Introduction
Thyroid cancer (TC) is the most frequent endocrine
malignant tumor, accounting for >90% of all endocrine
malignancies (1-3).
TC can be divided into three main histological types:
Differentiated (papillary and follicular TC), undifferentiated
(poorly differentiated and anaplastic TC) and medullary TC, which
originates from TC cells (4).
Anaplastic thyroid carcinoma (ATC), which is also termed
undifferentiated thyroid carcinoma, is a type of thyroid carcinoma
featuring high-grade morphology and high aggressiveness (5). ATC has an incidence of 1.5% in all
TCs and is the leading cause of all TC-related mortalities
(6). Due to the high levels of
extrathyroid invasion, distant metastasis and resistance to
conventional treatment, the median survival time of ATC is 3-9
months (7). Despite the use of
surgery, chemoradiotherapy, drug therapy and the development of
other novel methods for ATC treatment, the survival rate of ATC
patients is low because of high metastasis rate (8). Therefore, it is critical to explore
the molecular mechanisms involved in ATC and identify improved
therapeutic strategies for ATC.
K (lysine) acetyltransferase (KAT) is a class of
enzymes that catalyze the transfer of acetyl groups from acetyl-CoA
to the ε-amino groups of lysine found on specific protein
substrates (9). The acetylation of
lysine residues can affect the functions of certain proteins, such
as DNA-binding capacity, protein-protein interactions, enzyme
activity, chromosomal structure and the regulation of nuclear
transcription enzyme activity (10-12).
KAT can be divided into several families of enzymes, among which
Moz, YBF2, Sas2p and Tip (MYST), which is a pivotal one (13). Tip60, which is also termed KAT5, is
a main member of the MYST family of enzymes and acts as a crucial
participant in gene transcription, regulation of the cell cycle,
autophagy, tumorigenesis and metastasis (14). A previous study has shown that KAT5
silencing can inhibit proliferation and induce apoptosis of liver
cancer cells (15). KAT5 is also
involved in the suppression of the cell cycle and in the induction
of apoptosis of lung cancer cells (16). Overexpression of KAT5 increases the
aggressiveness of papillary thyroid cancer cells (17). In addition, KAT5 has been shown to
positively affect the invasive and migratory activities of ATC
cells (18). However, whether KAT5
affects the induction of cell apoptosis in ATC is unknown.
Therefore, the present study was designed to explore the effects of
KAT5 in ATC and investigate its underlying mechanism of action.
Materials and methods
Cell culture and treatment
The human ATC cell lines 8505C, FRO, KHM-5M and C643
and the normal thyroid cell line Nthy-ori 3-1 were provided by the
American Type Culture Collection and were cultured in DMEM
(Shanghai Zeye Biotechnology Co., Ltd.) in the presence of 10% FBS
(Cytiva) and 1% antibiotics (MilliporeSigma) at 37˚C with 5%
CO2. To investigate the autophagic pathway, 10 mM
3-methyladenine (3-MA) was applied to treat ATC cells for 2 h at
37˚C.
Cell transfection
The kinesin family member 11 (KIF11)-specific pcDNA
overexpression vector (ov-KIF11) and corresponding negative control
(ov-NC), the small interfering (si)-RNA targeting KAT5 (si-KAT5-1:
GGACAGCUCUGAUGGAAUACC; si-KAT5-2: GCAAUAAAUUGUUUCUAUAUG) and the
corresponding negative control (si-NC: 5'-UUCUCCGAACGUGUCACGU-3')
were provided by Genepharm, Inc. Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) was used for the
transfection of the aforementioned recombinants into 8505C cells
according to the manufacturer's protocols.
Cell Counting Kit-8 (CCK-8) assay
The evaluation of 8505C cell viability was assessed
utilizing the CCK-8 assay. Following incubation in 96-well plates,
the transfected cells were cultured in DMEM with 10% FBS.
Subsequently, the cells were incubated with 10 µl CCK-8 solution
(Abcam) for an additional 2 h. The absorbance was measured with a
microplate reader at λ=450 nm.
5-Ethynyl-2'-deoxyuridine (EdU)
assay
The 8505C cells were plated in six-well plates at a
density of 4x105 cells/well and then incubated at 37˚C
overnight. 8505C cells were mixed with 4% polyformaldehyde for 15
min at room temperature, fixed and incubated further in the
presence of 0.5% Triton X-100 for 15 min at room temperature.
Subsequently, the cells were stained with Cell-Light™ EdU Cell
Proliferation Detection Assay (Thermo Fisher Scientific, Inc.) and
counterstained with 4',6-diamidino-2-phenylindole (DAPI) (Beyotime
Institute of Biotechnology) for 10 min at room temperature.
Fluorescent microscopy was applied for counting the positive
cells.
Flow cytometry
Cell apoptosis was detected by fluorescein
isothiocyanate (FITC) Annexin V/propidium iodide (PI) Apoptosis
Detection kit I (Guangzhou RiboBio Co., Ltd.). Briefly, the
PBS-rinsed 8505C cells were re-suspended in binding buffer.
Following 10 min of exposure to 5 µl of Annexin V-FITC according to
the manufacturer's instructions, the cells were incubated with 10
µl PI at 4˚C for 30 min in the dark. Samples were run on a FC500MPL
flow cytometer (Beckman Coulter, Inc.) and FlowJo vX.0.7 software
(FlowJo LLC) was employed for the analysis of cell apoptosis. The
apoptotic rate was calculated using the formula: Apoptotic
rate=Q2+Q3. The results of the cell apoptosis analysis were
obtained from three different replications.
Reverse transcription-quantitative PCR
(RT-qPCR)
The extraction of total RNA from 1x104
treated cells was conducted employing TRIzol® reagent
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. The synthesis of RNA into cDNA was performed by using
PrimeScript RT Master Mix (Takara Bio, Inc.) according to the
manufacturer's instructions, which was provided by Takara Bio, Inc.
Subsequently, the cDNA was amplified by RT-qPCR using the SYBR
Premix Ex Taq II kit (Takara Bio, Inc.) according to the
manufacturer's instructions. The thermocycling conditions were as
follows: Initial denaturation at 95˚C for 3 min; followed by 40
cycles of denaturation at 95˚C for 30 sec, annealing at 60˚C for 30
sec and extension at 72˚C for 30 sec. The primer sequences for PCR
are presented as below: KAT5: 5'-AGTGGAGGGAGGGAAGATGG-3' (forward)
and 5'-TCTTCGTTGTCCTGGTTCCG-3' (reverse); KIF11:
5'-GCTTTCTCCTCGGCTCACTT-3' (forward) and 5'-GTGGGCACAGAGCCATTAGT-3'
(reverse); GAPDH: 5'-GGGAAACTGTGGCGTGAT-3' (forward) and
5'-GAGTGGGTGTCGCTGTTGA-3' (reverse). The comparative Cq method was
employed for the estimation of the relative gene expression levels
(19). The experiments were
replicated for three times.
Chromatin immunoprecipitation (CHIP)
assay
The EZ CHIP kit (Shanghai Huzheng Biotechnology Co.,
Ltd.) was employed for the CHIP assays. The cross-linking of the
cells was performed with 1% formaldehyde, whereas their quenching
with 2.5 M glycine. Then, 1x106 cells were collected via
centrifugation at 300 x g for 3 min at 25˚C and washed twice with
pre-chilled PBS. DNA was immunoprecipitated from the sonicated cell
lysates using 2 µg anti-KAT5 (1:30; cat. no. ab300521; Abcam) at
4˚C overnight. The following day, the samples were conjugated with
Protein A agarose (Invitrogen; Thermo Fisher Scientific, Inc.) for
6 h at 4˚C. A ChIP DNA purification kit (Beyotime Institute of
Biotechnology) was used to purify the immunoprecipitated DNA and it
was amplified by qPCR. RT-qPCR was applied to analyze the enriched
DNA.
Immunofluorescence staining
Initially, 8505C cells were subjected to 4%
paraformaldehyde fixation as well as 0.25% Triton X-100
permeabilization (Triton X-100 was only used for α-smooth muscle
actin, not cluster of differentiation 31). The overnight incubation
of 8505C cells was performed at 4˚C, in the presence of 2% BSA and
the primary antibody microtubule-associated protein 1A/1B-light
chain 3 (LC3) (cat. no. ab239416; Abcam). Subsequently, the samples
were incubated with DAPI (BIOSS) and FITC-conjugated secondary
antibodies IgG (H + L) (ProteinTech Group, Inc.) for LC3 in the
dark. A fluorescence microscope was used for the visualization and
the imaging of the cells.
Western blot assay
Total protein was extracted from 8505C cells using
RIPA buffer (GCS Auragen). The determination of the concentration
levels of the proteins was performed using the Detergent Compatible
Bradford Protein Assay kit (Beyotime Institute of Biotechnology).
Following their separation by PAGE in 10% SDS gels, the proteins
(30 µg/lane) were transferred to polyvinylidene fluoride membranes.
Subsequently, the membranes were blocked in 5% non-fat milk diluted
in 0.1% Tris-buffered saline with Tween 20 at room temperature for
2 h and incubated overnight with primary antibodies specific to
KAT5 (1:1,000; cat. no. ab300521; Abcam), Bcl-2 (1:1,000; cat. no.
ab32124; Abcam), Bax (1:1,000; cat. no. ab32503; Abcam), cleaved
caspase-3 (1:500; cat. no. ab32042; Abcam), Beclin1 (1:2,000; cat.
no. ab207612; Abcam), p62 (1:10,000; cat. no. ab109012; Abcam),
LC3II/I (1:2,000; cat. no. ab192890; Abcam), KIF11 (1:1,000; cat.
no. ab254298; Abcam) and GAPDH (1:2,500; cat. no. ab9485; Abcam) at
4˚C. The following morning, the membranes were incubated for 1 h
with secondary antibodies (1:2,000; cat. no. ab6721; Abcam) at room
temperature. Finally, the visualization of the protein bands was
conducted by employing an ECL detection system (Shanghai Yeasen
Biotechnology Co., Ltd.). QuantityOne 4.5.0 software (Bio-Rad
Laboratories, Inc.) was used for densitometry.
Statistical analysis
The data are displayed in the format of mean ±
standard deviation. The analysis was performed with SPSS 22.0
software (IBM Corp.) and GraphPad Prism 6 software (GraphPad
Software, Inc.). Significant differences were analyzed by one-way
ANOVA followed by the Bonferroni post-hoc test for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
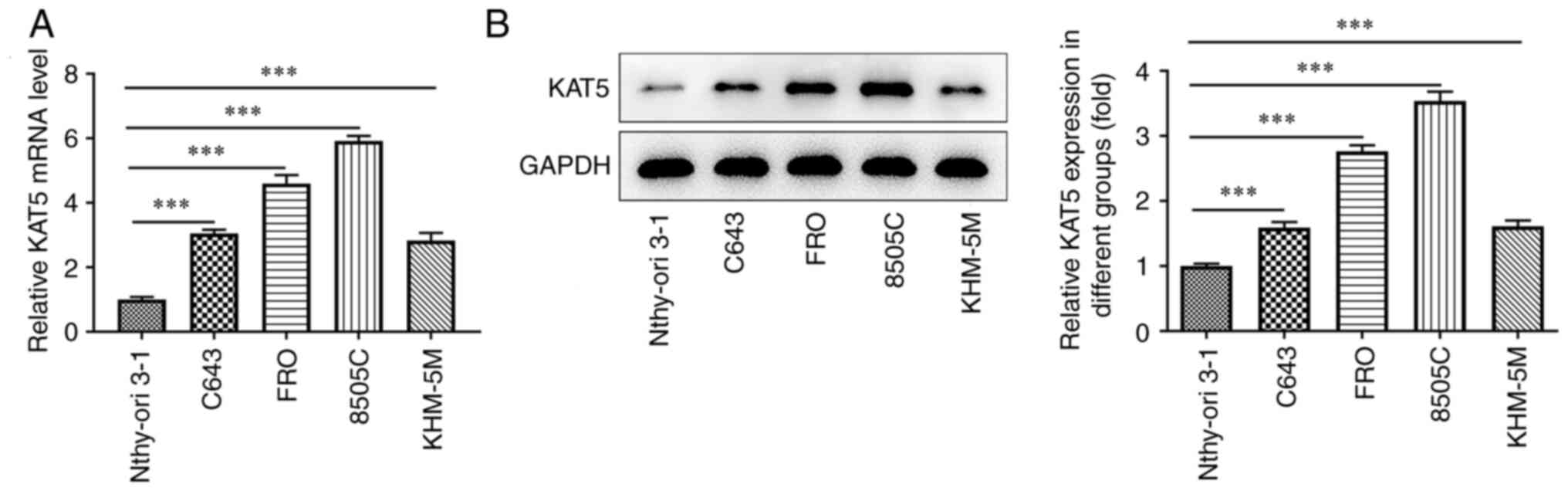
KAT5 expression is upregulated in ATC
cells
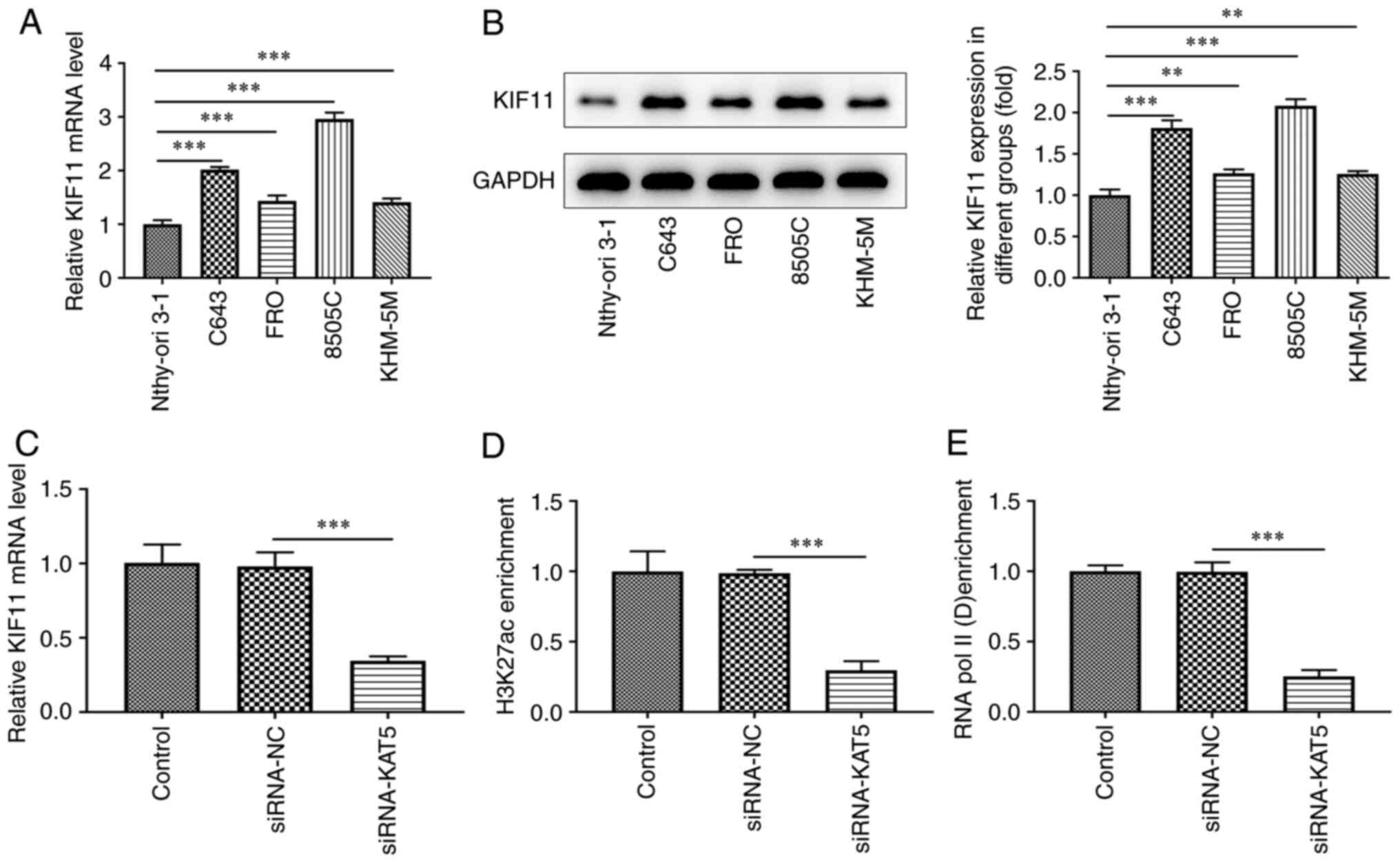
The expression levels of KAT5 in ATC cell lines were
initially appraised. The mRNA expression levels of KAT5 in the ATC
cell lines including 8505C, FRO and KHM-5M and in the C643 cell
line were increased relative to those noted in Nthy-ori 3-1 cells
(Fig. 1A). The results obtained
from the western blot assay indicated that KAT5 protein levels were
also upregulated in ATC cell lines compared with those of the
Nthy-ori 3-1 cell line (Fig. 1B).
Among these ATC cell lines, 8505C cells exhibited the highest
expression of KAT5. Therefore, 8505C cells were used for follow-up
studies.
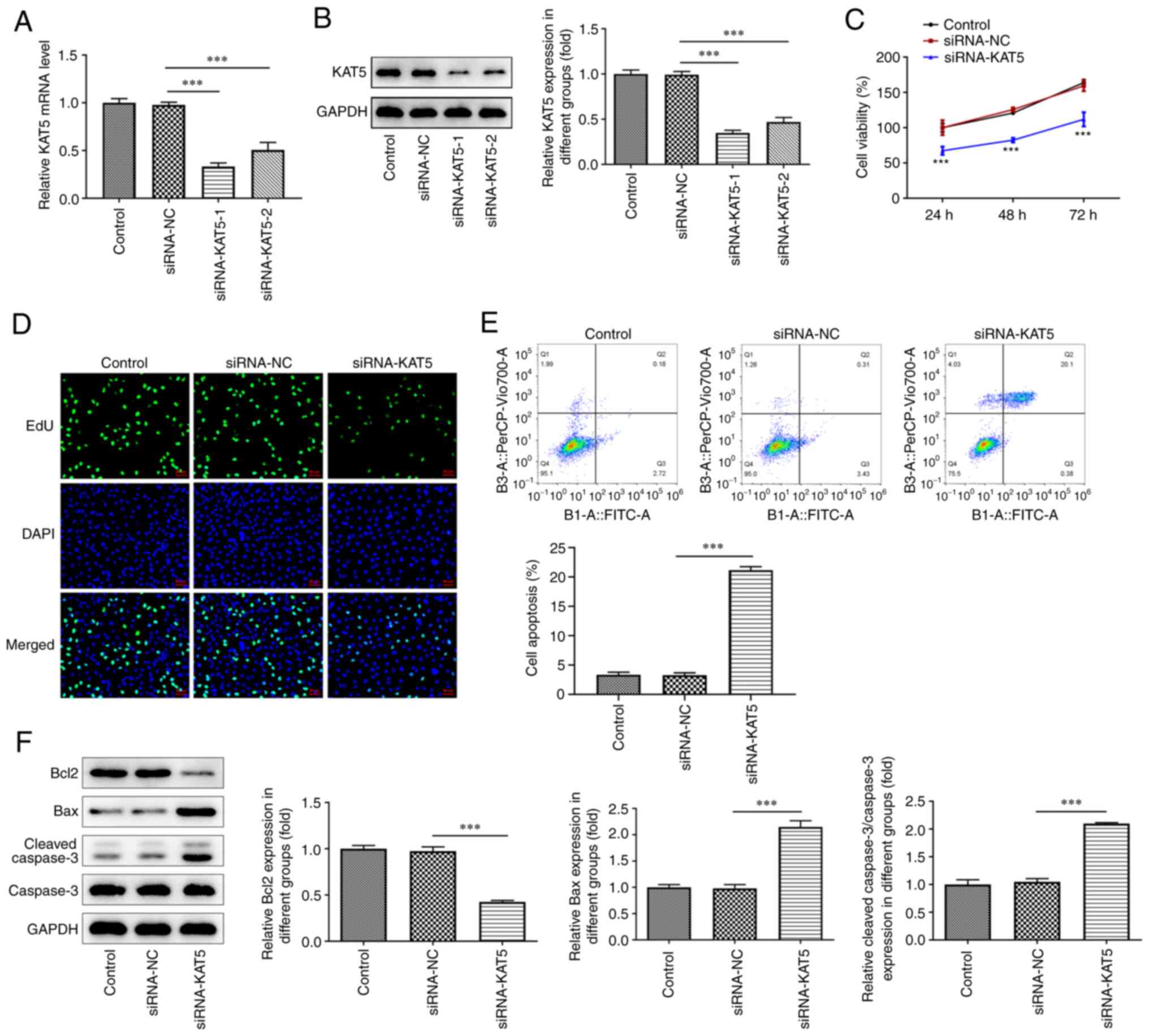
Knockdown of KAT5 expression inhibits
the proliferation and induces the apoptosis and autophagy of 8505C
cells
To explore the biological role of KAT5 in ATC,
si-KAT5-1/2 was transfected into 8505C cells to knockdown KAT5
expression. RT-qPCR and western blot analyses indicated that
si-KAT5-1 exhibited improved transfection efficiency. Therefore,
si-KAT5-1 was selected for subsequent analysis (Fig. 2A and B). The results obtained from the CCK-8
assay indicated that the capacity of cell proliferation was
remarkably inhibited following silencing of KAT5 compared with that
of the negative control samples (Fig.
2C). EdU staining further revealed that the number of positive
cells in the si-KAT5 group was significantly decreased (Fig. 2D). In addition, the induction of
cell apoptosis was notably promoted by knockdown of KAT5 expression
in 8505C cells (Fig. 2E). The data
from the western blot analysis indicated that KAT5 silencing
decreased Bcl-2 levels, whereas it also enhanced the contents of
Bax and cleaved caspase-3 (Fig.
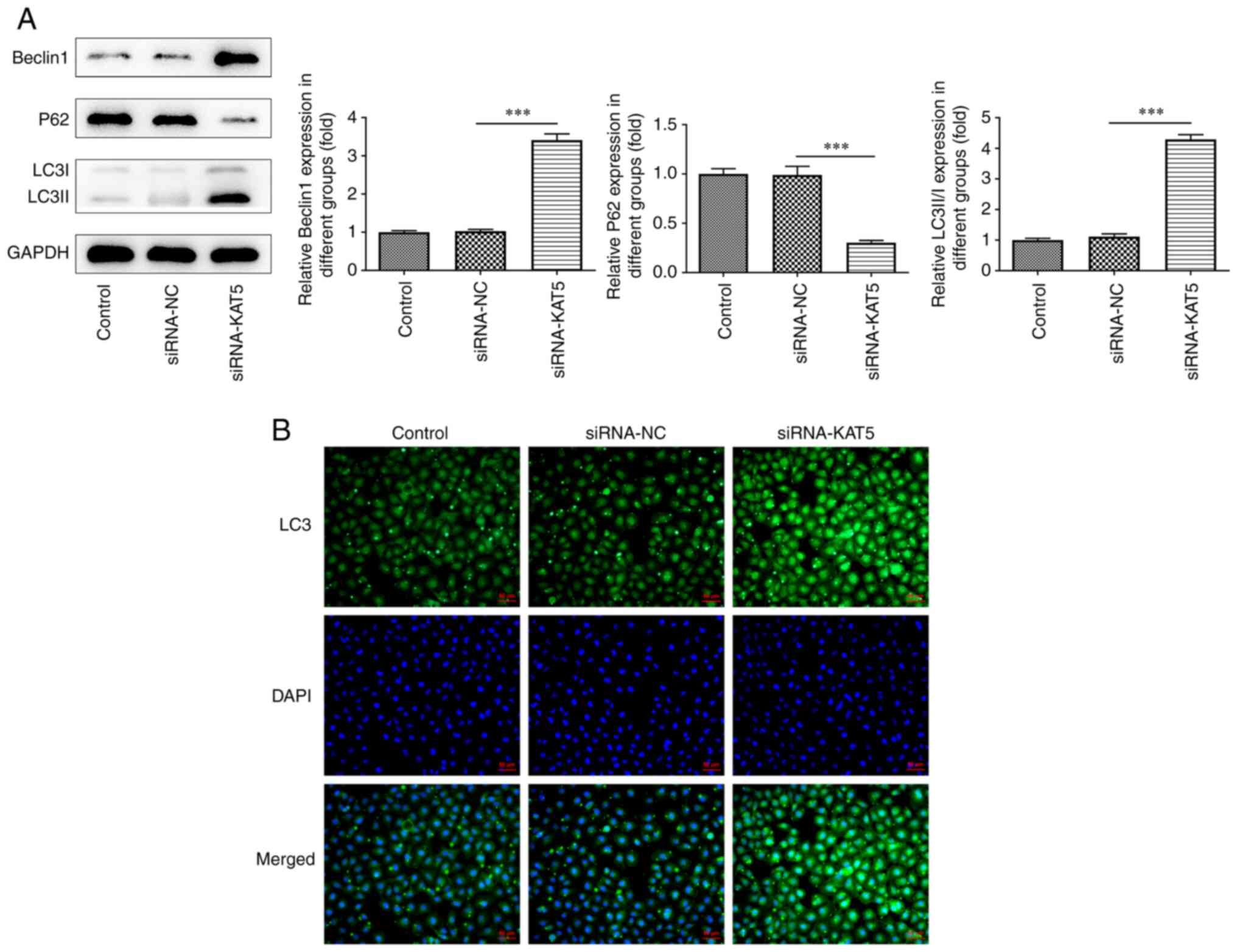
2F). In addition, silencing of KAT5 expression reduced p62
levels and promoted the levels of Beclin1 and LC3II/I compared with
those of the negative control samples (Fig. 3A), which was consistent with the
results of the immunofluorescence assay (Fig. 3B).
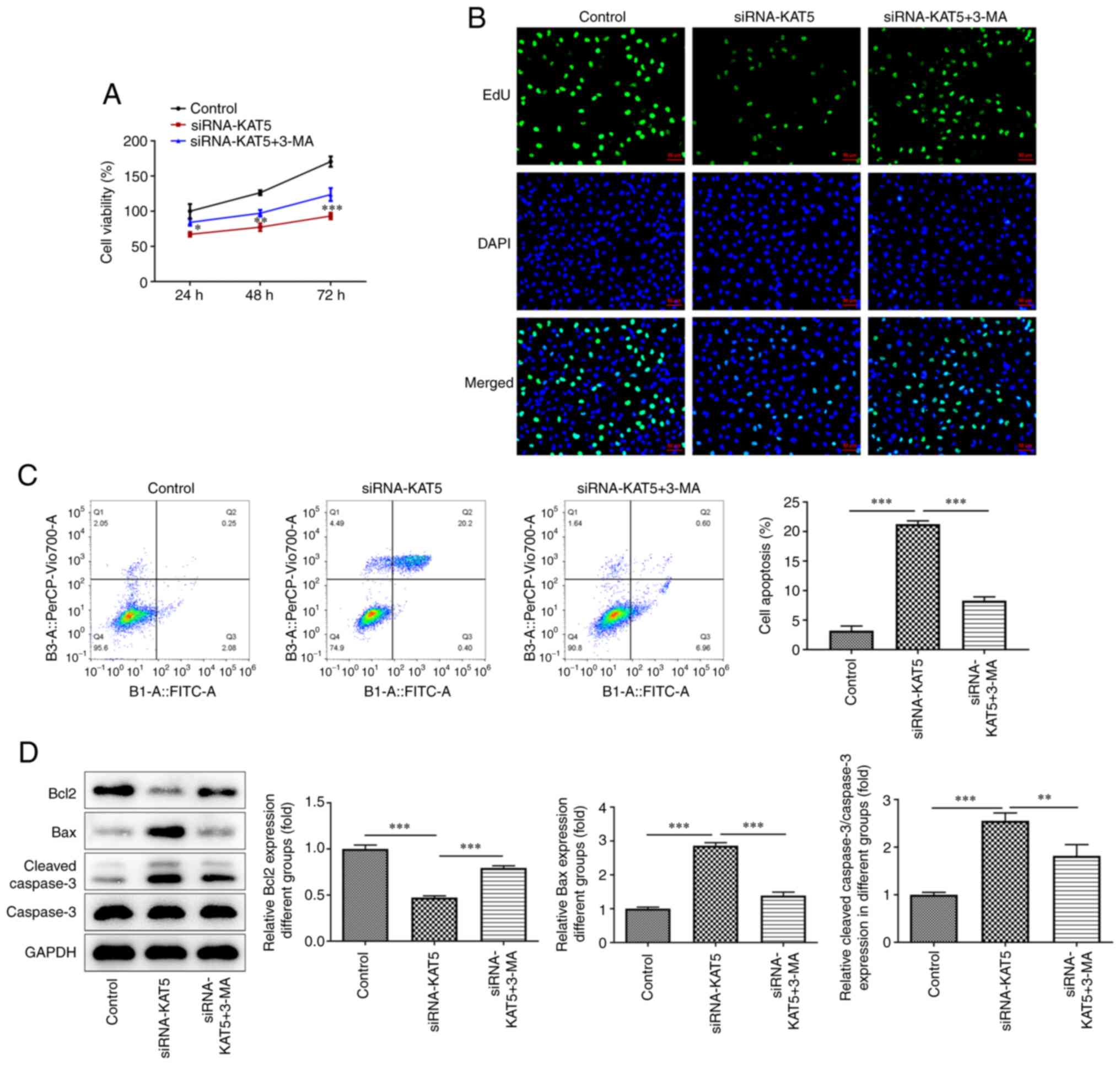
Silencing of KAT5 expression reduces
the proliferation and induces the apoptosis of 8505C cells by
promoting autophagy
To investigate the role of autophagy in
KAT5-mediated proliferation and apoptosis of 8505C cells, 10 mM of
the autophagy inhibitor 3-MA was administered to the cells.
Treatment of the cells with 3-MA enhanced their decreased
proliferation compared with the si-KAT5 group (Fig. 4A). Similarly, the EdU assay results
revealed that 3-MA treatment significantly increased the number of
positive cells (Fig. 4B). In
addition, 3-MA repressed the induction of apoptosis in
KAT5-silenced cells (Fig. 4C).
Subsequently, Bcl-2 levels were elevated, while Bax and cleaved
caspase-3 levels were decreased following the inhibition of
autophagy (Fig. 4D).
KAT5 depletion represses KIF11
expression in 8505C cells
Subsequently, the potential mechanism underlying
KAT5-mediated ATC cell apoptosis and autophagy was explored. The
mRNA and protein expression levels of KIF11 were elevated in the
ATC cell lines compared with those of the Nthy-ori 3-1 cell line
(Fig. 5A and B). The levels of KIF11 were suppressed by
KAT5 depletion (Fig. 5C). In
addition, the results obtained from the CHIP assay indicated that
the enrichment of H3K27ac and RNA pol II on the promoter region of
KIF11 was repressed following knockdown of KAT5 expression
(Fig. 5D and E).
Knockdown of KAT5 expression regulates
proliferation, apoptosis and autophagy by inhibiting KIF11
expression
Following the initial findings, the ability of KAT5
to affect ATC cell functions by targeting KIF11 expression was
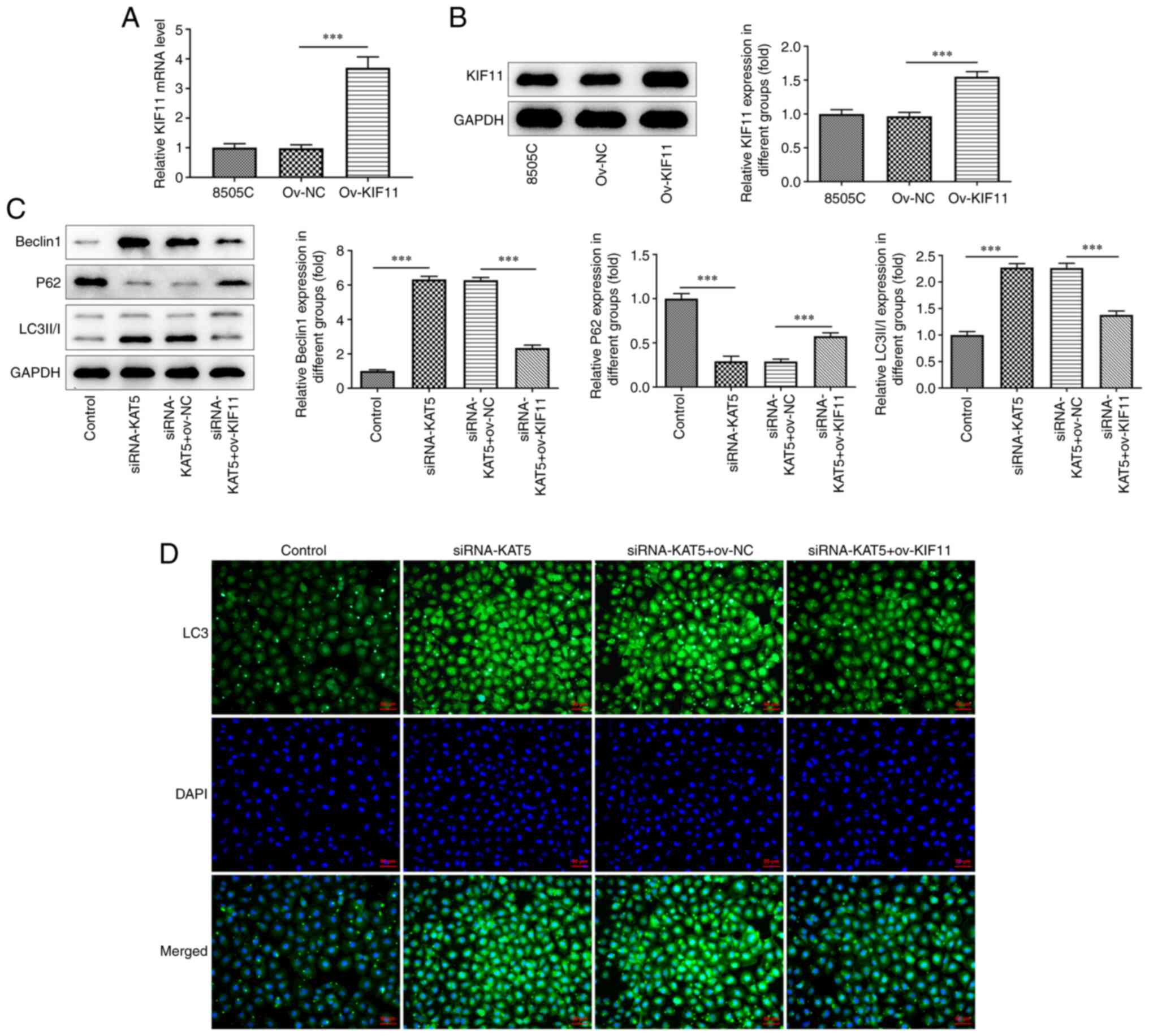
investigated. KIF11-overexpressing plasmids were constructed and
transfected into 8505C cells. RT-qPCR and western blot analyses
revealed that KIF11 overexpression promoted KIF11 expression
(Fig. 6A and B). KIF11 overexpression significantly
accelerated the production of p62 but reduced the levels of Beclin1
and LC3II/I in 8505C cells co-transfected with siRNA-KAT5 and
ov-KIF11 (Fig. 6C). The results of
the immunofluorescence assay revealed that KIF11 overexpression
evidently reduced the levels of LC3 compared with those of the
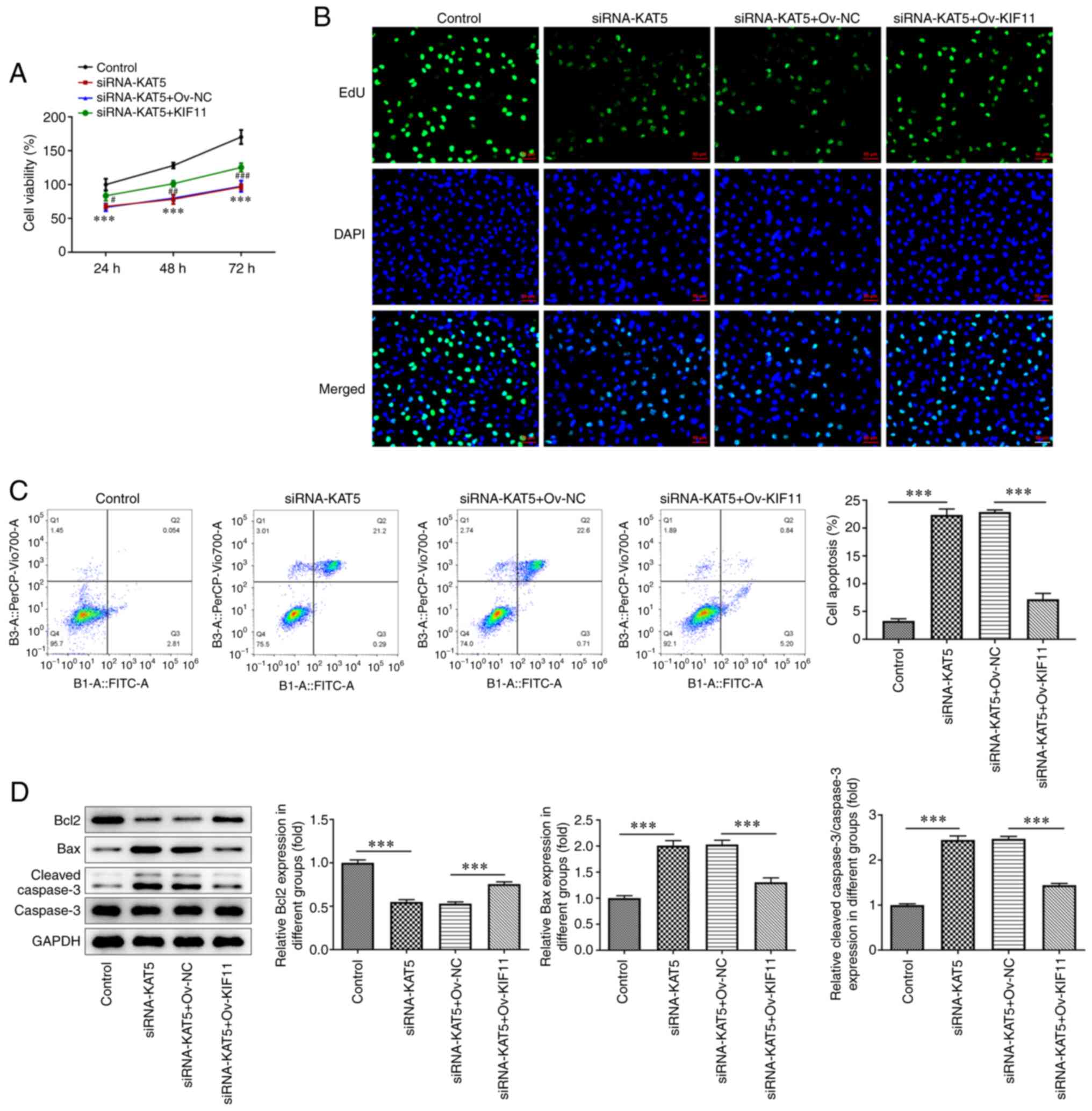
cells transfected with siRNA-KAT5 (Fig. 6D). Furthermore, the results of the
CCK-8 assay indicated that KIF11 overexpression reversed the
effects caused on cell proliferation compared with the negative
control cells (Fig. 7A); these
effects were accompanied with an increased number of positive 8505C
cells (Fig. 7B). In addition, the
cell apoptotic rate in KAT5-silenced cells was reversed following
transfection with ov-KIF11 plasmids (Fig. 7C). Finally, an increase in Bcl-2
levels and a reduction in the levels of Bax and cleaved caspase-3
were also noted in cells transfected with ov-KIF11 (Fig. 7D).
Discussion
As one of the deadliest human cancers, ATC accounts
for <2% of TCs (20). The
treatment for ATC includes surgery, chemotherapy, such as cisplatin
or doxorubicin, as well as accelerated hyperfractionated external
beam radiation therapy (21). Due
to the fact that ATC is a rare and aggressive tumor, it is still
challenging to predict the clinical responsiveness of patients to
treatment (22). Recently, several
genetic mutations and different molecular pathways, which are
involved in tumor progression, have been explored in ATC; moreover,
novel therapies acting on these molecular pathways have been
developed to improve the quality of life of patients with ATC
(23-25).
The present study aimed to explore the mechanism of KAT5/KIF11 in
ATC. The data demonstrated that KAT5 deficiency reduced KIF11
expression to promote the induction of apoptosis and autophagy in
ATC cells.
KATs are emerging as candidate therapeutic targets
for several cancer types; several inhibitors have been discovered
to target these proteins (26).
The MYST family, including KAT5-KAT8, has diverse functions that
affect a variety of cellular processes, including gene regulation,
cell cycle, stem cell homeostasis and development as well as the
DNA damage repair process (27).
KAT5, which is a critical element of the MYST family, is the
catalytic subunit of the NuA4 histone acetyltransferase complex.
This protein participates in the transcriptional activation of
specific genes (28) and the
complex is essential for triggering transcriptional programs which
have close relation with oncogene and proto-oncogene-modulated
growth induction, tumor suppressor-regulated growth arrest and
induction of apoptosis (29,30).
Feng et al (31)
demonstrated that KAT5 was involved in the progression of
gallbladder carcinoma tumorigenesis and was closely related to
disease prognosis. It was shown that KAT5 silencing induced
apoptosis of cells via the p38MAPK-mediated upregulation of cleaved
caspase-9. Kim and Lee (32)
demonstrated that KAT5 regulated the proliferation of prostate
cancer cells by a caspase-3-dependent apoptotic pathway, suggesting
that it was probably to be involved in the induction of apoptosis
of ATC cells. Therefore, the present study explored the mechanism
by which KAT5 participates in the induction of ATC apoptosis. The
data indicated that KAT5 expression was upregulated in ATC cells
compared with the corresponding expression in Nthy-ori 3-1 cells.
KAT5 interference repressed the proliferation of 8505C cells and
induced their apoptosis. Furthermore, the tumor-suppressive role of
KAT5 silencing was associated with the induction of autophagy.
As an acetyltransferase, KAT5 is involved in the
process of acetylation (33). It
has been suggested that histone acetylation can activate gene
transcription (34). For example,
KAT5 binds to programmed death-ligand 1 (PD-L1) and glutathione
peroxidase (GPX) 4 promoters through the H3K27ac acetylation site
to promote the transcription of PD-L1 and GPX4, indicating that it
may recruit transcription factors through the H3K27ac acetylation
site (35,36). In addition, Li et al
(37) revealed that KIF11 was
increased in ATC tissues. KIF11 has been shown to possess the
binding site of H3K27ac acetylation (38). A previous study has also shown that
KIF11 can activate the expression levels of AKT signaling members;
the activation of these proteins can inhibit the occurrence of
autophagy (39). A previous study
suggested that the activation of autophagy can induce apoptosis of
TC cells (40). In the present
study, it was found that silencing of KAT5 expression inhibited the
expression levels of KIF11 by repressing the enrichment of H3K27ac
and RNA pol II. KIF11 overexpression counteracted the effects of
KAT5 silencing on the proliferation, apoptosis and the induction of
autophagy of 8505C cells.
The present study concluded that silencing of KAT5
expression suppressed the proliferation and induced the apoptosis
and autophagy of ATC cells by targeting KIF11. The latter may serve
as a potential therapeutic target for ATC.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by The Growth Project
of Young Scientific and Technological Talents in General Colleges
and Universities in Guizhou Province, Qian Jiao He [grant number:
(2021) 186 and (2022) 242], National Natural Science Foundation of
China (grant number: 81960315), The Scientific and Technological
Plan Project of Guiyang City, Zhu Ke Contract [grant number: (2021)
43-7], The Science and Technology Plan Project in Guizhou Province,
Qian Ke He Foundation-ZK (grant number: 2022 General Project 403)
and The Graduate Research Fund in Guizhou Province, Qian Jiao He
[grant number: YJSKYJJ (2020) 144].
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LF and BZ designed the study, drafted and revised
the manuscript. LinZ, CL, PW, QW, JX and LixZ analyzed the data and
searched the literature. YG, SL, LS and XL confirm the authenticity
of all the raw data. All authors performed the experiments. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Laha D, Nilubol N and Boufraqech M: New
therapies for advanced thyroid cancer. Front Endocrinol (Lausanne).
11(82)2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Chmielik E, Rusinek D, Oczko-Wojciechowska
M, Jarzab M, Krajewska J, Czarniecka A and Jarzab B: Heterogeneity
of thyroid cancer. Pathobiology. 85:117–129. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Prete A, Borges de Souza P, Censi S, Muzza
M, Nucci N and Sponziello M: Update on fundamental mechanisms of
thyroid cancer. Front Endocrinol (Lausanne). 11(102)2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Bible KC, Kebebew E, Brierley J, Brito JP,
Cabanillas ME, Clark TJ Jr, Di Cristofano A, Foote R, Giordano T,
Kasperbauer J, et al: 2021 American thyroid association guidelines
for management of patients with anaplastic thyroid cancer. Thyroid.
31:337–386. 2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Molinaro E, Romei C, Biagini A, Sabini E,
Agate L, Mazzeo S, Materazzi G, Sellari-Franceschini S, Ribechini
A, Torregrossa L, et al: Anaplastic thyroid carcinoma: From
clinicopathology to genetics and advanced therapies. Nat Rev
Endocrinol. 13:644–660. 2017.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Gervasi R, Orlando G, Lerose MA, Amato B,
Docimo G, Zeppa P and Puzziello A: Thyroid surgery in geriatric
patients: A literature review. BMC Surg. 12 (Suppl
1)(S16)2012.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yang J and Barletta JA: Anaplastic thyroid
carcinoma. Semin Diagn Pathol. 37:248–256. 2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Fiorentino F, Mai A and Rotili D: Lysine
acetyltransferase inhibitors: Structure-activity relationships and
potential therapeutic implications. Future Med Chem. 10:1067–1091.
2018.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Numata A, Kwok HS, Zhou QL, Li J,
Tirado-Magallanes R, Angarica VE, Hannah R, Park J, Wang CQ,
Krishnan V, et al: Lysine acetyltransferase Tip60 is required for
hematopoietic stem cell maintenance. Blood. 136:1735–1747.
2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Li P, Ge J and Li H: Lysine
acetyltransferases and lysine deacetylases as targets for
cardiovascular disease. Nat Rev Cardiol. 17:96–115. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Proietti G, Wang Y, Punzo C and Mecinović
J: Substrate scope for human histone lysine acetyltransferase KAT8.
Int J Mol Sci. 22(846)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Li Y, Huang H, Zhu M, Bai H and Huang X:
Roles of the MYST family in the pathogenesis of Alzheimer's disease
via histone or non-histone acetylation. Aging Dis. 12:132–142.
2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Ghobashi AH and Kamel MA: Tip60: Updates.
J Appl Genet. 59:161–168. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yu Z, Chen T, Mo H, Guo C and Liu Q: BRD8,
which is negatively regulated by miR-876-3p, promotes the
proliferation and apoptosis resistance of hepatocellular carcinoma
cells via KAT5. Arch Biochem Biophys. 693(108550)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ren X, Yu J, Guo L and Ma H: Circular RNA
circRHOT1 contributes to pathogenesis of non-small cell lung cancer
by epigenetically enhancing C-MYC expression through recruiting
KAT5. Aging (Albany NY). 13:20372–20382. 2021.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Du J, Fu L, Ji F, Wang C, Liu S and Qiu X:
FosB recruits KAT5 to potentiate the growth and metastasis of
papillary thyroid cancer in a DPP4-dependent manner. Life Sci.
259(118374)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Wei X, Cai S, Boohaker RJ, Fried J, Li Y,
Hu L, Pan Y, Cheng R, Zhang S, Tian Y, et al: KAT5 promotes
invasion and metastasis through C-MYC stabilization in ATC. Endocr
Relat Cancer. 26:141–151. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Abe I and Lam AK: Anaplastic thyroid
carcinoma: Current issues in genomics and therapeutics. Curr Oncol
Rep. 23(31)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Perrier ND, Brierley JD and Tuttle RM:
Differentiated and anaplastic thyroid carcinoma: Major changes in
the American Joint Committee on Cancer eighth edition cancer
staging manual. CA Cancer J Clin. 68:55–63. 2018.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Lu X, Bao L, Pan Z and Ge M: Immunotherapy
for anaplastic thyroid carcinoma: The present and future. Zhejiang
Da Xue Xue Bao Yi Xue Ban. 50:675–684. 2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Haroon Al Rasheed MR and Xu B: Molecular
alterations in thyroid carcinoma. Surg Pathol Clin. 12:921–930.
2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bozorg-Ghalati F and Hedayati M: Molecular
biomarkers of anaplastic thyroid carcinoma. Curr Mol Med.
17:181–188. 2017.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cabanillas ME, Zafereo M, Gunn GB and
Ferrarotto R: Anaplastic thyroid carcinoma: Treatment in the age of
molecular targeted therapy. J Oncol Pract. 12:511–518.
2016.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Shvedunova M and Akhtar A: Modulation of
cellular processes by histone and non-histone protein acetylation.
Nat Rev Mol Cell Biol. 23:329–349. 2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Sapountzi V and Côté J: MYST-family
histone acetyltransferases: Beyond chromatin. Cell Mol Life Sci.
68:1147–56. 2011.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Doyon Y and Côté J: The highly conserved
and multifunctional NuA4 HAT complex. Curr Opin Genet Dev.
14:147–154. 2004.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Bird AW, Yu DY, Pray-Grant MG, Qiu Q,
Harmon KE, Megee PC, Grant PA, Smith MM and Christman MF:
Acetylation of histone H4 by Esa1 is required for DNA double-strand
break repair. Nature. 419:411–415. 2002.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Van Den Broeck A, Nissou D, Brambilla E,
Eymin B and Gazzeri S: Activation of a Tip60/E2F1/ERCC1 network in
human lung adenocarcinoma cells exposed to cisplatin.
Carcinogenesis. 33:320–325. 2012.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Feng FL, Yu Y, Liu C, Zhang BH, Cheng QB,
Li B, Tan WF, Luo XJ and Jiang XQ: KAT5 silencing induces apoptosis
of GBC-SD cells through p38MAPK-mediated upregulation of cleaved
Casp9. Int J Clin Exp Pathol. 7:80–91. 2013.PubMed/NCBI
|
|
32
|
Kim CH and Lee DH: KAT5 Negatively
regulates the proliferation of prostate cancer LNCaP cells via the
caspase 3-dependent apoptosis pathway. Anim Cells Syst (Seoul).
23:253–259. 2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Song ZM, Lin H, Yi XM, Guo W, Hu MM and
Shu HB: KAT5 acetylates cGAS to promote innate immune response to
DNA virus. Proc Natl Acad Sci USA. 117:21568–21575. 2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Martin BJE, Brind'Amour J, Kuzmin A,
Jensen KN, Liu ZC, Lorincz M and Howe LJ: Transcription shapes
genome-wide histone acetylation patterns. Nat Commun.
12(210)2021.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Xiao X, Zeng S, Li Y, Li L and Zhang J:
Aspirin Suppressed PD-L1 expression through suppressing KAT5 and
subsequently inhibited PD-1 and PD-L1 signaling to attenuate OC
development. J Oncol. 2022(4664651)2022.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Li H, Liu W, Zhang X, Wu F, Sun D and Wang
Z: Ketamine suppresses proliferation and induces ferroptosis and
apoptosis of breast cancer cells by targeting KAT5/GPX4 axis.
Biochem Biophys Res Commun. 585:111–116. 2021.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Li L, Zhu M, Huang H, Wu J and Meng D:
Identification of hub genes in anaplastic thyroid carcinoma:
Evidence from bioinformatics analysis. Technol Cancer Res Treat.
19(1533033820962135)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Wei D, Rui B, Qingquan F, Chen C, Ping HY,
Xiaoling S, Hao W and Jun G: KIF11 promotes cell proliferation via
ERBB2/PI3K/AKT signaling pathway in gallbladder cancer. Int J Biol
Sci. 17:514–526. 2021.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhou J, Jiang YY, Chen H, Wu YC and Zhang
L: Tanshinone I Attenuates the malignant biological properties of
ovarian cancer by inducing apoptosis and autophagy via the
inactivation of PI3K/AKT/mTOR pathway. Cell Prolif.
53(e12739)2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Long HL, Zhang FF, Wang HL, Yang WS, Hou
HT, Yu JK and Liu B: Mulberry anthocyanins improves thyroid cancer
progression mainly by inducing apoptosis and autophagy cell death.
Kaohsiung J Med Sci. 34:255–262. 2018.PubMed/NCBI View Article : Google Scholar
|