Introduction
Atherosclerosis is one of the most common
pathological mechanisms of cardiovascular diseases. Increasing
evidence suggested that patients with diabetes have a significantly
increased risk of atherosclerosis (1,2).
Even though antiplatelet and lipid-lowering treatments now
significantly improve the prognosis of patients with
atherosclerosis, there remains a residual cardiovascular risk in
certain patients, particularly those with diabetes. Therefore,
there is an urgent need to find new potential targets for
intervention against diabetic atherosclerosis.
There are increasing evidence demonstrated that
inflammation is a key factor in the development of atherosclerosis
(3). The CANTONS study noted that
anti-inflammatory therapy targeting IL-1β may provide a limited
reduction in the incidence of major adverse cardiovascular events
in patients with acute myocardial infarction (4). In addition, interventions with IL-6
and NLRP3 inflammasome have also been found to reduce the incidence
of atherosclerosis and complications. All of these data illustrated
that targeting inflammatory pathways may be an effective
intervention for atherosclerosis treatment (5,6). In
addition to classical inflammatory factors, including IL-1β, TNF-α
and IL-8, an increasing number of novel inflammatory factors are
being identified due to their cardiovascular protective effects
(7). IL-37 is a newly identified
anti-inflammatory factor; it was found that IL-37 is significantly
elevated in monocytes as well as in peripheral blood in patients
with coronary artery disease and acute myocardial infarction
(8,9). Moreover, IL-37 levels have also been
shown to be strongly associated with the long-term prognosis of
patients with acute myocardial infarction and acute stroke
(10-13).
Furthermore, it was also reported that overexpression of IL-37
significantly increases plaque instability in ApoE-/-
mice (14). Furthermore, in
vitro experiments demonstrated that IL-37 attenuated
atherosclerosis by suppressing T cell activation, inducing the
response of regulatory T cells and inhibiting vascular
calcification (11,15-17).
However, to the best of our knowledge, the effect and underlying
mechanism of IL-37 on diabetic atherosclerosis remains unclear.
Ferroptosis is a form of programmed death
characterized by lipid oxidation in the cell membrane. There is
growing evidence that ferroptosis is extensively involved in the
development and progression of atherosclerosis. It was reported
that high levels of uric acid promote the progression of
atherosclerosis by increasing levels of oxidative stress and
enhancing ferroptosis of macrophages (18). The use of a ferroptosis-specific
inhibitor, Fer-1, has also been found to be effective in inhibiting
the progression of atherosclerosis (19,20).
Glutathione peroxidase 4 (GPX4), is mainly responsible for
regulating glutathione synthesis in cells. When the expression of
GPX4 is inhibited in cells, glutathione synthesis in cells becomes
dysfunctional, which results in ferroptosis (21-23).
Thus, the decrease of GPX4 was also regarded as the marker of
ferroptosis (23). Several studies
have revealed that IL-37 has a potent ability to scavenge oxygen
radicals (8,11). However, whether IL-37 has a
protective effect on ferroptosis of macrophages and the related
mechanism remains unclear.
In the present study, the protective effect of IL-37
on diabetic atherosclerosis was confirmed; it was also found that
IL-37 may attenuate the progression of atherosclerosis by
suppressing ferroptosis of macrophages. The present study provided
new evidence that IL-37 protects against atherosclerosis.
Materials and methods
Animal model
The present experiment was approved (approval no.
SYDW2020-251) by the Ethics Committee of the Fourth Affiliated
Hospital of Harbin Medical University (Harbin, China). The mice
were housed in the animal center of the Second Affiliated Hospital
of Harbin Medical University (22±2˚C, 55±5% relative humidity with
a 12/12-h light/dark cycle). The mice were granted free access to
food and water. Male ApoE-/- mice (8-week-old, 20-25 g)
were randomly divided into two groups of 10 mice each. According to
a previous study (24), a diabetes
model was constructed using streptozotocin (STZ) intraperitoneal
injection (50 mg/kg/day for five consecutive days) after 4 weeks of
feeding using a high-fat diet. ApoE-/- mice with
continuous blood glucose >15 mM were used as a model of
diabetes. The mice were all administered a high-fat diet and
divided into STZ/HD and IL-37 groups. Saline and IL-37 were
administered intraperitoneally (1 µg/week), respectively (15). After 12 weeks, all animals were
anesthetized with an intraperitoneal injection of sodium
pentobarbital (35 mg/kg body weight) and then euthanized by
cervical dislocation; serum and vascular tissue were collected.
Cell culture
THP-1 cells were cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) and passaged once every 2-3
days. A total of 100 nM Phorbol-12-Myristate-13-Acetate was used to
treat THP-1 cells for 48 h to induce into macrophages. Macrophages
were divided into four groups: i) Control group (DMSO), ii) high
glucose (HG)/ox-LDL (100 ng/ml ox-LDL, 25 mM glucose for 24 h),
iii) IL-37/HG/ox-LDL (30 µM IL-37 for 0.5 h, 100 ng/ml ox-LDL, 25
mM glucose for 24 h) and iv) ML385 group (5 µM ML385 for 0.5 h, 30
µM IL-37 for 0.5 h, 100 ng/ml ox-LDL, 25 mM glucose for 24 h).
Western blot analysis
Macrophages and mouse aortic tissue were used to
extract protein lysates using RIPA (Beyotime Institute of
Biotechnology). Protein concentration was measured using a BCA kit
(Beyotime Institute of Biotechnology). Protein (20 µg/lane) was
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene fluoride
membranes (MilliporeSigma) using a semidry transblot apparatus
(Bio-Rad Laboratories, Inc.). Next, the polyvinylidene fluoride
membranes were blocked using 5% skimmed milk at room temperature
for 60 min and the membrane was washed 3 times with PBS for 10 min
each. PVDF membranes were incubated with the primary antibody
(anti-GPX4 antibody: cat. no. 59735; 1:1,000; CST Biological
Reagents Co., Ltd.; anti-NRF2 antibody: cat. no. 12721; 1:1,000;
CST Biological Reagents Co., Ltd.; anti-Histone H3 antibody: cat.
no. 14499; 1:2,000; CST Biological Reagents Co., Ltd.;
anti-β-actin: cat. no. AC006; 1:1,000; ABclonal Biotech Co., Ltd.)
overnight at 4˚C. The next day, three washes using PBS for 10 min
each followed. Subsequently, the membranes were incubated for 1 h
at room temperature using HRP-labeled Goat Anti-Rabbit IgG (cat.
no. A0208; 1:10,000; Beyotime Institute of Biotechnology). The
immuno-reactive bands were detected by chemiluminescence methods
and visualized using the Luminescent Imaging Workstation (Tanon
Science and Technology Co., Ltd.). The relative intensities of the
bands were measured and analyzed using the ImageJ software v.1.4
(National Institutes of Health).
Immunofluorescence staining
To detect nuclear translocation of nuclear factor
erythroid 2-related factor 2 (NRF2), THP-1 macrophages were treated
with 4% paraformaldehyde for 10 min at room temperature, washed
three times with PBS and then treated with 0.3% Triton for 5 min at
room temperature to disrupt the cell membrane. The cells were then
washed three times with PBS at room temperature and 5% bovine serum
albumin was added followed by incubation for 60 min to reduce
non-specific staining at room temperature. The macrophages were
incubated overnight at 4˚C with primary antibody against NRF2 (cat.
no. 12721; 1:100; CST Biological Reagents Co., Ltd.). Next, the
macrophages were washed with PBS and incubated for 60 min at room
temperature in the dark with a fluorescent Goat Anti-Rabbit
secondary antibody (cat. no. ab150077; 1:400; Abcam). The
macrophages were observed and images were captured on a confocal
laser microscope (LSM 800) at x200 magnification.
Malondialdehyde (MDA), reduced
glutathione (GSH) and superoxide dismutase (SOD measurement)
The lysates of macrophages were extracted according
to the manufacturer's instructions, and the levels of MDA (cat. no.
S0131S; Beyotime Institute of Biotechnology), GSH (cat. no. S0053;
Beyotime Institute of Biotechnology) and SOD (cat. no. S0101S;
Beyotime Institute of Biotechnology) were measured using kits, and
the absorbance (MDA, 532 nm; GSH, 412 nm; SOD, 450 nm) was analyzed
using an enzyme marker.
C11 BODIPY measurement
According to the manufacturer's instructions,
1x106 cells/ml were treated and digested using trypsin.
PBA wash was performed 3 times and the level of lipid oxidation in
the cell membrane was detected using C11 BODIPY (Thermo Fisher
Scientific, Inc.) staining reagent (1:1,000 with PBS). After
incubation for 30 min at room temperature and protected from light,
PBS was washed. The absorbance was detected using flow cytometry
(581/591).
Cell viability assays
Macrophages were cultured in 96 well plates with
10,000 cells per well. The Cell Counting Kit-8 (CCK-8) (Beyotime
Institute of Biotechnology) assays were performed according to the
manufacturer's instructions. Briefly, after preparing the
experimental groups and performing the cell treatments, the CCK-8
working solution was diluted 10-fold and 100 µl was removed and
added to 96-well plates. The plates were placed in an incubator
(37˚C, 60 min) to allow the reaction to occur. After the
incubation, absorbance was measured at 450 nm using a microplate
spectrophotometer (Tecan Group, Ltd.).
Lactate dehydrogenase (LDH)
assays
Macrophages were cultured in 96 well plates with
10,000 cells per well. According to the LDH Assay Kit (cat. no.
C0016; Beyotime Institute of Biotechnology) instructions, the cell
culture plates after drug stimulation were centrifuged at 400 x g
for 5 min using a multi-well plate centrifuge at room temperature.
The LDH-releasing reagent provided in the kit was diluted 10-fold
with PBS and mixed well. The cell supernatant was aspirated and 150
µl of the diluted LDH-releasing reagent was added to each cell
sample. The plates were shaken to assure the LDH-releasing reagent
in each well was mixed well. The plates were then incubated (37˚C)
for 1 h. The cell culture plates were subsequently centrifuged for
5 min at room temperature at 400 x g in a multi-well plate
centrifuge. The supernatant (120 µl) was collected from each well
and separately added to the corresponding wells of a new 96-well
plate. Thereafter the absorbance was measured at 490 nm.
Hoechst/propidium iodide (PI)
staining
Cells were seeded into 24-well plates with 50,000
cells per well. After the cells were treated according to the
experimental design, the cells were washed three times with PBS.
Then, 5 µl Hoechst staining solution was added, followed by 5 µl PI
stain. The cells were mixed and incubated in an ice bath at 4˚C for
20-30 min. The cells were carefully washed three times with PBS and
an anti-fluorescence quenching agent (Beyotime Institute of
Biotechnology) was added. Cells were observed under a fluorescence
microscope at x200 magnification and images captured. To calculate
the percentage of PI-positive cells, the number of PI-positive
cells was divided by the number of Hoechst-positive cells. Three
random fields were chosen for this calculation.
HE staining
The artery tissue from mice were fixed in 4%
paraformaldehyde at 4˚C and processed for paraffin or optimal
cutting temperature embedding. Embedded tissues were sliced to 8
µM, then HE staining were performed following the manufacturer's
instructions (cat. no. C0105S; Beyotime Institute of
Biotechnology). Eosin was used for 5 min at room temperature and
hematoxylin for 5 min at room temperature.
Statistical analysis
All experiments were independently repeated more
than three times. All statistical analyses were performed using the
GraphPad Prism 8.0 software (Dotmatics) and were presented as the
mean ± standard deviations (SDs). Statistical differences among
groups were determined using unpaired Student's t-test or by
one-way ANOVA followed by Tukey's post hoc analysis. Each
experiment was repeated at least three times, and differences with
P<0.05 were considered statistically significant.
Results
IL-37 improves the decreased cell
viability in macrophages induced by HG/ox-LDL
To determine the working concentration of IL-37,
THP-1 derived macrophages were treated using 5, 10, 20, 30 and 40
µM of IL-37. Cell viability was assayed after 24 h. It was found
that IL-37 at <30 µM did not exhibit significant cytostatic
effects on macrophages (Fig. 1A).
To mimic the HG and high fat environment in diabetic mice, HG (25
mM) and ox-LDL (100 ng/ml) were combined to stimulate macrophages.
CCK-8 assay demonstrated that IL-37 treatment significantly
improved that decrease of cell viability in macrophages induced by
HG/ox-LDL (Fig. 1B). Oxidative
damage to cell membranes is considered to be a key feature of
ferroptosis (18). LDH assay and
Hoechst-PI staining were used to detect the function of the cell
membrane. A significant increase was found in PI-positive cells in
the HG/ox-LDL group and a significant increase in LDH levels in the
supernatant. However, these changes were attenuated by IL-37
treatment (Fig. 1C-E).
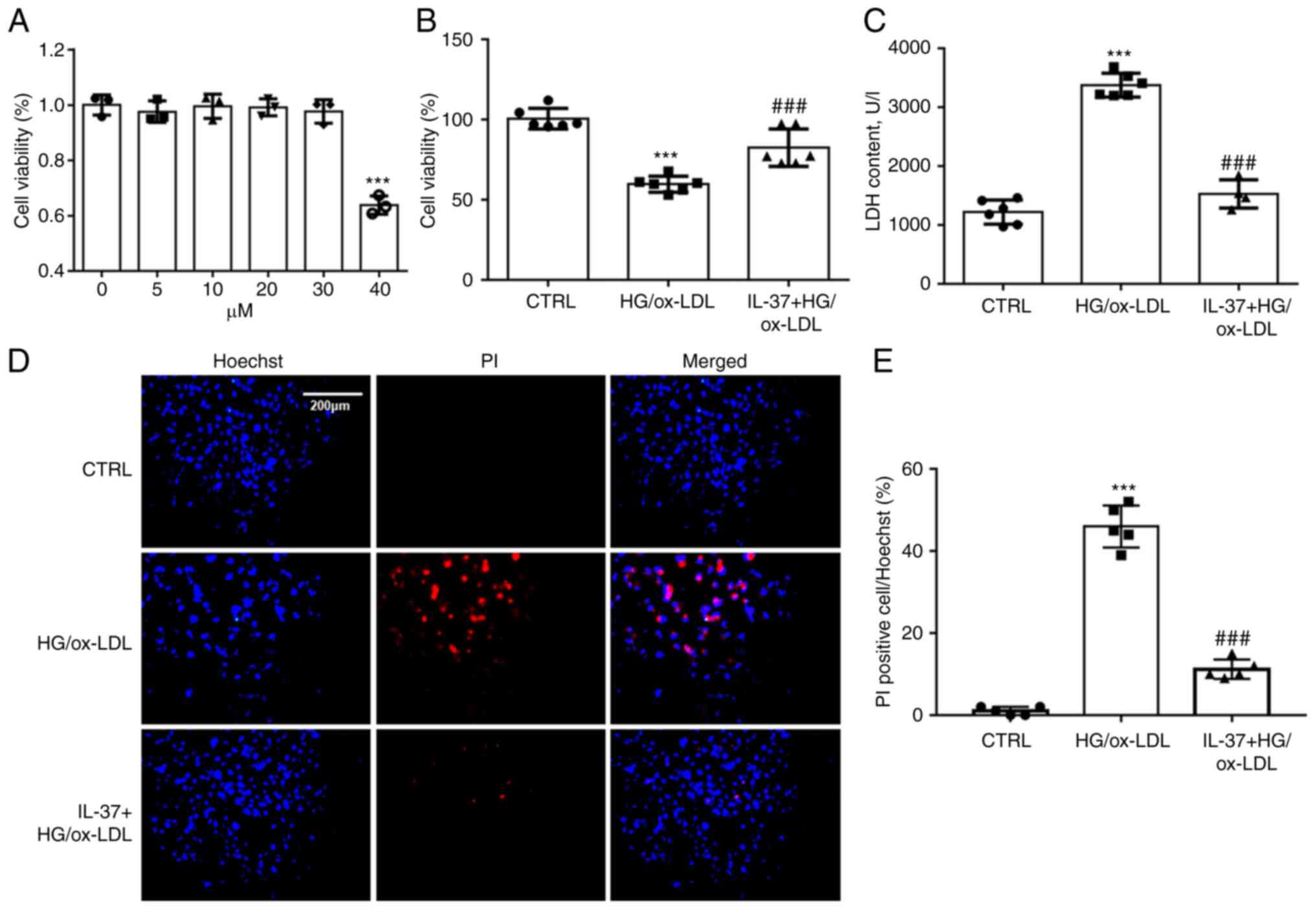 | Figure 1IL-37 ameliorates the decrease in
cell viability caused by HG/ox-LDL. (A) CCK-8 assay, the
macrophages were treated with 0, 5, 10, 20, 30 and 40 µM IL-37 for
24 h. The macrophages were divided into three groups: i) CTRL
group; ii) HG/ox-LDL group: The macrophages were treated with 100
ng/ml ox-LDL and 25 mM glucose for 24 h; iii) IL-37 group: 30 µM
IL-37 for 0.5 h, 100 ng/ml ox-LDL, 25 mM glucose for 24 h. (B)
CCK-8 assay. (C) LDH assay. (D and E) Hoechst-PI staining.
***P<0.001 vs. control group and
###P<0.001 vs. HG/ox-LDL. Each dot represents a
biological repetition. HG, high glucose; CCK-8, Cell Counting
Kit-8; LDH, lactate dehydrogenase. |
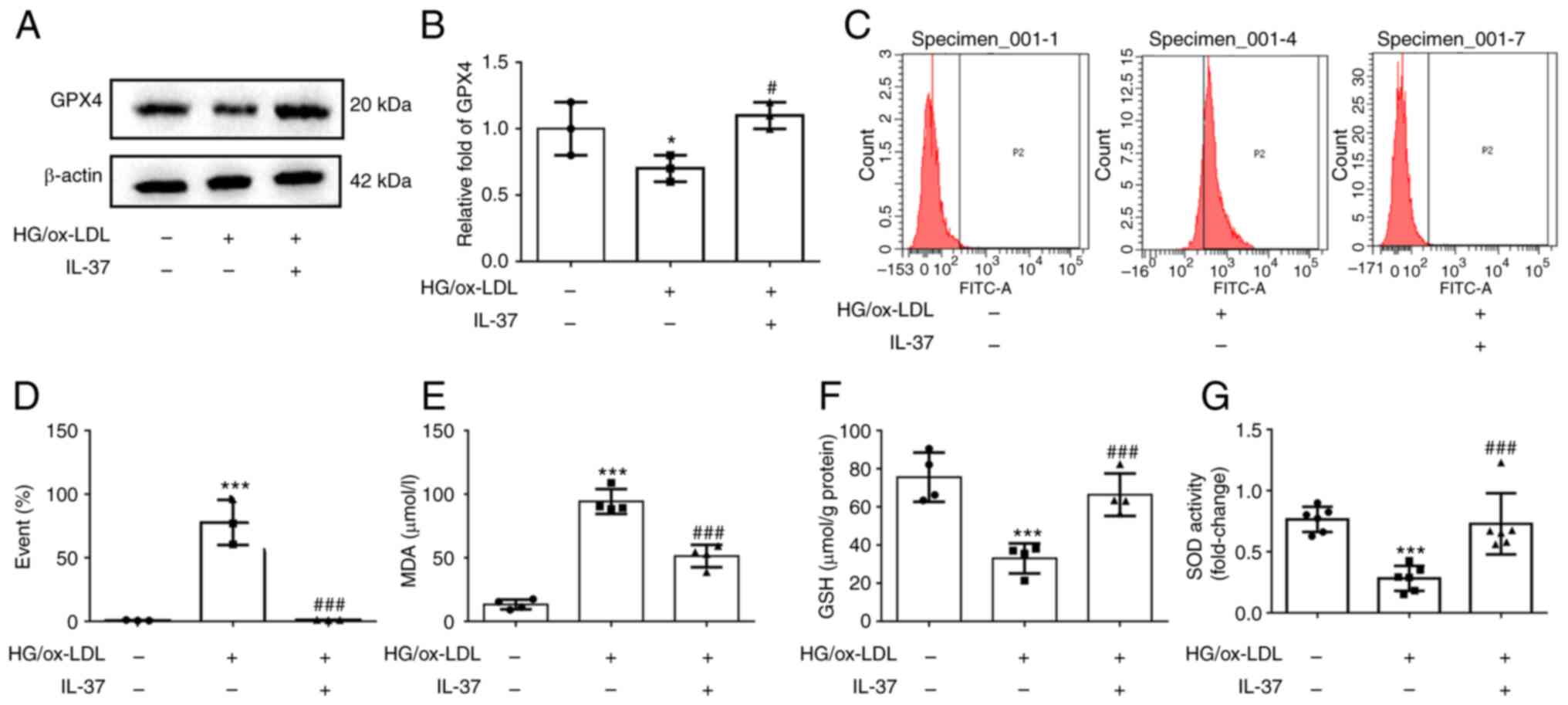
IL-37 suppresses macrophage
ferroptosis by improving oxidative stress
Western blot analysis was used to evaluate the
expression of GPX4, the marker of ferroptosis. The results
demonstrated that GPX4 was significantly decreased in HG/ox-LDL
group, while IL-37 treatment significantly increased the GPX4 level
(Fig. 2A and B). Increased cell membrane phospholipid
oxidation and MDA levels are key events in ferroptosis (18). C11 staining was next used to
evaluate the oxidation of macrophage cell membranes, and it was
found that IL-37 almost reversed the cell membrane oxidation caused
by HG/ox-LD treatment (Fig. 2C and
D). Moreover, IL-37 also decreased
the elevated MDA level in the HG/ox-LDL group (Fig. 2E). The GSH level and SOD activity
in macrophages were also determined. These results demonstrated
that IL-37 treatment significantly increased the GSH level and SOD
activity in the HG/ox-LDL group (Fig.
2F and G). These results
revealed that IL-37 suppressed macrophage ferroptosis by improving
oxidative stress. Furthermore, it was also found that IL-37
inhibited HG/ox-LDL-induced ferroptosis in mouse bone
marrow-derived macrophages (Fig.
S1).
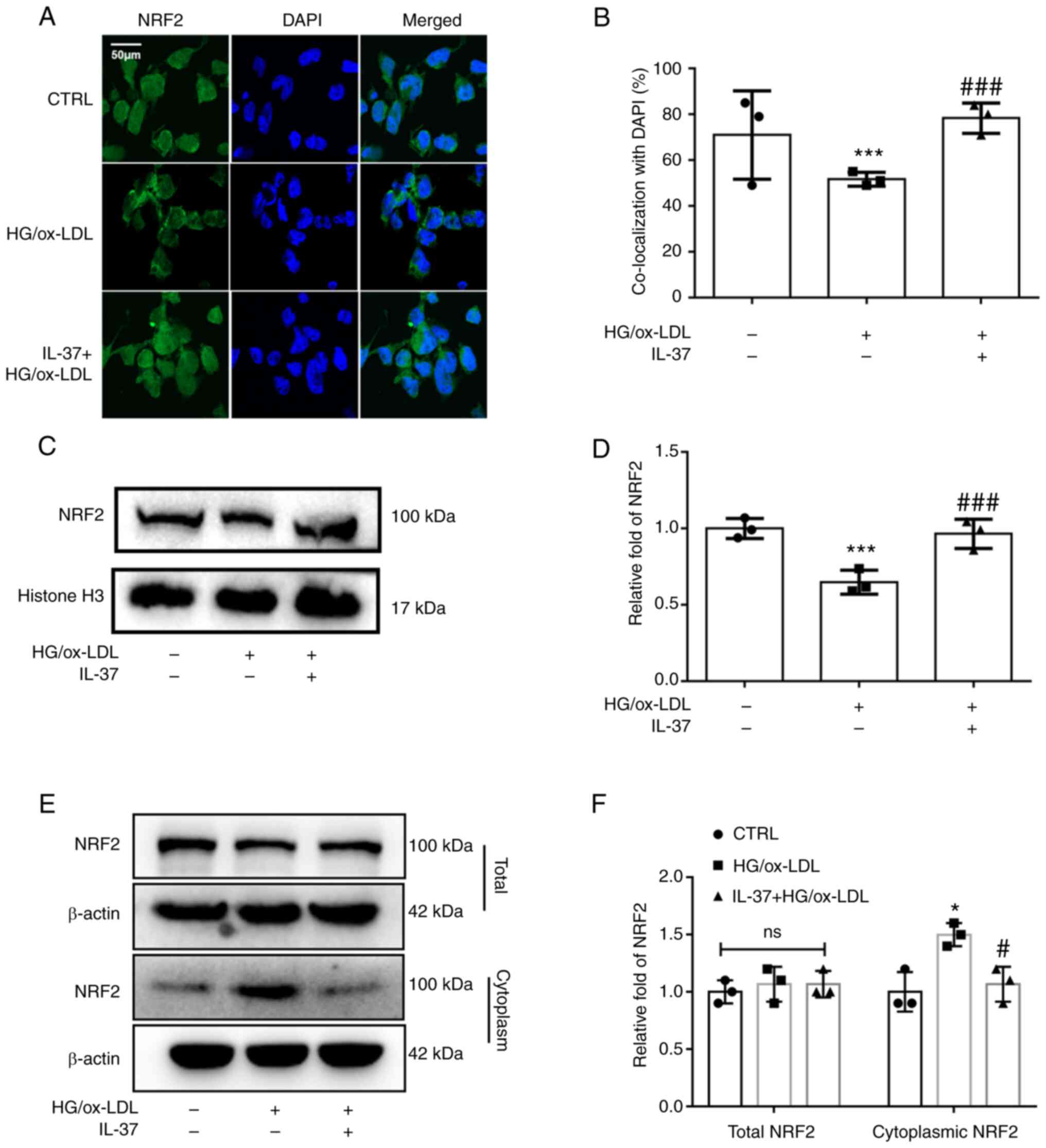
IL-37 promotes NRF2 activation via
enhancing NRF2 nuclear translocation
NRF2 acts as one of the most common cytosolic
transcription factors regulating oxidative stress metabolism and
inflammation. Under physiological conditions, NRF2 is located in
the cell nucleus. It is mainly responsible for regulating the
transcription of antioxidant proteins (18). From the immunofluorescence
staining, it was observed that HG/ox-LDL significantly inhibited
NRF2 nuclear translocation, while IL-37 attenuated this change
(Fig. 3A and B). Furthermore, cellular nuclear proteins
were extracted and western blot analysis was used to detect NRF2
levels. It was similarly found that NRF2 levels in the nucleus were
significantly decreased in the HG/ox-LDL group and significantly
increased after IL-37 treatment (Fig.
3C and D). In addition, it was
revealed that IL-37 did not affect the total NRF2 content in
macrophages, but reduced the level of NRF2 in the cytoplasm of
macrophages compared with the HG/ox-LDL group (Fig. 3E and F). These results suggested that IL-37
promotes NRF2 transfer from the cytoplasm to the nucleus.
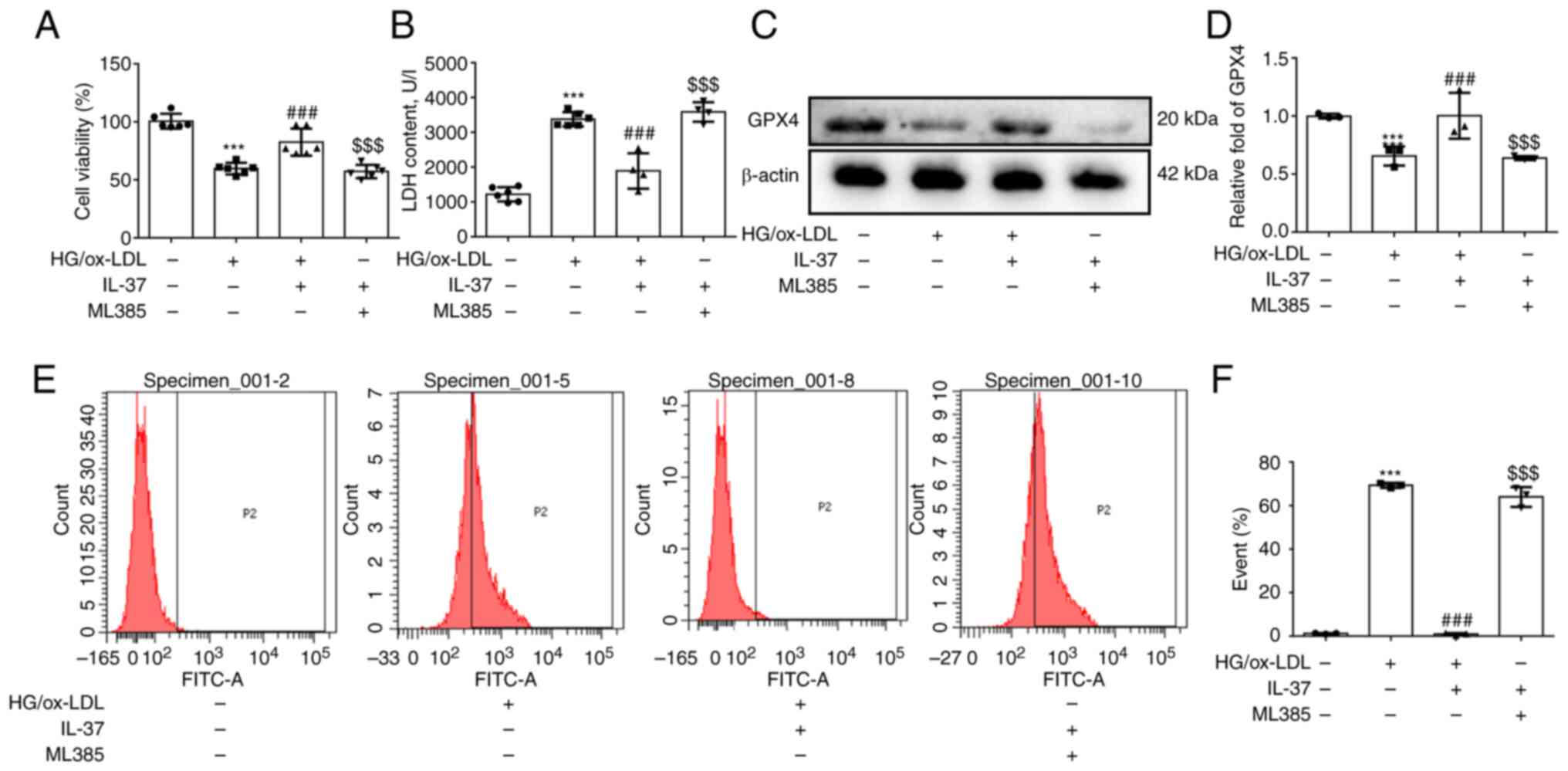
IL-37 inhibits ferroptosis of
macrophages by activating the NRF2 pathway
To further confirm if IL-37 inhibits ferroptosis of
macrophages caused by HG/ox-LDL by promoting NRF2 activation, ML385
(a specific inhibitor of NRF2) was used to pretreat macrophages.
Firstly, it was found that ML385 attenuated the protective effect
of IL-37 against macrophage viability (Fig. 4A). Secondly, after ML385
pretreatment, the LDH level in IL-37 group was also significantly
increased (Fig. 4B). Thirdly, it
was demonstrated that ML385 significantly decreased the GPX4 level
(Fig. 4C and D). Finally, the inhibitory effect of
IL-37 on cell membrane phospholipid oxidation was completely
abolished by ML385 (Fig. 4E and
F). Furthermore, the promotion of
NRF2 nuclear translocation by IL-37 was significantly reversed by
ML385 (Fig. S2). Overall, these
results strongly confirmed that IL-37 inhibits ferroptosis of
macrophage macrophages through activation of the NRF2 pathway.
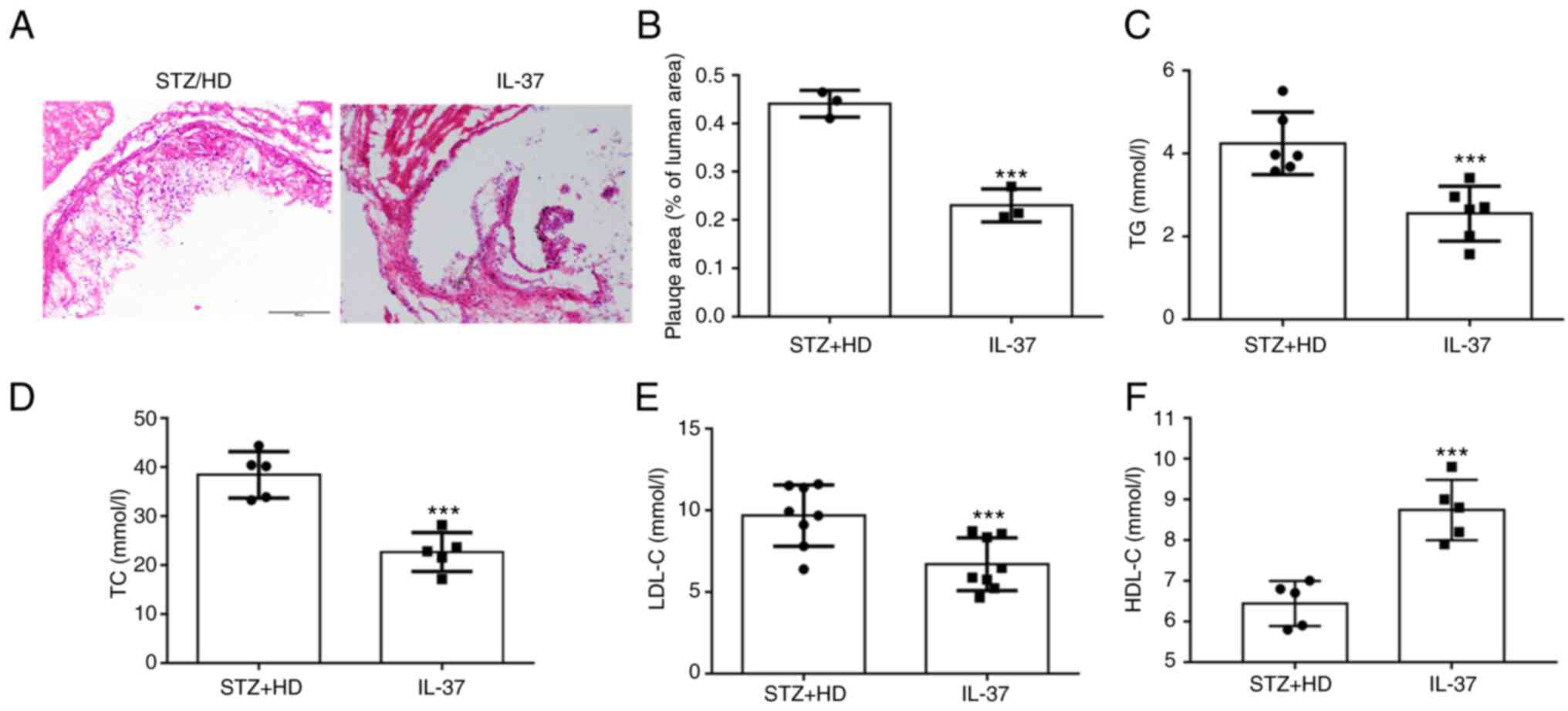
IL-37 attenuates atherosclerosis
progression in diabetic ApoE-/- mice
In the current study, STZ was used to construct a
model of diabetes. These mice were administered high-fat feeding
for 12 weeks. For H&E staining at the aortic sinus, it was
observed that IL-37 treatment significantly reduced the plaque area
(Fig. 5A and B). Moreover, lipid levels were evaluated
in mice and it was found that IL-37 treatment significantly reduced
triglycerides, total cholesterol and LDL-C levels (Fig. 5C-E). In addition, IL-37 increased
the level of HDL-C in HD/STZ group (Fig. 5D). These data indicated that IL-37
effectively inhibited the progression of atherosclerosis and
improved blood lipid levels in diabetic ApoE-/-
mice.
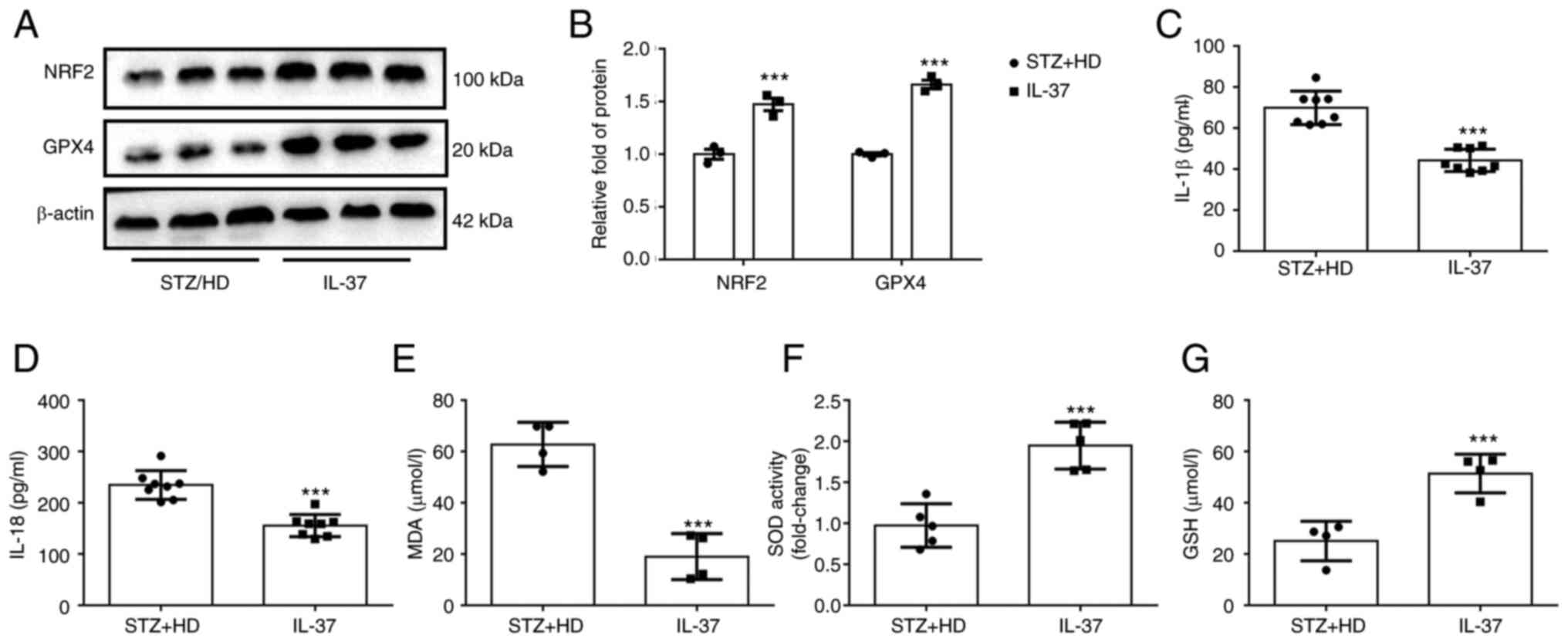
IL-37 inhibits ferroptosis of
macrophages, oxidative stress and inflammatory response in diabetic
ApoE-/- mice
To further confirm the protective effect of IL-37
against ferroptosis in vivo, lysate was extracted from mouse
aorta and the levels of NRF2 and GPX4 were detected using western
blot analysis. The results revealed that the protein levels of GPX4
and NRF2 were significantly increased in IL-37-treated mice
(Fig. 6A and B). Furthermore, the IL-18 and IL-1β
levels in serum were detected and it was found that IL-37 treatment
significantly decreased the IL-18 and IL-1β levels in diabetic
ApoE-/- mice (Fig. 6C and D). Lastly, the serum levels of MDA, GSH
and SOD in mice were evaluated. The data demonstrated that IL-37
treatment increased the GSH level, enhanced SOD activity and
decreased the MDA level in diabetic ApoE-/- mice
(Fig. 6E-G). These in vivo
results confirmed that IL-37 has the ability to inhibit ferroptosis
and inflammation and improve oxidative stress.
Discussion
The main findings of the present study are as
follows: i) IL-37 inhibited ferroptosis in macrophages caused by
HG/ox-LDL; ii) IL-37 inhibits macrophage ferroptosis by activating
the NRF2 pathway; and iii) IL-37 attenuated diabetic
atherosclerosis by inhibiting macrophage ferroptosis. The present
results suggested that IL-37 may serve as a new potential
intervention target for atherosclerosis therapy.
Ferroptosis is a newly discovered mode of programmed
cell death. In recent years, studies from autopsies and animal
experiments have reported that ferroptosis is extensively involved
in the development and progression of atherosclerosis (25-27).
Evidence from clinical researches suggested that HMOX1, a key gene
of ferroptosis, is strongly associated with the prognosis of
patients with atherosclerosis (28). Clinical data also suggested that
genes for ferroptosis can be used as biomarkers of atherosclerosis
(29). It was reported that
erythroid lineage Jak2V617F expression contributes to the
progression of atherosclerosis by enhancing macrophage ferroptosis
(30). Moreover, it was also found
that inhibition of ferroptosis can effectively improve
atherosclerosis. Yang et al (31) reported that prenyl-diphosphate
synthase subunits suppressed the of endothelial cells ferroptosis
in atherosclerosis by activating the NRF2 pathway. A previous study
(32) showed that Fer-1 inhibits
atherosclerosis progression in ApoE-/- mice by
suppressing ferroptosis in endothelial cells. These results all
suggested that either macrophage or endothelial cell ferroptosis is
involved in the development of atherosclerosis; thus ferroptosis
may be a potential target for intervention in atherosclerosis. In
the present study, both in vivo and in vitro evidence
found that IL-37 inhibits ferroptosis in macrophages caused by
HG/ox-LDL. It was also revealed that IL-37 could improve oxidative
stress caused by HG/ox-LDL. Previous studies also demonstrated that
IL-37 reduced HG-induced inflammation and oxidative stress. It was
previously reported (33) that
IL-37 could ameliorate oxidative damage in epithelial cells in
diabetic kidney injury via the STAT3 pathway. Furthermore, it was
found that the level of IL-37 was negatively correlated with
oxidative stress in patients with Hashimoto's thyroiditis, which
also suggests that IL-37 has an inhibitory effect on oxidative
stress (34). The results of the
present study found that IL-37 also has a broad antioxidant
capacity in atherosclerosis, and in addition, it was further
identified that IL-37 may attenuate ferroptosis of macrophages by
ameliorating oxidative stress. In addition, it was found that IL-37
improved diabetic atherosclerosis, and IL-37 improved macrophage
ferroptosis in both in vivo and in vitro experiments.
However, this does not directly indicate that IL-37 improves
diabetic atherosclerosis by inhibiting ferroptosis in macrophages.
Previous studies have identified the role of macrophage iron death
in promoting atherosclerosis (18,30,35).
Furthermore, it was reported that the ferroptosis inhibitor, Fer-1,
can significantly improve atherosclerosis (32). These evidence directly confirm that
macrophage ferroptosis promotes atherosclerosis, which also
supports the present conclusion to certain extent. On the other
hand, IL-37 may improve atherosclerosis by inhibiting macrophages
in other ways, such as inhibiting apoptosis of macrophages and
promoting autophagy. Further experiments are needed to confirm
this.
As a newly discovered inflammatory factor, the
anti-inflammatory effect of IL-37 is gradually being investigated.
Several studies demonstrated that IL-37 has potential protective
effects against inflammatory diseases, including colitis and
gastritis (36-38).
Furthermore, it was also reported that IL-37 improved the function
of plasma dendritic cells in patients with acute coronary syndrome
(11). The inflammatory response
is one of the key mechanisms of diabetic atherosclerosis. However,
no previous studies have focused on the role of IL-37 on the level
of inflammation in diabetic atherosclerotic mice. The current data
revealed that the levels of classical pro-inflammatory factors,
IL-1β and IL-18, in peripheral blood were significantly decreased
in diabetic atherosclerotic mice after continuous intraperitoneal
injection of IL-37. The results of the present study confirmed the
potent anti-inflammatory ability of IL-37 in atherosclerosis, which
is consistent with the phenomenon found in clinical patients. Thus,
anti-inflammation may be one of the potential mechanisms by which
IL-37 exerts athero-protective effects. However, the mechanism of
IL-37 inhibition of inflammation in HG/ox-LDL-treated macrophages
was not further investigated. This is one of the limitations to the
current study. Previous studies have confirmed that IL-37 is a
potential inhibitor of P65 and may exert anti-inflammatory effects
by inhibiting the phosphorylation and nuclear translocation of P65
(39-41).
P65 activation has also been suggested as a key mechanism for
increased inflammation levels in diabetic atherosclerotic mice
(42,43). Therefore, further studies are
needed to focus on the mechanisms underlying the role of IL-37 on
P65 activation in diabetic atherosclerotic mice.
NRF2 is a classical transcription factor. NRF2
regulates proteins of inflammation and oxidative stress and has
received a lot of attention for this reason. Under physiological
conditions, NRF2 is localized in the nucleus and is responsible for
binding the promoter of the antioxidant protein progenitor to
maintain oxidative stress homeostasis in the cell. In the present
study, it was found that macrophages treated with HG/ox-LDL showed
a significant decrease in NRF2 levels in the nucleus and a
significant downregulation of GPX4, GSH and SOD levels, compared
with the normal group. IL-37 treatment not only increased the
expression of NRF2 in the nucleus, but also upregulated the levels
of GPX4, GSH and SOD. However, ML383 abolished these changes. These
results provide strong evidence that IL-37 inhibits macrophage
ferroptosis by activating the NRF2 pathway. As a classical
transcription factor, a large number of studies have reported that
the transfer of NRF2 to the nucleus is the key to the biological
function of NRF2 (18,28,44).
It was reported that lycopene ameliorates DEHP exposure-induced
renal pyroptosis through enhancing NRF2 nuclear translocation
(45). In the present study, the
content of NRF2 was evaluated in the nucleus of ML385 treatment,
and it was found that the nuclear translocation of NRF2 caused by
IL-37 treatment was significantly inhibited by ML385. Furthermore,
ML385 inhibits the activity of NRF2, which is activated by IL-37.
Therefore, it was hypothesized that IL-37 may activate NRF2 pathway
by promoting NRF2.
It was reported that NRF2 plays a critical role in
mitigating lipid peroxidation and ferroptosis (46). It was found that exogenous
activation of NRF2 inhibits acute lung injury due to intestinal
ischemia via SLC7A11/HO-1(47).
Furthermore, inhibition of NRF2 has been reported to exacerbate
acute myocardial injury caused by doxorubicin (48). These results are consistent with
the findings of the present study. Moreover, it was observed that
IL-37 increased NRF2 expression of IL-37 in diabetic mice, but
in vitro experiment found that IL-37 had no effect on total
NRF2 level in macrophages. This has been previously reported, as
Luo et al (49)
demonstrated that after ox-LDL treatment of macrophages, the total
protein level of NRF2 did not change and NRF2 was reduced in the
nucleus, and quercetin treatment could inhibit this phenomenon.
However, in high-fat fed ApoE-/- mice, total vascular
NRF2 decreased and quercetin increased total vascular NRF2
expression. At the cellular level, there may be feedback regulation
in macrophages treated with HG/ox-LDL, resulting in unchanged NRF2
level and only decreased NRF2 level in the nucleus. Then, in
ApoE-/- mice, atherosclerotic plaques had occurred, and
the feedback regulatory effect of macrophages was weakened or lost,
which led to a decrease in NRF2 expression, but IL-37 could
ameliorate this phenomenon.
There are certain limitations to the present study.
It was only found that IL-37 improved diabetic atherosclerosis and
that IL-37 improved macrophage ferroptosis. Nevertheless, this does
not directly indicate that IL-37 improves diabetic atherosclerosis
by inhibiting ferroptosis in macrophages. Macrophage-deletion mice
are needed to further confirm that IL-37 inhibits ferroptosis in
macrophages to protect atherosclerosis. Secondly, at the animal
level, NrF2-deficient knockout mice confirmed that IL-37 improves
ferroptosis in macrophages by activating NRF2.
In conclusion, it was identified that IL-37
suppresses ferroptosis of macrophages to attenuate atherosclerosis
via activating the NRF2 pathway.
Supplementary Material
IL-37 inhibits mouse bone marrow
derived macrophages ferroptosis. The mouse bone marrow derived
macrophages were divided into three groups: i) CTRL group; ii)
HG/ox-LDL group: The macrophages were treated with 100 ng/ml ox-LDL
and 25 mM glucose for 24 h; and iii) IL-37 group: 30 μM
IL-37 for 0.5 h, 100 ng/ml ox-LDL, 25 mM glucose for 24 h. (A) Cell
Counting Kit-8 assay. (B and C) Western blotting was used to
analyze the protein level of GPX4. (D) GSH level in different
groups. (E) MDA level in different groups. (F) SOD level in
different group. *P<0.05 vs. control group and
#P<0.05 vs. HG/ox-LDL. Each dot represents a
biological repetition. HG, high glucose; GPX4, glutathione
peroxidase 4; GSH, reduced glutathione; MDA, malondialdehyde; SOD,
superoxide dismutase.
ML385 inhibits NRF2 nuclear
translocation induced by IL-37. The mouse bone marrow-derived
macrophages were divided into four groups: i) CTRL group; ii)
HG/ox-LDL group: The macrophages were treated with 100 ng/ml ox-LDL
and 25 mM glucose for 24 h; iii) IL-37 group: 30 μM IL-37
for 0.5 h, 100 ng/ml ox-LDL, 25 mM glucose for 24 h and iv) ML385
group: 5 μM ML385 for 0.5 h, 30 μM IL-37 for 0.5 h,
100 ng/ml ox-LDL, 25 mM glucose for 24 h. (A and B) Macrophage
nuclear proteins were extracted and western blotting was used to
evaluate the level of NRF2 in the nucleus. *P<0.05
vs. control group, #P<0.05 vs. HG/ox-LDL and
$P<0.05 vs. IL-37/HG/ox-LDL. Each dot represents a
biological repetition. NRF2, nuclear factor erythroid 2-related
factor 2; HG, high glucose.
Acknowledgements
The authors would like to thank Dr Gang Wang (The
Second Affiliated Hospital of Harbin Medical University, Harbin,
China) for providing help and would also like to thank Dr Steve
Sunny (University of Bristol, Bristol, UK) for help in revising the
manuscript.
Funding
Funding: The present study was supported by the Natural Science
Foundation of Heilongjiang (grant no. LH2020H063).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZC developed the analysis plan and wrote the
manuscript. QZ, JX, XH and NX performed the main experiments and
data analysis. ZC and XJ confirm the authenticity of all the raw
data. All authors contributed to manuscript revision, and have read
and approved the final manuscript.
Ethics approval and consent to
participate
The present experiment was approved (approval no.
SYDW2020-251) by the Ethics Committee of the Fourth Affiliated
Hospital of Harbin Medical University (Harbin, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lopez-Jimenez F, Almahmeed W, Bays H,
Cuevas A, Di Angelantonio E, le Roux CW, Sattar N, Sun MC, Wittert
G, Pinto FJ and Wilding JPH: Obesity and cardiovascular disease:
mechanistic insights and management strategies. A joint position
paper by the world heart federation and world obesity federation.
Eur J Prev Cardiol. 29:2218–2237. 2022.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Boswell L, Serés-Noriega T, Mesa A, Perea
V, Pané A, Viñals C, Blanco J, Giménez M, Vinagre I, Esmatjes E, et
al: Carotid ultrasonography as a strategy to optimize
cardiovascular risk management in type 1 diabetes: A cohort study.
Acta Diabetol. 59:1563–1574. 2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Wang C, Liu C, Shi J, Li H, Jiang S, Zhao
P, Zhang M, Du G, Fu S, Li S, et al: Nicotine exacerbates
endothelial dysfunction and drives atherosclerosis via
extracellular vesicle-miRNA. Cardiovasc Res.
25(cvac140)2022.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Everett BM, MacFadyen JG, Thuren T, Libby
P, Glynn RJ and Ridker PM: Inhibition of interleukin-1β and
reduction in atherothrombotic cardiovascular events in the cantos
trial. J Am Coll Cardiol. 76:1660–1670. 2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ferencik M, Mayrhofer T, Lu MT, Bittner
DO, Emami H, Puchner SB, Meyersohn NM, Ivanov AV, Adami EC, Voora
D, et al: Coronary atherosclerosis, cardiac troponin, and
interleukin-6 in patients with chest pain: The PROMISE trial
results. JACC Cardiovasc Imaging. 15:1427–1438. 2022.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Andreotti F, Maggioni AP, Campeggi A,
Iervolino A, Scambia G and Massetti M: Anti-inflammatory therapy in
ischaemic heart disease: From canakinumab to colchicine. Eur Heart
J Supp. 23:E13–E18. 2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Liberale L, Kraler S, Puspitasari YM,
Bonetti NR, Akhmedov A, Ministrini S, Montecucco F, Marx N, Lehrke
M, Hartmann NUK, et al: SGLT-2 inhibition by empagliflozin has no
effect on experimental arterial thrombosis in a murine model of
low-grade inflammation. Cardiovasc Res. 22(cvac126)2022.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Li H, Shen C, Chen B, Du J, Peng B, Wang
W, Chi F, Dong X, Huang Z and Yang C: Interleukin-37 is increased
in peripheral blood mononuclear cells of coronary heart disease
patients and inhibits the inflammatory reaction. Mol Med Rep.
21:151–160. 2020.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Law CC, Puranik R, Fan J, Fei J, Hambly BD
and Bao S: Clinical implications of IL-32, IL-34 and IL-37 in
atherosclerosis: Speculative role in cardiovascular manifestations
of COVID-19. Front Cardiovasc Med. 8(630767)2021.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Zhang F, Zhu T, Li H, He Y, Zhang Y, Huang
N, Zhang G, Li Y, Chang D and Li X: Plasma Interleukin-37 is
elevated in acute ischemic stroke patients and probably associated
with 3-month functional prognosis. Clin Interv Aging. 15:1285–1294.
2020.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhu R, Zhang F, Pan C, Yu K, Zhong Y and
Zeng Q: Role of IL-37- and IL-37-treated dendritic cells in acute
coronary syndrome. Oxid Med Cell Long. 2021(6454177)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Rafiei A, Ahmadi R, Kazemian S,
Rahimzadeh-Fallah T, Mohammad-Rezaei M, Azadegan-Dehkordi F, Sanami
S, Mirzaei Y, Aghaei F and Bagheri N: Serum levels of IL-37 and
correlation with inflammatory cytokines and clinical outcomes in
patients with coronary artery disease. J Invest Med. 70:1720–1727.
2022.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Liu K, Tang Q, Zhu X and Yang X: IL-37
increased in patients with acute coronary syndrome and associated
with a worse clinical outcome after ST-segment elevation acute
myocardial infarction. Clin Chim Acta. 468:140–144. 2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu J, Lin J, He S, Wu C, Wang B, Liu J,
Duan Y, Liu T, Shan S, Yang K, et al: Transgenic overexpression of
IL-37 protects against atherosclerosis and strengthens plaque
stability. Cell Physiol Biochem. 45:1034–1050. 2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Ji Q, Meng K, Yu K, Huang S, Huang Y, Min
X, Zhong Y, Wu B, Liu Y, Nie S, et al: Exogenous interleukin 37
ameliorates atherosclerosis via inducing the Treg response in
ApoE-deficient mice. Sci Rep. 7(3310)2017.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Chai M, Ji Q, Zhang H, Zhou Y, Yang Q,
Zhou Y, Guo G, Liu W, Han W, Yang L, et al: The protective effect
of interleukin-37 on vascular calcification and atherosclerosis in
apolipoprotein e-deficient mice with diabetes. J Interferon
Cytokine Res. 35:530–539. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Lotfy H and Moaaz M and Moaaz M: The novel
role of IL-37 to enhance the anti-inflammatory response of
regulatory T cells in patients with peripheral atherosclerosis.
Vascular. 28:629–642. 2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yu W, Liu W, Xie D, Wang Q, Xu C, Zhao H,
Lv J, He F, Chen B, Yamamoto T, et al: High level of uric acid
promotes atherosclerosis by targeting NRF2-mediated autophagy
dysfunction and ferroptosis. Oxid Med Cell Longev.
2022(9304383)2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ouyang S, You J, Zhi C, Li P, Lin X, Tan
X, Ma W, Li L and Xie W: Ferroptosis: The potential value target in
atherosclerosis. Cell Death Dis. 12(782)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Hu H, Chen Y, Jing L, Zhai C and Shen L:
The link between ferroptosis and cardiovascular diseases: A novel
target for treatment. Front Cardiovasc Med.
8(710963)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yu Y, Yan Y, Niu F, Wang Y, Chen X, Su G,
Liu Y, Zhao X, Qian L, Liu P and Xiong Y: Ferroptosis: A cell death
connecting oxidative stress, inflammation and cardiovascular
diseases. Cell Death Disc. 7(193)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Duan JY, Lin X, Xu F, Shan SK, Guo B, Li
FXZ, Wang Y, Zheng MH, Xu QS, Lei LM, et al: Ferroptosis and its
potential role in metabolic diseases: A curse or revitalization?
Front Cell Dev Biol. 9(701788)2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhou Y, Zhou H, Hua L, Hou C, Jia Q, Chen
J, Zhang S, Wang Y, He S and Jia E: Verification of ferroptosis and
pyroptosis and identification of PTGS2 as the hub gene in human
coronary artery atherosclerosis. Free Radical Biol Med. 171:55–68.
2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang G, Han B, Zhang R, Liu Q, Wang X,
Huang X, Liu D, Qiao W, Yang M, Luo X, et al: C1q/TNF-related
protein 9 attenuates atherosclerosis by inhibiting
hyperglycemia-induced endothelial cell senescence through the
AMPKα/KLF4 signaling pathway. Front Pharmacol.
12(758792)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Cui Z and Zhao X, Amevor FK, Du X, Wang Y,
Li D, Shu G, Tian Y and Zhao X: Therapeutic application of
quercetin in aging-related diseases: SIRT1 as a potential
mechanism. Front Immunol. 13(943321)2022.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Meng Q, Xu Y, Ling X, Liu H, Ding S, Wu H,
Yan D, Fang X, Li T and Liu Q: Role of ferroptosis-related genes in
coronary atherosclerosis and identification of key genes:
Integration of bioinformatics analysis and experimental validation.
BMC Cardiovasc Dis. 22(339)2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Guo Y, Zhang W, Zhou X, Zhao S, Wang J,
Guo Y, Liao Y, Lu H, Liu J, Cai Y, et al: Roles of ferroptosis in
cardiovascular diseases. Front Cardiovasc Med.
9(911564)2022.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wu D, Hu Q, Wang Y, Jin M, Tao Z and Wan
J: Identification of HMOX1 as a Critical Ferroptosis-Related Gene
in Atherosclerosis. Front Cardiovasc Med. 9(833642)2022.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Liu H, Xiang C, Wang Z and Song Y:
Identification of potential ferroptosis-related biomarkers and
immune infiltration in human coronary artery atherosclerosis. Int J
Gen Med. 15:2979–2990. 2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Liu W, Östberg N, Yalcinkaya M, Dou H,
Endo-Umeda K, Tang Y, Hou X, Xiao T, Fidler T, Abramowicz S, et al:
Erythroid lineage Jak2V617F expression promotes atherosclerosis
through erythrophagocytosis and macrophage ferroptosis. J Clin
Invest. 132(e155724)2022.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Yang K, Song H and Yin D: PDSS2 inhibits
the ferroptosis of vascular endothelial cells in atherosclerosis by
activating Nrf2. J Cardiovasc Pharmacol. 77:767–776.
2021.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Bai T, Li M, Liu Y, Qiao Z and Wang Z:
Inhibition of ferroptosis alleviates atherosclerosis through
attenuating lipid peroxidation and endothelial dysfunction in mouse
aortic endothelial cell. Free Radic Biol Med. 160:92–102.
2020.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Zhang X, Zhu Y, Zhou Y and Fei B:
Interleukin 37 (IL-37) reduces high glucose-induced inflammation,
oxidative stress, and apoptosis of podocytes by inhibiting the
STAT3-Cyclophilin A (CypA) signaling pathway. Med Sci Monit.
26(e922979)2020.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ruggeri RM, Cristani M, Vicchio TM,
Alibrandi A, Giovinazzo S, Saija A, Campennì A, Trimarchi F and
Gangemi S: Increased serum interleukin-37 (IL-37) levels correlate
with oxidative stress parameters in Hashimoto's thyroiditis. J
Endocrinol Invest. 42:199–205. 2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen Y, Luo X, Xu B, Bao X, Jia H and Yu
B: Oxidative stress-mediated programmed cell death: A potential
therapy target for atherosclerosis. Cardiovasc Drugs Ther 16:
10.1007/s10557-022-07414-z, 2022.
|
|
36
|
Ahmadnia Z, Ranaee M, Abandansari RM,
Bagheri N and Shirzad H: Evaluating the MicroRNA expression of
IL-35 and IL-37 in helicobacter pylori-infected patients with
gastritis and gastric ulcer. Iran J Allergy Asthma Immunol.
21:20–26. 2022.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Qin H, Sun C, Zhu Y, Qin Y, Ren S, Wang Z,
Li C, Li X, Zhang B, Hao J, et al: IL-37 overexpression promotes
endometrial regenerative cell-mediated inhibition of cardiac
allograft rejection. Stem Cell Res Ther. 13(302)2022.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Cong J, Wu D, Dai H, Ma Y, Liao C, Li L,
Ye L and Huang Z: Interleukin-37 exacerbates experimental colitis
in an intestinal microbiome-dependent fashion. Theranostics.
12:5204–5219. 2022.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ding Y, Wang Y, Cai Y, Pan C, Yang C, Wang
M, Qi X, Ye J, Ji Q, Yu J, et al: IL-37 expression in patients with
abdominal aortic aneurysm and its role in the necroptosis of
vascular smooth muscle cells. Oxid Med Cell Longev.
11(1806513)2022.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Liu T, Liu J, Lin Y, Que B, Chang C, Zhang
J, Liang Z, Gao X, Liu S, Liu L, et al: IL-37 inhibits the
maturation of dendritic cells through the IL-1R8-TLR4-NF-κB
pathway. Biochim Biophys Acta Mol Cell Biol Lipids. 1864:1338–1349.
2019.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Huang N, Liu K, Liu J, Gao X, Zeng Z,
Zhang Y and Chen J: Interleukin-37 alleviates airway inflammation
and remodeling in asthma via inhibiting the activation of NF-κB and
STAT3 signalings. Int Immunopharmacol. 55:198–204. 2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Xie M, Tang Q, Nie J, Zhang C, Zhou X, Yu
S, Sun J, Cheng X, Dong N, Hu Y and Chen L: BMAL1-downregulation
aggravates porphyromonas gingivalis-induced atherosclerosis by
encouraging oxidative stress. Circ Res. 126:e15–e29.
2020.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Wang Z, Liu B, Zhu J, Wang D and Wang Y:
Nicotine-mediated autophagy of vascular smooth muscle cell
accelerates atherosclerosis via nAChRs/ROS/NF-κB signaling pathway.
Atherosclerosis. 284:1–10. 2019.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Wei Z, Jing Z, Pinfang K, Chao S and
Shaohuan Q: Quercetin inhibits pyroptosis in diabetic
cardiomyopathy through the Nrf2 pathway. J Diabetes Res.
31(9723632)2022.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Li MZ, Zhao Y, Dai XY, Talukder M and Li
JL: Lycopene ameliorates DEHP exposure-induced renal pyroptosis
through the Nrf2/Keap-1/NLRP3/Caspase-1 axis. J Nutr Biochem.
113(109266)2023.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Dodson M, Castro-Portuguez R and Zhang DD:
NRF2 plays a critical role in mitigating lipid peroxidation and
ferroptosis. Redox Biol. 23(101107)2019.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Dong H, Qiang Z, Chai D, Peng J, Xia Y, Hu
R and Jiang H: Nrf2 inhibits ferroptosis and protects against acute
lung injury due to intestinal ischemia reperfusion via regulating
SLC7A11 and HO-1. Aging. 12:12943–12959. 2020.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Wang Y, Yan S, Liu X, Deng F, Wang P, Yang
L, Hu L, Huang K and He J: PRMT4 promotes ferroptosis to aggravate
doxorubicin-induced cardiomyopathy via inhibition of the Nrf2/GPX4
pathway. Cell Death Differ. 29:1982–1995. 2022.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Luo X, Weng X, Bao X, Bai X, Lv Y, Zhang
S, Chen Y, Zhao C, Zeng M, Huang J, et al: A novel
anti-atherosclerotic mechanism of quercetin: Competitive binding to
KEAP1 via Arg483 to inhibit macrophage pyroptosis. Redox Biol.
57(102511)2022.PubMed/NCBI View Article : Google Scholar
|