Introduction
Sepsis is an infection-induced systemic inflammatory
response syndrome accompanied by multiple organ injury and failure
(1). Timely treatment is critical
for sepsis; otherwise, it can develop into severe systemic
inflammation and eventually result in multiple organ injury and
dysfunction (2). A previous
extensive study revealed that activation of NOD-like receptors,
particularly the NLR family pyrin domain containing 3 (NLRP3)
inflammasome, plays an important role in lipopolysaccharide-induced
systemic inflammation. Additionally, activation of the NLRP3
inflammasome and its downstream inflammatory factors, including
interleukin (IL)-1β and IL-18, is involved in caspase-dependent
pyroptosis (3). Pyroptosis is a
type of programmed cell death that is associated with inflammation.
It is morphologically characterised by apoptosis and necrosis and
has also been identified as a major cause of sepsis-induced tissue
damage in previous research (4).
Therefore, appropriate suppression of pyroptosis-related proteins
may be a possible therapeutic strategy for sepsis and
sepsis-induced organ injury.
Neutrophils, which are the most abundant cells in
the immune system, are responsible for maintaining the equilibrium
of the body through repeated programmed cell death. Neutrophils
arrive at the site of infection and release a considerable number
of inflammatory cytokines and reactive oxygen species (ROS) during
sepsis, which exacerbates the inflammatory response (5). With the development of sepsis,
multiple organs, such as the heart, lung, kidney and spleen, are
infiltrated with a large number of activated neutrophils that
secrete a variety of enzymes and inflammatory mediators that are
partly dependent on pyroptosis in neutrophils and further
accelerate organ injury (6).
Hence, the regulation of pyroptosis in neutrophils may play an
important role in sepsis treatment.
The NLRP3 inflammasome is a multiprotein complex
that responds to a wide range of danger signals from different
sources, including microorganisms (7). Caspase-1 self-shearing and activation
are aided by the formation of the NLRP3 inflammasome. The
maturation of proinflammatory cytokines such as IL-1β and IL-18 is
reduced by activated caspase-1(8).
Furthermore, activated caspase-1 is associated with cleaved
gasdermin D (GSDMD). The cleaved N-terminal domain of GSDMD
translocates to the cell membrane and forms pores, which facilitate
the release of inflammatory cytokines and trigger the classical
signals of pyroptosis in the circulating blood (9,10).
Previous studies have reported that the inhibition or activation of
NLRP3 assembly or activation effectively improves sepsis (11,12).
In the present study, the effect of MCC950 on relieving LPS-induced
inflammation was investigated, and it was also determined whether
the effect of MCC950 was mediated by inhibiting pyroptosis in
neutrophils.
Materials and methods
Animals
A total of 60 male C57BL/6J mice (8-10 weeks old;
weight, 25-30 g) were purchased from the Beijing Weitong Lihua
Animal Technology Co., Ltd. and fed in a specific-pathogen-free
environment with a controlled temperature (22˚C) and relative
humidity of 40-70%. The physiological characteristics of female
mice are affected by periodic endocrine changes, such as estrus
cycle, pregnancy and delivery, which may cause significant
variations in their physiological indicators, and thus were not
selected for the present study. Food and water were available ad
libitum, and a 12-h light/dark cycle was maintained. All
experiments were approved by the Ethics Committee of Yangzhou
University (Yangzhou, China) (approval no. 202201001). All authors
have read the ARRIVE guidelines, and the study was performed
according to the ARRIVE guidelines. All mice were anaesthetised
with 2% and sacrificed with 5% isoflurane.
Mouse model of systemic
inflammation
The septic mouse model was established by
administering 10 mg/kg LPS (O55:B5; Sigma-Aldrich; Merck KGaA) by
intraperitoneal (i.p.) injection. The mice were randomised into the
following three groups: i) In the control group (CON), the mice
received a volume of sterile saline equal to the volume of LPS; ii)
in the LPS-induced inflammatory group (LPS), the mice received 10
mg/kg LPS via i.p. injection; and iii) in the MCC950 treatment
group (LPS + MCC950), the mice were treated with 10 mg/kg MCC950
twice: 6 h pre- and post-LPS. The mice were sacrificed 24 h after
LPS injection. Each group contained 10 mice.
Serum sample preparation
The mice were sacrificed via isoflurane, and blood
samples were collected from an ocular vein. The serum was separated
by centrifugation at 1,500 x g for 15 min (4˚C), and the upper
layer of the serum was harvested and stored at -80˚C for further
analysis. Blood urea nitrogen (BUN) (Elabscience Biotechnology,
Inc.; cat. no. E-BC-K329-S) and creatinine (Cre) (Elabscience
Biotechnology, Inc.; cat. no. E-BC-K188-M), which are biomarkers of
kidney toxicity, as well as alanine transaminase (ALT) (Elabscience
Biotechnology, Inc.; cat. no. E-BC-K235-S) and aspartate
aminotransferase (AST) (Elabscience Biotechnology, Inc.; cat. no.
E-BC-K236-M), which are indicators of liver function, were analysed
using respective kits based on colorimetric principles according to
the manufacturer's instructions.
Neutrophil isolation
According to the manufacturer's instructions,
neutrophil isolation was performed using the MACS Neutrophil
Isolation Kit (Miltenyi Biotec, Inc.; cat. no. 130-097-658).
Briefly, 2 ml of buffer A was added to a bottle of MACSxpress whole
blood cell isolation cocktail and gently pipetted. Buffer B was
then mixed with the cocktail. Subsequently, 1 ml of isolation mix
was merged with anticoagulated whole blood and incubated for 5 min
at room temperature with gentle shaking. The tube was then placed
in a magnetic grate for 15 min to separate the labelled cells. The
supernatant was collected, and the residual erythrocytes were
removed by erythrocyte lysis buffer (Wuhan Servicebio Technology
Co., Ltd.; cat. no. G2015-500ML).
Flow cytometry
The neutrophils were collected and stained with
Ly6G-FITC (Biolegend, Inc.; cat. no. 127605) to determine the
isolation efficiency. Briefly, the isolated cells were suspended in
diluted Ly6G-FITC staining solution at a concentration of 1
million/ml. After 30 min of incubation at 37˚C, the cells were
washed with PBS supplemented with 0.5% FBS three times and detected
by flow cytometry to define the purity and cell activity. The
oxidative stress levels of isolated cells were analysed using an
ROS assay kit (Beyotime Institute of Biotechnology; cat. no.
S0033S) according to the manufacturer's instructions. To analyse
cell necrosis, the isolated cells were stained with Annexin V/PI
using the Annexin V/PI staining kit (Beyotime Institute of
Biotechnology; cat. no. C1062S) and incubated for 10-20 min at room
temperature in the dark. Data were collected on a multicolor flow
cytometer (BD Fortessa; BD Biosciences) and analyzed with FlowJo
software (version 10.8; FlowJo LLC).
Histopathology analysis
Lung, liver, kidney, spleen, and heart tissues were
collected in 4% PFA (24 h at room temperature) and embedded in
paraffin. The tissues were cut into 5-µm thick sections and stained
with haematoxylin and eosin (H&E). The sections were observed
and photographed using a light microscope (Olympus
Corporation).
Enzyme-linked immunosorbent assay
(ELISA)
According to the manufacturer's instructions, the
concentrations of IL-1β, IL-6, IL-18, and TNF-α were determined
using ELISA kits (Mlbio; Shanghai Enzyme-linked Biotechnology Co.,
Ltd.; cat. nos. ml098416, IC50325-1, ml002294 and mIC50536-1,
respectively) The values were measured using a Tecan Spark plate
reader (Tecan Group, Ltd.).
Assessment of survival rates
For survival analyses, 30 male mice were divided
into three groups as aforementioned. The concentration of LPS was
changed to 15 mg/kg. Following the injection of LPS, the number of
mice that succumbed was recorded every 12 h for 72 h. The survival
rates were analysed using ImageJ software (v1.8.0; National
Institutes of Health). At the endpoint, all mice were euthanised
with isoflurane (2% isoflurane was used to induce anesthesia and 5%
was used for euthanasia, causing the mice to lose consciousness
quickly). After confirming that the mice had no movement, were not
breathing, and their pupils were dilated, the anesthesia machine
was closed, and then the mice were observed for another 2 min to
confirm cardiac arrest and death. If the mice lost >10% of their
body weight or ate <50% of their normal amount in 24 h, or were
unable to eat on their own, or were either excessively weak and/or
had hypothermia, the mice were euthanised as these were considered
as humane endpoints.
Western blotting
For western blotting, the isolated neutrophils were
lysed in RIPA lysis buffer (Beyotime Institute of Biotechnology)
supplemented with 1% protease and phosphatase inhibitor cocktail
(MedChemExpress) on ice for 15 min and centrifuged at 12,000 x g
for 15 min at 4˚C. The supernatants were collected for western
blotting. The protein concentration was determined using a BCA kit
(Beyotime Institute of Biotechnology). The proteins (15 µg/sample)
in each sample were separated using precast gels (Beyotime
Institute of Biotechnology) and then transferred to PVDF membranes
(Merck KGaA). The membranes were blocked with 5% BSA buffer (Wuhan
Servicebio Technology Co., Ltd.) for 1 h at room temperature and
then incubated with primary antibodies targeting full-length GSDMD,
cleaved GSDMD, IL-1β and IL-18 (all 1:1,000; ABclonal Biotech Co.,
Ltd.; cat. nos. A18281, A22523, A16288 and A16737, respectively),
NLRP3, caspase-1 and cleaved caspase-1 (all 1:1,000; Cell Signaling
Technology, Inc.; cat. nos. 15101, 24232S and 89332, respectively),
apoptosis-associated speck-like protein containing CARD (ASC)
(1:1,000; HUABIO; cat. no. ER6274) and β-actin (1:2,000; ABclonal
Biotech Co., Ltd.; cat. no. AC006) After overnight incubation at
4˚C, the blots were washed with TBST (TBS buffer with 1% Tween-20)
three times and then incubated with specific secondary antibodies
(goat anti-rabbit IgG-HRP; 1:2,000; cat. no. HA1001; and goat
anti-mouse IgG-HRP; 1:2,000; cat. no. HA1006; both HUABIO) for 2 h
at room temperature. Finally, the bands were visualised using an
enhanced chemiluminescence reagent (Bio-Rad Laboratories, Inc.).
The greyscale intensities of the proteins were analysed using
ImageJ software (v1.8.0; National Institutes of Health).
Statistical analysis
The results are presented as the means ± standard
deviations. All experiments were performed independently at least
three times. For mouse experiments, no specific blinding method was
used, but the mice in each sample group were selected randomly.
Statistical significance between groups was determined by one-way
analysis of variance (ANOVA) and Tukey's post hoc test. The
Kaplan-Meier method with log-rank test was used to analyze the
survival probabilities of the mice. P<0.05 was considered to
indicate a statistically significant difference. GraphPad Prism 8
software (Dotmatics) was used for analysis.
Results
MCC950 decreases the levels of
inflammatory cytokines in an LPS-induced inflammation mouse
model
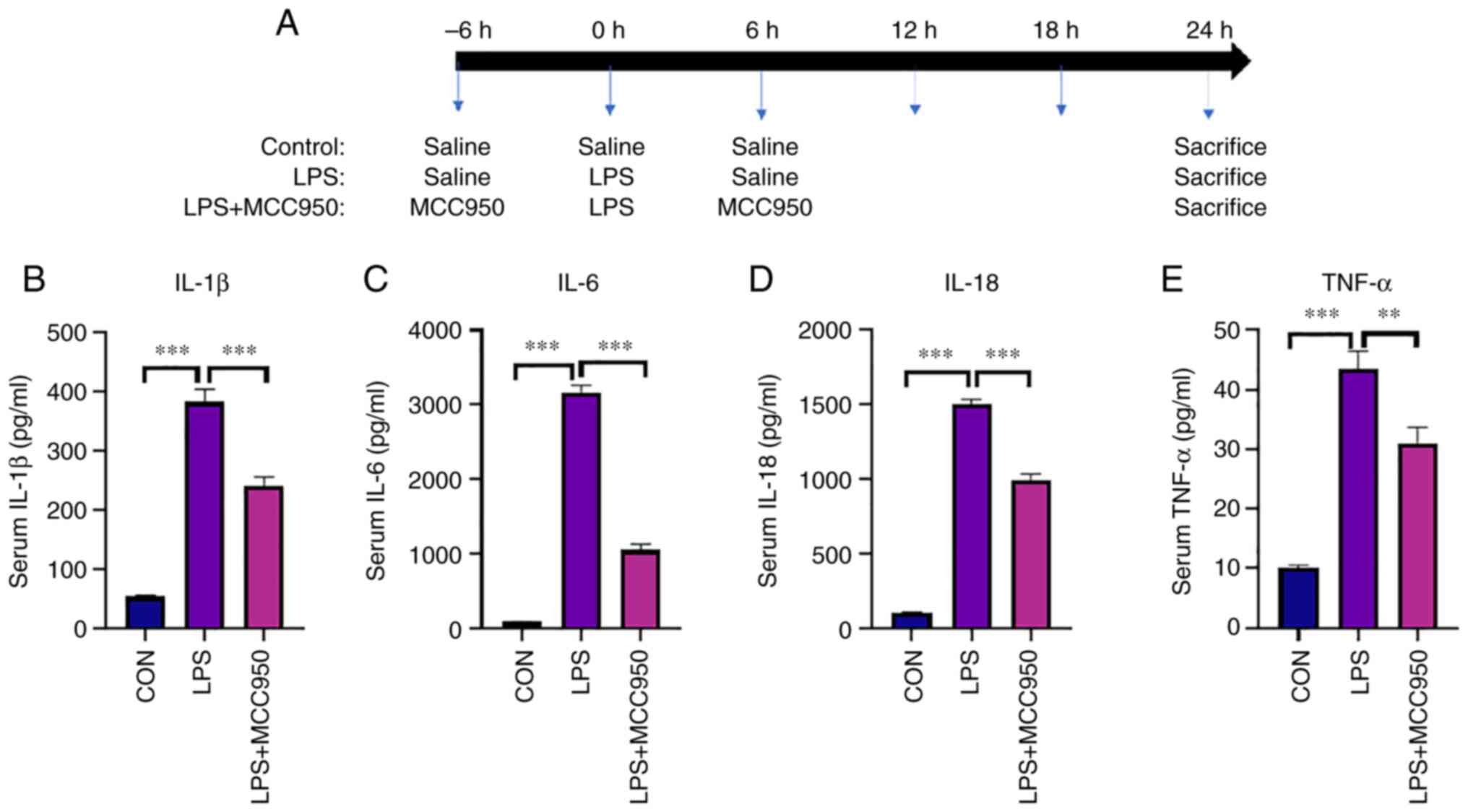
The mice were randomly divided into three groups.
The experimental process and the treatments are shown in Fig. 1A. A total of 24 h after LPS
injection, the mice were sacrificed, and blood was collected from
the inferior vena cava (~1.0 ml/mouse). The results revealed that
LPS markedly induced an inflammatory response in mice, including
the excessive secretion of inflammatory cytokines in the blood.
Compared with the control group (CON), the levels of IL-1β, IL-6,
IL-18 and TNF-α (Fig. 1B-E) in the
blood were upregulated in the LPS group. In the MCC950 + LPS group,
the levels of these cytokines were lower than those in the LPS
group (Fig. 1B-E).
MCC950 alleviates LPS-induced multiple
organ damage
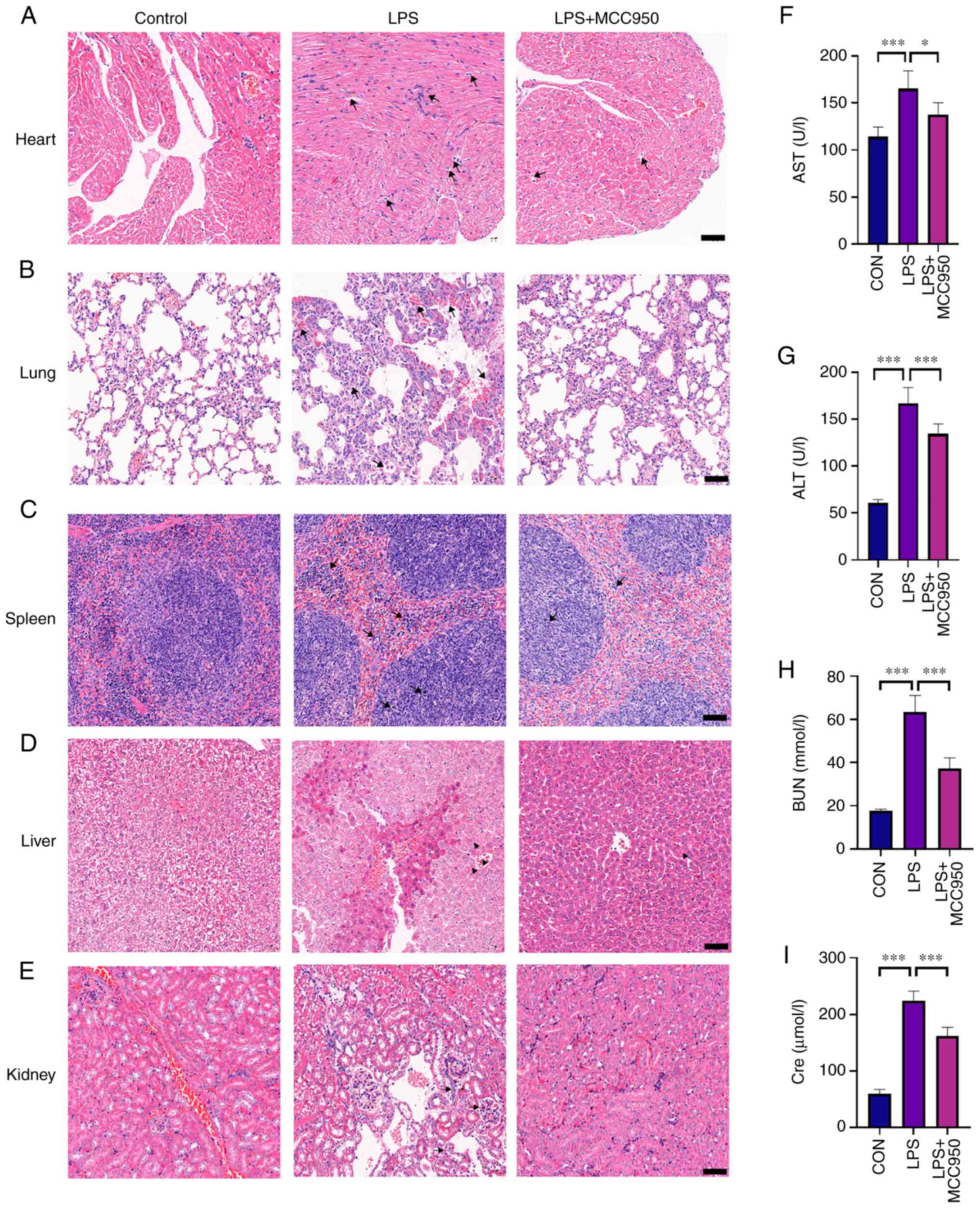
Organs such as the heart, liver, spleen, lung and
kidney were collected, embedded in paraffin, and stained with
H&E after LPS injection and MCC950 therapy. Histological images
revealed that LPS caused multiple organ damage.
As revealed in Fig.
2A, typical fibre bundles in the control group were neatly
organised and free of oedema in the interstitial space. In addition
to interstitial oedema, the fibre bundles in septic mouse cardiac
tissues 24 h after LPS administration were organised loosely,
fragmented and even disintegrated. In addition, some cardiomyocytes
deteriorated and dissolved, while necrosis and moderate
inflammatory cell infiltration were observed. Inflammatory cell
infiltration and fibre bundles were normal after MCC950
treatment.
The histological images in Fig. 2B depict the lung damage that
occurred during sepsis. The lung tissues were intact and clear, and
the cells were neatly distributed without oedema or indications of
harm in the control group. LPS injection significantly increased
inflammatory cell infiltration, interstitial oedema, and phagocyte
infiltration in the alveolar space. MCC950 decreased inflammatory
cell infiltration and interstitial oedema.
As revealed in Fig.
2C, lymphoid tissue proliferation was not significant in the
control group, and the red pulp was mainly composed of lymphocytes.
In the sepsis group, lymphoid hyperplasia and apoptosis were
observed in the white pulp of the spleen. In addition, acute and
chronic inflammatory cell infiltration was observed in the red pulp
of mice with sepsis. MCC950 administration alleviated LPS-induced
inflammatory infiltration, lymphoid hyperplasia, and cell
apoptosis.
As shown Fig. 2D,
the liver lobules of mice in the control group were intact and
clear, the cells were neatly distributed and free of oedema, and
the liver stripes were regular and clear. In the LPS-treated group,
the liver lobules were damaged, the liver cells were swollen with
spot necrosis, and the intracellular space was increased. When
LPS-treated mice were treated with MCC950, morphological analysis
revealed that liver tissue damage was significantly decreased. In
addition, LPS-treated mice exhibited a high level of neutrophil
infiltration, whereas MCC950 treatment reduced neutrophil
infiltration. ALT and AST levels, which are indicators of liver
function, were also assessed. The results demonstrated that MCC950
reduced the levels of ALT and AST, which were increased by LPS
(Fig. 1F and G).
As revealed in Fig.
2E, the kidney tissues were clear and intact, and the cells
were arranged neatly without oedema or injury in the control group.
LPS-treated mice exhibited deeply damaged kidney lobules, and the
cells were swollen with increased intracellular spaces. In
addition, neutrophil infiltration was observed in the kidney
tissues in the LPS group. MCC950 treatment alleviated the damage to
the kidney induced by LPS, as indicated by clearer nephrons, less
vacuolization, and less infiltration. In addition, the
concentrations of BUN and Cre, which are markers of kidney damage,
were detected. The results showed that LPS increased Cre and BUN
concentrations in serum, which were significantly decreased by
MCC950 treatment (Fig. 1H and
I).
MCC950 alleviates the damage to
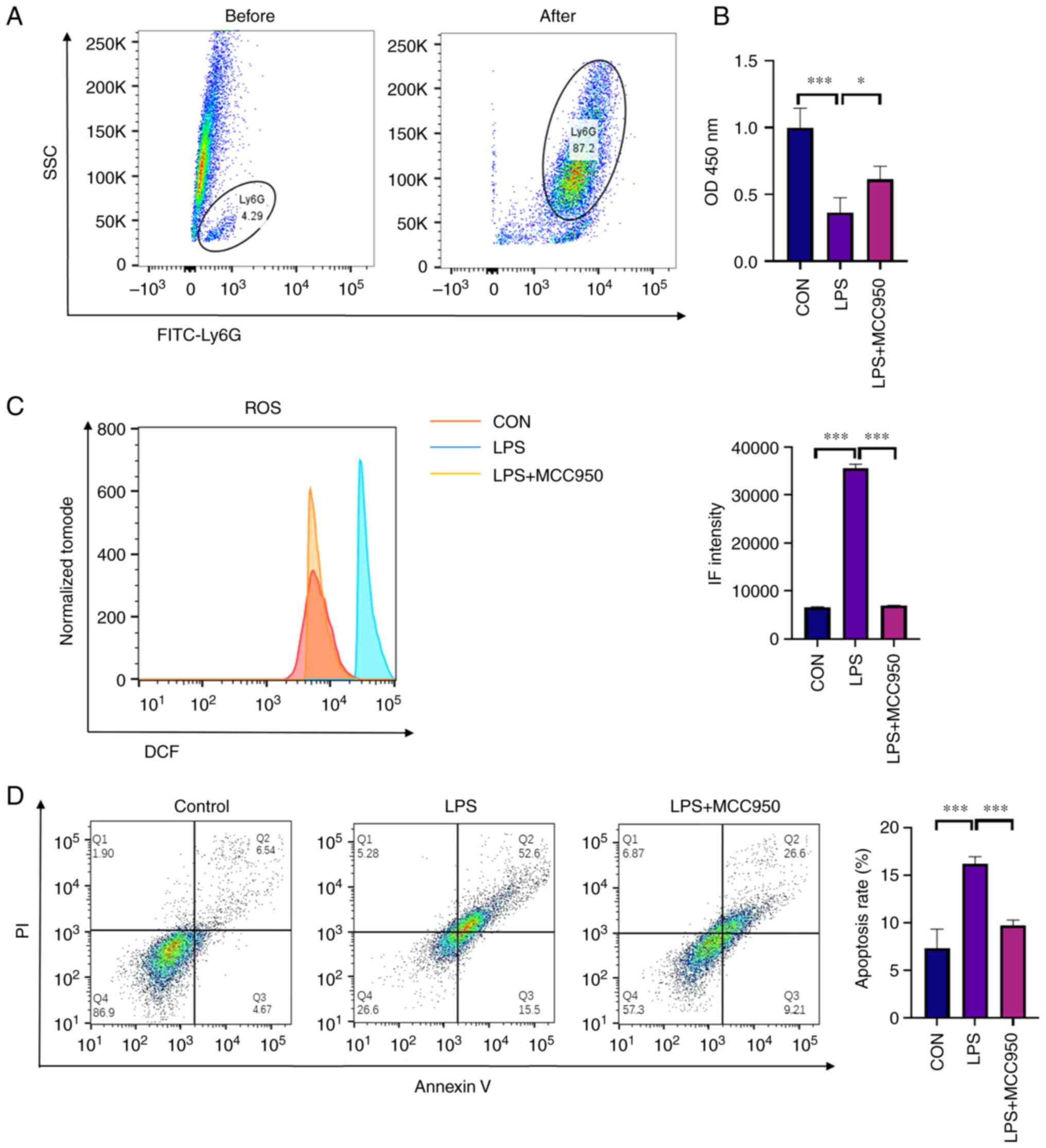
neutrophils in peripheral blood induced by sepsis
To explore the mechanism of MCC950 in the treatment
of LPS-induced inflammation, further experiments were performed.
Based on the morphological analysis of multiple organs by H&E
staining, it is considered that neutrophils play an important role
in sepsis. First, the purity of neutrophils (stained with Ly6G
antibody) isolated from mouse blood was determined using flow
cytometry. The results showed that neutrophils accounted for ~5% of
the peripheral blood lymphocytes in mice before isolation. After
isolation, the purity of neutrophils was detected by FACS analysis,
and the purity of neutrophils was ~87% among the collected cells
(Fig. 3A). In addition, the
viability of the isolated neutrophils was analysed using the CCK-8
assay. The results indicated that LPS decreased the viability of
neutrophils in peripheral blood, while MCC950 alleviated cell
damage (Fig. 3B). The ROS levels
in neutrophils isolated from each group of mice, were also
assessed. The results revealed that LPS increased ROS levels in
neutrophils, while MCC950 alleviated the oxidative stress induced
by LPS (Fig. 3C). To determine the
proportion of cells, Annexin V/PI staining was performed and FACS
was used for detection. The results revealed that LPS injection
promoted cell damage, including early-stage apoptosis (PI-negative
and Annexin V-positive) and late-stage apoptosis (PI-positive and
Annexin V-positive), while MCC950 alleviated the early and late
stage of apoptosis. Thus, it was determined that MCC950 treatment
alleviated damage in neutrophils (Fig.
3D).
MCC950 attenuates pyroptosis in
neutrophils by inhibiting the ROS/NLRP3/caspase-1 pathway
In order to investigate the mechanism of MCC950 in
LPS-induced neutrophil pyroptosis, proteins related to pyroptosis
were analysed by western blotting. The NLRP3 protein level was
increased in the LPS group, and MCC950 downregulated its
expression. Consistent with the change in NLRP3 expression, the
levels of cleaved caspase-1 and ASC were upregulated by LPS, while
MCC950 decreased their expression (Fig. 4A). In addition, the inflammatory
cytokines IL-1β and IL-18, which are downstream of NLRP3, were
induced by LPS, and MCC950 reversed this trend (Fig. 4B). Cleaved GSDMD, a marker of
pyroptosis, was enhanced during LPS-induced inflammation, and
MCC950 treatment inhibited this process (Fig. 4C). Taken together, these data
indicated that the NLRP3 inflammasome played a critical role in
LPS-induced neutrophil pyroptosis and that MCC950 exerted a
protective effect on the ROS/NLRP3/caspase-1 axis.
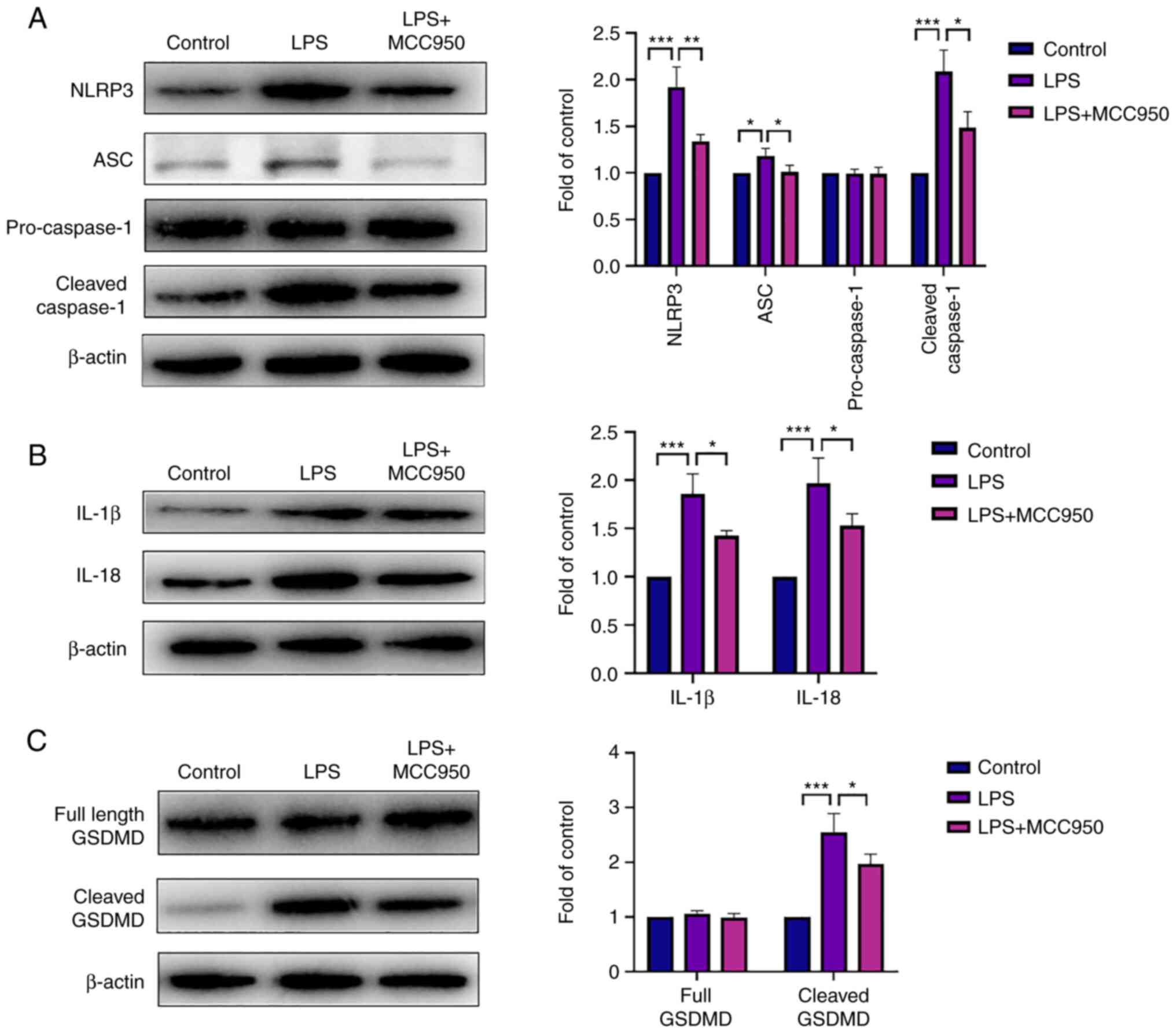 | Figure 4MCC950 attenuates pyroptosis in
neutrophils by inhibiting the NLRP3 inflammasome pathway. (A)
Representative bands of NLRP3, ASC, pro-caspase-1, cleaved
caspase-1, and β-actin (left panel). The expression of these
proteins (right panel). (B) Representative bands of mature IL-1β,
IL-18, and β-actin (left panel). The expression of these proteins
(right panel). (C) Representative bands of full-length GSDMD,
cleaved GSDMD, and β-actin (left panel). The expression of these
proteins (right panel). β-Actin was used as an internal control.
Data are shown as the mean ± SD; (n=6). *P<0.05,
**P<0.01 and ***P<0.001. NLRP3, NLR
family pyrin domain containing 3; IL, interleukin; GSDMD, gasdermin
D; LPS, lipopolysaccharide; ASC, apoptosis-associated speck-like
protein containing CARD. |
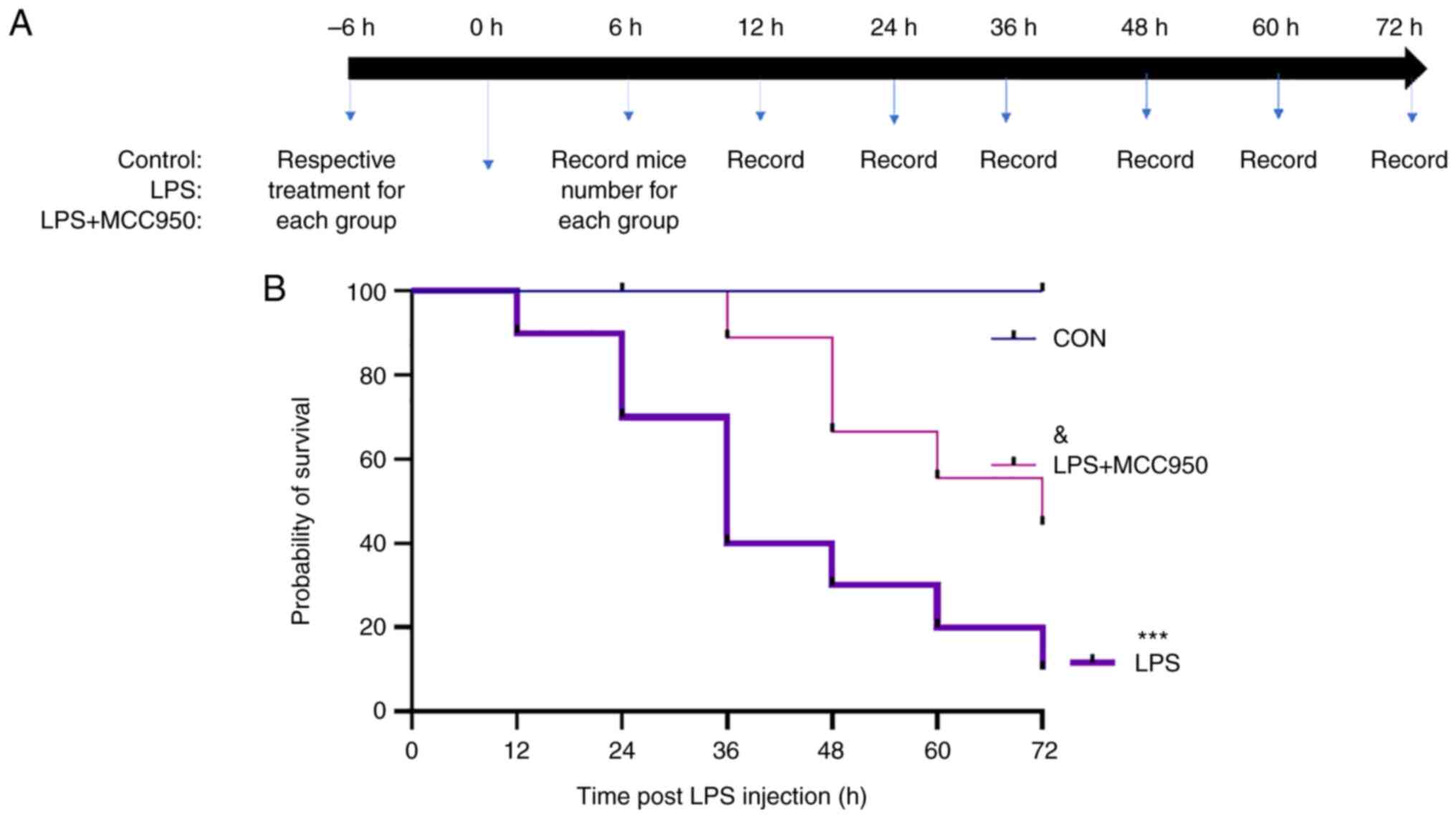
MCC950 improves the mortality of
septic mice
Survival rate analysis was performed according to
the schematic in Fig. 5A. The
control group exhibited a 100% survival rate. Compared with mice in
the LPS group, MCC950-treated mice exhibited significantly improved
survival during sepsis (Fig.
5B).
Discussion
Sepsis is a systemic inflammatory response syndrome
caused by the invasion of pathogenic microorganisms, such as
bacteria and viruses (13). The
primary syndromes of sepsis are systemic inflammation, organ
hypoperfusion, and even septic shock (14). With an increased understanding of
the Surviving Sepsis Campaign guidelines for diagnosis and
treatment, drugs and support methods for multiple organ function
have been developed and have increased the short-term mortality
rate of patients with sepsis to ~20% (15). However, new drug applications are
still required. In the present study, evidence for a potential drug
for the treatment of sepsis was provided. It was revealed that
MCC950 alleviated LPS-induced systemic immune responses, including
neutrophil infiltration in multiple organs, inflammatory cytokine
secretion, and necrosis of blood neutrophils. Previous studies have
reported that MCC950 as a small-molecule inhibitor directly
inhibits the activation of NLRP3 inflammasome (16,17).
A possible mechanism for this effect of MCC950 may involve the
relief of pyroptosis in blood neutrophils by reducing oxidative
stress, decreasing NLRP3 inflammasome levels, and reducing cleaved
GSDMD (7). In short, the findings
of the present study revealed a protective effect of MCC950 on
neutrophils and provide a potential use for MCC950 as a therapeutic
agent for sepsis.
A previous study reported that sepsis induced severe
multiple organ injury (18). To
evaluate the effect of MCC950 on LPS-induced inflammation, the
levels of ALT and AST (the indicators of liver damage), BUN and Cre
(the indicators of kidney damage), and the inflammatory cytokines
IL-1β, IL-6, IL-18, and TNF-α in serum, were analysed. The results
showed that MCC950 decreased the concentration of these indicators
of damage and alleviated the systemic symptoms of multiple organ
damage. These results were consistent with H&E staining of
these organs, which showed the infiltration of neutrophils in the
kidney and liver. In addition, the mortality of mice in the three
groups 72 h after LPS administration was observed. The results
revealed that MCC950 decreased the mortality of septic mice, which
was consistent with previous research in rats (19).
Neutrophils play a critical role in defence against
bacterial infection (20,21). The mechanism of the effect of
MCC950 on neutrophil function in sepsis is unclear. Pyroptosis is a
form of programmed cell death that occurs via a classical caspase-1
or noncaspase-1 pathway. A previous study reported that LPS induced
the caspase-1-dependent pathway via NLRP3 inflammasome activation
(22). The results of the present
study confirmed this finding and further revealed the mechanism of
this process. In addition, it was demonstrated that the components
NLRP3, ASC, and cleaved caspase-1 were increased in the blood
neutrophils of LPS-induced septic mice. In addition, ROS levels in
isolated cells were examined, and LPS administration aggravated
oxidative stress in blood neutrophils. A previous study reported
that the nicotine-NLRP3-ASC-pyroptosis pathway was activated by
ROS, since ROS scavenger, N-acetyl-cysteine (NAC), prevented
endothelial cell pyroptosis (23).
Previous research has revealed that Nur77 functions as an
intracellular LPS sensor, binding mitochondrial DNA and LPS to
activate the non-canonical NLRP3 inflammasome (24). In the present study, MCC950
treatment alleviated oxidative stress and downregulated downstream
NLRP3 inflammasome activation. After NLRP3 inflammasome activation,
the Gasdermin-D domain is cleaved to activate N-terminal GSDMD.
Activated GSDMD forms pores in the cell membrane, causing cell
necrosis and the release of various inflammatory cytokines, such as
IL-1β and IL-18 (25,26). The results of the present study
revealed that LPS-induced GSDMD was cleaved and further promoted
the maturation of IL-1β and IL-18. Annexin V/PI staining
demonstrated that neutrophil necrosis was increased in LPS-treated
mice. MCC950 treatment decreased the necrosis level of blood
neutrophils induced by LPS and decreased IL-1β and IL-18 levels.
These results indicated that MCC950 alleviated LPS-induced sepsis
by regulating neutrophil pyroptosis.
In conclusion, the present study revealed that
MCC950 could alleviate LPS-induced systemic immune responses,
including neutrophil infiltration in multiple organs, inflammatory
cytokine secretion, and necrosis in blood neutrophils. A possible
mechanism for this effect of MCC950 may involve the relief of
pyroptosis in blood neutrophils by inhibiting the ROS/NLRP3/IL-1β
pathway. The findings of the present study revealed a protective
effect of MCC950 on neutrophils and provide a possible use of
MCC950 as a therapeutic agent for sepsis.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Natural Science
Foundation of Yangzhou (grant no. YZ2019077).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
RM and JH conceived and designed the study. RM
performed the experiments and the data analysis. RM and JH wrote
the main manuscript text and prepared the figures. Both authors
read and approved the final version of the manuscript. RM and JH
confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
All experiments were approved by the Ethics
Committee of Yangzhou University (Yangzhou, China) (approval no.
202201001). All authors have read the ARRIVE guidelines and the
study was performed according to the ARRIVE guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Shen X, Cao K, Zhao Y and Du J: Targeting
neutrophils in sepsis: From mechanism to translation. Front
Pharmacol. 12(644270)2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Singer M, Deutschman CS, Seymour CW,
Shankar-Hari M, Annane D, Bauer M, Bellomo R, Bernard GR, Chiche
JD, Coopersmith CM, et al: The Third International consensus
definitions for sepsis and septic shock (Sepsis-3). JAMA.
315:801–810. 2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Zhaolin Z, Guohua L, Shiyuan W and Zuo W:
Role of pyroptosis in cardiovascular disease. Cell Prolif.
52(e12563)2019.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Pronin A, Pham D, An W, Dvoriantchikova G,
Reshetnikova G, Qiao J, Kozhekbaeva Z, Reiser AE, Slepak VZ and
Shestopalov VI: Inflammasome activation induces pyroptosis in the
retina exposed to ocular hypertension injury. Front Mol Neurosci.
12(36)2019.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Skoglund C, Appelgren D, Johansson I,
Casas R and Ludvigsson J: Increase of neutrophil extracellular
traps, mitochondrial DNA and Nuclear DNA in newly diagnosed type 1
diabetes children but not in high-risk children. Front Immunol.
12(628564)2021.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Phillipson M and Kubes P: The healing
power of neutrophils. Trends Immunol. 40:635–647. 2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Danielski LG, Giustina AD, Bonfante S,
Barichello T and Petronilho F: The NLRP3 inflammasome and its role
in sepsis development. Inflammation. 43:24–31. 2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Wang J, Sahoo M, Lantier L, Warawa J,
Cordero H, Deobald K and Re F: Caspase-11-dependent pyroptosis of
lung epithelial cells protects from melioidosis while caspase-1
mediates macrophage pyroptosis and production of IL-18. PLoS
Pathog. 14(e1007105)2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang YC, Liu QX, Zheng Q, Liu T, Xu XE,
Liu XH, Gao W, Bai XJ and Li ZF: Dihydromyricetin alleviates
sepsis-induced acute lung injury through inhibiting NLRP3
inflammasome-dependent pyroptosis in mice model. Inflammation.
42:1301–1310. 2019.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Gao YL, Zhai JH and Chai YF: Recent
advances in the molecular mechanisms underlying pyroptosis in
sepsis. Mediators Inflamm. 2018(5823823)2018.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zheng X, Chen W, Gong F, Chen Y and Chen
E: The role and mechanism of pyroptosis and potential therapeutic
targets in sepsis: A review. Front Immunol.
12(711939)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Zhang H, Du Y, Guo Y, Wang Z, Li H, Lv Z,
Zeng L, Chen Y, Xie Z and Li R: TLR4-NLRP3-GSDMD-Mediated
pyroptosis plays an important role in aggravated liver injury of
CD38(-/-) sepsis mice. J Immunol Res. 2021(6687555)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Minasyan H: Sepsis: Mechanisms of
bacterial injury to the patient. Scand J Trauma Resusc Emerg Med.
27(19)2019.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Park GL, Park M, Min JK, Park YJ, Chung SW
and Lee SJ: Anisomycin protects against sepsis by attenuating IκB
kinase-dependent NF-κB activation and inflammatory gene expression.
BMB Rep. 54:545–550. 2021.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Fleischmann C, Scherag A, Adhikari NK,
Hartog CS, Tsaganos T, Schlattmann P, Angus DC and Reinhart K:
International Forum of Acute Care Trialists. Assessment of global
incidence and mortality of hospital-treated sepsis. Current
estimates and limitations. Am J Respir Crit Care Med. 193:259–272.
2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Coll RC, Hill JR, Day CJ, Zamoshnikova A,
Boucher D, Massey NL, Chitty JL, Fraser JA, Jennings MP, Robertson
AAB and Schroder K: MCC950 directly targets the NLRP3
ATP-hydrolysis motif for inflammasome inhibition. Nat Chem Biol.
15:556–559. 2019.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Sui A, Chen X, Shen J, Demetriades AM, Yao
Y, Yao Y, Zhu Y, Shen X and Xie B: Inhibiting the NLRP3
inflammasome with MCC950 ameliorates retinal neovascularization and
leakage by reversing the IL-1β/IL-18 activation pattern in an
oxygen-induced ischemic retinopathy mouse model. Cell Death Dis.
11(901)2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Garofalo AM, Lorente-Ros M, Goncalvez G,
Carriedo D, Ballén-Barragán A, Villar-Fernández A, Peñuelas Ó,
Herrero R, Granados-Carreño R and Lorente JA: Histopathological
changes of organ dysfunction in sepsis. Intensive Care Med Exp.
7(Suppl 1)(S45)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhang XY, Chen X, Zhang HF, Guan S, Wen
SH, Huang WQ and Liu ZM: Propofol does not reduce pyroptosis of
enterocytes and intestinal epithelial injury after
lipopolysaccharide challenge. Dig Dis Sci. 63:81–91.
2018.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ji J and Fan J: Neutrophil in reverse
migration: Role in sepsis. Front Immunol. 12(656039)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Qi X, Yu Y, Sun R, Huang J, Liu L, Yang Y,
Rui T and Sun B: Identification and characterization of neutrophil
heterogeneity in sepsis. Crit Care. 25(50)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Kelley N, Jeltema D, Duan Y and He Y: The
NLRP3 inflammasome: An overview of mechanisms of activation and
regulation. Int J Mol Sci. 20(3328)2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Wu X, Zhang H, Qi W, Zhang Y, Li J, Li Z,
Lin Y, Bai X, Liu X, Chen X, et al: Nicotine promotes
atherosclerosis via ROS-NLRP3-mediated endothelial cell pyroptosis.
Cell Death Dis. 9(171)2018.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhu F, Ma J, Li W, Liu Q, Qin X, Qian Y,
Wang C, Zhang Y, Li Y, Jiang D, et al: The orphan receptor Nur77
binds cytoplasmic LPS to activate the non-canonical NLRP3
inflammasome. Immunity. 36:753–767.e8. 2023.PubMed/NCBI View Article : Google Scholar
|
|
25
|
He WT, Wan H, Hu L, Chen P, Wang X, Huang
Z, Yang ZH, Zhong CQ and Han J: Gasdermin D is an executor of
pyroptosis and required for interleukin-1β secretion. Cell Res.
25:1285–1298. 2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Tapia-Abellán A, Angosto-Bazarra D,
Martínez-Banaclocha H, de Torre-Minguela C, Cerón-Carrasco JP,
Pérez-Sánchez H, Arostegui JI and Pelegrin P: Addendum: MCC950
closes the active conformation of NLRP3 to an inactive state. Nat
Chem Biol. 17(361)2021.PubMed/NCBI View Article : Google Scholar
|