Introduction
Asthma is a chronic airway inflammatory disease that
threatens human health. At present, >300 million individuals are
diagnosed with asthma worldwide and the total number of patients
with asthma is expected to exceed 400 million by 2025(1). The pathological features of asthma
include airway remodelling and bronchial hyperresponsiveness
(2). As the physical barrier of
the airway, the bronchial epithelium is the first line of defence
against pathogens, environmental pollutants and allergens.
Moreover, the bronchial epithelium is not only involved in airway
mucus hypersecretion and airway remodelling but is also related to
the airway inflammatory response in asthma (3-5).
When bronchial epithelial cells are activated by inflammatory
stimuli, they initiate inflammatory cascades and release a variety
of pro-inflammatory factors (6).
The excessive release of pro-inflammatory factors leads to
pulmonary dysfunction, high mucin production and
protease-antiprotease imbalance (7,8).
Circular RNAs (circRNAs) are a specific class of
non-coding RNAs (ncRNAs), lacking the 5' cap and 3' poly(A) tail of
strand RNAs, with covalently closed-loop structures and are
resistant to exonuclease degradation (9-10).
Several studies suggested that dysregulated circRNAs interfere with
protein-coding transcripts at the post-transcriptional level by
competing with microRNAs (miRNAs or miRs), thereby affecting
development of asthma (11,12).
Huang et al (13) reported
that in CD4+ T cells, highly expressed hsa_circ_0005519
affects the secretion of IL-13/IL-6 by competitively binding to
let-7a-5p, thereby regulating the inflammatory process mediated by
T cells. circRNA ERBB2 promotes the proliferation and migration of
airway smooth muscle cells (ASMCs) via the miR-98-5p/insulin-like
growth factor 1 receptor axis (14). In addition, circVPS33A is highly
expressed in plasma of patients with asthma and an in vitro
model of airway epithelial inflammation where it regulates
high-mobility group box 1 expression via miR-192-5p, resulting in
epithelial cell damage (15).
miRNAs are short ncRNAs ~22 nucleotides in length
and are closely related to the pathogenesis of asthma (16,17).
miRNAs can bind to mRNA in the 3'-untranslated region, which
degrades mRNA and inhibits its translation, thereby altering the
function of other cells (18). By
acting on bronchial epithelial cells, ASMCs and immune cells,
miRNAs cause type 1 T helper (Th1)/Th2 polarisation, airway
remodelling and chronic epithelial inflammation (19). Recent studies have reported
increased miR-338-3p expression in the sera of patients with asthma
treated with anti-IL-5 agents (20,21).
However, the specific role of miR-338-3p in the inflammatory
response in asthma remains unclear.
E26 transformation specific-1 (ETS1) is a
proto-oncogene in various cancers (22). The differential expression of ETS1
has also recently been detected in various types of inflammatory
disease (23). Th2-polarised
CD4+ T cells show increased ETS1 expression and the
expression of inflammatory factors is upregulated after the cells
are transfected to overexpress ETS1(24). A previous study showed that
miR-338-3p targets ETS1 mRNA and inhibits ETS1 expression in
gallbladder cancer (25). However,
the roles of miR-338-3p and ETS1 in asthma remain unclear.
The present study aimed to investigate the role of
circRNA targeting dehydrogenase E1 (circDHTKD1)/miR-338-3p/ETS1
axis in a model of airway epithelial inflammation. The present
results may provide understanding of the role of circDHTKD1 in the
pathogenesis of asthma and provide novel insights into the
molecular mechanisms and therapeutic targets for asthma
research.
Materials and methods
Cell culture and treatments
Human immortalized bronchial epithelial cell line
(BEAS-2B cell line) was purchased from the Type Culture Collection
of the Chinese Academy of Science (Shanghai, China) and cultured
using RPMI-1640 supplemented with 10% foetal bovine serum (both
Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Beijing Solarbio Science & Technology
Co., Ltd.) at 37˚C with 5% CO2. To establish an in
vitro model of asthma epithelial inflammation, cells were
treated with 0, 1, 5 and 10 µg/ml lipopolysaccharide (LPS,
Sigma-Aldrich; Merck KGaA) for 24 h at 37˚C when cells had reached
a confluence of 70-80%. BEAS-2B cells were used as controls
following treatment with an equal volume of PBS for 24 h at
37˚C.
Cell Counting Kit-8 (CCK-8) assay
Cell activity was measured using CCK-8 assay
(Dojindo Laboratories, Inc.) according to the manufacturer's
instructions. BEAS-2B cells were resuspended in PBS and the density
was adjusted to 2x104 cells/ml before plating onto
96-well plates at a density of 5,000 cells/well. After 48 h at
37˚C, 10 µl CCK-8 reagent was added to each well. After 2 h,
absorbance values at 450 nm were measured with a microplate reader
(BioTek Instruments, Inc.).
Cell transfection
BEAS-2B cells were plated into 6-well plates
(5x105 cells/well) and cultured overnight to 60-70%
confluency before transfection. All RNA sequences used for cell
transfection were obtained from Guangzhou RiboBio Co., Ltd.
Transfection complexes containing 50 nM RNAs were transfected into
BEAS-2B cells using Lipofectamine 3000® (Invitrogen;
Thermo Fisher Scientific, Inc.) and serum-free Opti-MEM (Thermo
Fisher Scientific, Inc.) and incubated for 6 h at 37˚C according to
the manufacturer's protocols. Reverse transcription-quantitative
(RT-q)PCR or western blot analysis was performed 48 h after
transfection to assess transfection efficiency. The sequences of
small interfering (si)RNAs and miRNA mimics and inhibitors are
listed in Table I.
 | Table IsiRNA and miR sequences. |
Table I
siRNA and miR sequences.
| RNA | Sense sequence,
5'→3' | Antisense sequence,
5'→3' |
|---|
| si-NC |
UUCUCCGAACGUGUCACGUTT |
ACGUGACACGUUCGGAGAATT |
| si-circ#1 |
GCUGGAAUCUCAGUUGAUCAUTT |
AUGAUCAACUGAGAUUCCAGCTT |
| si-circ#2 |
UGCUGGAAUCUCAGUUGAUCATT |
UGAUCAACUGAGAUUCCAGCATT |
| miR-338-3p
mimics |
UCCAGCAUCAGUGAUUUUGUUG |
CAACAAAAUCACUGAUGCUGGA |
| miR-338-3p mimics
NC |
UCACAACCUCCUAGAAAGAGUAGA |
UCUACUCUUUCUAGGAGGUUGUGA |
| miR-338-3p
inhibitor |
CAACAAAAUCACUGAUGCUGGA | - |
| miR-338 inhibitor
NC |
UUCUCCGAACGUGUCACGUTT | - |
| si-ETS1 #1 |
GAGCUACGAUAGUUGUGAUTT |
AUCACAACUAUCGUAGCUCTT |
| si-ETS1 #2 |
GGAAUUACUCACUGAUAAATT |
UUUAUCAGUGAGUAAUUCCTT |
| si-ETS1 #3 |
AGGGCACCUUCAAGGACUATT |
UAGUCCUUGAAGGUGCCCUTT |
RNA-sequencing (RNA-seq)
BEAS-2B cells were treated with LPS at a
concentration of 5 µg/ml whereas PBS was used in the control group
for 24 h at 37˚C. Total RNA was isolated with RNAiso PLUS (Takara)
reagent and mRNA was enriched by Ribo-off rRNA Depletion kit (cat.
no. N406; Vazyme Biotech Co., Ltd.). 1% agarose gel electrophoresis
and NanoDrop ND-1000 spectrophotometry (Thermo Fisher Scientific,
Inc.) were employed to verify quality and integrity of the
extracted RNAs. RNA-seq libraries were prepared using the
VAHTS® Stranded mRNA-seq V2 Library Prep kit for
Illumina® (cat. no. NR612; Vazyme Biotech Co., Ltd.)
following the manufacturer's recommendations. Briefly, RNA samples
were fragmented and RT of first- and second-strand cDNAs was
performed using with random hexamer primers; cDNAs were purified
and RNA Adapters were ligated at the 5' and 3' ends of DNA
fragments. The quality of the established libraries were examined
using 8% agarose gel electrophoresis, then quantified by Qubit 2.0
Fluorometer (Invitrogen). The final strand-specific cDNA library
(420 µl of a 10 pM library) was subjected to Illumina®
Hiseq X Ten (Illumina, Inc.) platform for 150 bp pair-end
sequencing using HiSeq X Ten Reagent Kit (cat. no. FC-501-2501).
Sequencing reads from RNA-seq were mapped to the human reference
genome (hg19) from UCSC genome database (genome.ucsc.edu/). circRNA identification was
performed using CIRI2 software (version 2.0.6; sourceforge.net/projects/ciri/) based on
the RNA-seq data.
RNA extraction and RT-qPCR
Total RNA was extracted from BEAS-2B cells placed in
a 6-well plate using RNAiso PLUS reagent (Takara, Dalian, China).
The cRNA first strand was synthesized by RT using PrimeScript™ RT
kit (cat. no. RR037A; Takara Biotechnology Co., Ltd.) under the
following reaction conditions: 37˚C for 15 min and 85˚C for 5 sec.
qPCR was performed on an MX3000P qPCR system (Agilent Technologies,
Inc.) using the ChamQ Universal SYBR qPCR® Master Mix
kit (cat. no. Q711; Vazyme Biotech Co., Ltd.), followed by 40
cycles at 95˚C for 15 sec and 64˚C for 30 sec. U6 and GAPDH were
used as internal reference genes to normalize the data. Relative
RNA expression levels were calculated using the 2-ΔΔCq
method (26). The primers were
synthesized by Shanghai Bioengineering Co., Ltd. The forward and
reverse sequences of the primers used for qPCR are shown in
Table II.
| Table IIPCR primer pair sequences. |
Table II
PCR primer pair sequences.
| Gene | Forward sequence,
5'→3' | Reverse sequence,
5'→3' |
|---|
| ETS1 |
AGGGACAGAGCGGAACTCAAC |
AATTGGTCCGCTTCCTGTGTAG |
| GAPDH |
CATGAGAAGTATGACAACAGCCT |
AGTCCTTCCACGATACCAAAGT |
| miR-338-3p |
CGCGTCCAGCATCAGTGATT |
AGTGCAGGGTCCGAGGTATT |
| U6 |
CTCGCTTCGGCAGCACA |
AACGCTTCACGAATTTGCGT |
| IL-6 |
CACTGGTCTTTTGGAGTTTGAG |
GGACTTTTGTACTCATCTGCAC |
| IL-8 |
AACTGAGAGTGATTGAGAGTGG |
ATGAATTCTCAGCCCTCTTCAA |
| TNF-α |
AAGGACACCATGAGCACTGAAAGC |
AGGAAGGAGAAGAGGCTGAGGAAC |
| VEGF |
AGGGCAGAATCATCACGAAGT |
AGGGTCTCGATTGGATGGCA |
| circDHTKD1 |
CTCCCAACTTCAGAGCCAGG |
GCAAGGCCATGATCAACTGAG |
| circRAPGEF5 |
CGTCTAACATCTGCGACTCT |
TCTGGGAAGGCGTATGTTCA |
| circETNK1 |
GGCTTTGGGACTGAAGTTACT |
GTTTCCCACGTAACAGCCAA |
| circNID2 |
GAAGCTACAGGTGTGAGTGC |
TCAGATGCCAAAACTGCCTG |
| circTNS3 |
GCCAGTCAGCACAAAGGAG |
CTTCCTGAGTCTCCCGCC |
| circPDIA3 |
TTTAGAGATGGTGAAGAAGC |
ACTCAGGTGCAAGTCTCTTG |
| circGNB2L1 |
CCCTGGGTGTGTGCAAATAC |
GTTGGTCTCATCCCTGGTCA |
| circTBCD |
AGTGACAAGGCCCGAGATG |
CAACAAGTTCATCATCCATTCTG |
RNA isolation of nuclear and
cytoplasmic fractions
PARIS kit (Ambion Life Technologies) was used to
isolate nuclear and cytoplasmic RNA from BEAS-2B cells following
the manufacturer's instructions. The subcellular localization of
circDHTKD1 was analysed using RT-qPCR as aforementioned. GAPDH and
NEAT1 were used as cytoplasmic and cytosolic controls,
respectively.
Nucleic acid electrophoresis and
Sanger sequencing
PCR products were separated by electrophoresis on a
2% agarose gel and gels were cut and sent to Shanghai Sangon
Bioengineering Co., Ltd. for sequencing following observation in a
UV imaging system.
Dual-luciferase reporter assay
circRNA fragments containing the full length of
circDHTKD1 with either the wild-type (WT) or mutated (MUT) binding
site for miR-338-3p were synthesized and inserted into the
psiCHECK2 vector (Promega Corporation) to generate
psiCHECK2-circ-WT and psiCHECK2-circ-MUT, respectively. BEAS-2B
were co-transfected with the aforementioned luciferase reporter
plasmid and miR-338-3p mimics/miR-338-3p mimics negative control
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instruction.
Following 48 h transfection, the cells were lysed immediately and
the relative luciferase activity of firefly and Renilla
luciferase was determined using a Dual-Luciferase Reporter assay
kit (cat. no. FR201; TransGen Biotech Co., Ltd.) following the
manufacturer's protocol.
Western blot analysis
Total protein was isolated from BEAS-2B cells using
RIPA Lysis Buffer (cat. no. 20-188; Merck KGaA), then quantified
with BCA Protein Assay (cat. no. P0012; Beyotime Institute of
Biotechnology) and the protein was denatured in a 95˚C water bath
for 5 min. Proteins (30 µg/lane) were separated by 10% SDS-PAGE,
transferred onto PVDF membranes (Millipore; Merck KGaA), blocked
with 5% non-fat milk for 2 h at room temperature and incubated
overnight at 4˚C with the following primary antibodies: Anti-ETS1
(cat. no. ab220361; Abcam; 1/1,000); anti-phosphorylated (p)-ERK1/2
(cat. no ab201015; Abcam; 1/1,000); anti-ERK1/2 (cat. no. ab184699;
Abcam; 1/8,000) and anti-GAPDH (cat. no. ab181602; Abcam;
1/10,000). This was followed by incubation with rabbit anti-mouse
horseradish peroxidase-conjugated secondary antibodies (cat. no.
ab6721; Abcam; 1/5,000) for 60 min at room temperature. Protein
bands were scanned and imaged using Immobilon® Western
HRP Substrate ECL chemiluminescence reagent (cat. no. WBKLS0500;
Merck KGaA). Densitometry analysis was performed using ImageJ
software (Version 1.53e; National Institutes of Health) with GAPDH
as the loading control.
ELISA
BEAS-2B cells were seeded into 12-well plates
(1x10^5 cells/well) and treated with or without PBS for 48 h at
37˚C. Subsequently, the cell supernatant was collected and
centrifuged at 1,000 x g for 5 min at 37˚C. According to the
manufacturer's instruction, the concentrations of inflammatory
cytokines IL-6 (cat. no. PI330), IL-8 (cat. no. PI640), TNF-α (cat.
no. PT518) and VEGF (cat. no. PV963) were detected using ELISA kits
(Beyotime Institute of Biotechnology).
Bioinformatics analysis
Cancer-Specific CircRNAs Database (CSCD) (gb.whu.edu.cn/CSCD/) was applied to annotate and
identify exonic circRNAs. The circbank (circbank.cn/) and circRNA interactome (circinteractome.nia.nih.gov/) databases were used
to predict downstream miRNAs. In addition, the binding site of
miR-338-3p ETS1 and ERK2 was predicted using the TargetScan
database (targetscan.org/). The Kyoto Encyclopedia
of Genes and Genomes (KEGG) pathway enrichment analysis of
miR-338-3p target genes was conducted using the Metascape online
tool (version 3.5; metascape.org/) (27).
Statistical analysis
The experimental data from three independent
replicates are expressed as the mean ± standard deviation. Data
were analysed using SPSS 22.0 (IBM Corp.) and GraphPad Prism 8.0
software (GraphPad Software, Inc.; Dotmatics). Paired Student's
t-test was used to assess the difference between two groups, while
one-way ANOVA was used to analyse differences between ≥3 groups
followed by Tukey's post hoc test for pairwise comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of LPS on human airway
epithelial cell viability and inflammatory cytokine secretion
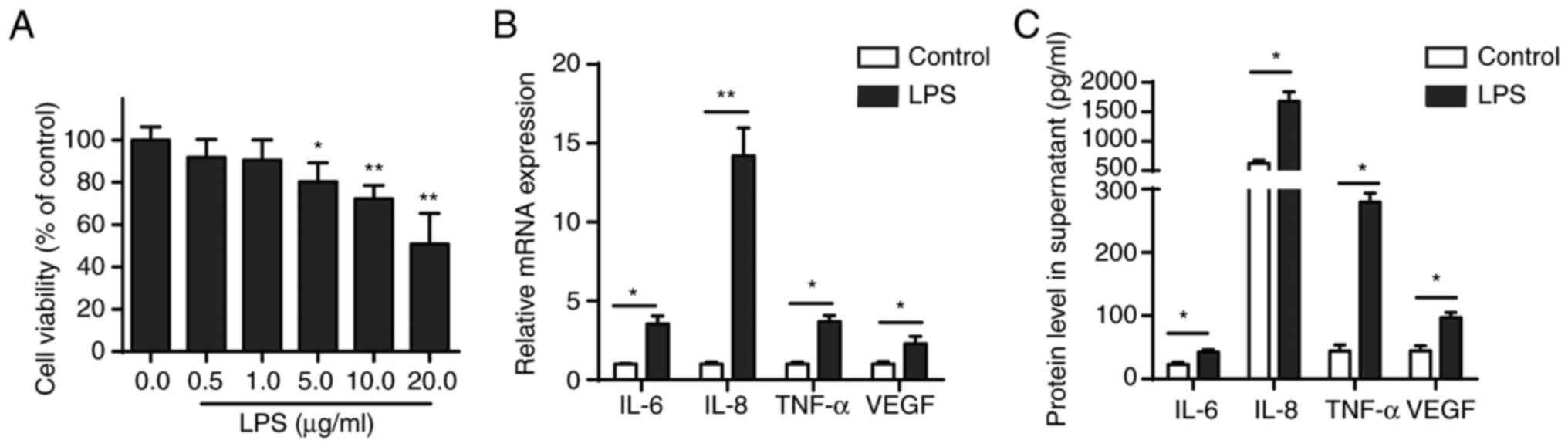
CCK-8 assay was performed to evaluate the effect of
different concentrations of LPS (0.1, 1.0, 10.0 and 100.0 µg/ml) on
the viability of BEAS-2B cells, which were shown to be
concentration-dependent. LPS concentration ≥5 µg/ml significantly
inhibited viability of BEAS-2B cells after 24 h treatment (Fig. 1A). Furthermore, RT-qPCR and ELISA
confirmed that 5 µg/ml LPS increased secretion of pro-inflammatory
cytokines (IL-6, IL-8, TNF-α and VEGF) in BEAS-2B cells (Fig. 1B and C).
circDHTKD1 is significantly
upregulated in LPS-stimulated BEAS-2B cells
The transcriptome of LPS-treated BEAS-2B cells was
sequenced using CIRI2 software for circRNA expression profiling,
followed by selection of exonic circRNAs using CSCD (gb.whu.edu.cn/CSCD/). Finally, the top eight
upregulated exonic circRNAs identified in BEAS-2B cells between
LPS-induced and normal-control were annotated in the circBase
database (circbase.org/; Table III). Changes in expression
profile were analysed by RT-qPCR and showed a significant
upregulation of circDHTKD1 (hsa_circ_0017724) in LPS-induced
BEAS-2B cells compared with that in the control group. The fold
change was increased compared with that of other circRNAs (Fig. 2A). Based on University of
California, Santa Cruz, genome browser (genome.ucsc.edu/) and circBase databases, circDHTKD1
was shown to be circularised from exons 2 and 3 of the DHTKD1 gene
on chromosome 10p14 with a splicing length of 368 bp (Fig. 2B). The back-splice junction of
circDHTKD1 was confirmed by Sanger sequencing. The RT-PCR product
of circDHTKD1 had head-to-tail splicing of the expected size (137
bp fragment) (Fig. 2C), indicating
that the product was a circRNA. In addition, the subcellular
localisation of circDHTKD1 in BEAS-2B cells was identified by
nucleocytoplasmic separation and circDHTKD1 was found to be
significantly enriched in the cytoplasm of BEAS-2B cells (Fig. 2D). These data demonstrated that
circDHTKD1 was upregulated in LPS-induced BEAS-2B cells and was
present in cytoplasm in a circular form.
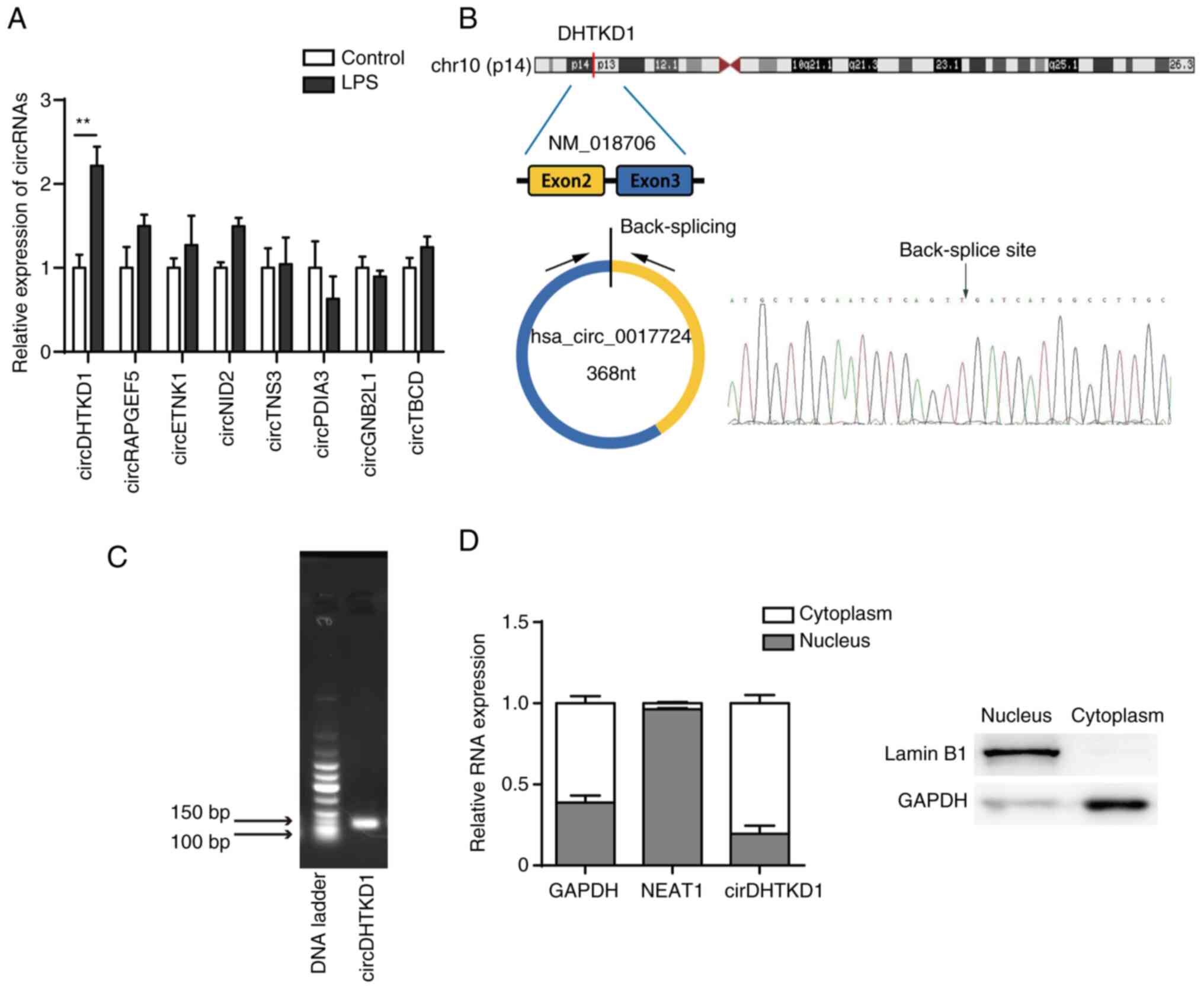 | Figure 2circDHTKD1 is upregulated in
LPS-treated BEAS-2B cells. (A) circRNA expression in BEAS-2B cells
treated with LPS was determined by RT-qPCR. (B) Schematic of
biogenesis of circDHTKD1 and Sanger sequencing of the back-splice
junction site of circDHTKD1. (C) Agarose gel electrophoresis of
circDHTKD1. (D) circDHTKD1 was primarily located in the cytoplasm
of BEAS-2B cells, according to the nuclear-cytoplasmic
fractionation RNA analysis performed by reverse
transcription-quantitative PCR. **P<0.01. LPS,
lipopolysaccharide; DHTKD1, dehydrogenase E1; NEAT1, nuclear
enriched abundant transcript 1; circ, circular; RAPGEF5, Rap
Guanine Nucleotide Exchange Factor 5; ETNK1, Ethanolamine Kinase 1;
NID2, Nidogen 2; TNS3, Tensin-3; PDIA3, Protein Disulfide Isomerase
Family A Member 3; GNB2L1, Guanine Nucleotide-Binding Protein
Subunit Beta-2-Like 1; TBCD, Tubulin Folding Cofactor D. |
| Table IIITop eight upregulated exonic
circRNAs. |
Table III
Top eight upregulated exonic
circRNAs.
| circRNA | Gene | Position, hg
19 | Spliced sequence
length |
|---|
|
hsa_circ_0005281 | Tubulin folding
cofactor D |
chr17:80721840-80730383 | 403 |
| | | strand: + | |
|
hsa_circ_0001681 | Rap guanine
nucleotide exchange factor 5 |
chr7:22330793-22357656 | 516 |
| | | strand: - | |
|
hsa_circ_0002151 | Protein disulphide
isomerase family A member 3 |
chr15:44046021-44048964 | 197 |
| | | strand: + | |
|
hsa_circ_0017724 | Dehydrogenase
E1 |
chr10:12123470-12126750 | 368 |
| | | strand: + | |
|
hsa_circ_0008664 | Ethanolamine kinase
1 |
chr12:22796696-22826594 | 789 |
| | | strand: + | |
|
hsa_circ_0075402 | Guanine
nucleotide-binding protein subunit β-2-like 1 |
chr5:180668491-180669345 | 320 |
| | | strand: - | |
|
hsa_circ_0031930 | Nidogen 2 |
chr14:52495439-52527074 | 1996 |
| | | strand: - | |
|
hsa_circ_0006229 | Tensin-3 |
chr7:47384352-47385954 | 369 |
| | | strand: - | |
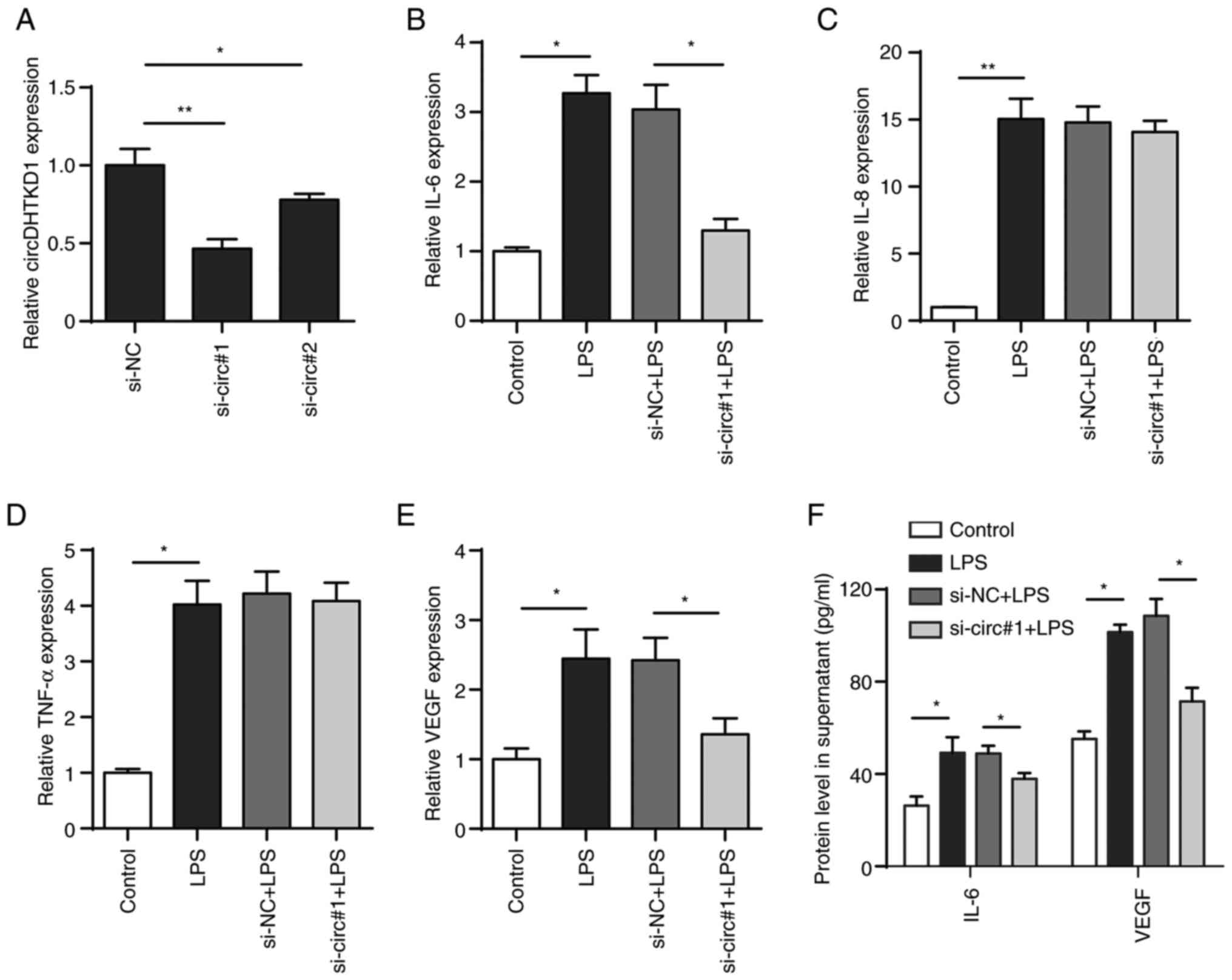
Knockdown of circDHTKD1 inhibits
LPS-induced inflammatory response
To determine the role of circDHTKD1 in the
LPS-induced inflammatory response of BEAS-2B cells, two pairs of
siRNAs targeting circDHTKD1 (si-circ#1 and si-circ#2) were
designed, both of which effectively interfered with the expression
of circDHTKD1. The knockdown of circDHTKD1 caused by si-circ#1 was
notably more efficient compared with that caused by si-circ#2
(Fig. 3A). The qPCR results showed
that the circDHTKD1 knockdown partially decreased mRNA levels of
IL-6 and VEGF but had no significant effect on the mRNA levels of
IL-8 and TNF-α (Fig. 3B-E). ELISA
confirmed the results of the circDHTKD1 knockdown (Fig. 3F). These results indicated that
circDHTKD1 was involved in the regulation of LPS-induced
inflammatory responses in BEAS-2B cells.
circDHTKD1 serves as a sponge for
miR-338-3p
As circRNAs act as miRNA sponges and circDHTKD1 is
primarily localised to the cytoplasm, it was hypothesised that
circDHTKD1 may be involved in regulation of LPS-induced BEAS-2B
cell inflammatory injury response via a competing endogenous RNA
(ceRNA) mechanism. The target miRNAs that bind to circDHTKD1 were
predicted through two databases, namely circBank (circbank.cn/) and circRNA interactome (circinteractome.nia.nih.gov/). miR-338-5p and
miR-338-3p were shared miRNAs in both databases (Fig. 4A). RT-qPCR results showed that the
expression of miR-338-3p significantly decreased following 24 h LPS
induction, whereas expression of miR-338-5p did not change
significantly (Fig. 4B). The
dual-luciferase reporter assay showed that overexpression of
miR-338-3p significantly reduced the luciferase activity of plasmid
psiCHECK2-circ-WT but failed to decrease luciferase activity of
psiCHECK2-circ-MUT (Fig. 4C and
D). In addition, expression of
miR-338-3p was increased after circDHTKD knockdown, whereas no
significant changes in circDHTKD1 expression were detected after
miR-338-3p overexpression (Fig. 4E
and F). The results suggested that
circDHTKD1 interfered with the function of miR-338-3p by acting as
a sponge for miR-338-3p.
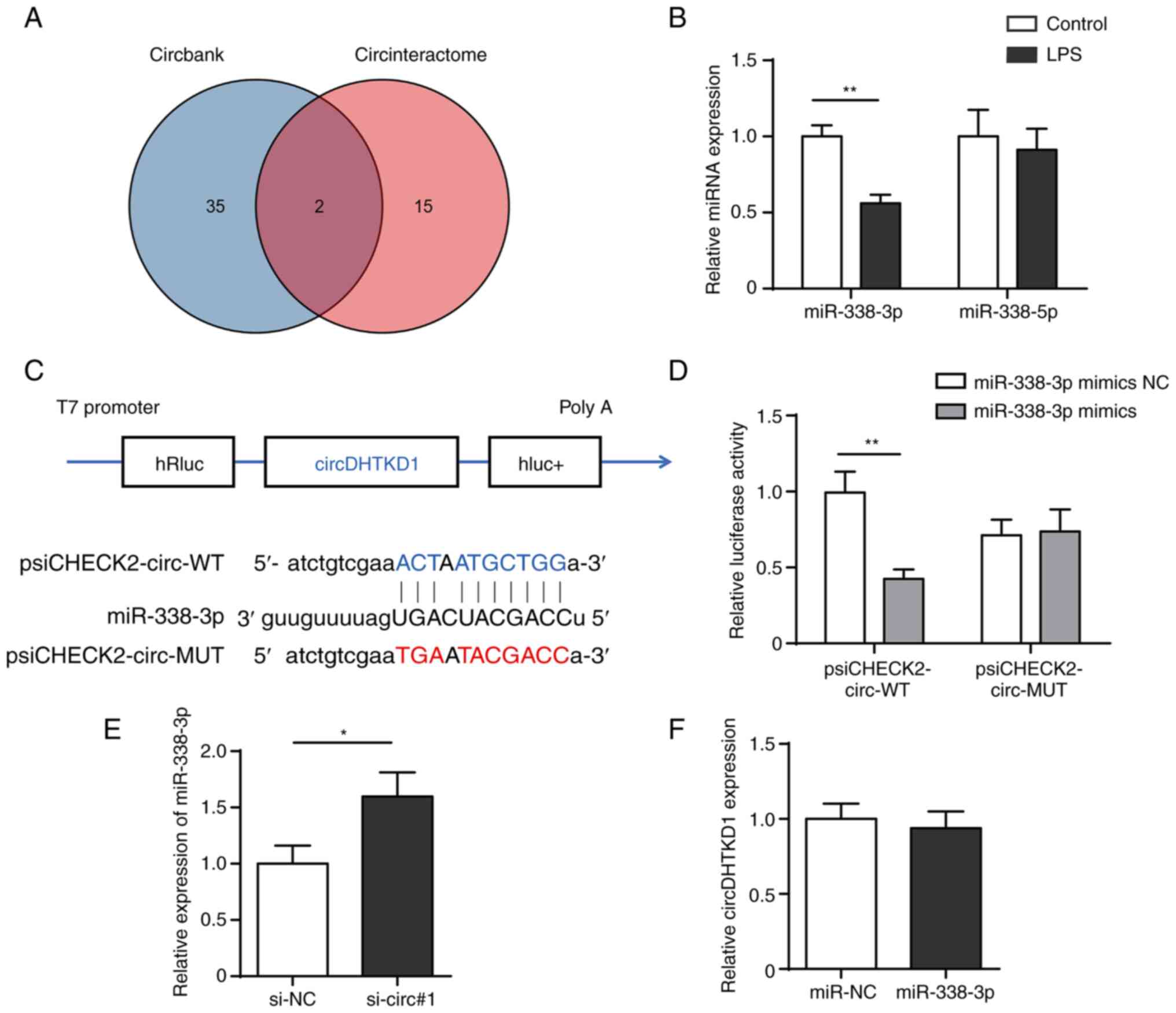 | Figure 4circDHTKD1 serves as a sponge for
miR-338-3p. (A) CircBank and CircInteractome databases were used to
screen miRNAs that potentially interact with circDHTKD1. (B)
RT-qPCR analysis of expression of miR-338-3p and miR-338-5p in
LPS-treated BEAS-2B cells. (C) Schematic of predicted binding sites
of circDHTKD1 and miR-338-3p. (D) Dual-luciferase reporter assay
was used to validate the interaction between circDHTKD1 and
miR-338-3p. (E) miR-338-3p and (F) circDHTKD1 levels were detected
using RT-qPCR. *P<0.05, **P<0.01.
RT-qPCR, reverse transcription-quantitative PCR; hRluc,
Renilla luciferase reporter gene; WT, wild-type; MUT,
mutant; NC, negative control; si, small interfering; DHTKD1,
dehydrogenase E1; circ, circular; miR, microRNA; LPS,
lipopolysaccharide. |
ETS1 is a downstream target gene of
miR-338-3p
TargetScan (targetscan.org/) was used to predict the target genes
of miR-338-3p while Metascape (metascape.org/) was used for cluster analysis of
target genes of miR-338. The results suggested that MAPK signaling
pathway was significantly enriched (Fig. 5A). Further searches using Kyoto
Encyclopedia of Genes and Genomes database (kegg.jp/)
revealed that ERK pathway genes (ERK2 and ETS1) may be regulated by
miR-338-3p (Fig. 5B). The present
results showed that transfection of miR-338-3p mimics in BEAS-2B
cells decreased ETS1 expression levels, while inhibition of
miR-338-3p increase ETS1 expression levels (Fig. 5C and D). In addition, western blotting showed
that ETS1 protein levels significantly decreased following circRNA
knockdown in BEAS-2B cells (Fig.
5E).
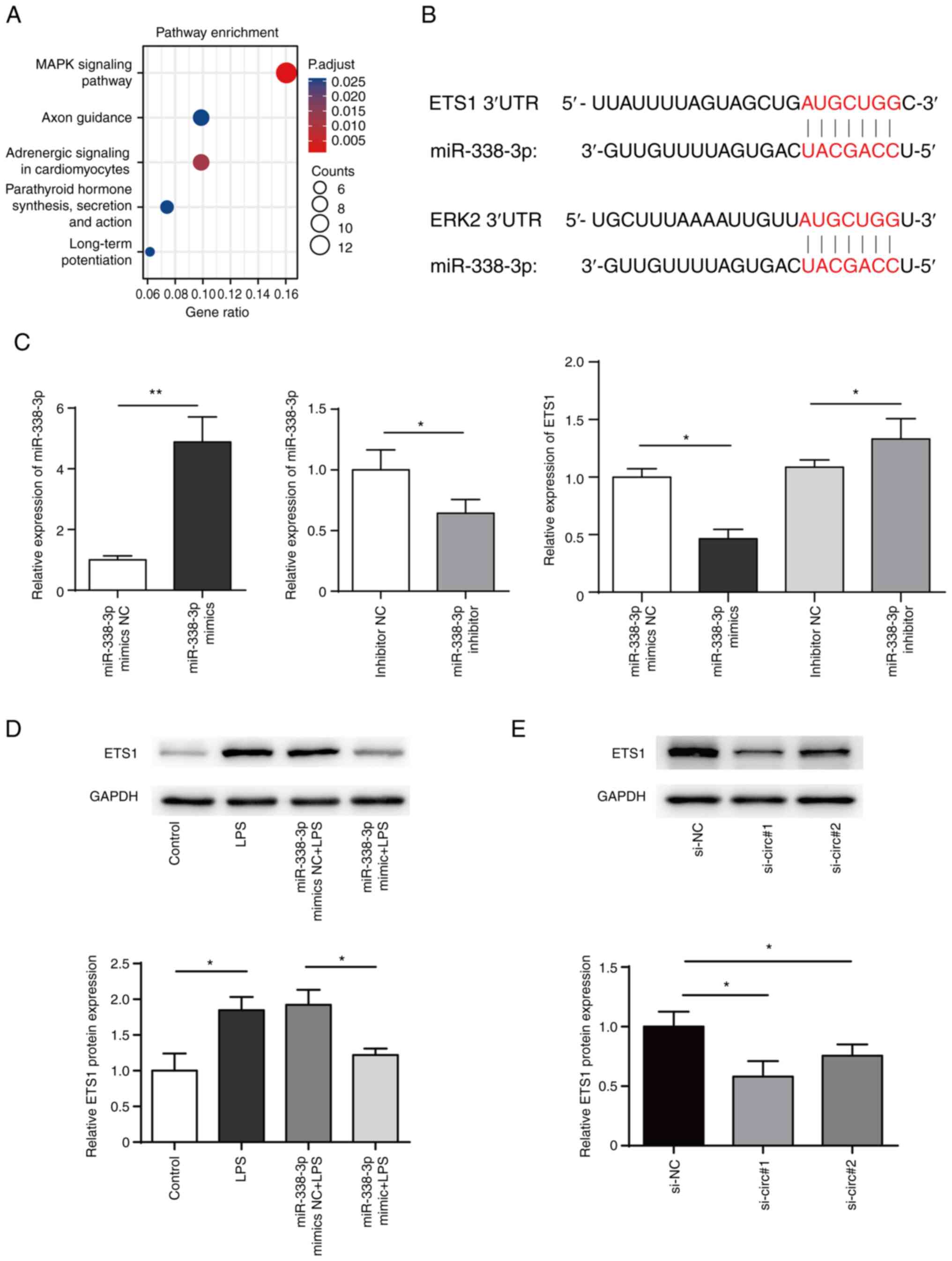 | Figure 5ETS1 is a target of miR-338-3p. (A)
Pathway enrichment of miR-338-3p. (B) Bioinformatics prediction of
the binding site of miR-338-3p to mRNA 3' untranslated region of
ETS1 and ERK2. (C) miR-338-3p mimics, miR-338-3p mimics NC,
miR-338-3p inhibitor or miR-338-3p inhibitor NC were transfected
into BEAS-2B cells and mRNA levels of ETS1 were measured. (D)
Expression of ETS1 in BEAS-2B cells transfected with miR-338-3p
mimics was measured by western blotting. (E) Expression of ETS1 in
BEAS-2B cells transfected with siRNA targeting circDHTKD1 or its
NC. *P<0.05, **P<0.01. ETS1, E26
transformation specific-1; NC, negative control; si, small
interfering; LPS, lipopolysaccharide; 3'UTR, 3' untranslated
region; DHTKD1, dehydrogenase E1; circ, circular. |
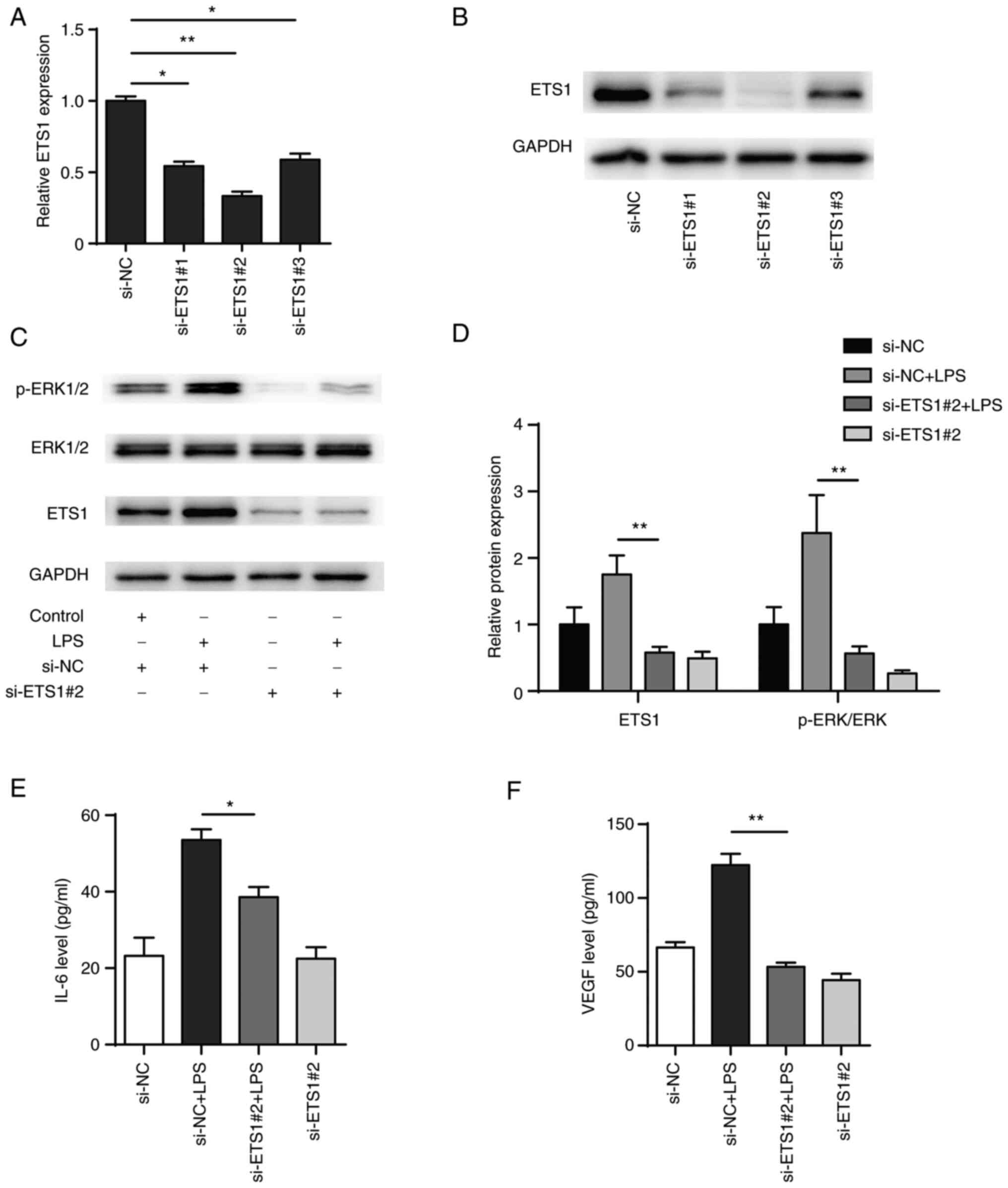
Knockdown of ETS1 attenuates
LPS-induced inflammatory responses in BEAS-2B cells
To explore the role of ETS1 in LPS-induced BEAS-2B
cells, three siRNA pairs targeting ETS1 were designed (si-ETS1#1,
si-ETS1#2 and si-ETS1#3) and transiently transfected into BEAS-2B
cells. Following 48 h transfection, the mRNA and protein levels of
ETS1 were significantly decreased in all transfection groups, with
the si-ETS1#2 transfection showing the highest efficiency in
knocking down ETS1 (Fig. 6A and
B). In addition, western blotting
showed that phosphorylation of ERK1/2 in BEAS-2B cells increased
after LPS stimulation and knockdown of ETS1 significantly inhibited
both the phosphorylation of ERK1/2 induced by LPS and activation of
the ERK signalling pathway (Fig.
6C and D). Also, knockdown of
ETS1 decreased the expression levels of IL-6 and VEGF (Fig. 6E and F). The present results indicated that
ETS1 was involved in the regulation of inflammatory factors IL-6
and VEGF, as well as activation of the ERK pathway.
circDHTKD1 enhances LPS-induced
inflammatory responses in BEAS-2B cells via the miR-338-3p/ETS1
axis
The functional recovery assay showed that LPS
treatment resulted in increased expression of p-ERK compared with
that in the negative control and that p-ERK levels were
significantly decreased when circDHTKD1 was knocked down. This
effect was reversed by transfection with miR-338-3p inhibitor
(Fig. 7A and B). In addition, transfection with
miR-338-3p inhibitor reversed the decrease in IL-6 and VEGF protein
levels that was induced by the circDHTKD1 knockdown (Fig. 7C and D). These results collectively
demonstrated that circDHTKD1 enhanced LPS-induced inflammatory
responses in BEAS-2B cells via the miR-338-3p/ETS1 axis.
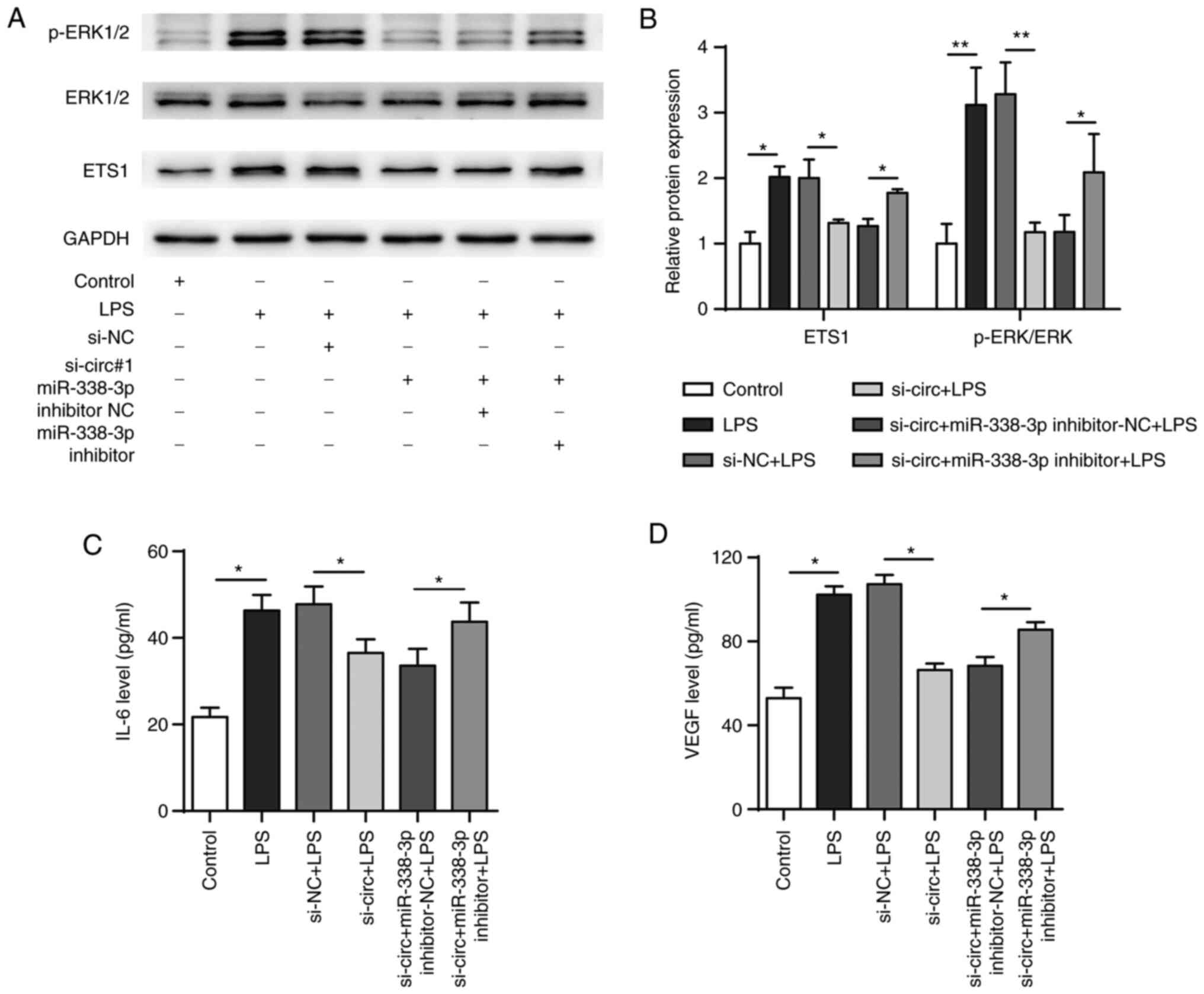 | Figure 7circDHTKD1 promotes LPS-induced
BEAS-2B cell inflammation response by regulating the
miR-338-3p/ETS1 axis. (A) Combined with LPS treatment, BEAS-2B
cells were transfected with si-NC, si-circ#1, si-circ#1 +
inhibitor-NC or si-circ#1 + miR-338-3p inhibitor and ETS1, ERK and
p-ERK protein expression levels detected by western blot and (B)
semi-quantitative analysis (B). Protein expression levels of (C)
IL-6 and (D) VEGF in the culture supernatant of BEAS-2B cells were
detected by ELISA. *P<0.05, **P<0.01.
ETS1, E26 transformation specific-1; NC, negative control; si,
small interfering; LPS, lipopolysaccharide; p, phosphorylated;
circ, circular; DHTKD1, dehydrogenase E1. |
Discussion
Airway epithelial cells are involved in the
pathogenesis of asthma (28).
Repeated damage to epithelial cells triggers and activates airway
remodelling via signalling cascades (29). Several studies have demonstrated
that significantly higher levels of IL-6 and VEGF in
bronchoalveolar lavage fluid and induced sputum in asthmatic
patients are associated with lung function compared with those in
the normal population (30-33).
Serum TNF-α and IL-8 levels are significantly higher in patients
with asthma in acute exacerbation compared with those in stable
patients (34). The present study
treated BEAS-2B cells with LPS and demonstrated that LPS
significantly decreased cell viability while inflammatory factors
were significantly upregulated at the mRNA and protein levels,
demonstrating that LPS induced bronchial epithelial cells to
produce an inflammatory phenotype.
circRNAs are ncRNAs with a closed-loop structure
formed by back splicing that perform gene regulation at the
transcriptional or post-transcriptional level (35). Bao et al (11) characterised the circRNA expression
profile of a mouse asthma model and observed that differentially
expressed circRNAs are involved in lipid metabolism, cell adhesion
and endocytosis and the pathogenesis of allergic asthma. Several
studies have revealed the role of circRNAs in asthmatic airway
epithelial inflammation: Jia et al (36) reported that circRNA_406961
regulates particulate matter 2.5 (PM2.5)-induced inflammation in
human bronchial epithelial cells by interacting with IL
enhancer-binding factor 2 and activating the c-Jun N-terminal
kinase/signal transducer and activator of transcription 3 pathway.
In addition, circRNA104250 and lncRNAuc001.dgp.1 promote
PM2.5-induced inflammatory responses in BEAS-2B cells by
co-targeting miR-3607-5p (37).
The present study showed that circDHTKD1 was upregulated in BEAS-2B
cells treated with LPS. Knockdown of circDHTKD1 resulted in
decreased expression levels of pro-inflammatory cytokines.
Therefore, circDHTKD1 may serve an inflammatory role in asthma
progression.
Aberrantly expressed miR-338-3p is associated with
the pathogenesis of various diseases. miR-338-3p is downregulated
in the lung tissue of patients with pulmonary fibrosis (38). In addition, miR-338-3p is
negatively correlated with VEGF expression levels in hepatoma and
vascular endothelial cells (39,40).
Recent studies showed that the miR-338-3p expression is increased
in the serum of patients with severe asthma treated with anti-IL-5
agents, potentially in association with the MAPK and transforming
growth factor β signalling pathways (20,21).
circRNAs regulate gene expression by competitively binding to
miRNAs and serving as ceRNAs for miRNAs (41,42).
The present study showed that the expression of circDHTKD1 was
upregulated in LPS-induced BEAS-2B cells, whereas miR-338-3p was
downregulated. It was hypothesized that circDHTKD1 regulates the
synthesis of IL-6 and VEGF by targeting miR-338-3p. Although online
bioinformatics analysis showed that miR-338-3p cannot target and
bind IL-6 and VEGF-encoding mRNA, it may affect IL-6 and VEGF
protein synthesis and secretion by regulating activation of the ERK
pathway.
ETS1 is considered a proto-oncogene and several
studies have shown that ETS1 may also be involved in the
pathogenesis of inflammatory disease (23,43).
In addition, Yang et al (44) reported that ETS1 was targeted by
miR-338-3p in 768-O cells. In acute ischemic stroke, the
miR-338-3p/ETS1 axis exerts protective effects on cell viability,
apoptosis and inflammation (45).
Colás-Algora et al (46)
reported that the expression of the transcription factor ETS1 is
upregulated in endothelial cells stimulated with LPS and ETS1
knockdown decreases expression of E-cadherin and exacerbates
disruption of the cell membrane. Studies have demonstrated the
involvement of ETS1 in asthma: Zook et al (47) demonstrated that ETS1 is required
for IL-33 to induce IL-5 and IL-13 production. Wang et al
(24) demonstrated that ETS1
expression is increased in Th2-polarised CD4+ T cells
and overexpression of ETS1 results in increased expression of
Th2-type cytokines (IL-5 and IL-13). Therefore, ETS1 may regulate
the secretion of inflammatory factors in LPS-induced airway
epithelial cells. The present findings suggested that ETS1 was
involved in the regulation of the production of inflammatory
factors IL-6 and VEGF. ETS1 is a key downstream gene of the ERK
pathway and is positively regulated by the ERK pathway (48); however, studies have reported that
ETS1 is also a key gene in the activation of the ERK pathway
(49,50). The present study showed that the
circDHTKD1/miR-338-3p/ETS1 axis promoted activation of the
LPS-induced ERK pathway and inflammatory factor secretion.
Altogether, the present study demonstrated that
circDHTKD1 was upregulated in LPS-induced BEAS-2B cells and
circDHTKD1 knockdown alleviated LPS-induced cellular inflammatory
responses via the miR-338-3p/ETS1 axis. Although circDHTKD1 may be
a potential target for the treatment of asthma, the role of the
circ_0001679/miR-338-3p/ETS1 axis in asthma needs validation in
clinical samples and animal models.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets generated and/or analysed during the
current study are available in the Genome Sequence Archive
(Genomics, Proteomics & Bioinformatics 2021) in National
Genomics Data Centre (Nucleic Acids Res 2022), China National
Centre for Bioinformation/Beijing Institute of Genomics, Chinese
Academy of Sciences repository, www.ngdc.cncb.ac.cn/gsa-human/browse/HRA003481.
Authors' contributions
FQ, SH and JW designed the study and wrote and
edited the manuscript. FQ and JW revised the manuscript. SH and XY
performed the experiments and analyzed the data. XC and SZ were
involved in the conception of the study. All authors have read and
approved the final manuscript. SH and JW confirm the authenticity
of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Busse WW and Rosenwasser LJ: Mechanisms of
asthma. J Allergy Clin Immunol. 111:S799–S804. 2003.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Barcik W, Boutin R, Sokolowska M and
Finlay BB: The role of lung and gut microbiota in the pathology of
Asthma. Immunity. 52:241–255. 2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hellings PW and Steelant B: Epithelial
barriers in allergy and asthma. J Allergy Clin Immunol.
145:1499–1509. 2020.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Gohy S, Hupin C, Ladjemi MZ, Hox V and
Pilette C: Key role of the epithelium in chronic upper airways
diseases. Clin Exp Allergy. 50:135–146. 2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Hammad H and Lambrecht BN: The basic
immunology of asthma. Cell. 184:1469–1485. 2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Gras D, Chanez P, Vachier I, Petit A and
Bourdin A: Bronchial epithelium as a target for innovative
treatments in asthma. Pharmacol Ther. 140:290–305. 2013.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ursini CL, Cavallo D, Fresegna AM, Ciervo
A, Maiello R, Tassone P, Buresti G, Casciardi S and Iavicoli S:
Evaluation of cytotoxic, genotoxic and inflammatory response in
human alveolar and bronchial epithelial cells exposed to titanium
dioxide nanoparticles. J Appl Toxicol. 34:1209–1219.
2014.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Speth JM, Bourdonnay E, Penke LR, Mancuso
P, Moore BB, Weinberg JB and Peters-Golden M: Alveolar epithelial
cell-derived prostaglandin E2 serves as a request signal for
macrophage secretion of suppressor of cytokine signaling 3 during
innate inflammation. J Immunol. 196:5112–5120. 2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9(e1003777)2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Bao H, Zhou Q, Li Q, Niu M, Chen S, Yang
P, Liu Z and Xia L: Differentially expressed circular RNAs in a
murine asthma model. Mol Med Rep. 22:5412–5422. 2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Wang X, Chen H, Liu J, Gai L, Yan X, Guo Z
and Liu F: Emerging advances of non-coding RNAs and competitive
endogenous RNA regulatory networks in Asthma. Bioengineered.
12:7820–7836. 2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Huang Z, Cao Y, Zhou M, Qi X, Fu B, Mou Y,
Wu G, Xie J, Zhao J and Xiong W: Hsa_circ_0005519 increases
IL-13/IL-6 by regulating hsa-let-7a-5p in CD4+ T cells
to affect asthma. Clin Exp Allergy. 49:1116–1127. 2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Huang JQ, Wang F, Wang LT, Li YM, Lu JL
and Chen JY: Circular RNA ERBB2 contributes to proliferation and
migration of airway smooth muscle cells via miR-98-5p/IGF1R
signaling in asthma. J Asthma Allergy. 14:1197–1207.
2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Su Y, Geng L, Ma Y, Yu X and Kang Z and
Kang Z: Identification of circular RNA circVPS33A as a modulator in
house dust mite-induced injury in human bronchial epithelial cells.
Exp Lung Res. 47:368–381. 2021.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Williams AE, Larner-Svensson H, Perry MM,
Campbell GA, Herrick SE, Adcock IM, Erjefalt JS, Chung KF and
Lindsay MA: MicroRNA expression profiling in mild asthmatic human
airways and effect of corticosteroid therapy. PLoS One.
4(e5889)2009.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Feng MJ, Shi F, Qiu C and Peng WK:
MicroRNA-181a, -146a and -146b in spleen CD4+ T lymphocytes play
proinflammatory roles in a murine model of asthma. Int
Immunopharmacol. 13:347–353. 2012.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Specjalski K and Niedoszytko M: MicroRNAs:
Future biomarkers and targets of therapy in asthma. Curr Opin Pulm
Med. 26:285–292. 2020.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Rebane A and Akdis CA: MicroRNAs in
allergy and asthma. Curr Allergy Asthma Rep. 14(424)2014.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Cañas JA, Valverde-Monge M, Rodrigo-Muñoz
JM, Sastre B, Gil-Martínez M, García-Latorre R, Rial MJ,
Gómez-Cardeñosa A, Fernández-Nieto M, Pinillos-Robles EJ, et al:
Serum microRNAs as tool to predict early response to Benralizumab
in severe Eosinophilic Asthma. J Pers Med. 11(76)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rial MJ, Cañas JA, Rodrigo-Muñoz JM,
Valverde-Monge M, Sastre B, Sastre J and Del Pozo V: Changes in
serum MicroRNAs after Anti-IL-5 biological treatment of severe
Asthma. Int J Mol Sci. 22(3558)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Zhou X, Zhou R, Zhou H, Li Q, Hong J, Meng
R, Zhu F, Zhang S, Dai X, Peng G, et al: ETS-1 induces
endothelial-like differentiation and promotes metastasis in
non-small cell lung cancer. Cell Physiol Biochem. 45:1827–1839.
2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Jiang L, Liang J, Wang T, Meng F and Duan
W: ETS proto-oncogene 1 modulates PTP1B expression to participate
in high glucose-mediated endothelial inflammation. Acta Biochim
Biophys Sin (Shanghai). 54:565–573. 2022.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang T, Zhou Q and Shang Y: Downregulation
of miRNA-451a promotes the differentiation of CD4+ T cells towards
Th2 cells by upregulating ETS1 in childhood Asthma. J Innate Immun.
13:38–48. 2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhang L, Yan R, Zhang SN, Zhang HZ, Ruan
XJ, Cao Z and Gu XZ: MicroRNA-338-3p inhibits the progression of
bladder cancer through regulating ETS1 expression. Eur Rev Med
Pharmacol Sci. 23:1986–1995. 2019.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Zhou Y, Zhou B, Pache L, Chang M,
Khodabakhshi A, Tanaseichuk O, Benner C and Chanda SK: Metascape
provides a biologist-oriented resource for the analysis of
systems-level datasets. Nat Commun. 10(1523)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Lambrecht BN and Hammad H: The airway
epithelium in asthma. Nat Med. 18:684–692. 2012.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chung KF: p38 mitogen-activated protein
kinase pathways in asthma and COPD. Chest. 139:1470–1479.
2011.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Neveu WA, Allard JL, Raymond DM, Bourassa
LM, Burns SM, Bunn JY, Irvin CG, Kaminsky DA and Rincon M:
Elevation of IL-6 in the allergic asthmatic airway is independent
of inflammation but associates with loss of central airway
function. Respir Res. 11(28)2010.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Zhang JG, Chen XJ, Liu T and Jiang SJ:
FOXP3+ associated with the pro-inflammatory regulatory T and T
helper 17 effector cells in asthma patients. Exp Ther Med.
12:2753–2758. 2016.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Papadaki G, Bakakos P, Kostikas K, Hillas
G, Tsilogianni Z, Koulouris NG, Papiris S and Loukides S: Vascular
endothelial growth factor and cysteinyl leukotrienes in sputum
supernatant of patients with asthma. Respir Med. 107:1339–1345.
2013.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Hoshino M, Nakamura Y and Hamid QA: Gene
expression of vascular endothelial growth factor and its receptors
and angiogenesis in bronchial asthma. J Allergy Clin Immunol.
107:1034–1038. 2001.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Jiang XG, Yang XD, Lv Z and Zhuang PH:
Elevated serum levels of TNF-α, IL-8, and ECP can be involved in
the development and progression of bronchial asthma. J Asthma.
55:111–118. 2018.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Chen LL: The expanding regulatory
mechanisms and cellular functions of circular RNAs. Nat Rev Mol
Cell Biol. 21:475–490. 2020.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Jia Y, Li X, Nan A, Zhang N, Chen L, Zhou
H, Zhang H, Qiu M, Zhu J, Ling Y and Jiang Y: Circular RNA 406961
interacts with ILF2 to regulate PM2.5-induced inflammatory
responses in human bronchial epithelial cells via activation of
STAT3/JNK pathways. Environ Int. 141(105755)2020.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Li X, Jia Y, Nan A, Zhang N, Zhou H, Chen
L, Pan X, Qiu M, Zhu J, Zhang H, et al: CircRNA104250 and
lncRNAuc001.dgp.1 promote the PM2.5-induced inflammatory response
by co-targeting miR-3607-5p in BEAS-2B cells. Environ Pollut.
258(113749)2020.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Rackow AR, Judge JL, Woeller CF, Sime PJ
and Kottmann RM: miR-338-3p blocks TGFβ-induced myofibroblast
differentiation through the induction of PTEN. Am J Physiol Lung
Cell Mol Physiol. 322:L385–L400. 2022.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhang T, Liu W, Zeng XC, Jiang N, Fu BS,
Guo Y, Yi HM, Li H, Zhang Q, Chen WJ and Chen GH: Down-regulation
of microRNA-338-3p promoted angiogenesis in hepatocellular
carcinoma. Biomed Pharmacother. 84:583–591. 2016.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Wei H, Cao C, Wei X, Meng M, Wu B, Meng L,
Wei X, Gu S and Li H: Circular RNA circVEGFC accelerates high
glucose-induced vascular endothelial cells apoptosis through
miR-338-3p/HIF-1α/VEGFA axis. Aging (Albany NY). 12:14365–14375.
2020.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Zhao X, Cai Y and Xu J: Circular RNAs:
Biogenesis, mechanism, and function in human cancers. Int J Mol
Sci. 20(3926)2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Kristensen LS, Andersen MS, Stagsted L,
Ebbesen KK, Hansen TB and Kjems J: The biogenesis, biology and
characterization of circular RNAs. Nat Rev Genet. 20:675–691.
2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yan M, Komatsu N, Muro R, Huynh NCN,
Tomofuji Y, Okada Y, Suzuki HI, Takaba H, Kitazawa R, Kitazawa S,
et al: ETS1 governs pathological tissue-remodeling programs in
disease-associated fibroblasts. Nat Immunol. 23:1330–1341.
2022.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Yang X, Zhang Y and Fan H: Downregulation
of SBF2-AS1 functions as a tumor suppressor in clear cell renal
cell carcinoma by inhibiting miR-338-3p-targeted ETS1. Cancer Gene
Ther. 28:813–827. 2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Chen C, Wang L, Wang L, Liu Q and Wang C:
LncRNA CASC15 promotes cerebral ischemia/reperfusion injury via
miR-338-3p/ETS1 axis in acute ischemic stroke. Int J Gen Med.
14:6305–6313. 2021.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Colás-Algora N, García-Weber D,
Cacho-Navas C, Barroso S, Caballero A, Ribas C, Correas I and
Millán J: Compensatory increase of VE-cadherin expression through
ETS1 regulates endothelial barrier function in response to TNFα.
Cell Mol Life Sci. 77:2125–2140. 2020.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Zook EC, Ramirez K, Guo X, van der Voort
G, Sigvardsson M, Svensson EC, Fu YX and Kee BL: The ETS1
transcription factor is required for the development and
cytokine-induced expansion of ILC2. J Exp Med. 213:687–696.
2016.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Dittmer J: The biology of the Ets1
proto-oncogene. Mol Cancer. 2(29)2003.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Plotnik JP, Budka JA, Ferris MW and
Hollenhorst PC: ETS1 is a genome-wide effector of RAS/ERK
signalling in epithelial cells. Nucleic Acids Res. 42:11928–11940.
2014.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Ning M, Qin S, Tian J, Wang Y and Liu Q:
LncRNA AFAP-AS1 promotes anaplastic thyroid cancer progression by
sponging miR-155-5p through ETS1/ERK pathway. Bioengineered.
12:1543–1554. 2021.PubMed/NCBI View Article : Google Scholar
|