Introduction
Primary central nervous system lymphoma (PCNSL) is a
rare and aggressive intracranial tumor that accounts for
approximately 2% of all primary central nervous system tumors.
PCNSL has an overall incidence rate of 0.43 per 100,000 people and
a slight predilection for men (1).
In particular, the incidence of PCNSL has increased in patients
over 70 years (2). PCNSL is
composed of diffuse large B-cell lymphoma (DLBCL), which is the
most common histopathological subtype (90-95%), followed by Burkitt
(5%), lymphoblastic (5%), marginal zone (3%), and T-cell lymphoma
(2-3%) (3).
In general, PCNSL tumors are sensitive to
radiotherapy and chemotherapy. High-dose methotrexate (MTX)-based
multiple chemotherapy, including rituximab and cytarabine, has been
used as a standard induction therapy in young patients, and elderly
patients who have multiple comorbidities with renal and bone marrow
dysfunction require a reduction in doses of methotrexate (2,4).
Considering the high radiosensitivity of PCNSL, whole-brain
radiotherapy (WBRT) is often selected as an alternative
consolidation therapy to high-dose chemotherapies. However, delayed
WBRT-related neurotoxicity effects, such as severe cognitive
dysfunction, affect the quality of life of survivors and are
observed more frequently in the elderly (2). Thus, WBRT tends to be deferred until
tumor recurrence or administered with a reduced irradiation dose to
avoid WBRT-related delayed neurotoxicity in the elderly. The median
survival of patients with PCNSL over 70 years of age has not
changed in the last few decades and remains in the range of 6-7
months (2). Although PCNSL is
radiosensitive, radiosensitizers have the potential to reduce the
irradiation dose, avoid delayed neurotoxicity, and maintain the
therapeutic effect of radiotherapy for PCNSL. Therefore, new drug
development for radiosensitizers is essential in PCNSL.
5-aminolevulinic acid (ALA) is a natural precursor
of heme (5). In the final step of
heme synthesis in the mitochondria, Fe2+ is inserted
into protoporphyrin IX (PpIX) to form heme. 5-ALA is rapidly
converted to heme in normal cells, whereas in various tumor cells,
PpIX is not converted to heme; consequently, PpIX selectively and
highly accumulates in the mitochondria of tumor cells (5). 5-ALA-induced PpIX is also a
photosensitizer and exhibits high tumor selectivity. Thus, 5-ALA
has been widely used as a live molecular fluorescence marker for
brain tumor surgery, denoted photodynamic diagnosis, for malignant
gliomas (6), meningiomas (7) and PCNSL (8). In a clinical study of stereotaxic
biopsy for intracranial lymphomas, histopathological analysis
showed that all samples with 5-ALA fluorescence (strong and vague)
contained diagnostic lymphoma tissue, resulting in a positive
predictive value of 100% (8).
A previous study demonstrated that PpIX could
enhance the production of reactive oxygen species (ROS) such as
superoxide, singlet oxygen, and hydroxyl radicals via water
radiolysis induced by ionizing irradiation (IR) (9). We confirmed that radiotherapy with
5-ALA administration enhanced mitochondrial ROS production and host
antitumor response in glioma in vitro and in vivo
(10-13).
This combination therapy is known as radiodynamic therapy (RDT),
and recent experimental studies have demonstrated the efficacy of
RDT using 5-ALA in various cancer cells, including melanoma
(14), colorectal cancer (15), prostate cancer (16), breast cancer (17) and lung cancer (18) cells in vitro and in
vivo. Therefore, we hypothesized that tumors that accumulate
5-ALA-induced PpIX are candidates for RDT. Although previous
studies on 5-ALA/RDT focused on radioresistant malignant neoplasms,
no study focused on radiosensitive malignant neoplasms such as
hematological malignancies.
In the present study, we assessed the synthesis of
5-ALA-induced PpIX and its radiodynamic effect under normal and
hypoxic conditions in lymphoma cells. We then evaluated lymphoma
cells' intracellular ROS production after exposure to IR. We also
discuss the possible mechanism of the radiodynamic effect of 5-ALA
in lymphoma cells and the potential of 5-ALA as a radiosensitizer
for PCNLS.
Materials and methods
Data collection
The present study was conducted at the Department of
Neurosurgery, University of Occupational and Environmental Health,
from January 2021 to July 2022.
Chemicals
5-ALA was purchased from Cosmo Bio Co. Ltd (Tokyo,
Japan). It was then dissolved in fresh culture medium at a final
concentration of 1 µM for intracellular PpIX imaging and 0.3 µM for
other in vitro studies. Among other materials,
2CM-H2DCFDA (DCFD) and Thiazolyl Blue tetrazolium
bromide were purchased from Sigma-Aldrich (Tokyo, Japan) and
Invitrogen (CA, USA), and MitoTracker Deep Red FM and NucBlue™ Live
Cell Stain were purchased from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). The DCFD was dissolved in Hank's Balanced Salt
Solution with calcium and magnesium and without red phenol 1X
(Invitrogen) at a final concentration of 10 µM. In addition, the
MitoTracker Deep Red FM was dissolved in fresh culture medium at a
final concentration of 50 nM, while the Thiazolyl Blue tetrazolium
bromide was dissolved in PBS (-) at a final concentration of 5
mg/ml.
Culture and treatment of cells
A human Burkitt lymphoma cell line (Raji) and two
human brain lymphoma cell lines (HKBML and TK) were used. The Raji
and HKBML lines were obtained from the ATCC and RIKEN BRC Cell
Bank, respectively, while the TK cell line was obtained from the
JCRB cell bank. Raji and TK cells were cultured for several days in
RPMI-1640 supplemented with 10% fetal bovine serum (FBS), while the
HKBML line was cultured in Ham's F12 supplemented with 15% FBS at
37˚C before use. The cell lines were maintained in a humidified
incubator with 5% CO2 at 37˚C. Hypoxic conditions were
induced by placing the cells in a multi-gas incubator with 5%
O2/balanced N2 for 24 h just before each
experiment at 37˚C. Finally, 5-ALA was dissolved in each medium and
incubated with the cell lines for 4 h under normoxic and hypoxic
conditions.
Flow cytometric analysis
After each incubation period, cells were collected
by centrifugation (800 x g for 5 min at 4˚C). Immediately
afterwards, the cells were re-suspended in cold PBS/FBS and
analyzed using a flow cytometer (EC800; Sony Biotechnology, Tokyo,
Japan). Overall, 3x106 cells from each sample were
evaluated. Analyses of flow cytometric data were performed using
FlowJo software (Tree Star Inc., Ashland, OR, USA). The median
fluorescence intensity (MFI) of 5-ALA-induced PpIX was relative to
that of untreated cells in each cell line, and then, the relative
MFI of 5-ALA-induced PpIX was compared in normoxic and hypoxic
conditions, as described previously (13).
Evaluation of PpIX fluorescence
intensity in lymphoma cells
Lymphoma cells were seeded in 35-mm culture flasks
and cultured in complete medium containing 0.3 µM 5-ALA for 4 h.
After 5-ALA treatment, the cells were immediately assessed using a
flow cytometer (excitation, 488 nm; emission, 640/30 nm band-pass
filter). The control cells were not exposed to 5-ALA under normoxic
conditions. The MFI of PpIX for the treated cells relative to that
of the 5-ALA-untreated cells was calculated for each cell line
using FlowJo software.
Simple Western experiments
We used the Simple Western system (ProteinSimple,
Inc., CA, USA), a non-gel-based and western blot-like substitute,
to efficiently and quantitatively analyze the protein expression
levels of the cells (19,20). Whole protein lysates were obtained
by re-suspending cell pellets in RIPA buffer (WAKO Pure Chemical
Co., Osaka, Japan, 182-02451) with a Halt protease inhibitor
cocktail kit (Thermo Fisher Scientific, 78410), and the final
concentration was standardized to 0.75 µg/ml. Simple western blot
analyses were performed with HIF1α antibody (1:50, Proteintech, IL,
USA, 20960-1-AP) and anti-beta actin antibody (1:50, Abcam,
Cambridge, UK, ab8227).
Detection of subcellular localization
of PpIX
Intracellular accumulation of PpIX was detected
using a confocal laser scanning microscope (LSM880; Carl Zeiss,
Jena, Germany), according to a modified method (10,13).
Briefly, lymphoma cells were seeded into a 35-mm flask culture in
fresh medium containing 1 µM 5-ALA and incubated in the dark at
37˚C for 4 h. The cells were re-suspended in PBS/FBS, seeded in
glass-bottom dishes (Asahi Techno Glass, Tokyo, Japan), and
observed immediately. PpIX fluorescence (excitation, 488 nm;
emission, 630-nm long-pass filter) was imaged using a confocal
laser-scanning microscope. All procedures were performed in the
dark.
Evaluation of cell responses to
ionizing irradiation
Lymphoma cell lines were seeded at a density of
1x106 cells/well in 6-well plates. Cells in hypoxic
groups were exposed to hypoxic conditions by placing the
appropriate plates in a multi-gas incubator with 5% O2
for 24 h before the IR treatment. Cells in the 5-ALA treatment
group were cultured in a complete medium containing 0.3 µM 5-ALA
for 4 h. The plates were stored in a light-protected humid field
chamber to avoid photoactivation of 5-ALA-induced PpIX. Cells were
exposed to 2 Gy of IR using a gamma irradiator (Gammacell 40
Extractor; Nordion International, Inc., Ontario, Canada) at a
dose-rate of 0.6 Gy/min. Cells under hypoxic conditions were
maintained at 5% O2 during IR exposure. The response of
cells to IR was evaluated using a colony-forming assay. After 8-10
days of IR exposure, the cells were stained with a Thiazolyl Blue
tetrazolium bromide solution. Three culture dishes were prepared
for each group, and each experiment was performed independently
three times. Only the colonies containing ≥50 cells were counted.
The plating efficiency was determined for the unirradiated controls
that were treated in the same way and maintained under the same
conditions. Finally, the relative survival rates were calculated
for each group.
Evaluation of intracellular levels of
ROS after ionizing irradiation in lymphoma cells
Intracellular production of ROS immediately (0 h)
and 12 h after IR was assessed using DCFD, an oxidant-sensitive
fluorescent probe, using a flow cytometer. Cells were seeded in
35-mm culture flasks and irradiated with 8 Gy in the dark using a
gamma irradiator. During the irradiation treatment, the culture
flasks were kept in the dark at room temperature. Control cells
were treated using the same procedure, but without exposure to
5-ALA and IR. Cells in the hypoxia groups were kept under 5%
hypoxic conditions during IR exposure, treated in the same way, and
maintained under the same conditions as the normoxic groups after
IR. Cells in the 5-ALA group were treated with 0.3 µM 5-ALA for 4 h
and then immediately exposed to IR. To evaluate intracellular level
of ROS immediately after IR and after 12 h, each cell was
re-suspended in 10 µM of DCFD solution after IR and incubated at
37˚C for 15 min before analysis. Then, DCFD fluorescence was
analyzed using a flow cytometer (excitation, 488 nm; emission,
525/50 nm band-pass filter), as described above. All procedures
were conducted in the dark. Analyses of flow cytometric data were
performed using FlowJo software. In addition, the MFI of DCFD for
treated cells relative to that of control cells was calculated for
each cell line using FlowJo software.
Evaluation of mitochondrial density in
lymphoma cells
Lymphoma cells were seeded into 35-mm glass-base
dish in fresh medium containing 50 nM MitoTracker working solution
and incubated in the dark at 37˚C for 30 min. To stain the nuclei,
NucBlue was added to the dish and incubated under the same
conditions for 15 min. MitoTracker (excitation, 488 nm; emission,
630-nm long-pass filter) and NucBlue (excitation, 405 nm; emission,
405/488 nm band-pass filter) fluorescence were imaged using a
confocal laser scanning microscope. All procedures were performed
in the dark. Mitochondrial density in lymphoma cells was calculated
using microdensitometry. Based on previous studies (11,21),
microdensitometry for quantitative evaluation of mitochondrial
density in lymphoma cells was performed on these images using the
public domain software ImageJ 1.53e (National Institute of Health,
Bethesda, MD, USA), with modifications to the original methods.
Briefly, the image data of all samples, including mitochondria and
nuclear fluorescence images, were transferred to ImageJ. The
‘freehand’ tool was used to delineate the whole cell and nucleus
and then measure the areas of the whole cell and nucleus,
respectively. The cytosol areas were calculated by subtracting the
areas of the nuclei from those of the whole cells. Next, the
mitochondrial fluorescence image was converted to an 8-bit
grayscale image using ImageJ. The freehand tool was then used to
delineate the whole cell as the region of interest (ROI), and the
mean gray value (MGV) of the mitochondrial fluorescence within the
ROI was plotted on a graph. Finally, mitochondrial density was
calculated by dividing the MGV of mitochondrial fluorescence by the
area of the cytosol.
Statistical analysis
Data are presented as the means ± standard error
(SE) of the mean. Statistical analyses were performed using EZR
version 1.54 (https://www.jichi.ac.jp/saitama-sct/SaitamaHP.files/statmed.html),
which is a graphical user interface for R version 4.0.3 (https://cran.r-project.org/) and R commander 2.7-1
(https://socialsciences.mcmaster.ca/jfox/Misc/Rcmdr/).
The relative MFI of the PpIX fluorescence, and the survival rate of
cells in the colony forming assay were analyzed using unpaired
Student's t-tests. The relative MFI of the DCFD fluorescence (ROS)
and the mitochondrial density were analyzed with a one-way analysis
of variance followed by Bonferroni's test. A P-value of <0.05
was considered to indicate statistical significance (*P<0.05,
**P<0.01)
Results
Evaluation of accumulation of
5-ALA-induced PpIX in lymphoma cells
We first examined the intracellular accumulation of
5-ALA-induced PpIX under normoxic and hypoxic conditions using flow
cytometric analyses of the lymphoma cells (Figs. 1A and S1). The relative MFI of PpIX
fluorescence in the 5-ALA-treated cells was obviously increased
compared to that in the 5-ALA-untreated group in each cell line,
with some variations. The relative MFI of PpIX fluorescence (mean ±
SE) was 26.9±2.69 in the Raji line, 8.04±0.97 in the HKBML line,
and 5.35±1.2 in the TK line under the normoxic conditions,
respectively. Under the hypoxic conditions, the relative MFI of
PpIX fluorescence (mean ± SE) was 35.9±2.97 in the Raji line,
7.04±1.42 in the HKBML line, and 14.28±2.66 in the TK line,
respectively. In the Raji and TK lines, the relative MFI of PpIX
fluorescence under the hypoxic conditions was significantly higher
than that under the normoxic conditions (P<0.05 in Raji, and
P<0.01 in TK). However, there was no significant difference in
PpIX fluorescence between the conditions for HKBML (P=0.57). In the
imaging examination of 5-ALA-induced PpIX, we confirmed PpIX
accumulation in the 5-ALA-treated lymphoma cells using a confocal
laser scanning microscope (LSM880, Zeiss). PpIX fluorescence
accumulated densely in the cytoplasm but not in the nucleus of each
cell line (Fig. 1B). Further, to
evaluate the effect of the hypoxic conditions on each cell line, we
confirmed the expression of HIF-1a using a Simple Western system.
All cells showed induction of HIF-1a under 5% O2
conditions (Fig. 1C).
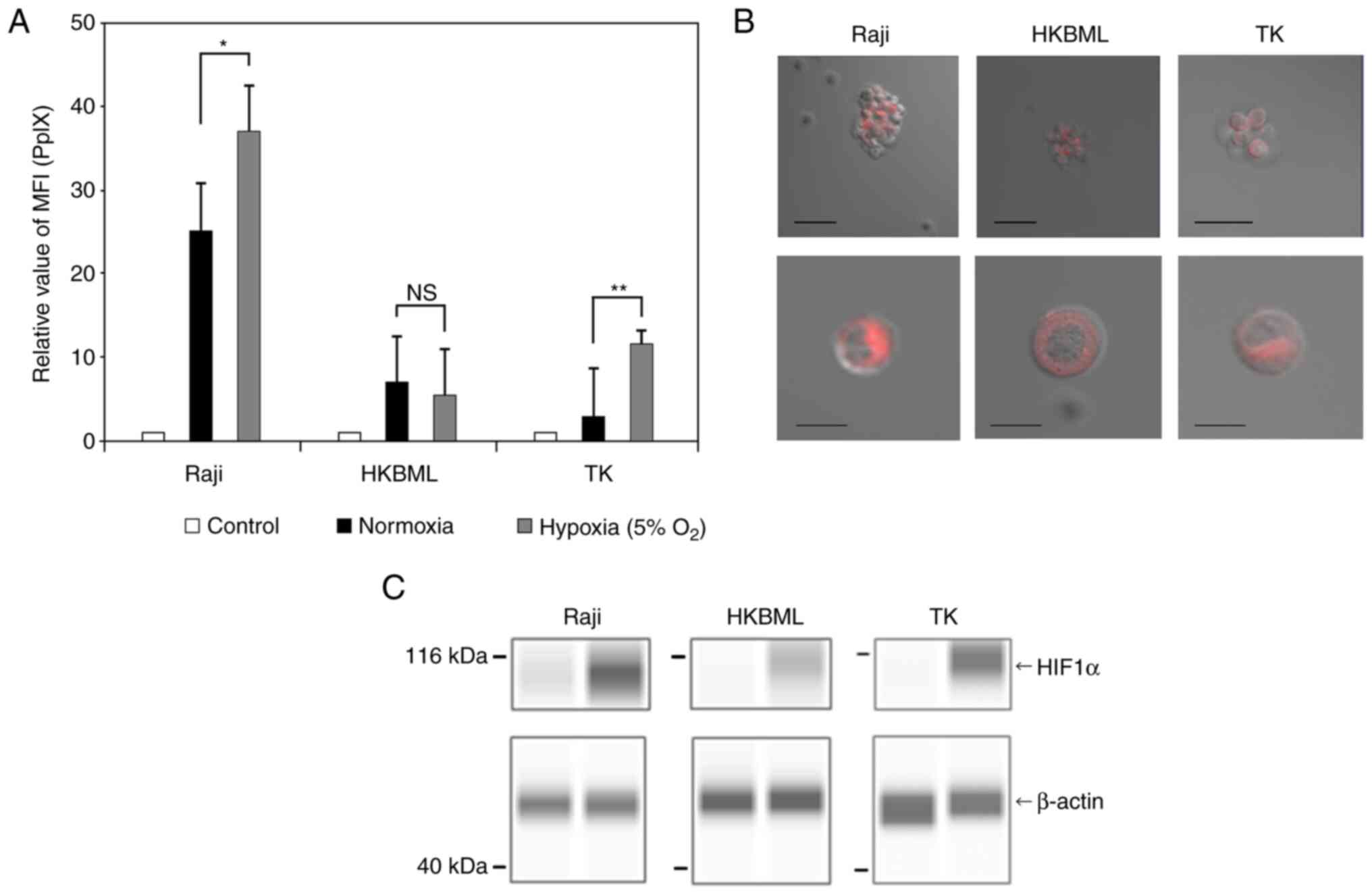 | Figure 1Intracellular accumulation and
visualization of 5-ALA-induced PpIX in lymphoma cells. The
intracellular PpIX fluorescence in lymphoma cells was determined
using flow cytometry. Lymphoma cells were treated with 0.3 mM 5-ALA
and incubated for 4 h. Hypoxic conditions were induced by placing
cells in a multi-gas incubator (5% O2) for 24 h before
the treatment with 5-ALA, and expression analysis of HIF1α. (A) The
relative MFI of 5-ALA-induced PpIX fluorescence was relative to
that of 5-ALA-untreated cells in each cell line, and the relative
MFI of 5-ALA-induced PpIX was compared under normoxic and hypoxic
conditions. The control group was treated without 5-ALA under
normoxic conditions. Columns, mean (n=8); error bars, SE. (B)
Visualization of intracellular PpIX in the lymphoma cells treated
with 1.0 mM 5-ALA for 4 h using a confocal laser scanning
microscope (excitation, 488 nm; emission, 630-nm long-pass filter).
Scale bar, 50 µm (top row) or 10 µm (bottom row). (C) Expression
analysis of HIF1α in lymphoma cells. HIF1α and β-actin were
detected using capillary western blotting, and the results are
shown as capillary western lane view images which were obtained
using the Simple Western system. *P<0.05;
**P<0.01. 5-ALA, 5-aminolevulinic acid; HIF1α,
hypoxia-induced factor 1α; MFI, median fluorescence intensity; NS,
not significant; PpIX, protoporphyrin IX. |
Evaluation of efficacy of radiodynamic
therapy with 5-ALA in lymphoma cells in vitro
We evaluated the efficacy of RDT with 5-ALA in
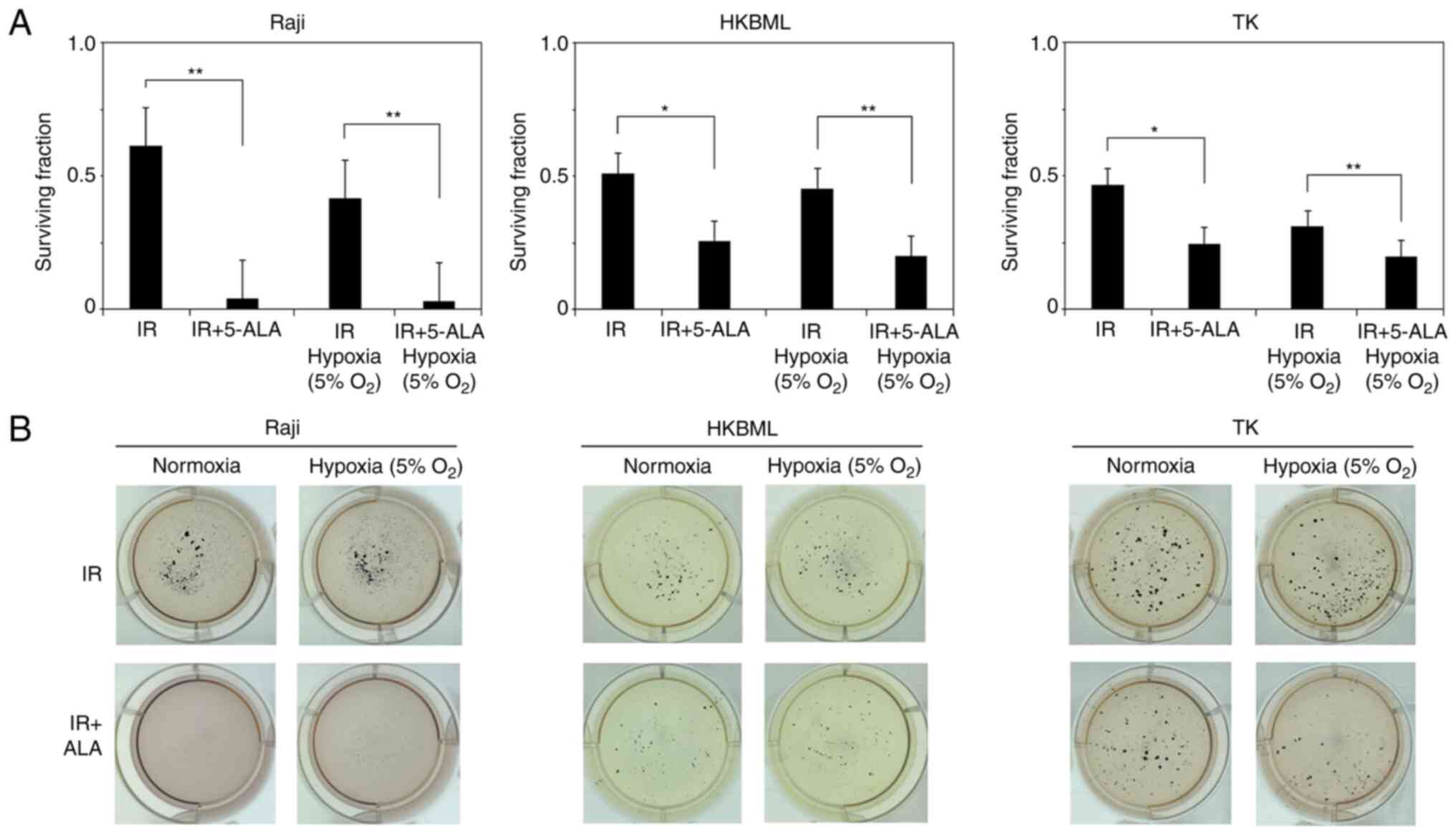
lymphoma cells using a colony-forming assay (Fig. 2). The relative survival rate of
cells in 5-ALA treated group (mean ± SE) were significantly
decreased compared with that in the 5-ALA-untreated group
(0.61±0.08 vs. 0.04±0.01, P<0.01 for the Raji line; 0.51±0.08
vs. 0.26±0.03, P<0.05 for the HKBML line; 0.47±0.07 vs.
0.25±0.02, P<0.05 for the TK line, 5-ALA-untreated group vs.
5-ALA treatment group, respectively) under the normoxic conditions.
Similarly, under the hypoxic conditions, the relative survival rate
of the cells in 5-ALA treatment group was significantly decreased
compared with that in the 5-ALA-untreated group (0.41±0.03 vs.
0.03±0.01, P<0.01 for the Raji line; 0.45±0.07 vs. 0.2±0.01,
P<0.01 for the HKBML line; 0.31±0.03 vs. 0.2±0.01, P<0.01 for
the TK line, 5-ALA-untreated group vs. 5-ALA treatment group,
respectively). Thus, 5-ALA had a radiodynamic effect on the
lymphoma cells under both normal and hypoxic conditions.
Temporal changes of ROS production
following RDT with 5-ALA in vitro
We evaluated temporal changes in intracellular ROS
production after IR in lymphoma cells under the normoxic and
hypoxic conditions (Fig. 3A). In
the cells without 5-ALA treatment, under the normoxic condition,
the relative MFI of DCFD fluorescence (mean ± SE) 12 h after IR was
significantly increased compared to that immediately after IR (0 h)
in each cell line (1.04±0.04 vs. 1.71±0.09, P<0.01 in the Raji
line; 0.93±0.03 vs. 1.13±0.04, P<0.01 in the HKBML line;
1.02±0.02 vs. 1.37±0.06, P<0.01 in the TK line; IR (0) vs. IR
(12), respectively) (Figs. 3B, and S2 and S3). Thus, we confirmed that
intracellular ROS production increased over time after IR without
5-ALA treatment in lymphoma cells, which is in agreement with
previous reports (10,13,22,23).
Meanwhile, the relative MFI of DCFD fluorescence (mean ± SE) in the
5-ALA treatment group was significantly higher than that in the
5-ALA-untreated group 12 h after IR in each cell (1.71±0.09 vs.
1.96±0.03, P<0.05 in the Raji line; 1.13±0.04 vs. 1.38±0.04,
P<0.01 in the HKBML line; 1.37±0.06 vs. 1.57±0.04, P<0.05 in
the TK line; 5-ALA-untreated group vs. 5-ALA treatment group,
respectively). Thus, under normoxic conditions, the ROS production
in the 5-ALA treatment group was significantly increased compared
to that in the 5-ALA-untreated group 12 h after IR (Figs. 3B, S2 and S3).
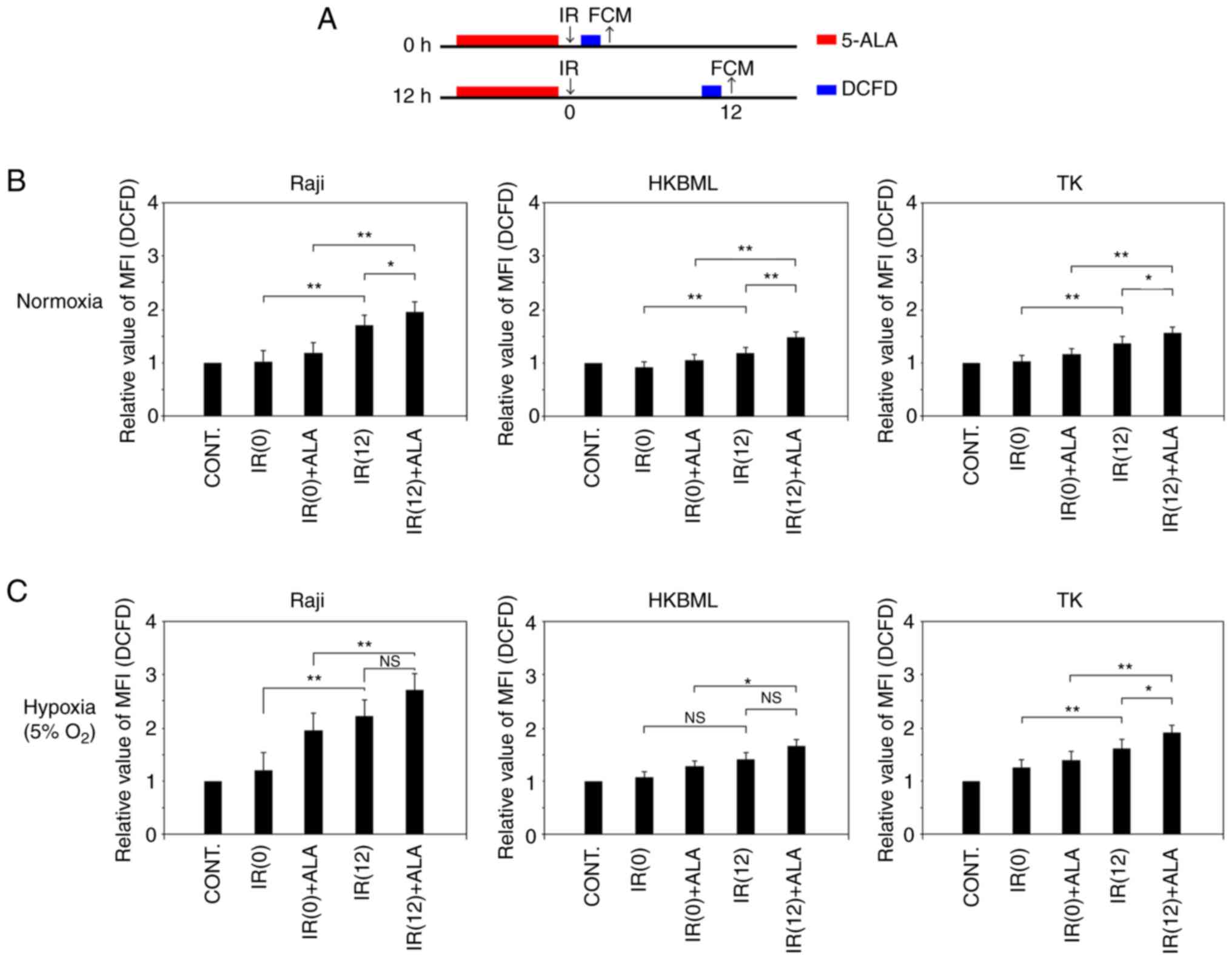 | Figure 3Temporal changes of ROS production in
radiodynamic therapy with 5-ALA in vitro. (A) Schedules of
the 5-ALA treatment, exposure to IR and detection of ROS
procedures. Cells were incubated with 0.3 mM 5-ALA for 4 h. Hypoxic
conditions were induced by placing cells in a multi-gas incubator
(5% O2) for 24 h before the treatment, and then, cells
were kept in hypoxic conditions during IR. Detection of ROS was
conducted using an oxidant-sensitive fluorescent probe (DCFD). The
control values were the intracellular ROS levels in the cells
without 5-ALA treatment or exposure to IR. The MFI of DCFD
fluorescence in the cells after IR exposure in relation to that of
the control was calculated in each cell line and then, the relative
MFI of DCFD fluorescence was compared in the cells immediately and
12 h after IR. Relative MFI of DCFD fluorescence (intracellular ROS
levels) after IR in lymphoma cells under (B) normoxic and (C)
hypoxic conditions. Columns, mean (n=5); error bars, SE.
*P<0.05; **P<0.01. 5-ALA,
5-aminolevulinic acid; CONT, control; DCFD, 2CM-H2DCFDA;
FCM, flow cytometry; IR, ionizing irradiation; MFI, median
fluorescence intensity; NS, not significant; ROS, reactive oxygen
species; IR (0), cells immediately after IR but without 5-ALA
treatment; IR (12), cells 12 h
after IR but without 5-ALA treatment; IR(0) + ALA, cells
immediately after IR with 5-ALA pretreatment; IR(12) + ALA, cells 12 h after IR with 5-ALA
pretreatment. |
In the 5-ALA-untreated cells under the hypoxic
condition, the relative MFI of DCFD fluorescence (mean ± SE) 12 h
after IR was significantly increased compared to that immediately
after IR (0 h) in the Raji and TK cell lines (1.22±0.07 vs.
2.22±0.13, P<0.01 in the Raji cell line; 1.26±0.05 vs.
1.63±0.07, P<0.01 in the cell TK line; IR (0) vs. IR (12), respectively) (Figs. 3C, S2A and C, and S3). There was no significant difference
in the relative MFI of DCFD fluorescence (mean ± SE) of the
5-ALA-untreated group between immediately after IR (0 h) and 12 h
after IR in the HKBML cell line (1.07±0.03 vs. 1.28±0.05,
P>0.05) (Figs. 3C, and S2B and S3). The relative MFI of DCFD
fluorescence (mean ± SE) in the 5-ALA treatment group was
significantly higher than that in the 5-ALA-untreated group 12 h
after IR in the TK cell line (1.63±0.07 vs. 1.91±0.09, P<0.05)
(Figs. 3C and S2C and S3). There was no significant difference
in the relative MFI of DCFD fluorescence (mean ± SE) 12 h after IR
between the 5-ALA-untreated group and the 5-ALA treatment group in
the Raji and HKBML cell lines (2.22±0.13 vs. 2.72±0.24, P>0.05
in the Raji cell line; 1.28±0.05 vs. 1.51±0.08, P>0.05 in the
HKBML cell line; 5-ALA-untreated vs. 5-ALA treatment group,
respectively) (Figs. 3C, S2A and B, and S3). Thus, under hypoxic conditions, only
TK cells exhibited significantly increased ROS production in the
5-ALA treatment group compared with the 5-ALA-untreated group 12 h
after IR.
Evaluation of mitochondrial density in
lymphoma cells
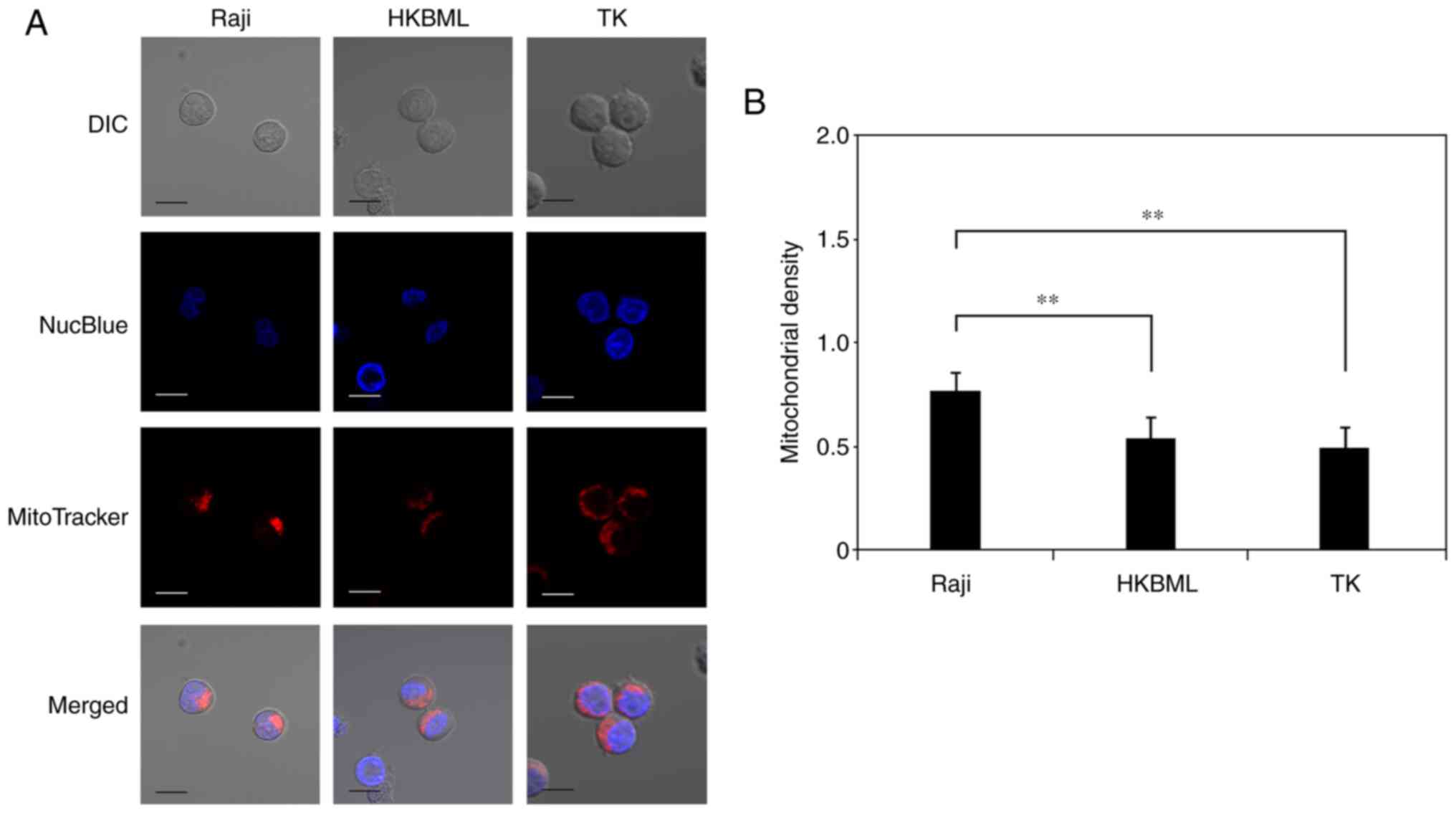
We evaluated the mitochondrial density in lymphoma
cells using microdensitometry with mitochondrial and nuclear
staining through confocal laser scanning microscopy (Fig. 4A). The mitochondrial density (mean
± SE) was 0.78±0.05 in the Raji line, 0.55±0.04 in the HKBML line,
and 0.5±0.06 in the TK line, respectively (Fig. 4B). The mitochondrial density in the
Raji cells was significantly higher than that in the HKBML and TK
cells (P<0.01) (Fig. 4B).
Discussion
We previously demonstrated that 5-ALA enhances
oxidative stress and delayed ROS production in mitochondria after
IR, and enhances cell death in proportion to 5-ALA-induced PpIX
accumulation in glioma cells in vitro (10,13).
In addition, we confirmed that 5-ALA enhanced the host antitumor
immune response and caused high inhibition of tumor growth in a rat
subcutaneous glioma model (11).
Similarly, the present study demonstrated that 5-ALA resulted in
high PpIX accumulation and enhanced cell death, with an increase in
delayed ROS production after IR in lymphoma cells. To the best of
our knowledge, this is the first study to investigate the
radiodynamic effect of 5-ALA on lymphoma cells.
Although many recent studies have demonstrated the
radiodynamic effect of 5-ALA in tumor cells (14-16,18),
the appropriate conditions for RDT with 5-ALA remain unclear. In a
prior study, we performed 5-ALA/RDT with 2 Gy/day for five
consecutive days (total 10 Gy) in a rat glioma subcutaneous model,
then confirmed the inhibition of tumor growth with strong
aggregation of Iba-1 positive macrophages within the tumor specimen
(11). Another recent study
performed 5-ALA/RDT with 2 Gy/day for five consecutive days x2
courses (total 20 Gy) in a mouse melanoma subcutaneous tumor model
and demonstrated the radiodynamic effect of 5-ALA via tumor growth
curve creation and microarray analysis of the gene expression
profiles in tumor specimens (14).
Another study also demonstrated the radiodynamic effect of 5-ALA at
4 Gy/day for three consecutive days (total 12 Gy) in a mouse
subcutaneous tumor model inoculated with prostate cancer cells
(16). That study reported
reductions in tumor volumes and mitotic activities by comparing the
pathological findings in a 5-ALA/RDT group and a control group. In
addition, they demonstrated that radiosensitization by 5-ALA was
not due to the photodynamic effect of light contamination by
examining subcutaneous tumors covered with high-performance black
masking tape (16). In contrast, a
recent study investigated the radiodynamic effect of 5-ALA using a
mouse brain tumor model inoculated with patient-derived
glioblastoma stem cells (24).
They performed 5-ALA/RDT at 2 Gy/day five times (3 times a week,
Monday/Wednesday/Friday; total 10 Gy) and evaluated tumor volume
using luciferin-based bioluminescence imaging (24). However, the bioluminescence of the
brain tumors in the 5-ALA/RDT group varied widely, and
consequently, they could not demonstrate the radiodynamic effect of
5-ALA in their mouse brain tumor model (24).
In terms of the oxidative stress in the mitochondria
of tumor cells, it may be important to increase ROS production over
the lethal level through different treatments (25). We confirmed that ROS production
after 5-ALA/RDT increased over time (within 12 h) in glioma and
lymphoma cell lines under the normoxic condition, including in the
present study (10,13). Meanwhile, a study utilizing
prostate cancer cells revealed that ROS production increased just
after 5-ALA/RDT and then gradually decreased over time (within 12
h) (26). Thus, the pattern of ROS
production after 5-ALA/RDT treatment varied in each cell line. Once
oxidative stress damages the mitochondria within tumor cells,
impaired mitochondria produce ROS via metabolic processes (22,23).
However, if these ROS do not reach a lethal level, the impaired
mitochondria may recover with time (more than 24 h), and then the
ROS production will gradually decrease (23). Consequently, the mitochondria may
regain normal function. Because the radiodynamic effect of 5-ALA
through single-dose IR is weak (12), the duration of each fraction of IR
may be important in RDT with 5-ALA. Taken together, we hypothesize
that IR exposure is needed before impaired mitochondria completely
regain their ability to produce ROS above lethal levels during
5-ALA/RDT. Namely, the duration of each fraction of IR is
recommended to be within 24 h following 5-ALA/RDT. Recently,
clinicians have considered the application of RDT with 5-ALA as an
adjuvant therapy for patients (27-29).
Thus, further studies are required to determine the appropriate
conditions of IR in 5-ALA/RDT.
Tumor hypoxia is associated with poor clinical
outcomes in many cancers, and hypoxic tumor cells are up to three
times more resistant to radiotherapy than normoxic tumor cells
owing to the absence of the oxygen enhancement effect (30,31).
Even very low levels of oxygen (approximately 2%) are sufficient to
yield an oxygen enhancement effect in radiotherapy (32), and many studies have defined oxygen
levels of 1-5% as hypoxic conditions in vitro (26,33,34).
First, we confirmed that the cell culture was unstable under
hypoxic conditions of 1% O2 in our experimental setting.
Thus, in the present study, we cultured cells in 5% O2
and confirmed the induction of HIF-1a in each cell line. 5-ALA
accumulated PpIX, and radiotherapy with 5-ALA clearly inhibited
cell proliferation compared to radiotherapy alone in colony
formation assay under hypoxic conditions in lymphoma cells. In the
present study, lymphoma cells revealed different survival rates in
5-ALA/RDT in each cell line, with some variation. 5-ALA is
metabolized and converted to heme in the mitochondria via the heme
synthesis in normal cells (5). In
tumor cells, the heme synthesis is stopped at the final step, and
PpIX is not converted to heme, which leads to accumulation of PpIX
in the mitochondria of tumor cells (5). Therefore, the accumulation of
5-ALA-induced PpIX is dependent on the cell metabolism in tumor
cells (5). In the present study,
lymphoma cells exhibited differences in 5-ALA-induced PpIX
accumulation under different oxygen culture conditions. In
addition, only TK cells exhibited enhancement of ROS production 12
h after IR in the 5-ALA-treated group under hypoxic conditions.
Although further studies evaluating the effect of 5-ALA on ROS
production after IR under hypoxic conditions are required, these
differences may depend on the amount of 5-ALA-induced PpIX
accumulation and mitochondrial density. A recent study demonstrated
that 5-ALA/RDT induced an increase in mitochondrial ROS production
and conversion of mitochondria-mediated tumor metabolism into
quiescent status via the inhibition of HIF-1, the mitochondrial
oxygen consumption rate (OCR), and the extracellular acidification
rate, and then it inhibited cancer stemness under hypoxic
conditions. Finally, 5-ALA helped prostate cancer cells overcome
the effects of hypoxia-induced radiation resistance (26). Although reoxygenation of the cell
culture cannot be completely excluded, we believe that 5-ALA can
induce a radiodynamic effect in lymphoma cells under hypoxic
conditions.
DLBCLs exhibit highly heterogeneous metabolic
features, such as BCR-dependent and OxPhos-DLBCLs (35). BCR-dependent subtype has greater
glycolytic flux, typical of the Warburg phenotype. Meanwhile,
OxPhos-subtype shows elevated electron transport chain activity,
ATP production, and fatty acid oxygenation. In addition, these
metabolic phenotypes are associated with a subtype selective
survival mechanism (35).
Considering the mechanism of the radiodynamic effect of 5-ALA,
5-ALA-induced PpIX is a key mediator, not 5-ALA itself. The cell
metabolism affects the conversion of 5-ALA into PpIX. In tumor
cells, 5-ALA treatment is associated with accumulation of PpIX in
the mitochondria due to the mitochondrial dysfunction, including
inactivation of heme converted enzyme (5). As previous studies demonstrated,
5-ALA-induced PpIX enhanced the antitumor effect induced by
radiotherapy in tumor cells (5,10-18).
In the present study, lymphoma cells showed differences in ROS
production after IR and 5-ALA-induced PpIX accumulation under
hypoxic conditions in each cell line. Accumulation of 5-ALA-induced
PpIX in Raji and TK cells but not HKBML cells under hypoxic
conditions was significantly increased compared with that under
normoxic conditions according to flow cytometric analysis. In
addition, only in TK cells, ROS production was increased 12 h after
IR in 5-ALA treated cells under hypoxic conditions. Taken together,
the differences in 5-ALA metabolism under different oxygen
conditions may affect the 5-ALA-induced PpIX accumulation and ROS
production after IR in lymphoma cells. A further study of the
metabolic changes within lymphoma cells in the surrounding
environment is needed. According to a recent clinical study using
physiologic magnetic resonance imaging, tumor microenvironment
mapping of patients demonstrated intertumoral heterogeneity factors
such as 65% glycolysis, 19% OxPhos, 9% hypoxia, and 7% necrosis in
PCNSL (36). Thus, recent
therapeutic strategies have been developed for each metabolic
target in DLBCL (37,38). Although further investigations are
required, 5-ALA/RDT may become a treatment option for PCNSL.
5-ALA has been widely used in clinical settings as a
live molecular marker for glioma and PCNSL (8,39),
and many experimental studies have demonstrated the radiodynamic
effect of 5-ALA in cancer cells. Nevertheless, the precise
mechanism of action of 5-ALA/RDT remains unclear. Recently, IR has
been shown to affect both the nucleus and mitochondria, which
includes activation of the DNA damage response and mitochondrial
signaling toward apoptosis (40).
Importantly, impaired mitochondria damaged by IR produce ROS
(mainly superoxide) via the metabolic process, then damage the rest
of the surrounding normal mitochondria, consequently propagating
oxidative stress within the cell, which is denoted as
‘intermitochondrial communication’ (41). This effect amplifies the oxidative
damage signal and further metabolic ROS production over time after
IR. In addition, a recent study demonstrated that mitochondria
damaged by IR activated an immune response in the nucleus (42).
PpIX enhances the production of ROS such as
superoxide, hydroxyl radicals, and singlet oxygen via water
radiolysis induced by IR (9), and
5-ALA accumulates PpIX in the mitochondria of cancer cells. In a
previous study, we confirmed that 5-ALA treatment after IR did not
enhance ROS production in glioma cells (13). In the present study, the Raji line
revealed a high accumulation of 5-ALA-induced PpIX and a high
5-ALA/RDT effect compared to the other cell lines. Therefore, we
propose that focal oxidative stress in the mitochondria is the
first step, and high accumulation of PpIX in the mitochondria is
important just before IR exposure in 5-ALA/RDT. In future studies,
oxidative stress in cancer cells after IR in 5-ALA/RDT should be
examined in detail.
Although the pattern of ROS production after IR
varies, we hypothesized that continuous oxidative stress above the
lethal level after IR in 5-ALA/RDT is important, which is in
agreement with another study (26). Namely, delayed ROS production via
effective ‘intermitochondrial communication’ may enhance the
therapeutic effects during 5-ALA/RDT. Thus, we speculated that
‘intermitochondrial communication’ was dependent on the
distance between each mitochondrion and examined the mitochondrial
density in lymphoma cells. In the present study, the Raji cells
showed significantly higher mitochondrial density values than the
other cells. As we described previously, we recently noticed the
importance of the application of 5-ALA/RDT for radiosensitive
malignant neoplasms. We, therefore, began this experimental in
vitro study using lymphoma cell lines. Although the mechanism
of 5-ALA/RDT is complicated, we believe that the mitochondria play
a main role in this process, and further investigation of the
mitochondrial response is required, including their roles in the
apoptotic signaling, metabolism, and dynamics of 5-ALA/RDT, and the
experimental tumor model in lymphoma.
This study has certain limitations. We confirmed the
radiodynamic effect of 5-ALA in lymphoma cells in vitro, but
not in an in vivo study. We have ever investigated the
radiodynamic effect of 5-ALA using a rat allogeneic subcutaneous
tumor model inoculated with 9L gliosarcoma, and confirmed
enhancement of the host antitumor immune response. It is possible
that, in an in vivo study of lymphoma, host immunity may
affect the results of 5-ALA/RDT.
In conclusion, 5-ALA has been widely used as a
fluorescent live marker for glioma and PCNSL in clinical settings.
Although PCNSL is radiosensitive, reduction of the radiation dose
is needed to avoid delayed radiation injury while maintaining
therapeutic effects, particularly in the elderly. 5-ALA accumulates
PpIX and induces a radiodynamic effect in lymphoma cells under both
normoxic and hypoxic conditions. Thus, 5-ALA/RDT is a potential
therapeutic option for PCNSL.
Supplementary Material
Representative flow cytometry plots
(upper and middle rows) and histograms (lower row) of
5-aminolevulinic acid-induced protoporphyrin IX fluorescence
intensity in lymphoma cells under the normoxic and hypoxic
conditions. The control group was treated without 5-ALA. The
percentage in each plot is the percentage of cells within the
indicated area. FS, forward scatter; SS, side scatter.
Representative flow cytometry plots of
DCFD fluorescence intensity after IR in lymphoma cells under
normoxic (upper row) and hypoxic (lower row) conditions. (A) Raji,
(B) HKBML and (C) TK cells. The control group was treated without
5-ALA and IR. The percentage in each plot is the percentage of
cells within the indicated area. 5-ALA, 5-aminolevulinic acid;
DCFD, 2CM-H2DCFDA; FS, forward scatter; IR, ionizing irradiation;
SS, side scatter; IR(0), cells immediately after IR but without
5-ALA treatment; IR(12), cells 12 h after IR but without 5-ALA
treatment; IR(0) + ALA, cells immediately after IR with 5-ALA
pretreatment; IR(12) + ALA, cells 12 h after IR with 5-ALA
pretreatment.
Representative histograms of flow
cytometry of 2CM-H2DCFDA fluorescence intensity after IR in
lymphoma cells under normoxic (upper row) and hypoxic (lower row)
conditions. IR, ionizing irradiation; IR(0), cells immediately
after IR but without 5-ALA treatment; IR(12), cells 12 h after IR
but without 5-ALA treatment; IR(0) + ALA, cells immediately after
IR with 5-ALA pretreatment; IR(12) + ALA, cells 12 h after IR with
5-ALA pretreatment.
Acknowledgements
Not applicable.
Funding
Funding: This work was supported by JSPS KAKENHI (grant no.
18K07307).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KS and JY drafted the final manuscript. KS, JY, KT
and RM made substantial contributions to the conception and design
of the study. KS, JY, KT and RM critically revised the manuscript
and provided constructive feedback. KS, KT and RM performed flow
cytometric analysis and interpreted the data. KS and JY performed
fluorescence cell imaging and interpreted the data. KS, JY and RM
confirm the authenticity of the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Han CH and Batchelor TT: Diagnosis and
management of primary central nervous system lymphoma. Cancer.
123:4314–4324. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Siegal T and Bairey O: Primary CNS
lymphoma in the elderly: The challenge. Acta Haematol. 141:138–145.
2019.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Ferreri AJ and Marturano E: Primary CNS
lymphoma. Best Pract Res Clin Haematol. 25:119–130. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Calimeri T, Steffanoni S, Gagliardi F,
Chiara A and Ferreri AJM: Erratum to ‘How we treat primary central
nervous system lymphoma’: [ESMO Open Volume 6, Issue 4, August
2021, 100213]. ESMO Open. 6(100326)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Ishizuka M, Abe F, Sano Y, Takahashi K,
Inoue K, Nakajima M, Kohda T, Komatsu N, Ogura S and Tanaka T:
Novel development of 5-aminolevurinic acid (ALA) in cancer
diagnoses and therapy. Int Immunopharmacol. 11:358–365.
2011.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Stummer W, Pichlmeier U, Meinel T,
Wiestler OD, Zanella F and Reulen HJ: ALA-Glioma Study group.
Fluorescence-guided surgery with 5-aminolevulinic acid for
resection of malignant glioma: A randomised controlled multicentre
phase III trial. Lancet Oncol. 7:392–401. 2006.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Stummer W, Holling M, Bendok BR, Vogelbaum
MA, Cox A, Renfrow SL, Widhalm G, Ezrin A, DeSena S, Sackman ML and
Wyse JW: The NXDC-MEN-301 Study on 5-ALA for meningiomas surgery:
An innovative study design for the assessing the benefit of
intra-operative fluorescence imaging. Brain Sci.
12(1044)2022.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Kiesel B, Millesi M, Woehrer A, Furtner J,
Bavand A, Roetzer T, Mischkulnig M, Wolfsberger S, Preusser M,
Knosp E and Widhalm G: 5-ALA-induced fluorescence as a marker for
diagnostic tissue in stereotactic biopsies of intracranial
lymphomas: Experience in 41 patients. Neurosurg Focus.
44(E7)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Takahashi J and Misawa M: Characterization
of reactive oxygen species generated by protoporphyrin IX under
X-ray irradiation. Radiat Phys Chem. 78:889–898. 2009.
|
|
10
|
Ueta K, Yamamoto J, Tanaka T, Nakano Y,
Kitagawa T and Nishizawa S: 5-Aminolevulinic acid enhances
mitochondrial stress upon ionizing irradiation exposure and
increases delayed production of reactive oxygen species and cell
death in glioma cells. Int J Mol Med. 39:387–398. 2017.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Yamamoto J, Ogura S, Shimajiri S, Nakano
Y, Akiba D, Kitagawa T, Ueta K, Tanaka T and Nishizawa S:
5-aminolevulinic acid-induced protoporphyrin IX with multi-dose
ionizing irradiation enhances host antitumor response and strongly
inhibits tumor growth in experimental glioma in vivo. Mol Med Rep.
11:1813–1819. 2015.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yamamoto J, Ogura S, Tanaka T, Kitagawa T,
Nakano Y, Saito T, Takahashi M, Akiba D and Nishizawa S:
Radiosensitizing effect of 5-aminolevulinic acid-induced
protoporphyrin IX in glioma cells in vitro. Oncol Rep.
27:1748–1752. 2012.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kitagawa T, Yamamoto J, Tanaka T, Nakano
Y, Akiba D, Ueta K and Nishizawa S: 5-Aminolevulinic acid strongly
enhances delayed intracellular production of reactive oxygen
species (ROS) generated by ionizing irradiation: Quantitative
analyses and visualization of intracellular ROS production in
glioma cells in vitro. Oncol Rep. 33:583–590. 2015.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Takahashi J, Murakami M, Mori T and
Iwahashi H: Verification of radiodynamic therapy by medical linear
accelerator using a mouse melanoma tumor model. Sci Rep.
8(2728)2018.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yamada K, Murayama Y, Kamada Y, Arita T,
Kosuga T, Konishi H, Morimura R, Shiozaki A, Kuriu Y, Ikoma H, et
al: Radiosensitizing effect of 5-aminolevulinic acid in colorectal
cancer in vitro and in vivo. Oncol Lett. 17:5132–5138.
2019.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Miyake M, Tanaka N, Hori S, Ohnishi S,
Takahashi H, Fujii T, Owari T, Ohnishi K, Iida K, Morizawa Y, et
al: Dual benefit of supplementary oral 5-aminolevulinic acid to
pelvic radiotherapy in a syngenic prostate cancer model. Prostate.
79:340–351. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Kaneko T, Tominaga M, Kouzaki R, Hanyu A,
Ueshima K, Yamada H, Suga M, Yamashita T, Okimoto T and Uto Y:
Radiosensitizing effect of 5-aminolevulinic acid and protoporphyrin
IX on Carbon-ion beam irradiation. Anticancer Res. 38:4313–4317.
2018.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang DM, Cvetkovic D, Chen L and Ma CC:
Therapeutic effects of in-vivo radiodynamic therapy (RDT) for lung
cancer treatment: A combination of 15MV photons and
5-aminolevulinic acid (5-ALA). Biomed Phys Eng Express.
8(1088/2057-1976/ac9b5c)2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang J, Valdez A and Chen Y: Evaluation of
automated Wes system as an analytical and characterization tool to
support monoclonal antibody drug product development. J Pharm
Biomed Anal. 139:263–268. 2017.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Baddela VS, Sharma A, Michaelis M and
Vanselow J: HIF1 driven transcriptional activity regulates
steroidogenesis and proliferation of bovine granulosa cells. Sci
Rep. 10(3906)2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Prall F, Maletzki C and Linnebacher M:
Microdensitometry of osteopontin as an immunohistochemical
prognostic biomarker in colorectal carcinoma tissue microarrays:
Potential and limitations of the method in ‘biomarker pathology’.
Histopathology. 61:823–832. 2012.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yamamori T, Yasui H, Yamazumi M, Wada Y,
Nakamura Y, Nakamura H and Inanami O: Ionizing radiation induces
mitochondrial reactive oxygen species production accompanied by
upregulation of mitochondrial electron transport chain function and
mitochondrial content under control of the cell cycle checkpoint.
Free Radic Biol Med. 53:260–270. 2012.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Saenko Y, Cieslar-Pobuda A, Skonieczna M
and Rzeszowska-Wolny J: Changes of reactive oxygen and nitrogen
species and mitochondrial functioning in human K562 and HL60 cells
exposed to ionizing radiation. Radiat Res. 180:360–366.
2013.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Dupin C, Sutter J, Amintas S, Derieppe MA,
Lalanne M, Coulibaly S, Guyon J, Daubon T, Boutin J, Blouin JM, et
al: An orthotopic model of glioblastoma is resistant to
radiodynamic therapy with 5-AminoLevulinic acid. Cancers (Basel).
14(4244)2022.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Galadari S, Rahman A, Pallichankandy S and
Thayyullathil F: Reactive oxygen species and cancer paradox: To
promote or to suppress? Free Radic Biol Med. 104:144–164.
2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Owari T, Tanaka N, Nakai Y, Miyake M, Anai
S, Kishi S, Mori S, Fujiwara-Tani R, Hojo Y, Mori T, et al:
5-Aminolevulinic acid overcomes hypoxia-induced radiation
resistance by enhancing mitochondrial reactive oxygen species
production in prostate cancer cells. Br J Cancer. 127:350–363.
2022.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Nordmann NJ and Michael AP:
5-Aminolevulinic acid radiodynamic therapy for treatment of
high-grade gliomas: A systematic review. Clin Neurol Neurosurg.
201(106430)2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Krivoshapkin A, Gaytan A, Abdullaev O,
Salim N, Sergeev G, Marmazeev I, Cesnulis E, Killeen T, Tyuryn V,
Kiselev R, et al: Prospective comparative study of intraoperative
balloon electronic brachytherapy versus resection with
multidisciplinary adjuvant therapy for recurrent glioblastoma. Surg
Neurol Int. 12(517)2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Pepper NB, Stummer W and Eich HT: The use
of radiosensitizing agents in the therapy of glioblastoma
multiforme-a comprehensive review. Strahlenther Onkol. 198:507–526.
2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Higgins GS, O'Cathail SM, Muschel RJ and
McKenna WG: Drug radiotherapy combinations: Review of previous
failures and reasons for future optimism. Cancer Treat Rev.
41:105–113. 2015.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Dhani N, Fyles A, Hedley D and Milosevic
M: The clinical significance of hypoxia in human cancers. Semin
Nucl Med. 45:110–121. 2015.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Ashton TM, McKenna WG, Kunz-Schughart LA
and Higgins GS: Oxidative phosphorylation as an emerging target in
cancer therapy. Clin Cancer Res. 24:2482–2490. 2018.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Ihata T, Nonoguchi N, Fujishiro T, Omura
N, Kawabata S, Kajimoto Y and Wanibuchi M: The effect of hypoxia on
photodynamic therapy with 5-aminolevulinic acid in malignant
gliomas. Photodiagnosis Photodyn Ther. 40(103056)2022.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Castillo CA, Leon D, Ruiz MA, Albasanz JL
and Martin M: Modulation of adenosine A1 and A2A receptors in C6
glioma cells during hypoxia: Involvement of endogenous adenosine. J
Neurochem. 105:2315–2329. 2008.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Caro P, Kishan AU, Norberg E, Stanley IA,
Chapuy B, Ficarro SB, Polak K, Tondera D, Gounarides J, Yin H, et
al: Metabolic signatures uncover distinct targets in molecular
subsets of diffuse large B cell lymphoma. Cancer Cell. 22:547–560.
2012.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Stadlbauer A, Marhold F, Oberndorfer S,
Heinz G, Zimmermann M, Buchfelder M, Heynold E and Kinfe TM:
Metabolic tumor microenvironment characterization of contrast
enhancing brain tumors using physiologic MRI. Metabolites.
11(668)2021.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Norberg E, Lako A, Chen PH, Stanley IA,
Zhou F, Ficarro SB, Chapuy B, Chen L, Rodig S, Shin D, et al:
Differential contribution of the mitochondrial translation pathway
to the survival of diffuse large B-cell lymphoma subsets. Cell
Death Differ. 24:251–262. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Noble RA, Thomas H, Zhao Y, Herendi L,
Howarth R, Dragoni I, Keun HC, Vellano CP, Marszalek JR and Wedge
SR: Simultaneous targeting of glycolysis and oxidative
phosphorylation as a therapeutic strategy to treat diffuse large
B-cell lymphoma. Br J Cancer. 127:937–947. 2022.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Farrell C, Shi W, Bodman A and Olson JJ:
Congress of neurological surgeons systematic review and
evidence-based guidelines update on the role of emerging
developments in the management of newly diagnosed glioblastoma. J
Neurooncol. 150:269–359. 2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Averbeck D and Rodriguez-Lafrasse C: Role
of mitochondria in radiation responses: Epigenetic, metabolic, and
signaling impacts. Int J Mol Sci. 22(11047)2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Kam WW and Banati RB: Effects of ionizing
radiation on mitochondria. Free Radic Biol Med. 65:607–619.
2013.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Tigano M, Vargas DC, Tremblay-Belzile S,
Fu Y and Sfeir A: Nuclear sensing of breaks in mitochondrial DNA
enhances immune surveillance. Nature. 591:477–481. 2021.PubMed/NCBI View Article : Google Scholar
|