Introduction
Preeclampsia (PE) is a pregnancy-associated
disorder, characterized by elevated maternal blood pressure and
proteinuria after 20 weeks of gestation (1). To date, it remains the major cause of
fetal and maternal mortality and morbidity worldwide (2). Although considerable efforts in
recent years have been made in the understanding of PE, little
improvement in disease diagnosis and treatment for PE remains
unsatisfactory (3,4). Although previous studies indicate
that trophoblastic dysfunction, including insufficient trophoblast
migration and invasion, is critical for the pathogenesis of PE, the
underlying regulatory mechanisms of trophoblast invasion remain
unclear (5,6).
MicroRNAs (miRs) are a group of non-coding small
RNAs (21-23 nucleotides), which can regulate gene expression
through binding with target mRNA (7). Several reports have demonstrated that
miRs play the key roles in the regulation of trophoblastic invasion
(8,9). For example, Liu et al
(10) showed that the miR-142-3p
expression levels is upregulated in placenta specimens obtained
from patients with PE and its upregulation inhibited trophoblast
cell invasion and migration through disruption of the TGF-β/SMAD3
signaling pathway. Gao et al (11) found that miR-299 suppresses the
invasion and migration of trophoblast cells partly via targeting
histone deacetylase 2. Notably, a recent study showed that low
expression of miR-424 in placenta is associated with severe PE
(12). Another report also found
that miR-424 is downregulated in primary human trophoblasts by
hypoxia and associated with hindered trophoblast differentiation
(13). However, these studies did
not investigate the effects and underlying regulatory mechanisms of
miR-424 on trophoblastic invasion. In addition, miR-424 has been
reported to serve as an oncogene in several types of human cancer
and is demonstrated to contribute to the migration and invasion of
tumor cells (14). miR-424
expression levels were found to be increased in laryngeal squamous
cell carcinoma (LSCC) tissues and its upregulation promoted
migration and invasion of LSCC cells (15). Wu et al (16) demonstrated that miR-424 increases
the migration and invasion in pancreatic cancer cells. Based on the
evidence from these previous studies, it is reasonable that miR-424
may affect trophoblastic invasion.
In the present study, the differentially expressed
miRs that were screened based on GSE96985 microarray data retrieved
from Gene Expression Omnibus (GEO) and miR-424 was selected for
further analysis. The regulatory mechanisms and effects of miR-424
on the invasion and migration of HTR-8/SVneo cells were further
investigated via in vitro experiments. The present findings
suggested that miR-424 may be a potential therapeutic target
against PE.
Materials and methods
Clinical samples
The placenta samples were collected from 60 pregnant
women undergoing cesarean section (30 patients with PE and 30
healthy pregnant women with uncomplicated pregnancies) in the
Department of Gynecology and Obstetrics, the Obstetrics &
Gynecology Hospital of Fudan University. Patients' main clinical
parameters are reported and summarized in Table I. Hypertension (≥140/90 mmHg) and
proteinuria (≥300 mg protein/24 h-urine sample) occurring after
20-week gestation were the necessary inclusion criteria for
patients with PE. Patients with cardiovascular diseases, diabetes,
metabolic syndrome, infections, kidney disease, congenital
malformations and chromosomal anomalies (number and/or structure)
were excluded. All samples were immediately frozen in liquid
nitrogen and stored at -80˚C until further use. The present study
was approved by the Research Ethics Committee of the Obstetrics
& Gynecology Hospital of Fudan University (approval no.
2018-013). Written informed consent was obtained from each
participant.
 | Table IClinical characteristics of patients
with preeclampsia and normal pregnant women. |
Table I
Clinical characteristics of patients
with preeclampsia and normal pregnant women.
|
Characteristics | Control, n=30 | GDM, n=30 | P-value |
|---|
| Age, years | 30.37±5.45 | 31.02±5.83 | 0.657 |
| MAP, mm Hg | 91.62±8.34 | 134.21±10.47 | <0.0001 |
| Urinary protein,
g/24 h | - | 2.4±0.2 | <0.0001 |
| Uric acid.
µmol/l | 251.46±34.63 | 396.76±39.03 | <0.0001 |
| Pregnancy,
weeks | 38.34±1.73 | 37.82±1.14 | 0.175 |
| Birth weight,
kg | 3.41±0.37 | 2.83±0.43 | <0.0001 |
| BMI,
kg/m2 | 23.12±1.43 | 22.54±1.66 | 0.153 |
Cell culture
Human trophoblast cell line HTR-8/SVneo was obtained
from Cell Culture Center of the Shanghai Institute (Shanghai,
China), which was shown to contain a heterogeneous population of
both trophoblast and mesenchymal cells (17,18).
Cells were maintained in RPMI-1640 medium supplemented with 10%
fetal bovine serum (FBS; Invitrogen; Thermo Fisher Scientific,
Inc.) and 1% penicillin/streptomycin (Sigma-Aldrich; Merck KGaA) in
a humidified incubator with 5% CO2 at 37˚C.
Immunofluorescent staining
Prior to immunofluorescent staining, the cells were
fixed with 4% paraformaldehyde for 15 min at 4˚C, followed by
incubation with 5% FBS containing 0.5% Triton X-100 for 5 min and
finally blocked with 2% bovine serum albumin (BSA; Sigma-Aldrich;
Merck KGaA) in PBS at 37˚C for 60 min. Subsequently, the cells were
incubated at 4˚C overnight with primary antibodies against
cytokeratin 7 (CK7; 1:4,000; cat no. ab9021; Abcam) and vimentin
(1:1,000; cat no. sc7557; Santa Cruz Biotechnology, Inc.).
Subsequently, the cells were washed and incubated with either Alexa
Fluor 488 (1:200; cat no. ab150077; Abcam) or 568 (1:200; cat no.
ab175473; Abcam) conjugated goat anti-mouse and goat anti-rabbit
IgG in blocking buffer for 60 min at room temperature. The cells
were mounted on slides with mounting buffer containing DAPI.
Immunofluorescence was detected using a fluorescence microscope
(Olympus Corporation).
miR microarray
The gene expression profiles of PE associated miRs
were retrieved from the NCBI GEO database (http://www.ncbi.nlm.nih.gov/geo) with accession number
GSE96985, which was conducted through GPL20712 platforms
(Agilent-070156 Human miRNA) and included 3 PE placenta tissues and
3 normal placenta tissue samples (19). The ‘limma’ package in R software
(version 4.2) was applied to perform variance analysis on miRs
expression in the microarray. |log Fold Change (FC)|>2 and
P<0.05 were regarded as the criteria for screening the
differentially expressed-miRs (DE-miRs). A heatmap of the obtained
DE-miRs was constructed using the ‘pheatmap’ package.
Reverse transcription-quantitative PCR
analysis
Total RNA was extracted from placenta tissues (100
mg) or cultured cells (1x107 cells) using a mirVana
miRNA Isolation Kit (Thermo Fisher Scientific, Inc.) as per
manufacturer's instructions. The concentration and quality of total
RNA was determined using a NanoDrop 2000 spectrophotometer (Thermo
Fisher Scientific, Inc.). Reverse transcription of miRs (1 µg) was
performed using the TaqMan MicroRNA Reverse Transcription kit
(Thermo Fisher Scientific, Inc.) with gDNA eraser kit (Takara Bio,
Inc.) and miRs expression levels were measured using TaqMan
MicroRNA Assay kit (Thermo Fisher Scientific, Inc.) on the ABI
PRISM 7300 system (Thermo Fisher Scientific, Inc.). For detection
of adenomatous polyposis coli (APC) mRNA, 1 µg RNA was
reverse-transcribed into cDNA using a Reverse Transcription kit
with gDNA eraser kit (Takara Bio, Inc.). Then a SYBR Premix Ex Taq
II (Takara Bio, Inc.) was used for qPCR. U6 and GAPDH were used as
internal controls for miR-424-5p and APC, respectively. The qPCR
thermocycling conditions were: Initial denaturation at 95˚C for 30
sec; 40 cycles of 95˚C for 10 sec and 60˚C for 30 sec. The relative
expression of each gene was calculated using the 2-∆∆Cq
method (20). The following primer
pairs were used for the qPCR: MiR-424 (MIMAT0001341; https://www.mirbase.org/) forward,
5'-CGACCAGCAGCAATTCATGT-3' and universal anti-sense,
5'-GCAGGGTCCGAGGTATTC-3'; miR-142-3p (MI0000434) forward,
5'-GGACGTGTAGTGTTTCCTACT-3'; miR-219a (MIMAT0004567) forward,
5'-GCTAGAGTTGAGTCTGGAC-3'; miR-15a-5p (MIMAT0000068) forward,
5'-GCGACCTAGCAGCACATAATGG-3'; miR-132 (MIMAT0000426) forward,
5'-GCGATAACAGTCTACAGCCA-3'; miR-483 (MIMAT0002173) forward,
5'-ATAGTCACTCCTCTCCTCCC-3'; miR-145-5p (MIMAT0000437) forward,
5'-AGCTCGTCCAGTTTTCCCAGG-3'; U6 forward,
5'-GCTTCGGCAGCACATATACTAAAAT-3' and reverse,
5'-CGCTTCACGAATTTGCGTGTCAT-3'; APC (RefSeq NM_001127511.3) forward,
5'-CGCTTCTGTACCACCCTCAG-3' and reverse, 5-ACCGCAGTTTTACTCCAGGG-3';
GAPDH (RefSeq NM_002046.3) forward, 5'-TCAACGACCCCTTCATTGACC-3' and
reverse, 5'-CTTCCCGTTGATGACAAGCTTC-3'.
Cell transfection
miR-424-5p mimics (sense
5'-CAGCAGCAAUUCAUGUUUUGAA-3' and anti-sense,
5'-CAAAACAUGAAUUGCUGCUGUU-3'), miR mimics negative controls (NC;
sense, 5'-UUCUCCGAACGUGUCACGUTT-3' and anti-sense,
5'-ACGUGACACGUUCGGAGAATT-3'), miR-424-5p inhibitor
(5'-UUCAAAACAUGAAUUGCUGCUG-3') and inhibitor NC
(5'-CAGUACUUUUGUGUAGUACAA-3') were synthesized by Shanghai
GenePharma Co. Ltd.. To overexpress APC, the full-length 3'UTR of
human APC gene was amplified by PCR using human cDNA isolated from
placenta tissues and inserted into pcDNA 3.1 vector (Invitrogen;
Thermo Fisher Scientific, Inc.). The APC targeted small interfering
RNAs (si-APC, 5'-AGUUCUUCUAAAUAUCCAGUA-3') and si-Scramble
(5'-CCAAUUGGUUACUAUAUAUCA-3') were synthesized and purified by
Guangzhou RiboBio Co., Ltd. Cells (1.0x106/well) were
seeded and grown overnight in six-well plates. When the cell
confluence reached 80%, Lipofectamine® 2000 Transfection
Reagent (Invitrogen; Thermo Fisher Scientific, Inc.) was used for
transient transfection of the cells with miR-424-5p mimics (50 nM),
miR-424-5p inhibitor (100 nM), NC (100 nM), 2 µg plasmids or 200 nM
si-RNA at 37˚C according to the manufacturer's guidelines, and cell
samples were collected 48 h later for subsequent experimental
studies, such as western blot and RT-qPCR assays.
Cell viability
Transfected HTR-8/SVneo cells were trypsinized to
prepare a single cell suspension, which was seeded in a 96-well
plate at a density of 5x103 cells per well. After 48 h
incubation at 37˚C, 10 µl CCK-8 solutions (CCK-8; Dojindo
Laboratories, Inc.), were added to each well and the cells were
incubated for further 2 h at 37˚C, then the absorbance was detected
at 450 nm using a microplate reader (Model 680; Bio-Rad
Laboratories, Inc.).
The activity of caspase 3
Caspase-3 activity was measured using a Caspase-3
Activity kit (Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. The optical density at 405 nm was then
detected using a microplate reader (Model 680; Bio-Rad
Laboratories, Inc.).
Transwell assay
Cell invasion was measured using a Transwell assay
(Costar; Corning, Inc.). In brief, 24 h after transfection, 100 µl
of serum-free medium containing 2x104 transfected
HTR-8/SVneo cells were added into the upper chamber precoated with
Matrigel (Becton, Dickinson and Company) at 37˚C for 24 h, while
the complete medium was added to the lower well at 37˚C. After 24 h
incubation at 37˚C, the cells at the bottom were fixed in 4%
formaldehyde solution for 15 min at room temperature and then
stained with 0.1% crystal violet for another 20 min at room
temperature. The cell numbers were counted from six random fields
in each well at magnification x400 under an inverted microscope
(Ts2r-FL; Nikon Corporation).
Wound healing assay
When transfected HTR-8/SVneo cells grown up to ~100%
confluence, the cells were starved for 6 h and wounded with a
sterile 100-µl pipette tip and then cells were cultured in
serum-free medium. Images were taken after 24 h of scratching at
magnification x200 under an inverted microscope (Ts2r-FL; Nikon
Corporation) and the scratch width was calculated using ImageJ
software (National Institutes of Health). The equation for
calculation of the percentage of wound closure is given below.
Wound closure (%)=((W0-Wt)/W0) x100, where W0
represents the wound area at 0 h and Wt represents the wound area
at 24 h (21).
Luciferase reporter assay
Luciferase reporters were generated based on the
Firefly luciferase expressing vector (pmirGLO; Promega
Corporation). The 3'-untranslated region (3'-UTR) of the human APC
gene and its mutant of the theoretical miR-424 binding site were
cloned into the pmirGLO vector to form the reporter vector, named
wild-type (Wt) and mutant-type (Mut) of APC 3'UTR, respectively. To
construct pmirGLO-APC-3'-UTR, a partial 3'-UTR of the APC segment
of human APC mRNA containing the putative miR-424 binding sites was
amplified and cloned into the vector pmirGLO. The human APC mRNA
was extracted from placenta tissues and reverse-transcribed to cDNA
using a Reverse Transcription Kit with gDNA eraser kit (Takara Bio,
Inc.). Mutations within potential miR-424 binding sites were
introduced using the QuikChange Site-Directed Mutagenesis kit (Life
Technologies; Thermo Fisher Scientific, Inc.). When the HTR-8/SVneo
cells grew to 60-70% confluency, the cells were co-transfected with
100 ng luciferase plasmid and 50 ng Renilla luciferase
plasmid along with 100 nM miR-424 mimics/inhibitor or miR-NC using
Lipofectamine 2000® (Invitrogen; Thermo Fisher
Scientific, Inc.). After transfection at 37˚C for 48 h, the firefly
luciferase activity was measured by dual-luciferase assays kit
(Promega Corporation) according to the manufacturer's instructions.
Firefly luciferase activity was normalized to Renilla
luciferase activity.
Western blot
Total proteins were extracted from placental tissues
(100 mg) and cultured cells (1x107 cells) using ice cold
RIPA lysis buffer (Beyotime Institute of Biotechnology) with a
protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA) and the
protein concentration was measured using a BCA kit (Beyotime
Institute of Biotechnology). Proteins (40 µg/lane) were separated
via SDS-PAGE on a 10% gels (Beyotime Institute of Biotechnology)
and transferred onto a PVDF membrane (MilliporeSigma). The PVDF
membrane was blocked with 1% BSA (Sigma-Aldrich; Merck KGaA) for 2
h at room temperature, followed by incubation overnight at 4˚C with
primary antibodies including rabbit monoclonal anti-human APC
(1:1,000; cat. no. 2504; Cell Signaling Technology, Inc.), rabbit
monoclonal anti-human β-catenin (1:1,000; cat. no. 8480; Cell
Signaling Technology), rabbit monoclonal anti-human MMP-9 (1:1,000;
cat. no. 13667; Cell Signaling Technology), rabbit monoclonal
anti-human MMP2 (1:1,000; cat. no. 40994; Cell Signaling
Technology), the antibodies against rabbit polyclonal anti-human
Wnt1 (1:1,000; cat. no. ab15251; Abcam) and rabbit polyclonal
anti-human β-actin (1:2,000; cat. no. ab8227; Abcam). Following
primary antibody incubation, the membrane was incubated with goat
polyclonal secondary antibody to rabbit IgG-H&L Alexa
Fluor® 488 (1:2,000; cat. no. ab150081; Abcam) for 1 h
at room temperature.
Protein bands were visualized using a
chemiluminescence detection system (MilliporeSigma) and protein
expression was quantified using Quantity One software (version
4.6.6, Bio-Rad Laboratories, Inc.). All experiments were conducted
in triplicate.
Statistical analysis
All statistical data were analyzed using GraphPad
Prism 5.0 software (GraphPad Software, Inc.). All experiments were
performed in triplicate and data are expressed as the mean ±
standard deviation. Statistical analysis was performed using
unpaired Student's t-test or one-way ANOVA followed by Tukey's post
hoc test. Pearson's method was used in correlation test. P<0.05
was considered to indicate a statistically significant
difference.
Results
miR-424 is downregulated in placenta
specimens from patients with PE
To identify unique miRs involved in PE, the
microarray data of GSE96985 downloaded from the GEO was first
analyzed and the DE-miRs were screened using R language software
(version 4.2; https://www.r-project.org/). In total, there were 58
differentially expressed miRs, including 31 upregulated and 27
downregulated in PE group in comparison with the normal group
(Fig. 1A). Subsequently, three
upregulated (miR-142-3p, miR-219a and miR-15a-5p) and four
downregulated miRs (miR-424, miR-132, miR-483 and miR-145-5p) were
selected to verify the results of this microarray data. miR-142-3p,
miR-219a and miR-15a-5p were significantly upregulated, while
miR-424, miR-132, miR-483 and miR-145-5p were downregulated in PE
group compared with normal group (Fig.
1B). These results are in line with previous reports (12,22-27),
suggesting the experimental reliability of this microarray results.
miR-424 has previously reported to be lowly expressed in the
placenta of patients with severe PE and closely associated with the
severity of PE (12). Another
report also found that hypoxia downregulates miR-424 in primary
human trophoblasts and miR-424 is associated with hindered
trophoblast differentiation (13).
However, the function and molecular mechanisms of miR-424 in
regulating the migration and invasion of trophoblasts remain
unclear. Therefore, miR-424 was selected for further
investigation.
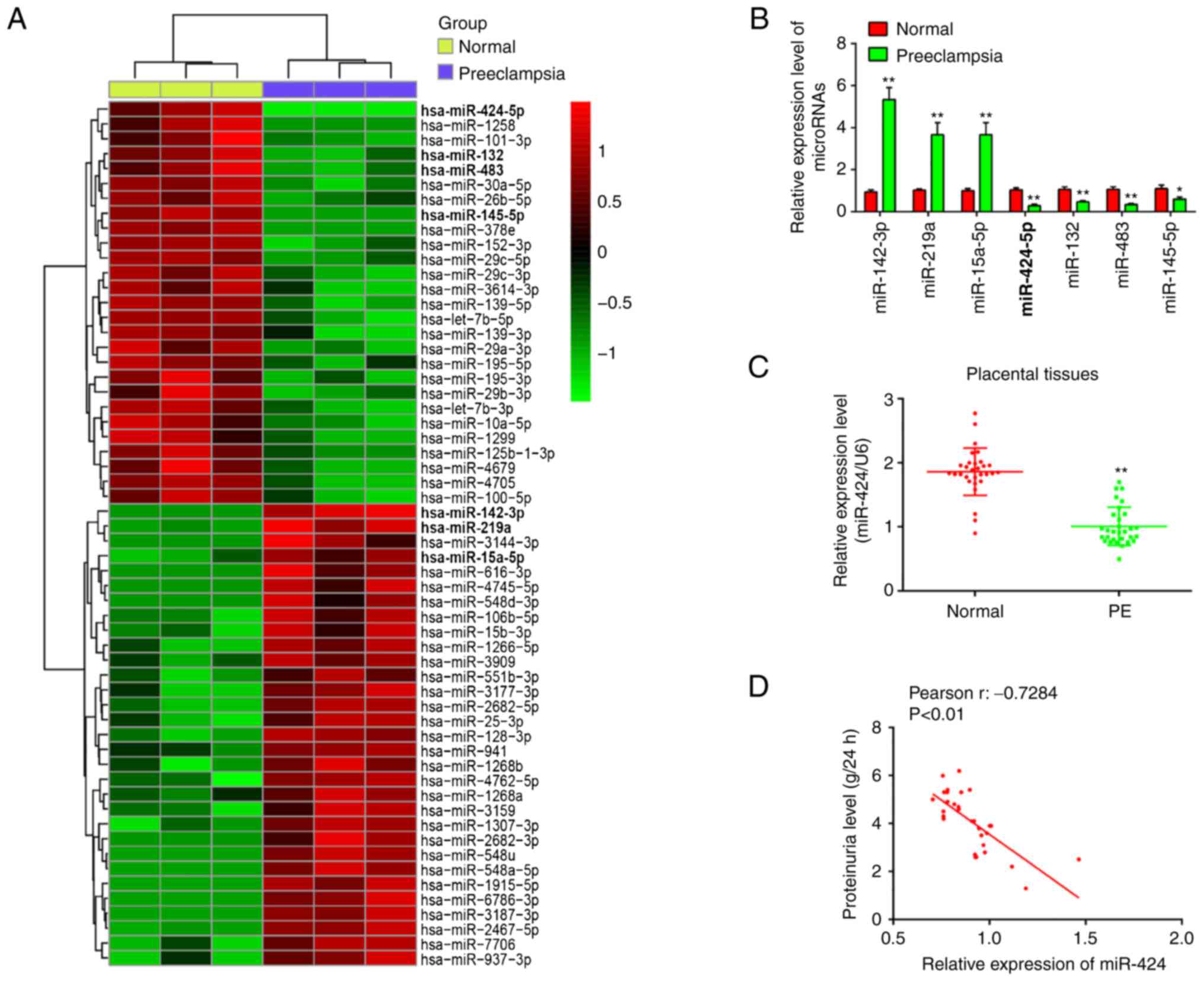 | Figure 1miR-424 is significantly
downregulated in placental tissues. (A) Differentially expressed
miRs between PE and normal group. Data are retrieved from Gene
Expression Omnibus dataset, accession no. GSE96985. The color code
in the heat map is linear and the upregulated miRs are shown in
green to red, whereas the downregulated miRs are shown from red to
green. (B) miR-142-3p, miR-219a, miR-15a-5p, miR-424, miR-132,
miR-483 and miR-145-5p were further analyzed using RT-qPCR. (C)
miR-424 expression was determined in placental tissues from PE
patients (n=30) and healthy pregnant women (n=30) using RT-qPCR.
(D) The positive correlation between miR-424 expression and
proteinuria level in PE pregnancies (r=-0.7284; P<0.01). Data
are presented as means ± standard deviation of three individual
experiments. *P<0.05 and **P<0.01 vs.
control. miRs, microRNA; RT-qPCR, reverse
transcription-quantitative PCR; PE, preeclampsia. |
To further verify the dysregulation of miR-424,
RT-qPCR analysis was performed based on 60 placenta samples
obtained from 30 patients with PE and 30 healthy pregnant women
with uncomplicated pregnancies. The result showed that miR-424
expression levels were much lower in patients with PE than that in
healthy controls (Fig. 1C). In
addition, by analyzing the correlation between miR-424 and
proteinuria levels, it was found that the expression of miR-424 was
negatively correlated with the proteinuria levels in PE pregnancies
(r=-0.7284; Fig. 1D).
Collectively, these data indicated that miR-424 may be a novel
target for the diagnosis of PE.
miR-424 promotes the migratory and
invasive abilities of HTR-8/SVneo cells
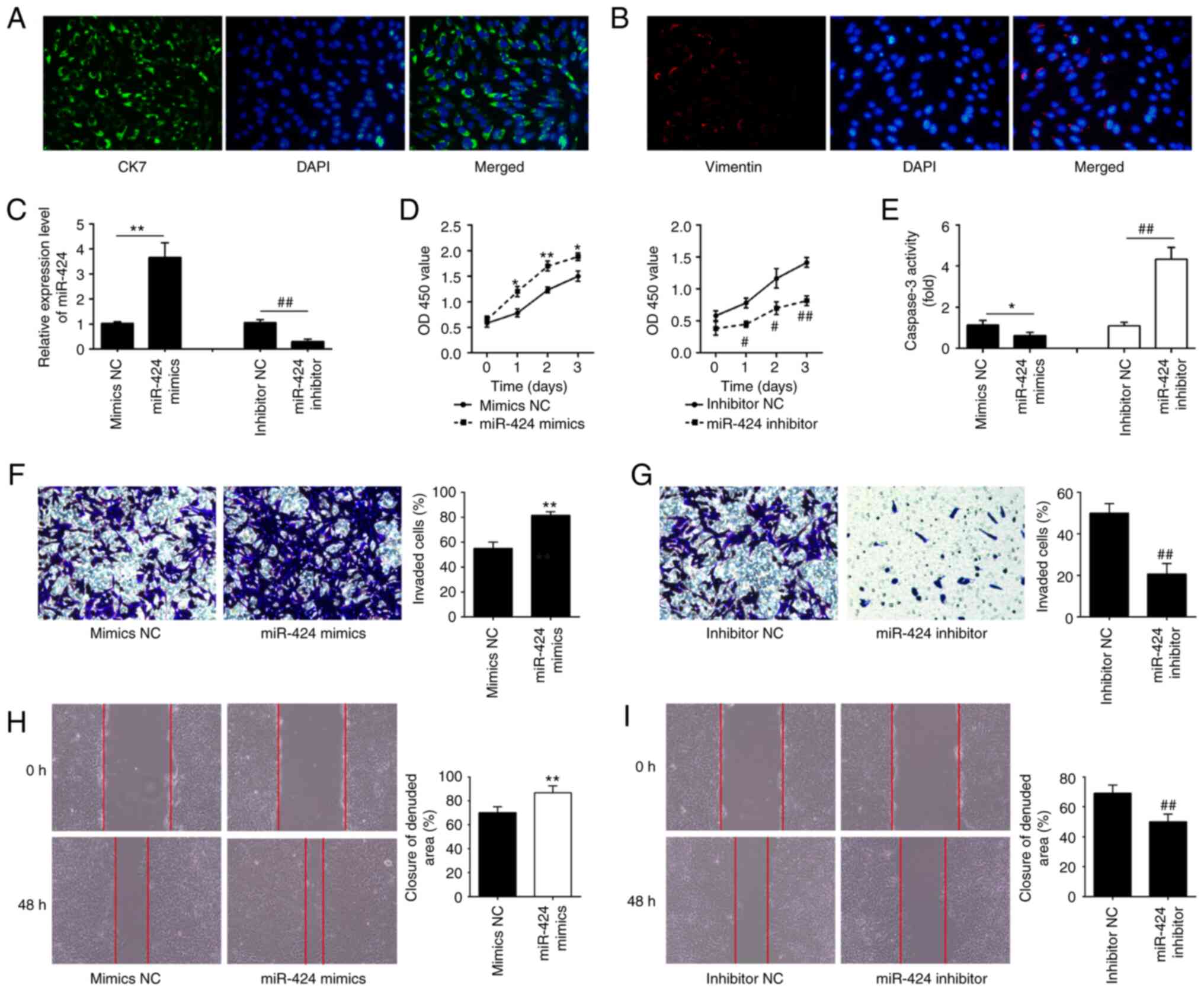
A previous study has shown that HTR-8/SV40neo cells
contain a heterogeneous population of both trophoblast and
mesenchymal cells (17). To
validate the proportion of trophoblast and mesenchymal cells in
this cell lines, CK7 epithelial and vimentin mesenchymal markers
were measured via immunofluorescence analysis. The majority of
cells were positive to CK7, while the percentage of
vimentin-positive cells was low, confirming a prevalence of
placental trophoblast cells (Fig.
2A and B). Given that this
cell line is largely used to study trophoblast invasion/migration
(28), the same was chosen for
subsequent experiments. To further evaluate the effects of miR-424
on HTR-8/SVneo cells invasion and migration in vitro,
miR-424 was overexpressed or knocked down in HTR-8/SVneo cells
using miR-424 mimics or inhibitor. miR-424 mimics caused a
significant upregulation (**P<0.01; Fig. 2C) and miR-424 inhibitor resulted in
a significant downregulation of miR-424 in HTR-8/SVneo cells
(##P<0.01; Fig. 2C).
Reduced activity of trophoblast cells including decreased cell
viability and increased apoptosis is well-recognized to lead to PE
progression (29). CCK-8 and
caspase 3 activity assays were performed to measure the cell
viability and apoptosis of HTR-8/SVneo cells, respectively.
Overexpression of miR-424 significantly enhanced the viability of
HTR-8/SVneo cells compared with that in the mimics NC-transfected
cells (Fig. 2D) and a remarkable
reduction of caspase 3 activity was observed in HTR-8/SVneo cells
(*P<0.05; Fig. 2E),
while miR-424 knockdown had opposite effects (Fig. 2D and E). Moreover, Transwell invasion and wound
healing assays were performed to measure the invasiveness of
HTR-8/SVneo cells. Overexpression of miR-424 significantly promoted
the cell invasion compared with mimics NC group
(**P<0.01; Fig. 2F),
while knockdown of miR-424 had opposite effects
(##P<0.01; Fig. 2G).
miR-424 overexpression significantly increased the migratory
activities of HTR-8/SVneo cells compared with mimics NC group
(**P<0.01; Fig. 2H),
whereas miR-424 knockdown led to a marked reduction of the
migratory activities of HTR-8/SVneo cells compared with the
inhibitor NC group (##P<0.01; Fig. 2I). These results indicated that
miR-424 may be involved in the pathogenesis of PE through
regulating the migration and invasion of HTR-8/SVneo cell.
APC was a target of miR-424 in
HTR-8/SVneo cells
To examine the molecular mechanisms by which miR-424
promoted the migration and invasion of HTR-8/SVneo cells,
TargetScan was used to predict the target genes of miR-424. Among
several candidates (https://www.targetscan.org/cgi-bin/targetscan/vert_80/targetscan.cgi?species=Human&gid=&mirsc=&mirc=&mirnc=&mirvnc=&mirg=hsa-miR-424-5p),
APC was chosen because it is a well-known negative regulator of the
Wnt/β-catenin signaling pathway that has been previously reported
to be associated with PE (30).
One potential binding site for miR-424 was found in the 3'-UTR
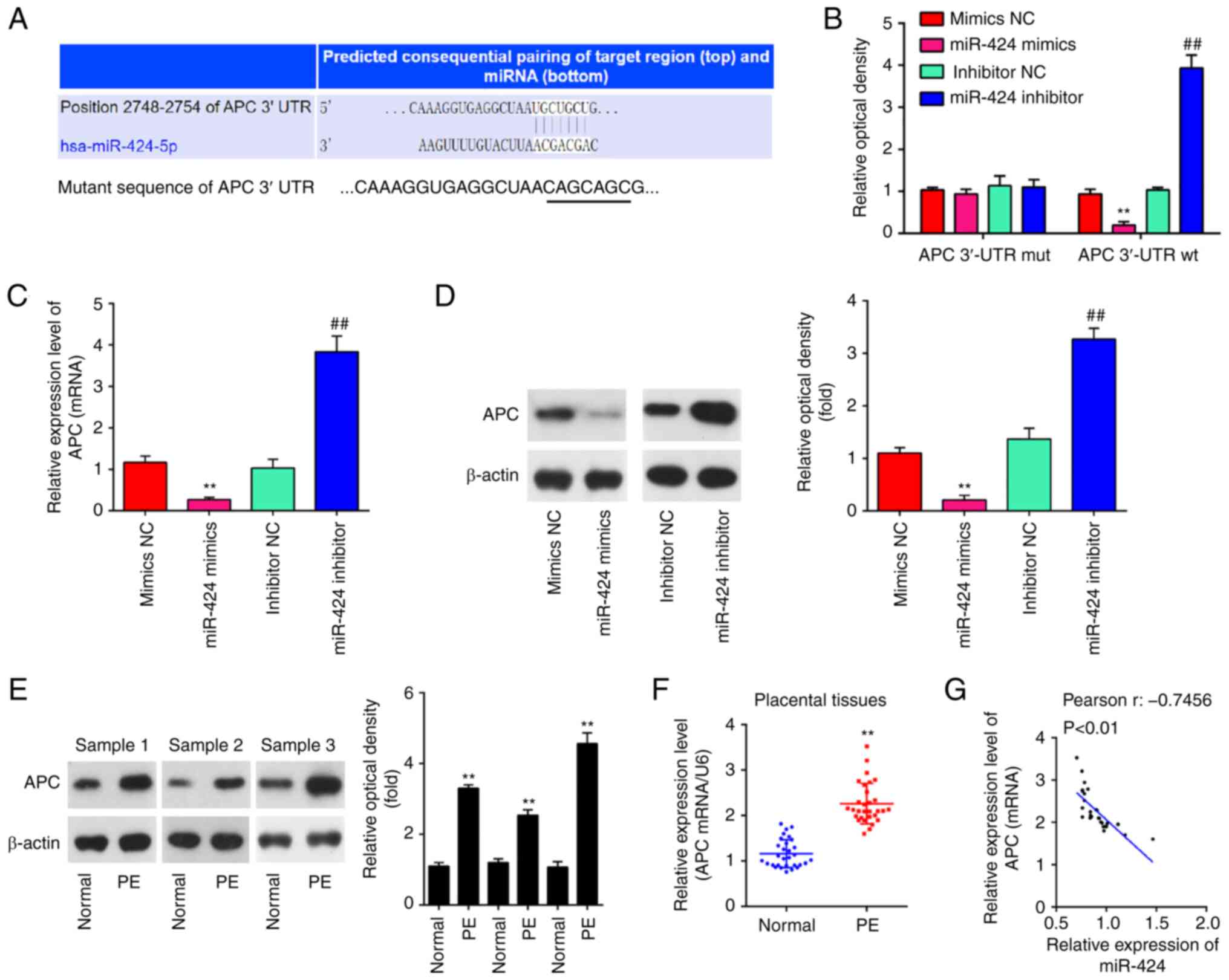
region of APC mRNA (Fig. 3A). To
test whether miR-424 targets APC, dual luciferase reporter assay
was performed. The miR-424 mimics significantly decreased the
luciferase activity, while miR-424 inhibitor increased the
luciferase activity of constructs containing the Wt APC 3'-UTR.
These effects were all lost if the miR-424 binding sites were
mutated (Fig. 3B). Subsequently,
the effect of miR-424 on the expression of APC was measured at the
mRNA and protein levels in HTR-8/SVneo cells using RT-qPCR and
western blot analysis. The mRNA and protein (**P<0.01
for both; Fig. 3C and D) levels of APC were significantly
decreased following the overexpression of miR-424, whereas was
upregulated following knockdown of miR-424 (##P<0.01
for both; Fig. 3C and D). In addition, the protein levels of APC
in placenta samples from 3 PE patients and 3 healthy pregnant women
were measured via western blot analysis. It was shown that APC
protein levels were significantly increased in patients with PE
compared with those in healthy controls (**P<0.01 for
all; Fig. 3E). APC mRNA expression
was significantly increased in patients with PE compared with that
in healthy controls (n=30; **P<0.01; Fig. 3F). Furthermore, a negative
correlation between the levels of expression of APC and miR-424 was
observed in placenta samples from PE patients (r=-0.7456;
P<0.01; Fig. 3G). These
findings indicate that APC is a functional target of miR-424 in
HTR-8/SVneo cells.
miR-424 suppresses the invasion and
migration of HTR-8/SVneo cells by targeting APC
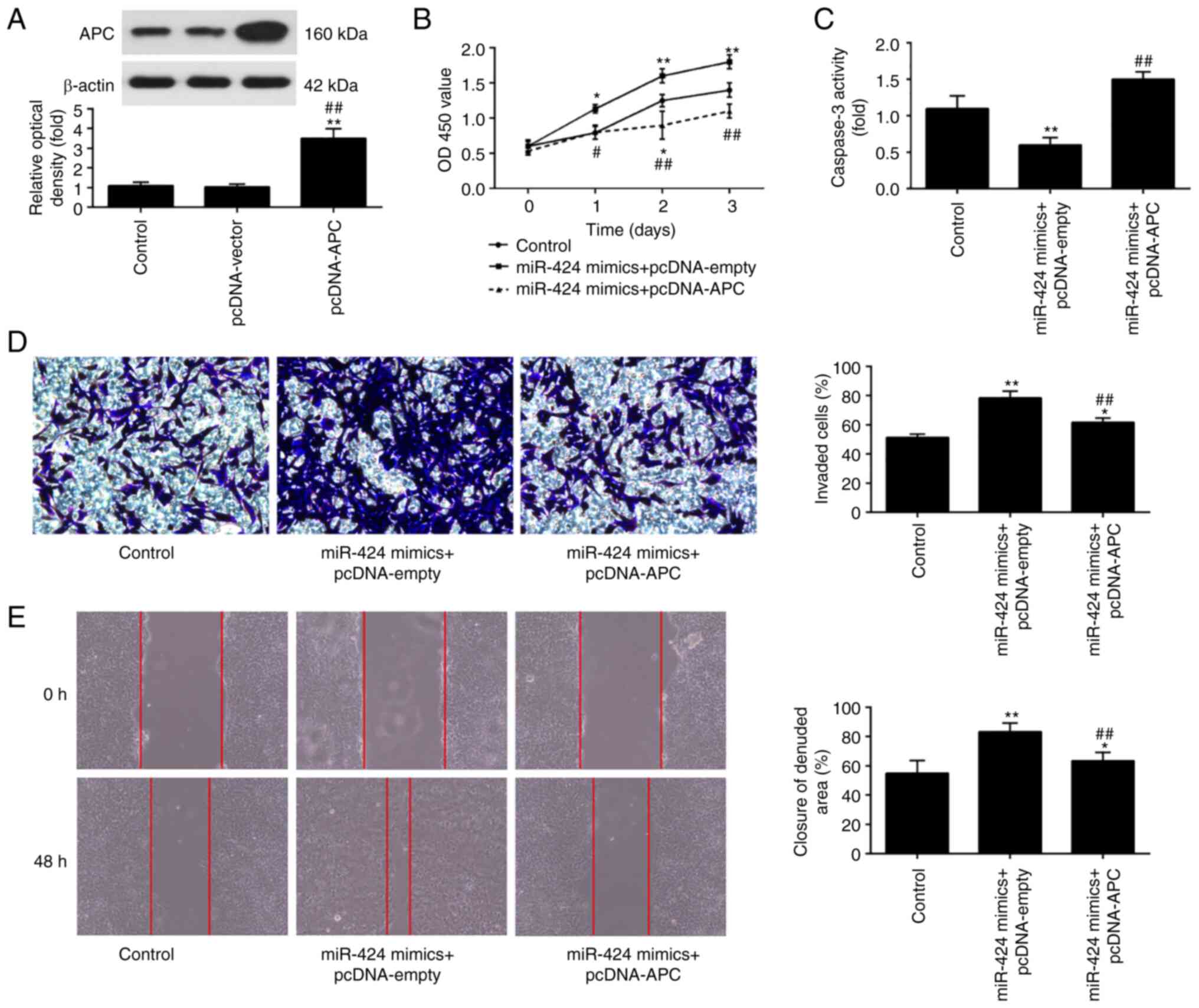
To examine whether APC mediates the positive effects
of miR-424 upregulation on HTR-8/SVneo cells, miR-424 mimics and
pcDNA-APC were co-transfected into HTR-8/SVneo cells. APC protein
expression levels were significantly increased after pcDNA-APC
transfection in HTR-8/SVneo cells (**P<0.01 vs.
control; ##P<0.01 vs. cpDNA-vector; Fig. 4A). Furthermore, it was shown that
miR-424 overexpression resulted in a significant increase in cell
viability, while the same was decreased following APC upregulation
(Fig. 4B). Meanwhile, APC
upregulation reversed the suppressive effect of miR-424 mimics on
the activity of caspase 3 (**P<0.01 vs. control;
##P<0.01 vs. miR-424 mimics + pcDNA-empty; Fig. 4C). In addition, the findings of
Transwell and wound healing assays indicated that APC upregulation
attenuated the promoting effect of miR-424 mimics on the invasion
and migration of HTR-8/SVneo cells (*P<0.05 and
**P<0.01 vs. control; ##P<0.01 vs.
miR-424 mimics + pcDNA-empry; Fig.
4D and E). These findings
demonstrated that upregulation of miR-424 may promote trophoblast
cell invasion and migration by downregulating APC expression.
However, the underlying molecular mechanism remains unclear and
need to be further invetigated.
miR-424 directly targets APC-mediated
Wnt/β-catenin signaling pathway
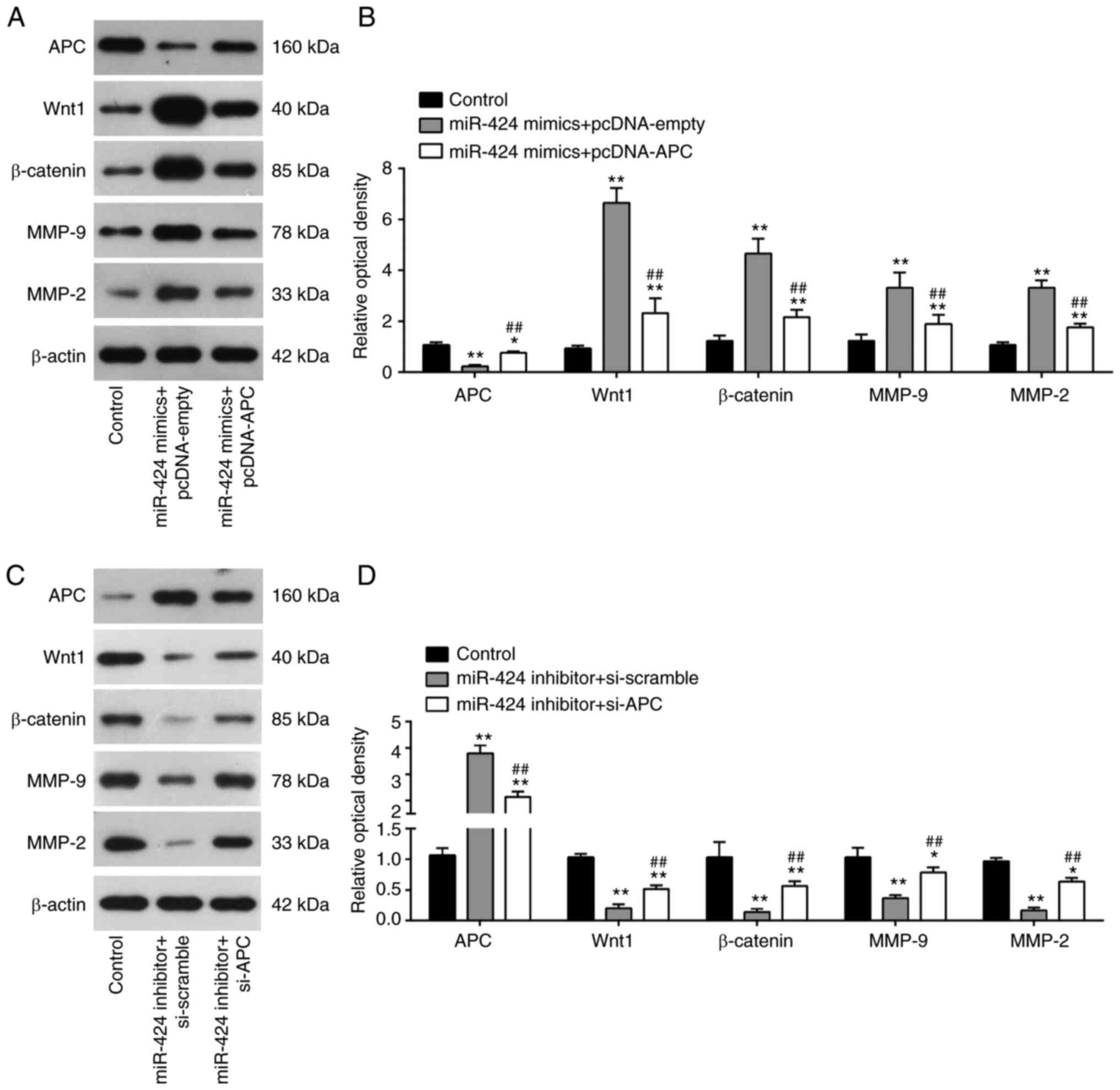
Since APC is a key transcriptional modulator of
Wnt/β-catenin signaling pathway, which play a critical role in the
regulation of trophoblasts migration and invasion (31-33),
western blot analysis was performed to examine if miR-424 affects
APC mediated Wnt/β-catenin signaling pathway in HTR-8/SVneo cells.
miR-424 overexpression significantly increased the expression
levels of Wnt1, β-catenin, MMP-9 and MMP-2, while APC upregulation
attenuated the promoting effects of miR-424 mimics on these
proteins (Fig. 5A and B). In the contrast, it was also found
that miR-424 knockdown markedly decreased the expression levels of
Wnt1, β-catenin, MMP-9 and MMP-2, while APC inhibition reversed the
inhibitory effects of miR-424 knockdown on these proteins (Fig. 5C and D). Thus, these findings indicated that
miR-424 could activate Wnt/β-catenin signaling pathway by directly
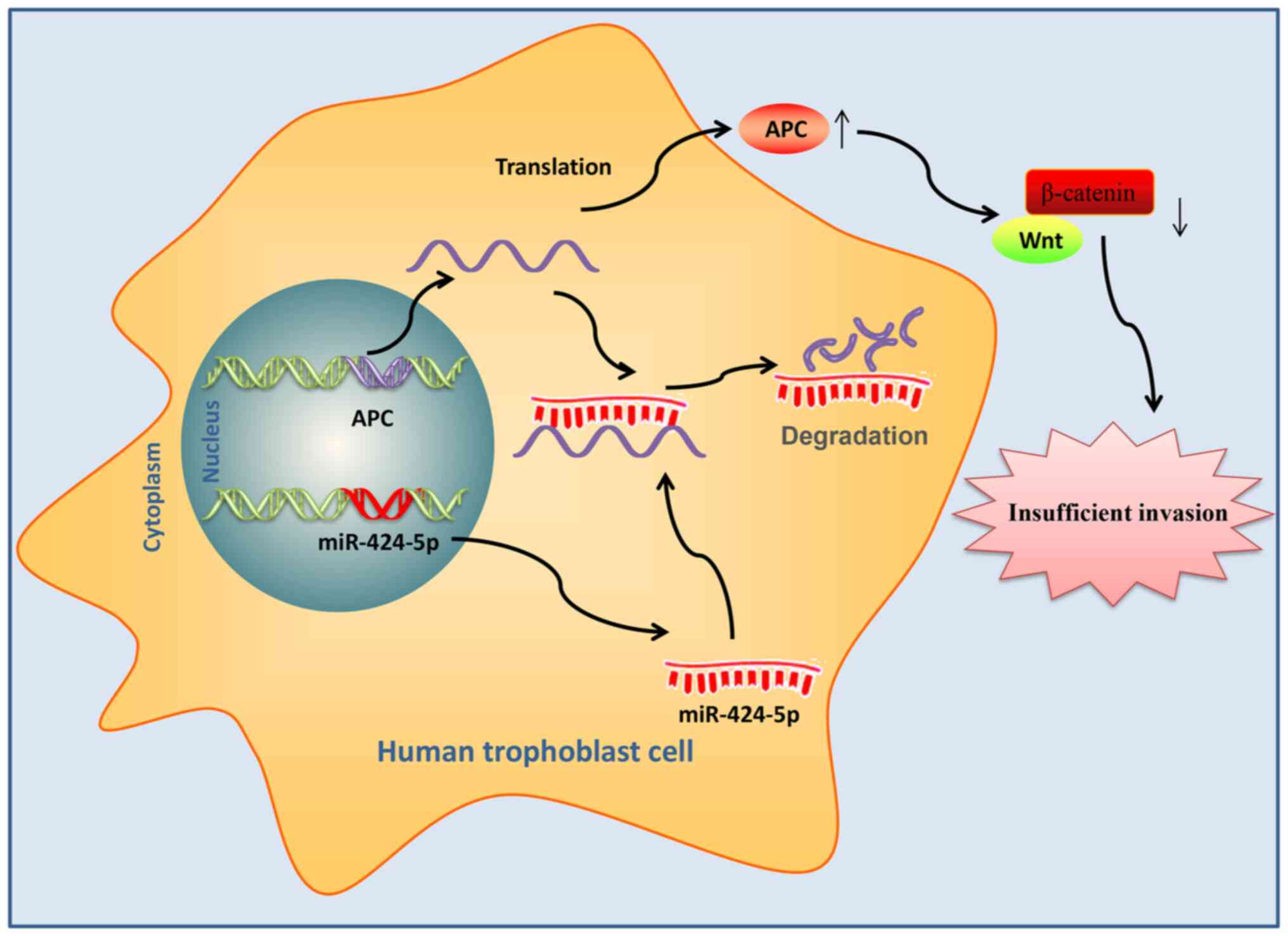
targeting APC (Fig. 6).
Discussion
In the present study, miR-424 was downregulated in
placenta samples obtained from patients with PE and negatively
correlated with proteinuria levels. Moreover, miR-424
overexpression enhanced cell viability, suppressed cell apoptosis
and promoted the migration and invasion of HTR-8/SVneo cells by
targeting APC-mediated Wnt/β-catenin signaling pathway. The present
findings suggested that miR-424/APC axis may serve as a novel
therapeutic target for patients with PE.
It is well-known that shallow trophoblast invasion
is closely associated with the pathogenesis of PE (34). Therefore, in-depth analysis of the
mechanisms of inadequate trophoblast invasion is crucial in order
to improve PE. Several studies have indicated the involvement of
miRs during pregnancy in the regulation of trophoblast invasion
(35,36). Jiang et al (35) showed that miR-520g upregulation
inhibits the invasion of HTR-8/SVneo cells via inhibition of MMP2.
Xiao et al (36) reported
that miR-144 overexpression promotes the trophoblastic cell
migration and invasion through targeting PTEN in PE. A recent study
showed that the miR-424 expression levels were decreased in
placentas of patients with severe PE, suggesting that miR-424 may
play an important role in the development of PE (12). Thus, miR-424-5p was selected in
this study for further investigation.
A number of studies have previously focused on the
roles of miR-424 in the tumor metastasis in various types of human
cancers, such as hepatocellular carcinoma and gastric cancer
(14,37). Liu et al (38) demonstrated that miR-424-5p is
increased in thyroid cancer cells and miR-424-5p overexpression
enhances the invasion and migration of thyroid cancer cells by
inactivating Hippo signaling pathway. In addition, miR-424 has also
been found to be downregulated and act as a tumor suppressor to
repress the migration and invasion in basal-like breast cancer
cells (39). It is generally
considered that trophoblast cells are invasive and share behavioral
characteristics with tumor cells (34). Therefore, we hypothesize that
miR-424-5p may have similar role in the trophoblast invasion and
migration. In the present study, in vitro assays confirmed
that miR-424 overexpression enhances cell viability, suppresses
cell apoptosis and promotes the invasion and migration of
HTR-8/SVneo cells, while miR-424 knockdown exerts opposite effects.
This is in agreement with the findings of Li and Li (40) where miR-424-5p was found to
regulate the proliferation, migration and invasion of trophoblast
cells in PE. However, the exact molecular mechanism by which
miR-424 affects trophoblast invasion and migration has not been
studied in depth.
Previous studies have demonstrated that the
importance of WNT signaling pathway in implantation and placental
development (41,42). Moreover, aberrant Wnt/β-catenin
signaling pathway has been reported to be engaged in the
pathogenesis of PE and blocking the Wnt/β-catenin pathway results
in dysregulation of trophoblast cell proliferation and invasion
(43). A previous study has
demonstrated that stathmin-1 can inhibit the activation of
Wnt/β-catenin signaling pathway and in turn downregulates MMPs
expressions, leading to abnormal trophoblast invasion (44). The abnormal activation of Wnt
signaling may contribute to the process of PE through regulation of
trophoblast cells. The present study revealed that miR-424 can
target APC, a well-known negative regulator of the Wnt/β-catenin
signaling pathway (45). The
regulatory relationship between miR-424 and APC in PE was further
investigated and the results showed that miR-424 negatively
regulates the expression levels of APC in HTR-8/SVneo cells and
that there is an inverse association between the expression levels
of miR-424 and APC in placenta tissues. To the best of our
knowledge, the present results showed for the first time that the
overexpression of APC effectively attenuates the promoting
functions of miR-424 on trophoblast migration and invasion.
Moreover, the present data showed that miR-424 upregulation raises
the levels of Wnt/β-catenin signaling pathway proteins by
suppressing APC. MMPs are proved to be involved in extracellular
matrix remodeling and can degrade extracellular matrix (ECM)
through the function of protease, which in turn affects placental
vascular remodeling and trophoblast cell invasion (46). Notably, MMP-2 and MMP-9 are
gelatinase proteins in the MMPs family (47). The effects of miR-424 on the
expression of MMP-2 and MMP-9 was also evaluated in the present
study. The miR-424 upregulation increased the levels of MMP-2 and
MMP-9 by targeting APC. The present results suggest that miR-424
activates APC-mediated Wnt/β-catenin signaling pathway to promote
trophoblast cells migration and invasion.
Previous studies have shown that the approach for
miR-424 to exert its biological functions is mainly through binding
mRNA to regulate downstream pathways (48,49).
In the present study, the targets of miR-424 were predicted using
miR target prediction tools like TargetScan and microRNA.org. Among these targets, miR-424 regulates
estrogen-related receptor γ to suppress trophoblast proliferation
and invasion (50). Moreover,
miR-424 targeted Wnt3a to influence Wnt/β-catenin signaling pathway
and then affected the proliferation, apoptosis, migration and
invasion of trophoblast cell line HTR-8/SVneo (51). In addition to Wnt/catenin signaling
pathway, miR-424 also wields its function by regulating other
pathways, such as PI3K/AKT signaling pathway in PE (40). In the future, it would be
interesting to clarify if miR-424 plays its role via these
mechanisms in PE.
Although some interesting results were found in the
present study, there were still some limitations. First, this study
only used HTR-8/SVneo cell line, which is the most common cells
used to study trophoblastic dysfunction, therefore key experiments
need to be further performed in different cell lines, such as
TEV-1, ACH-3P, SGHPL-5 and HIPEC65, to corroborate the findings
published in the present study. Additionally, although several
differently expressed miRs were screened based on previously
published microarray data retrieved from GEO database (GSE96985),
only miR-424 was investigated in the present study. Further studies
are required to investigate the functions of other miRs. Third, PE
is a complex pathological process involving several miRs and target
genes (52). The present study
focused exclusively on the miR-424 and APC/Wnt/β-catenin signaling
pathway. The underlying relationship between this pathway and other
related miRs needs further investigations.
In conclusion, the present study demonstrated that
miR-424 promotes trophoblast migration and invasion by targeting
APC and activating Wnt/β-catenin signaling. These findings suggest
that miR-424 upregulation may serve as a potential diagnostic
target for patients with PE. In future, in vivo studies and
clinical trial data are required to validate the preliminary in
vitro results obtained in this study.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WZ performed the experiments, contributed to data
analysis and wrote the paper. XC analysed the data. XC
conceptualized the study design, contributed to data analysis and
experimental materials. WZ and XC confirm the authenticity of all
the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of the Obstetrics & Gynecology Hospital of
Fudan University (approval no. 2018-013). All individuals provided
written informed consent for the use of human specimens for
clinical research.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wagner LK: Diagnosis and management of
preeclampsia. Am Fam Physician. 70:2317–2324. 2004.PubMed/NCBI
|
|
2
|
Kanasaki K and Kalluri R: The biology of
preeclampsia. Kidney Int. 76:831–837. 2009.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Giguère Y, Massé J, Thériault S, Bujold E,
Lafond J, Rousseau F and Forest JC: Screening for pre-eclampsia
early in pregnancy: Performance of a multivariable model combining
clinical characteristics and biochemical markers. BJOG.
122:402–410. 2015.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ray JG, Vermeulen MJ, Schull MJ and
Redelmeier DA: Cardiovascular health after maternal placental
syndromes (CHAMPS): Population-based retrospective cohort study.
Lancet. 366:1797–1803. 2005.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Meekins JW, Pijnenborg R, Hanssens M,
McFadyen IR and van Asshe A: A study of placental bed spiral
arteries and trophoblast invasion in normal and severe
pre-eclamptic pregnancies. Br J Obstet Gynaecol. 101:669–674.
1994.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Sircar M, Thadhani R and Karumanchi SA:
Pathogenesis of preeclampsia. Curr Opin Nephrol Hypertens.
24:131–138. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297.
2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Takahashi H, Ohkuchi A, Kuwata T, Usui R,
Baba Y, Suzuki H, Chaw Kyi TT, Matsubara S, Saito S and Takizawa T:
Endogenous and exogenous miR-520c-3p modulates CD44-mediated
extravillous trophoblast invasion. Placenta. 50:25–31.
2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Fang M, Du H, Han B, Xia G, Shi X, Zhang
F, Fu Q and Zhang T: Hypoxia-inducible microRNA-218 inhibits
trophoblast invasion by targeting LASP1: Implications for
preeclampsia development. Int J Biochem Cell Biol. 87:95–103.
2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Liu E, Liu Z, Zhou Y, Chen M, Wang L and
Li J: MicroRNA1423p inhibits trophoblast cell migration and
invasion by disrupting the TGF-β1/Smad3 signaling pathway. Mol Med
Rep. 19:3775–3782. 2019.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Gao Y, She R, Wang Q, Li Y and Zhang H:
Up-regulation of miR-299 suppressed the invasion and migration of
HTR-8/SVneo trophoblast cells partly via targeting HDAC2 in
pre-eclampsia. Biomed Pharmacother. 97:1222–1228. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tang Q, Gui J, Wu X and Wu W:
Downregulation of miR-424 in placenta is associated with severe
preeclampsia. Pregnancy Hypertens. 17:109–112. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Mouillet JF, Donker RB, Mishima T,
Cronqvist T, Chu T and Sadovsky Y: The unique expression and
function of miR-424 in human placental trophoblasts. Biol Reprod.
89(25)2013.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Wei S, Li Q, Li Z, Wang L, Zhang L and Xu
Z: miR-424-5p promotes proliferation of gastric cancer by targeting
Smad3 through TGF-β signaling pathway. Oncotarget. 7:75185–75196.
2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Li Y, Liu J, Hu W, Zhang Y, Sang J, Li H,
Ma T, Bo Y, Bai T, Guo H, et al: miR-424-5p promotes proliferation,
migration and invasion of laryngeal squamous cell carcinoma. Onco
Targets Ther. 12:10441–10453. 2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Wu K, Hu G, He X, Zhou P, Li J, He B and
Sun W: MicroRNA-424-5p suppresses the expression of SOCS6 in
pancreatic cancer. Pathol Oncol Res. 19:739–748. 2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Abou-Kheir W, Barrak J, Hadadeh O and
Daoud G: HTR-8/SVneo cell line contains a mixed population of
cells. Placenta. 50:1–7. 2017.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Graham CH, Hawley TS, Hawley RG,
MacDougall JR, Kerbel RS, Khoo N and Lala PK: Establishment and
characterization of first trimester human trophoblast cells with
extended lifespan. Exp Cell Res. 206:204–211. 1993.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Chen D, He B, Zheng P, Wang S, Zhao X, Liu
J, Yang X and Cheng W: Identification of mRNA-, circRNA- and
lncRNA- associated ceRNA networks and potential biomarkers for
preeclampsia from umbilical vein endothelial cells. Front Mol
Biosci. 8(652250)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Bahar E and Yoon H: Modeling and
predicting the cell migration properties from scratch wound healing
assay on cisplatin-resistant ovarian cancer cell lines using
artificial neural network. Healthcare (Basel).
9(911)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Mao Y, Hou B, Shan L, Sun X and Wang L:
Aberrantly up-regulated miR-142-3p inhibited the proliferation and
invasion of trophoblast cells by regulating FOXM1. Placenta.
104:253–260. 2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Zhou G, Li Z, Hu P, Wang J, Fu J, Wei B
and Zhang Y: miR-219a suppresses human trophoblast cell invasion
and proliferation by targeting vascular endothelial growth factor
receptor 2 (VEGFR2). J Assist Reprod Genet. 38:461–470.
2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang Y, Du X and Wang J: Transfer of
miR-15a-5p by placental exosomes promotes pre-eclampsia progression
by regulating PI3K/AKT signaling pathway via CDK1. Mol Immunol.
128:277–286. 2020.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wang YP, Zhao P, Liu JY, Liu SM and Wang
YX: MicroRNA-132 stimulates the growth and invasiveness of
trophoblasts by targeting DAPK-1. Eur Rev Med Pharmacol Sci.
24:9837–9843. 2020.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Han L, Luo QQ, Peng MG, Zhang Y and Zhu
XH: miR-483 is downregulated in pre-eclampsia via targeting
insulin-like growth factor 1 (IGF1) and regulates the PI3K/Akt/mTOR
pathway of endothelial progenitor cells. J Obstet Gynaecol Res.
47:63–72. 2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Lv Y, Lu X, Li C, Fan Y, Ji X, Long W,
Meng L, Wu L, Wang L, Lv M and Ding H: miR-145-5p promotes
trophoblast cell growth and invasion by targeting FLT1. Life Sci.
239(117008)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Li X, Li C, Wang Y, Cai J, Zhao L, Su Z
and Ye H: IGFBP1 inhibits the invasion, migration, and apoptosis of
HTR-8/SVneo trophoblast cells in preeclampsia. Hypertens Pregnancy.
41:53–63. 2022.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yu Y, An X and Fan D: Histone deacetylase
sirtuin 2 enhances viability of trophoblasts through p65-mediated
microRNA-146a/ACKR2 axis. Reprod Sci. 28:1370–1381. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Li L, Peng W, Zhou Q, Wan JP, Wang XT and
Qi HB: LRP6 regulates Rab7-mediated autophagy through the
Wnt/β-catenin pathway to modulate trophoblast cell migration and
invasion. J Cell Biochem. 121:1599–1609. 2020.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Douchi D, Ohtsuka H, Ariake K, Masuda K,
Kawasaki S, Kawaguchi K, Fukase K, Oikawa M, Motoi F, Naitoh T, et
al: Silencing of LRRFIP1 reverses the epithelial-mesenchymal
transition via inhibition of the Wnt/β-catenin signaling pathway.
Cancer Lett. 365:132–140. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Clevers H and Nusse R: Wnt/β-catenin
signaling and disease. Cell. 149:1192–1205. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Anastas JN and Moon RT: WNT signalling
pathways as therapeutic targets in cancer. Nat Rev Cancer.
13:11–26. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Carter AM, Enders AC and Pijnenborg R: The
role of invasive trophoblast in implantation and placentation of
primates. Philos Trans R Soc Lond B Biol Sci.
370(20140070)2015.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Jiang L, Long A, Tan L, Hong M, Wu J, Cai
L and Li Q: Elevated microRNA-520g in pre-eclampsia inhibits
migration and invasion of trophoblasts. Placenta. 51:70–75.
2017.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Xiao J, Tao T, Yin Y, Zhao L, Yang L and
Hu L: miR-144 may regulate the proliferation, migration and
invasion of trophoblastic cells through targeting PTEN in
preeclampsia. Biomed Pharmacother. 94:341–353. 2017.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Lu M, Kong X, Wang H, Huang G, Ye C and He
Z: A novel microRNAs expression signature for hepatocellular
carcinoma diagnosis and prognosis. Oncotarget. 8:8775–8784.
2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Liu X, Fu Y, Zhang G, Zhang D, Liang N, Li
F, Li C, Sui C, Jiang J, Lu H, et al: miR-424-5p promotes anoikis
resistance and lung metastasis by inactivating hippo signaling in
thyroid cancer. Mol Ther Oncolytics. 15:248–260. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Wang J, Wang S, Zhou J and Qian Q:
miR-424-5p regulates cell proliferation, migration and invasion by
targeting doublecortin-like kinase 1 in basal-like breast cancer.
Biomed Pharmacother. 102:147–152. 2018.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Li C and Li Q: Circular RNA circ_0111277
serves as ceRNA, targeting the miR-424-5p/NFAT5 axis to regulate
the proliferation, migration, and invasion of trophoblast cells in
preeclampsia. Reprod Sci. 29:923–935. 2022.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wang X, Zhang Z, Zeng X, Wang J, Zhang L,
Song W and Shi Y: Wnt/β-catenin signaling pathway in severe
preeclampsia. J Mol Histol. 49:317–327. 2018.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Fitzgerald JS, Germeyer A, Huppertz B,
Jeschke U, Knöfler M, Moser G, Scholz C, Sonderegger S, Toth B and
Markert UR: Governing the invasive trophoblast: Current aspects on
intra- and extracellular regulation. Am J Reprod Immunol.
63:492–505. 2010.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Zhang Z, Wang X, Zhang L, Shi Y, Wang J
and Yan H: Wnt/β-catenin signaling pathway in trophoblasts and
abnormal activation in preeclampsia (review). Mol Med Rep.
16:1007–1013. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Tian FJ, Qin CM, Li XC, Wu F, Liu XR, Xu
WM and Lin Y: Decreased stathmin-1 expression inhibits trophoblast
proliferation and invasion and is associated with recurrent
miscarriage. Am J Pathol. 185:2709–2721. 2015.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Xing Y, Clements WK, Le Trong I, Hinds TR,
Stenkamp R, Kimelman D and Xu W: Crystal structure of a
beta-catenin/APC complex reveals a critical role for APC
phosphorylation in APC function. Mol Cell. 15:523–533.
2004.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Sahay AS, Jadhav AT, Sundrani DP, Wagh GN,
Mehendale SS and Joshi SR: Matrix metalloproteinases-2 (MMP-2) and
matrix metalloproteinases-9 (MMP-9) are differentially expressed in
different regions of normal and preeclampsia placentae. J Cell
Biochem. 119:6657–6664. 2018.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Li Z, Takino T, Endo Y and Sato H:
Activation of MMP-9 by membrane type-1 MMP/MMP-2 axis stimulates
tumor metastasis. Cancer Sci. 108:347–353. 2017.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Dastmalchi N, Hosseinpourfeizi MA,
Khojasteh SMB, Baradaran B and Safaralizadeh R: Tumor suppressive
activity of miR-424-5p in breast cancer cells through targeting
PD-L1 and modulating PTEN/PI3K/AKT/mTOR signaling pathway. Life
Sci. 259(118239)2020.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Dastmalchi N, Safaralizadeh R,
Hosseinpourfeizi MA, Baradaran B and Khojasteh SMB: MicroRNA-424-5p
enhances chemosensitivity of breast cancer cells to Taxol and
regulates cell cycle, apoptosis, and proliferation. Mol Biol Rep.
48:1345–1357. 2021.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Zou Z, He Z, Cai J, Huang L, Zhu H and Luo
Y: Potential role of microRNA-424 in regulating ERRgamma to
suppress trophoblast proliferation and invasion in fetal growth
restriction. Placenta. 83:57–62. 2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Liu C, Li H, Zhang Y and Ding H: Long
intergenic noncoding RNA 00473 promoting migration and invasion of
trophoblastic cell line HTR-8/SVneo via regulating
miR-424-5p-mediated wnt3a/β-catenin signaling pathway. J Obstet
Gynaecol Res. 47:3034–3046. 2021.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Selvaraj S, Lakshmanan G, Kalimuthu K and
Sekar D: Role of microRNAs and their involvement in preeclampsia.
Epigenomics. 12:1765–1767. 2020.PubMed/NCBI View Article : Google Scholar
|