchy α-SMA
immunofluorescence. By contrast, the treatment of fibroblasts with
CM from H/R HUVEC (H/R CM) led to the formation of more intense and
fibrous α-SMA immunofluorescence, typical of stress fibers
(Fig. 1B). The effect of soluble
CTGF on fibroblast differentiation was then examined using rhCTGF
(Fig. 2). Fibroblasts were treated
with 0.5 or 1 µg/ml rhCTGF, before α-SMA immunofluorescence was
observed. Treatment with rhCTGF dose-dependently stimulated the
formation of α-SMA stress fibers in fibroblasts, indicating that
rhCTGF directly induced the differentiation of fibroblasts into
myofibroblasts.
Function-blocking antibody against
integrin αVβ3 abolishes the effect of CM from
CTGF-overexpressing HUVECs on α-SMA fiber formation
To investigate how CTGF from endothelial cells
affects neighboring fibroblasts, function-blocking antibodies
against integrin α6 and αVβ3 were used alongnside CM from
CTGF-overexpressing HUVECs (Fig.
3). CTGF mediates downstream effects by binding to integrins,
such as α6, β1, αV and β3(15).
HUVECs were first infected with a control adenovirus (Ad-mock) or
an adenovirus expressing human CTGF (Ad-CTGF), before CTGF
overexpression was confirmed using western blot analysis (Fig. 3A). HUVEC CM was then collected and
used to treat fibroblasts and immunofluorescence staining for α-SMA
was performed (Fig. 3B). CM from
CTGF-overexpressing HUVECs potently stimulated α-SMA stress fiber
formation, which was significantly inhibited by a function-blocking
antibody against integrin αVβ3 (Fig.
3B). The function-blocking antibody against integrin α6 could
not block stress fiber formation (Fig.
3B and C), suggesting that
CTGF from endothelial cells stimulates the differentiation of
fibroblasts to myofibroblasts through integrin αVβ3.
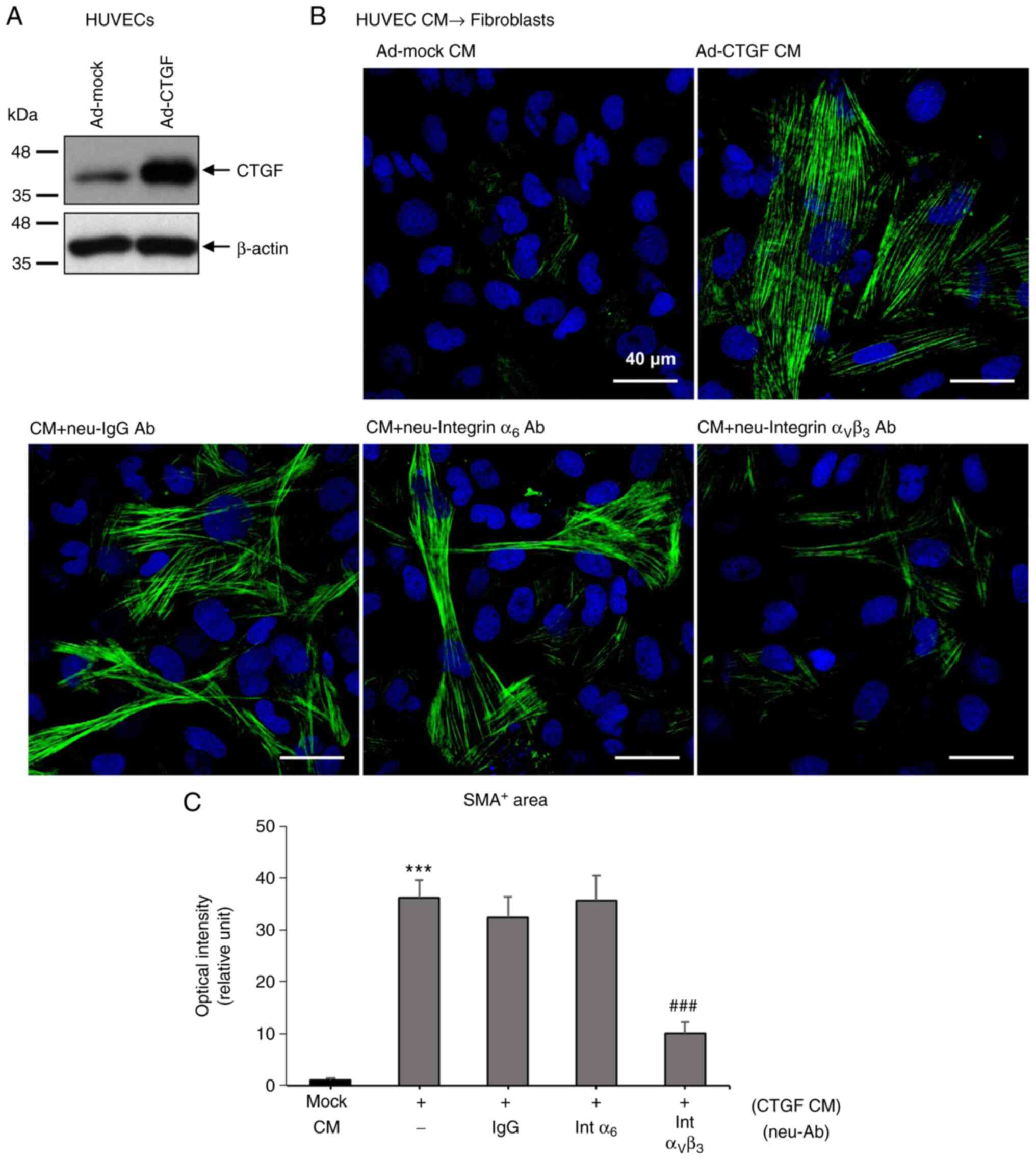 | Figure 3CM from CTGF-overexpressing HUVECs
stimulates α-SMA stress fiber formation through
αVβ3-integrin in fibroblasts. (A) CTGF
protein expression was increased in HUVECs infected with Ad-CTGF.
CTGF overexpression was confirmed using western blot analysis.
Ad-mock HUVECs were infected with a control Ad containing a control
shuttle vector (pShuttle-CMV-lacZ). (B) Function-blocking
Int αVβ3 diminished the effects of CTGF CM on
α-SMA fiber formation (green) in fibroblasts. Intα6, Int
αVβ3 or purified mouse IgG (each 10 µg/ml)
was added to the CM directly. Nuclei were stained with DAPI (blue).
Magnification, x400. (C) Quantification of the α-SMA+
area (n=5-6 per group). ***P<0.001 vs. mock CM.
###P<0.001 vs. CTGF CM only. CM, conditioned medium;
Ad, adenovirus; CTGF, connective tissue growth factor; IgG,
immunoglobulin G; neu, neutralizing; Ab, antibody;
Intα6, anti-integrin α6 antibody; Int
αVβ3, anti-integrin
αVβ3 antibody; SMA, smooth muscle actin. |
CTGF protein stability is increased
under H/R in HUVECs
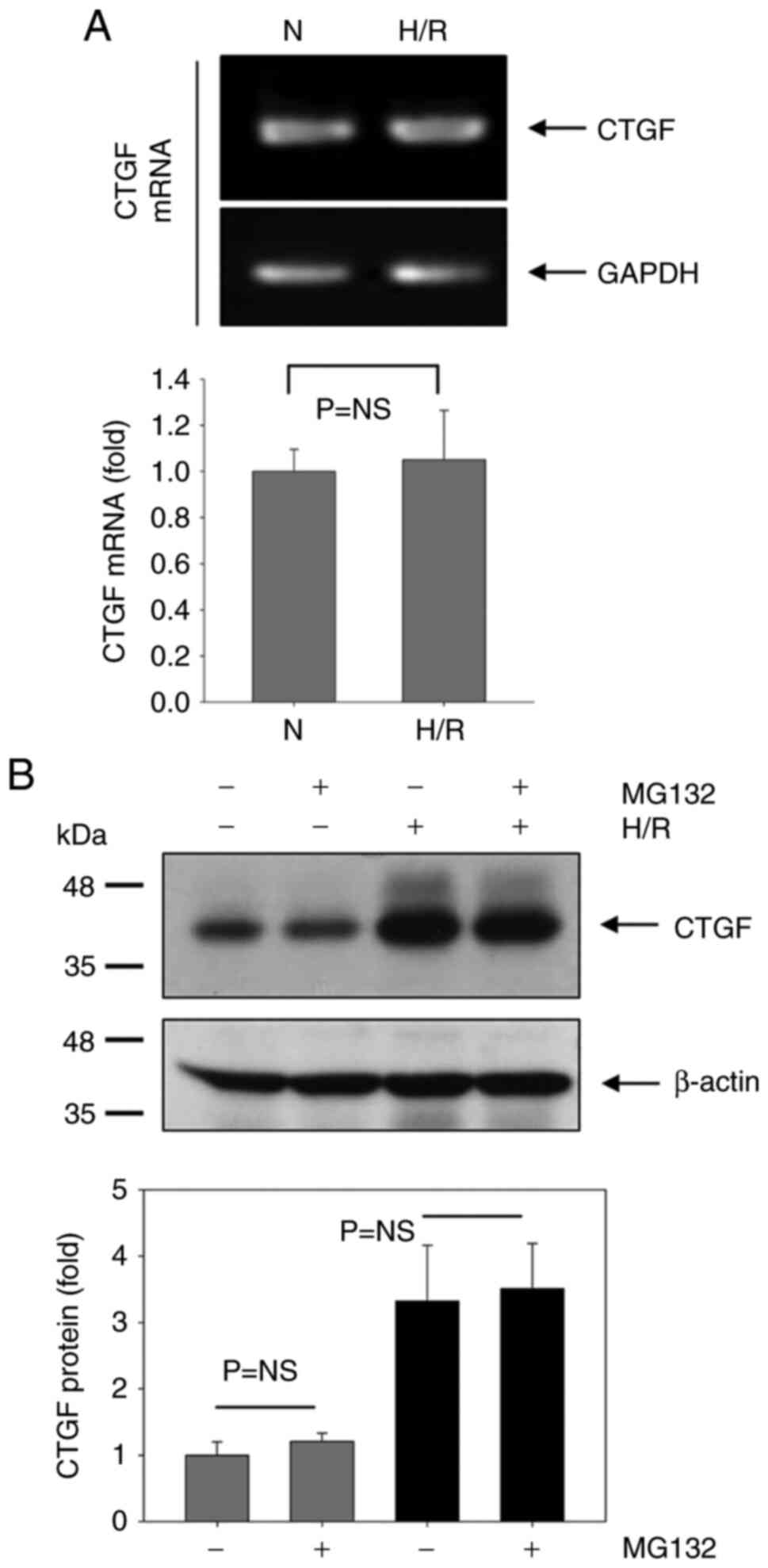
Subsequently, the mechanism underlying CTGF
upregulation by H/R in HUVECs was evaluated (Fig. 4). The CTGF mRNA level was first
examined to test whether H/R could affect the transcription of
CTGF. However, CTGF mRNA was not changed under H/R conditions
(Fig. 4A), although CTGF protein
expression was markedly increased by H/R (Fig. 4B). This suggests that increased
CTGF protein expression was not due to any changes in CTGF mRNA
levels under H/R conditions. The protein synthesis of CTGF by H/R
was therefore tested after blocking protein degradation with the
proteasome inhibitor MG132 (Fig.
4B). No significant differences in the protein synthesis of
CTGF was detected between the normoxia and H/R groups with the
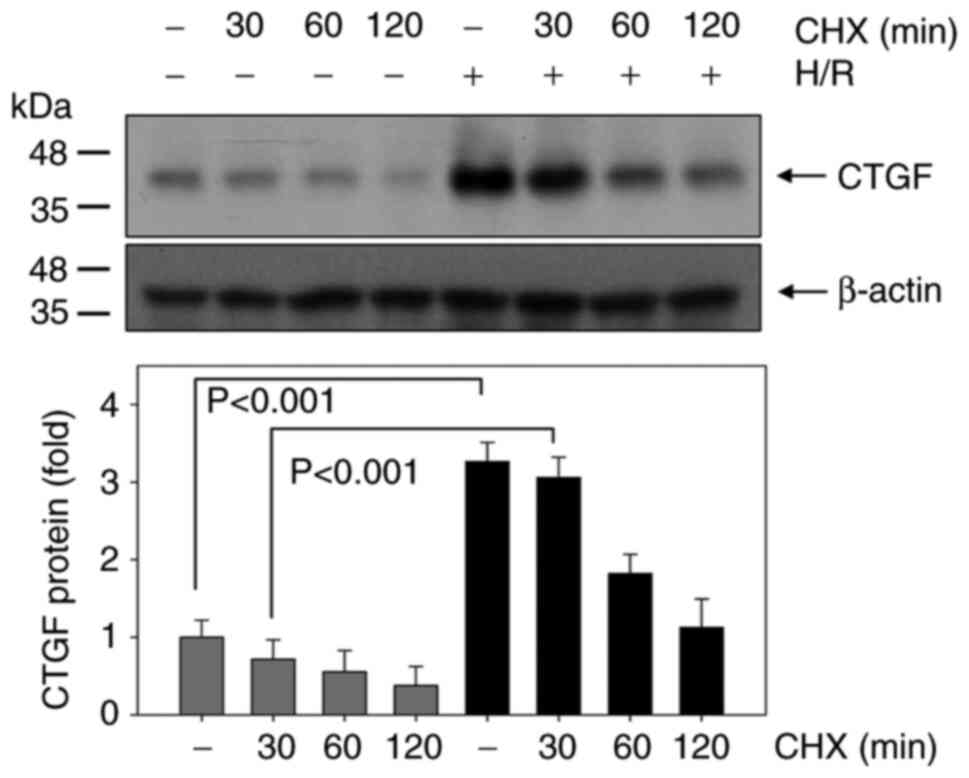
presence or absence of MG132. Changes in CTGF stability following
H/R was then examined using CHX, a protein translation blocker
(Fig. 5). Notably, CTGF stability
was significantly higher in the H/R group compared with that in the
normoxic control group at 30 and 60 min (Fig. 5).
Discussion
The tumor microenvironment can contain fibroblasts,
immune cells, blood vessels and the ECM (8). Fibroblasts are typically quiescent
but can be activated into myofibroblasts during the wound-healing
response (6). In addition, CAFs
can directly regulate cancer cell proliferation, tumor immunity,
angiogenesis, ECM remodeling and metastasis, suggesting that they
can be a target for anti-cancer therapy (8). Several preclinical studies have
reported CAFs to be possible targets for anti-cancer therapy in
lymphoma, Lewis lung cancer, melanoma and gastrointestinal cancer
(6-8).
α-SMA is a marker that can be used to reflect the myofibroblast
population of CAFs, such that docetaxel-conjugate nanoparticles
have been shown to target α-SMA+ stromal-suppressed
metastases in a mouse model of breast cancer (24). Furthermore, selective depletion of
myofibroblasts has been documented to attenuate angiogenesis in a
pancreatic ductal adenocarcinoma mouse model. However, depletion of
α-SMA+ myofibroblasts in mouse pancreatic cancer can
also increase the population of immunosuppressive
CD4+Foxp3+ Tregs, leading to more invasive
tumors (25). In the present
study, CM from HUVECs under H/R conditions, in addition to that
from CTGF-overexpressing HUVECs, was found to activate fibroblasts
into α-SMA+ myofibroblasts. This suggests that blood
vessels can promote neighboring fibroblasts into differentiating
into myofibroblasts. A clinical trial of bevacizumab targeting
endothelial cell precursors with CAFs has previously been
conducted; the addition of bevacizumab to the standard of care
significantly improved overall survival in malignant pleural
mesothelioma (26). Although
targeting α-SMA+ myofibroblasts therapeutically remains
to be a controversial topic (24,25),
targeting only CAFs, CAFs with endothelial cells or other types of
cells is a promising strategy (26). However, future studies are required
to define this strategy more precisely.
CTGF is mainly secreted from endothelial cells and
can modulate complex biological processes during normal embryonic
development and tissue repair (10). Abnormal CTGF expression profiles
have been observed in several diseases, including tissue fibrosis
(of the lung, heart and liver), systemic sclerosis and tumors in
major organs (23,27). It has been previously reported that
>30 types of human cancers are associated with the dysregulated
aberrant expression of CTGF (27).
Higher CTGF expression is associated with more aggressive
inflammatory colorectal cancer, whilst CTGF expression has also
been found to be increased in breast cancer, chondrosarcoma and
glioma (10,27). By contrast, CTGF can also function
as a tumor suppressor. CTGF expression has been observed to be
reduced in non-small cell lung cancer cells, where decreased CTGF
expression may be involved in lung tumorigenesis (10,28).
In the present study, CTGF from HUVECs stimulated fibroblast
differentiation, suggesting a possible association of HUVECs with
CAF generation. Further investigations on the functional roles of
CTGF-expressing endothelial cells and myofibroblasts in tumors or
ischemic diseases are required.
CTGF can bind to several types of receptors, such as
integrins, heparan sulfate proteoglycans, lipoprotein
receptor-related proteins and tyrosine kinase receptors (29). However, integrins are known to be
the principal CTGF receptors (29). CTGF mediates downstream effects
through α6, β1, αV and
β3 integrins (15).
Physiologically, CTGF enhances the lactogenic differentiation of
mammary epithelial cells by binding to integrins α6 and
β1, and to a lesser degree β3 integrin
(15). In addition, CTGF can
activate β1 integrin signaling in primary skin
fibroblasts (30) and pancreatic
stellate cells through αVβ3 (13,31).
In the present study, CM from CTGF-overexpressing endothelial cells
stimulated fibroblast differentiation, which was inhibited by a
function-blocking antibody against integrin
αVβ3, but not by a function-blocking antibody
against integrin α6.
The present study has a number of limitations.
Although it was demonstrated that H/R increased CTGF expression in
HUVECs and had a direct effect on the differentiation of
fibroblasts to myofibroblasts through integrin
αVβ3, the mechanism of integrin-mediated
fibroblast differentiation by H/R endothelial CM was not explored.
Integrin-linked kinase (ILK) is a key mediator of integrin
signaling that interacts with the cytoplasmic domain of β-integrins
(32). Therefore, ILK may be a
downstream candidate in this case. It has previously been reported
that integrin αVβ3 is involved in the stress
fiber formation through signaling molecules, such as focal adhesion
kinase (FAK), PKCα and RhoA (33,34).
In human aortic smooth muscle cells, osteoprotegerin, a ligand for
integrin αVβ3, mediated the phosphorylation
of FAK and actin cytoskeleton reorganization (34). Integrin αVβ3
triggers the formation of focal adhesions and stress fibers through
the activation of the transforming protein RhoA in astrocytes
(33). Another limitation of the
present study is that the binding of CTGF onto integrin
αVβ3 on fibroblasts was not confirmed.
CTGF-overexpressing HUVEC CM facilitated the differentiation of
fibroblasts into myofibroblasts, which was inhibited by a
functional blocking antibody against integrin
αVβ3. Therefore, it is highly likely that
CTGF will bind to αVβ3. However, more
specific assays, such as a CTGF adhesion assay (15), are required to investigate the
possible interaction between CTGF and integrin
αVβ3. The previous report by Morrison et
al (15) demonstrated the
interaction between CTGF and integrin complexes using an adhesion
assay. HC11 epithelial cells adhered onto CTGF-coated wells, where
function-blocking antibodies against both α6 and
β1 interrupted this CTGF-mediated epithelial cell
adhesion (15). Further studies
are required to determine the underlying mechanism(s) of
CTGF/integrin αVβ3-mediated fibroblast
differentiation, especially the stress fiber formation in
fibroblasts.
Furthermore, another limitation of the present study
is that the proteasomal degradation and stability of CTGF was
analyzed using cellular homogenates. Since CTGF is a secreted
protein, it is not sufficient to analyze proteasomal degradation
and stability using cell lysates. Interestingly, whilst CTGF is a
secreted protein, CTGF can also strongly bind to heparin and other
matrix components, rendering it detectable in the supernatants or
in cellular homogenates, depending on the cell type investigated
(35,36). Although CTGF (38 kDa) was readily
detected in cell lysates, it was not detectable in the conditioned
medium (35). Upon stimulation of
CTGF with serotonin, enhanced levels of CTGF protein were detected
in the cellular homogenates, whereas no protein was detectable in
cell culture supernatants (37).
Therefore, the regulatory mechanisms associated with proteasomal
degradation or stability of CTGF, especially secreted CTGF, under
H/R and additional conditions require additional in-depth
investigations.
To conclude, the supernatants of CTGF-overexpressing
HUVECs stimulated fibroblast differentiation, which was
significantly inhibited by a function-blocking antibody against
integrin αVβ3. These findings suggest that
communication between CTGF-secreting endothelial cells and
neighboring fibroblasts can lead to the development of
myofibroblast-associated diseases, which may be a potential
therapeutic target for the treatment of cancer or ischemic
diseases.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the National
Research Foundation of Korea (grant no. 2020R1A2C1012564), funded
by the Government of South Korea (Ministry of Science and ICT).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SL was responsible for designing the research,
conducting the experiments, performing the data analysis and
writing the manuscript. SL confirms the authenticity of all the raw
data. SL has read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The author declares that they have no competing
interests.
References
|
1
|
Servais C and Erez N: From sentinel cells
to inflammatory culprits: Cancer-associated fibroblasts in
tumour-related inflammation. J Pathol. 229:198–207. 2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
McAnulty RJ: Fibroblasts and
myofibroblasts: Their source, function and role in disease. Int J
Biochem Cell Biol. 39:666–671. 2007.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Otranto M, Sarrazy V, Bonte F, Hinz B,
Gabbiani G and Desmouliere A: The role of the myofibroblast in
tumor stroma remodeling. Cell Adh Migr. 6:203–219. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Tomasek JJ, Gabbiani G, Hinz B, Chaponnier
C and Brown RA: Myofibroblasts and mechano-regulation of connective
tissue remodelling. Nat Rev Mol Cell Biol. 3:349–363.
2002.PubMed/NCBI View
Article : Google Scholar
|
|
5
|
Schafer M and Werner S: Cancer as an
overhealing wound: An old hypothesis revisited. Nat Rev Mol Cell
Biol. 9:628–638. 2008.PubMed/NCBI View
Article : Google Scholar
|
|
6
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ziani L, Chouaib S and Thiery J:
Alteration of the antitumor immune response by cancer-associated
fibroblasts. Front Immunol. 9(414)2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kobayashi H, Enomoto A, Woods SL, Burt AD,
Takahashi M and Worthley DL: Cancer-associated fibroblasts in
gastrointestinal cancer. Nat Rev Gastroenterol Hepatol. 16:282–295.
2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Leask A: Potential therapeutic targets for
cardiac fibrosis: TGFbeta, angiotensin, endothelin, CCN2, and PDGF,
partners in fibroblast activation. Circ Res. 106:1675–1680.
2010.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Chen Z, Zhang N, Chu HY, Yu Y, Zhang ZK,
Zhang G and Zhang BT: Connective tissue growth factor: From
molecular understandings to drug discovery. Front Cell Dev Biol.
8(593269)2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bradham DM, Igarashi A, Potter RL and
Grotendorst GR: Connective tissue growth factor: A cysteine-rich
mitogen secreted by human vascular endothelial cells is related to
the SRC-induced immediate early gene product CEF-10. J Cell Biol.
114:1285–1294. 1991.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Chuva de Sousa Lopes SM, Feijen A, Korving
J, Korchynskyi O, Larsson J, Karlsson S, ten Dijke P, Lyons KM,
Goldschmeding R, Doevendans P and Mummery CL: Connective tissue
growth factor expression and Smad signaling during mouse heart
development and myocardial infarction. Dev Dyn. 231:542–550.
2004.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Gao R and Brigstock DR: Connective tissue
growth factor (CCN2) induces adhesion of rat activated hepatic
stellate cells by binding of its C-terminal domain to integrin
alpha(v)beta(3) and heparan sulfate proteoglycan. J Biol Chem.
279:8848–8855. 2004.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Heng EC, Huang Y, Black SA Jr and Trackman
PC: CCN2, connective tissue growth factor, stimulates collagen
deposition by gingival fibroblasts via module 3 and alpha6- and
beta1 integrins. J Cell Biochem. 98:409–420. 2006.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Morrison BL, Jose CC and Cutler ML:
Connective Tissue Growth Factor (CTGF/CCN2) enhances lactogenic
differentiation of mammary epithelial cells via integrin-mediated
cell adhesion. BMC Cell Biol. 11(35)2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Becker TC, Noel RJ, Coats WS, Gómez-Foix
AM, Alam T, Gerard RD and Newgard CB: Use of recombinant adenovirus
for metabolic engineering of mammalian cells. Methods Cell Biol.
43:161–189. 1994.PubMed/NCBI View Article : Google Scholar
|
|
17
|
He TC, Zhou S, da Costa LT, Yu J, Kinzler
KW and Vogelstein B: A simplified system for generating recombinant
adenoviruses. Proc Natl Acad Sci USA. 95:2509–2514. 1998.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Smith RC, Branellec D, Gorski DH, Guo K,
Perlman H, Dedieu JF, Pastore C, Mahfoudi A, Denèfle P, Isner JM
and Walsh K: p21CIP1-mediated inhibition of cell proliferation by
overexpression of the gax homeodomain gene. Genes Dev.
11:1674–1689. 1997.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Deng YZ, Chen PP, Wang Y, Yin D, Koeffler
HP, Li B, Tong XJ and Xie D: Connective tissue growth factor is
overexpressed in esophageal squamous cell carcinoma and promotes
tumorigenicity through beta-catenin-T-cell factor/Lef signaling. J
Biol Chem. 282:36571–36581. 2007.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhou Y, Capuco AV and Jiang H: Involvement
of connective tissue growth factor (CTGF) in insulin-like growth
factor-I (IGF1) stimulation of proliferation of a bovine mammary
epithelial cell line. Domest Anim Endocrinol. 35:180–189.
2008.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rehn M, Veikkola T, Kukk-Valdre E,
Nakamura H, Ilmonen M, Lombardo C, Pihlajaniemi T, Alitalo K and
Vuori K: Interaction of endostatin with integrins implicated in
angiogenesis. Proc Natl Acad Sci. USA. 98:1024–1029.
2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Ma W, Tavakoli T, Derby E, Serebryakova Y,
Rao MS and Mattson MP: Cell-extracellular matrix interactions
regulate neural differentiation of human embryonic stem cells. BMC
Dev Biol. 8(90)2008.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Lee SW, Won JY, Kim WJ, Lee J, Kim KH,
Youn SW, Kim JY, Lee EJ, Kim YJ, Kim KW and Kim HS: Snail as a
potential target molecule in cardiac fibrosis: Paracrine action of
endothelial cells on fibroblasts through snail and CTGF axis. Mol
Ther. 21:1767–1777. 2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Murakami M, Ernsting MJ, Undzys E, Holwell
N, Foltz WD and Li SD: Docetaxel conjugate nanoparticles that
target α-smooth muscle actin-expressing stromal cells suppress
breast cancer metastasis. Cancer Res. 73:4862–4871. 2013.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Ozdemir BC, Pentcheva-Hoang T, Carstens
JL, Zheng X, Wu CC, Simpson TR, Laklai H, Sugimoto H, Kahlert C,
Novitskiy SV, et al: Depletion of carcinoma-associated fibroblasts
and fibrosis induces immunosuppression and accelerates pancreas
cancer with reduced survival. Cancer Cell. 28:831–833.
2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Zalcman G, Mazieres J, Margery J,
Greillier L, Audigier-Valette C, Moro-Sibilot D, Molinier O, Corre
R, Monnet I, Gounant V, et al: Bevacizumab for newly diagnosed
pleural mesothelioma in the Mesothelioma Avastin Cisplatin
Pemetrexed Study (MAPS): A randomised, controlled, open-label,
phase 3 trial. Lancet. 387:1405–1414. 2016.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ramazani Y, Knops N, Elmonem MA, Nguyen
TQ, Arcolino FO, van den Heuvel L, Levtchenko E, Kuypers D and
Goldschmeding R: Connective tissue growth factor (CTGF) from basics
to clinics. Matrix Biol. 68-69:44–66. 2018.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Chien W, Yin D, Gui D, Mori A, Frank JM,
Said J, Kusuanco D, Marchevsky A, McKenna R and Koeffler HP:
Suppression of cell proliferation and signaling transduction by
connective tissue growth factor in non-small cell lung cancer
cells. Mol Cancer Res. 4:591–598. 2006.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Jun JI and Lau LF: Taking aim at the
extracellular matrix: CCN proteins as emerging therapeutic targets.
Nat Rev Drug Discov. 10:945–963. 2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Chen CC, Chen N and Lau LF: The angiogenic
factors Cyr61 and connective tissue growth factor induce adhesive
signaling in primary human skin fibroblasts. J Biol Chem.
276:10443–10452. 2001.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gao R and Brigstock DR: A novel integrin
alpha5beta1 binding domain in module 4 of connective tissue growth
factor (CCN2/CTGF) promotes adhesion and migration of activated
pancreatic stellate cells. Gut. 55:856–862. 2006.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Naska S, Park KJ, Hannigan GE, Dedhar S,
Miller FD and Kaplan DR: An essential role for the integrin-linked
kinase-glycogen synthase kinase-3 beta pathway during dendrite
initiation and growth. J Neurosci. 26:13344–13356. 2006.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Avalos AM, Valdivia AD, Muñoz N,
Herrera-Molina R, Tapia JC, Lavandero S, Chiong M, Burridge K,
Schneider P, Quest AF and Leyton L: Neuronal Thy-1 induces
astrocyte adhesion by engaging syndecan-4 in a cooperative
interaction with alphavbeta3 integrin that activates PKCalpha and
RhoA. J Cell Sci. 122:3462–3471. 2009.PubMed/NCBI View Article : Google Scholar
|
|
34
|
He Y, Zou P, Lu Y, Jia D, Li X, Yang H,
Tang L, Zhu Z, Tu T, Tai S, et al: Osteoprotegerin promotes intimal
hyperplasia and contributes to in-stent restenosis: Role of an
αVβ3/FAK dependent YAP pathway. J Mol Cell Cardiol. 139:1–13.
2020.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Steffen CL, Ball-Mirth DK, Harding PA,
Bhattacharyya N, Pillai S and Brigstock DR: Characterization of
cell-associated and soluble forms of connective tissue growth
factor (CTGF) produced by fibroblast cells in vitro. Growth
Factors. 15:199–213. 1998.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Boes M, Dake BL, Booth BA, Erondu NE, Oh
Y, Hwa V, Rosenfeld R and Bar RS: Connective tissue growth factor
(IGFBP-rP2) expression and regulation in cultured bovine
endothelial cells. Endocrinology. 140:1575–1580. 1999.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Hahn A, Heusinger-Ribeiro J, Lanz T,
Zenkel S and Goppelt-Struebe M: Induction of connective tissue
growth factor by activation of heptahelical receptors. Modulation
by Rho proteins and the actin cytoskeleton. J Biol Chem.
275:37429–37435. 2000.PubMed/NCBI View Article : Google Scholar
|