Introduction
Ischemia-reperfusion injury (IRI) is a frequent
occurrence seen in various diseases and surgical procedures such as
coronary heart disease, kidney transplants, and liver transplants
(1-3).
Interruption or reduction of blood flow leads to cellular ischemia
and hypoxia, resulting in tissue and/or organ damage. The
metabolites and mediators produced during ischemia promote further
damage, as reflected by the serious dysfunction of the cell
ultrastructure, metabolism, electrophysiology, and function
(4). The primary causes of renal
ischemia-reperfusion injury (RIRI) are cardiac arrest (systemic
hypoperfusion) and renal surgical intervention (5). In the case of kidney transplants,
kidney injury occurs following a short period of warm ischemia
following the removal of the organ from the donor, a prolonged
period of cold ischemia during cryopreservation, and a final stage
of warm ischemia during recipient implantation.
Post-revascularization, renal blood reperfusion triggers a series
of events that can exacerbate renal injury (6), resulting in hemodynamic fluctuations,
inflammation, and damage to tubular epithelial cells (7). The pathogenesis of RIRI may be
attributed to structural and functional dysmetabolism of the
kidneys induced by a range of factors, such as excessive production
of oxygen-free radicals, intracellular calcium overload,
inflammatory factors and transmitters, apoptosis, membrane lipid
peroxidation, and alterations in nitric oxide content (8-10).
Salidroside (SA) is a natural active ingredient
extracted from Rhodiola rosea (11), renowned for its diversified
biological activities, including anti-inflammatory, anti-oxidative,
anti-tumorigenic, and anti-radiation properties (11). As a result of its broad
pharmacological effects, SA has attracted the attention both
domestically and internationally. Numerous studies have documented
the protective effects of SA against a range of organ injuries,
primarily by reducing oxidative stress, inhibiting apoptosis,
reducing intracellular calcium overload, and enhancing
mitochondrial function (11-13).
Prior studies have shown SA to have a protective effect on
myocardial cell injury caused by hypoxia, as well as having
anti-arrhythmic properties. Furthermore, it inhibits the
proliferation and contraction of vascular smooth muscle cells. Its
mechanisms possibly involve reducing creatine phosphate (CK)
activity and malonaldehyde (MDA) content, increasing superoxide
dismutase (SOD) activity, increasing sarcoplasmic reticulum
Ca2+-ATPase (SERCA) activity, reducing Ca2+
overload, and suppressing apoptosis-related gene expression
(14-16).
Although numerous studies have confirmed that SA has a protective
effect on IRI of myocardium, testis, and the brain, its effect on
RIRI and the underlying mechanisms remain unknown.
Materials and methods
Animals
Male Sprague-Dawley rats (weighing 200-220 g) were
purchased from the Animal Center of Wuhan University. The rats were
housed in a temperature and humidity-controlled environment with a
12-h light/dark cycle and provided with standard food and water.
This study was performed in accordance with the Helsinki
Declaration II (17) and was
approved by the Institutional Review Boards of Renmin Hospital of
Wuhan University (approval no. K2021-08-012).
Establishment of the RIRI model
The rats were subjected to an 8 h fast prior to the
surgical procedure. Thereafter, sodium pentobarbital (50 mg/kg,
intraperitoneal injection) was injected into the abdominal cavity
to anesthetize the rats, and the limbs were immobilized during the
operation. An abdominal incision was used to reveal the renal
pedicle, and the bilateral renal arteries were ligated for 45 min.
During reperfusion, the hemostatic clamp was removed, and changes
in kidney color were assessed. Blood flow to the kidney generally
resumed to normal within a few min, which was then followed by
suturing of the incision. The sham group underwent the same
surgical procedure without clamping of the renal pedicle. After 24
h, rats were humanely sacrificed under anesthesia by
intraperitoneal injection with 1% pentobarbital sodium (150 mg/kg).
The kidneys were harvested, and serum samples were collected.
In vivo treatment protocols
All rats were randomly divided into three groups:
Sham group (n=6), IR group (n=6), and IR+SA group (n=18). To
explore the effects of SA on RIRI, SA group rats were given 1, 10,
or 100 mg/kg SA (cat. no. 10338-51-9; MedChemExpress) daily by
gavage for 7 days before the IR procedure (n=6 per group).
Cell culture
The rat renal tubular epithelial cell line, NRK
cells, were cultured in DMEM (cat. no. 12430112; Sigma-Aldrich;
Merck KGaA; Thermo Fisher Scientific, Inc.) with 1%
Penicillin-Streptomycin solution (cat. no. 15140122; Sigma-Aldrich;
Merck KGaA; Thermo Fisher Scientific, Inc.) and 10% FBS (cat. no.
16140089; Sigma-Aldrich; Merck KGaA; Thermo Fisher Scientific,
Inc.) and maintained in a humidified incubator at 37˚C incubator
supplied with 5% CO2.
Establishment of the
hypoxia-reoxygenation (HR) model
NRK cells were starved in serum-free DMEM for 12 h,
and then incubated in a thermostatic tri-gas incubator supplied
with 95% N2 for 8 h to simulate a hypoxic environment.
Subsequently, the cells were moved to a normoxic incubator for a
period of 12 h, followed by the replacement of the media with
supplemented with DMEM for 12 h of reoxygenation.
In vitro treatment protocols
To explore the effects of SA and the PI3K/AKT
signaling pathway, NRK cells were incubated with SA (1-1,000 µM) or
PI3K/AKT-IN-1 (2.62 µM; cat. no. HY-144806; MedChemExpress) at the
onset of reoxygenation. Cells were randomly divided into one of
four groups: Control group, HR group, HR+SA group, and
HR+SA+PI3K/AKT-IN-1 group.
Cell viability assay
Cell viability was determined using a CCK-8 assay
(cat. no. C0038; Sigma-Aldrich; Merck KGaA Institute of
Biotechnology). Briefly, NRK cells were seeded into 96-well plates
at a density of 1x104 cells/well and incubated for 12 h
in a thermostatic incubator. Upon treating cells with SA and/or
PI3K/AKT-IN-1 for 4 h, 10 µl CCK-8 solution was added to each well,
and cells were cultured for an additional 1 h. Subsequently, the
absorbance at 450 nm was measured using a microplate reader
(PerkinElmer; Thermo Fisher Scientific, Inc.).
Renal function analysis and
histopathological analysis
Renal function was evaluated by measuring serum
creatinine (Scr) and blood urea nitrogen (BUN) levels in plasma at
the Department of Clinical Laboratories of Renmin Hospital, Wuhan
University. Kidney samples were procured, fixed with
paraformaldehyde, and then embedded in paraffin. The samples were
sectioned into 4-µm thick slices, and then stained with hematoxylin
for 5 min and eosin for 3 min at room temperature. The severity of
RIRI was assessed using the Paller scoring system (18), with histopathological changes
graded as follows: i) 1 point, tubular epithelial smoothness or
tubular expansion; ii) 1 or 2 points, loss of brush-like edge; iii)
1 or 2 points, obstruction of the tubular lumen; iv) 1 point,
cytoplasmic vacuolization; and v) 1 point, cell necrosis. The
histological data were analyzed and graded independently by two
observers who were blinded to the experimental groups.
Western blotting
Proteins from kidney tissues or NRK cells were
extracted and lysed in RIPA buffer supplemented with protease and
phosphatase inhibitors. Protein content was measured using a BCA
kit (cat no. A045-4-2, Nanjing Jiancheng Bioengineering Institute).
Subsequently, 50 µg protein samples were loaded on 10% SDS gels,
resolved using SDS-PAGE, and subsequently transferred to a PVDF
membrane. The membranes were blocked with 5% BSA for 1 h at room
temperature and incubated with one of the following primary
antibodies: Rabbit anti-GPX4 (1:1,000; cat. no. 67763-1-Ig,
Sigma-Aldrich; Merck KGaA Group, Inc.), rabbit anti-ACSL4 (1:1,000.
cat. no.22401-1-AP; Sigma-Aldrich; Merck KGaA Group, Inc.), rabbit
anti-PI3K antibody (1:1,000; cat. no. 4249; Cell Signaling
Technology, Inc.), rabbit anti-p-PI3K antibody (1:1,000; cat. no.
17366; Cell Signaling Technology, Inc.), rabbit anti-AKT (1:1,000;
cat. no. 9272; Cell Signaling Technology, Inc.), rabbit anti-p-AKT
antibody (1:1,000; cat. no. 4060; Cell Signaling Technology, Inc.),
rabbit anti-SOD1 antibody (1:1,000; cat. no. 37385; Cell Signaling
Technology, Inc.), rabbit anti-SOD2 antibody (1:1,000; cat. no.
13141; Cell Signaling Technology, Inc.) and rabbit anti-GAPDH
antibody (1:10,000; cat. no. 5174; Cell Signaling Technology, Inc.)
overnight at 4˚C. The following day, the membranes were washed in
TBST and incubated for 2 h at room temperature with the
corresponding secondary antibodies. Signals were visualized using
an ECL system and ECL kit (cat. no. W028-2-1; Nanjing Jiancheng
Bioengineering Institute), and densitometry analysis was calculated
using ImageJ version 1.8.0 (National Institutes of Health).
Detection of the oxidative stress
indicators
The levels of malondialdehyde (MDA) and superoxide
dismutase (SOD) in kidney tissues or NRK cells were detected using
an MDA assay kit (cat. no. S0131S; Sigma-Aldrich; Merck KGaA
Institute of Biotechnology) and SOD assay kit (cat. no. S0101S;
Sigma-Aldrich; Merck KGaA Institute of Biotechnology) according to
the manufacturer's protocol.
Measurement of ROS levels in the
kidney tissues
The levels of ROS were analyzed in the tissues
utilizing the dihydroethidium (DHE) fluorescent probe (cat. no.
D7008; MilliporeSigma), according to the manufacturer's protocol.
The sections were stained by incubation with 50 µM DHE for 1 h in
the dark at room temperature, and then stained with 1 mg/ml DAPI
for 10 min at 4˚C. The sections were rinsed three times with PBS. A
fluorescent microscope (magnification, x200) was used to evaluate
the fluorescence intensity at an excitation wavelength of 525 nm
and an emission wavelength of 610 nm.
Evaluation of intracellular lipid
hydroperoxide (LPO)
C11-BODIPY581/591 (cat. no. HY-D1301;
MedChemExpress, Inc.) was used as a fluorescent probe to assess
intracellular LPO levels. The probe integrates into the lipid
membrane and is oxidized by intracellular LPO, fluorescing green
once oxidized. A total of 5 µM C11-BODIPY581/591 was
used, and kidney slices were incubated for 30 min at 4˚C. After
washing with PBS, the slices were examined by flow cytometer BD
FACSaria II (version 7.6.1, BD Biosciences).
Evaluation of ROS levels in NRK
cells
The 2',7'-dichlorofluorescein diacetate (DCFH-DA)
fluorescent probe (cat. no. D6883; MilliporeSigma-Aldrich; Merck
KGaA) was used to detect ROS levels in NRK cells according to the
manufacturer's protocols. After incubating with 10 µM DCFH-DA for 1
h at 4˚C in the dark, NRK cells were treated with 1 µg/ml DAPI for
10 min at 4˚C. The cells were observed using a fluorescent
microscope at a wavelength of 485 nm.
Statistical analysis
GraphPad Prism version 8.0.1 (GraphPad Software,
Inc.) was used to analyze the data. Data are presented as the mean
± SD of three repeats. Differences between groups were compared
using one-way ANOVA followed by a post hoc Tukey's. P<0.05 was
considered to indicate a statistically significant difference.
Results
SA protects against RIRI in a
dose-dependent manner
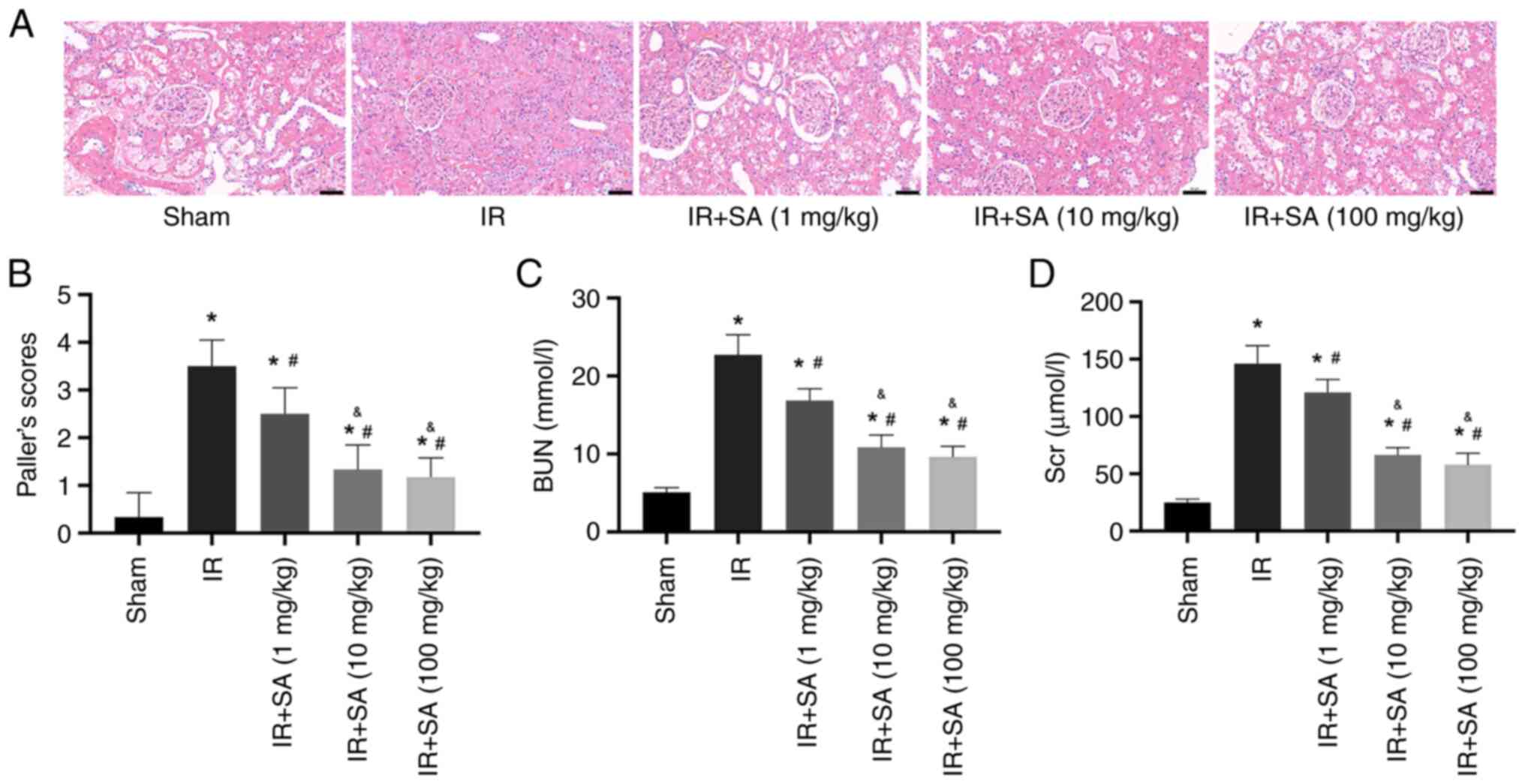
To assess the role of SA in RIRI, rats were
subjected to different doses of SA (1, 10, or 100 mg/kg/day),
administered via gavage for 7 days before undergoing the
establishment of RIRI. Renal function was assessed following this
treatment. The results of hematoxylin and eosin staining and renal
injury score analysis demonstrated reduced IR-induced renal tubular
injury in the rats pretreated with SA. In the RIRI rats,
pretreatment with 1, 10, and 100 mg/kg of SA significantly
decreased renal tubular injury, with the optimal effect apparent at
10 and 100 mg/kg, whereas the renal protective effect of 1 mg/kg SA
pretreatment was less prominent (Fig.
1A and B). Accordingly, 10
mg/kg SA was used for all subsequent experiments. Moreover, the
concentrations of Scr and BUN in the IR group were notably higher
than those in the sham group. Compared to the IR group,
pretreatment with SA significantly reduced Scr and BUN levels in
the serum (Fig. 1C and D).
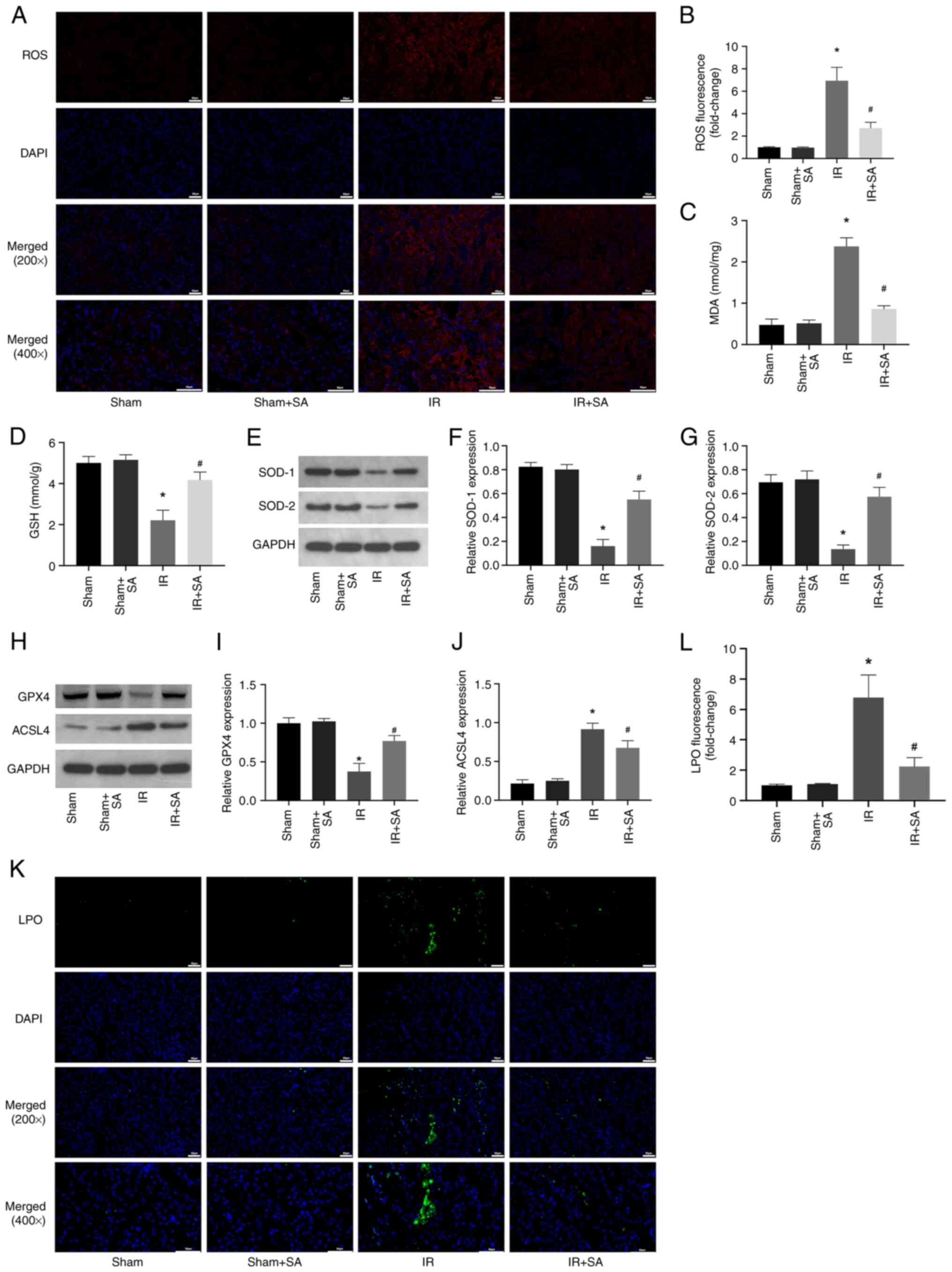
SA inhibits IR-induced oxidative
stress and ferroptosis in rats
Next, the effect of SA pretreatment on the levels of
ROS in the IR-induced kidneys was examined. The results showed that
SA pretreatment inhibited IR-induced increases in ROS and MDA while
increasing the levels of GSH, SOD1, and SOD2 (Fig. 2A-G). Numerous studies have shown
that high levels of oxidative stress can induce ferroptosis
(19-21).
Therefore, subsequent investigations focused on the effect of SA on
the ferroptosis of renal cells during IR. The results showed that
SA pretreatment inhibited the decrease in the expression of GPX4
and the increase in the expression of ACSL4 induced by RIRI
(Fig. 2H-J). Furthermore, the
levels of LPO were detected using the lipid peroxidation probe
C11-BODIPY581/591. The significant increase in LPO
induced by IR was prevented by treatment with SA (Fig. 2K and L). Together, the results suggested that
SA pretreatment conferred its renal protective effects through
anti-oxidative and anti-ferroptotic means.
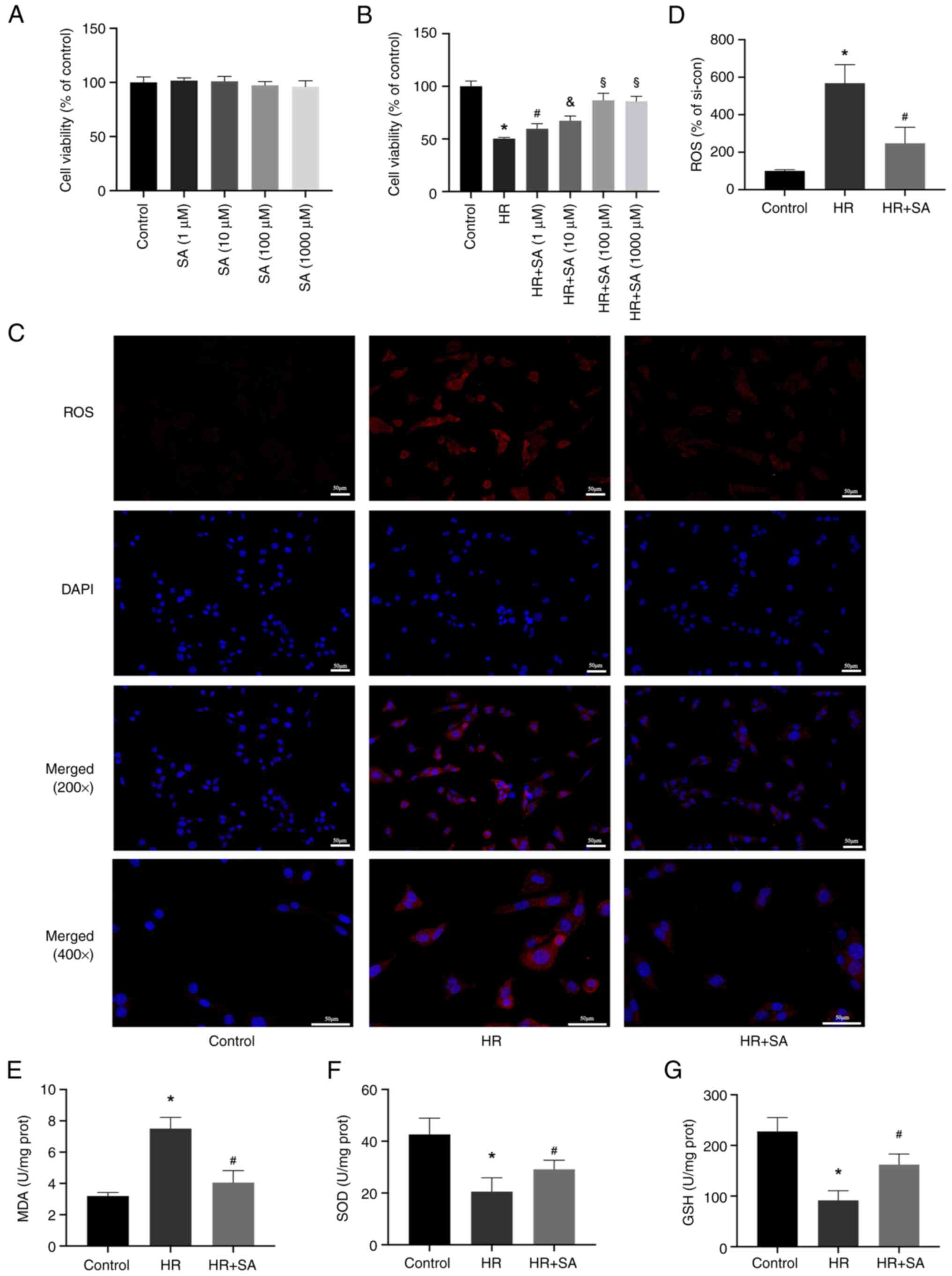
SA reduces oxidative stress induced by
HR injury in NRK cells
Subsequently, in vitro experiments were
performed to evaluate the effect of SA on the human renal tubular
epithelial cell line NRK. First, the toxicity of SA in NRK cells
was evaluated and it was found that SA did not noticeably impact
cell viability after 24 h (Fig.
3A). Additionally, we discovered that pretreatment with SA
reduced HR-induced cell death. Treatment with 1-1,000 µM SA exerted
a renal protective effect, with the optimal effect evident at 100
µM (Fig. 3B). Therefore, a
concentration of 100 µM of SA was used for further experiments. DHE
staining and MDA detection kits were used to assess the effects of
SA on ROS levels induced by HR. The results showed that reduced ROS
levels in NRK cells were associated with increased SOD and GSH
activity and decreased activity of MDA (Fig. 3C-G). In agreement with the in
vivo experiments, the outcomes of the in vitro
experiments showed that SA protected renal tubular epithelial cells
from oxidative damage through suppression of ROS buildup.
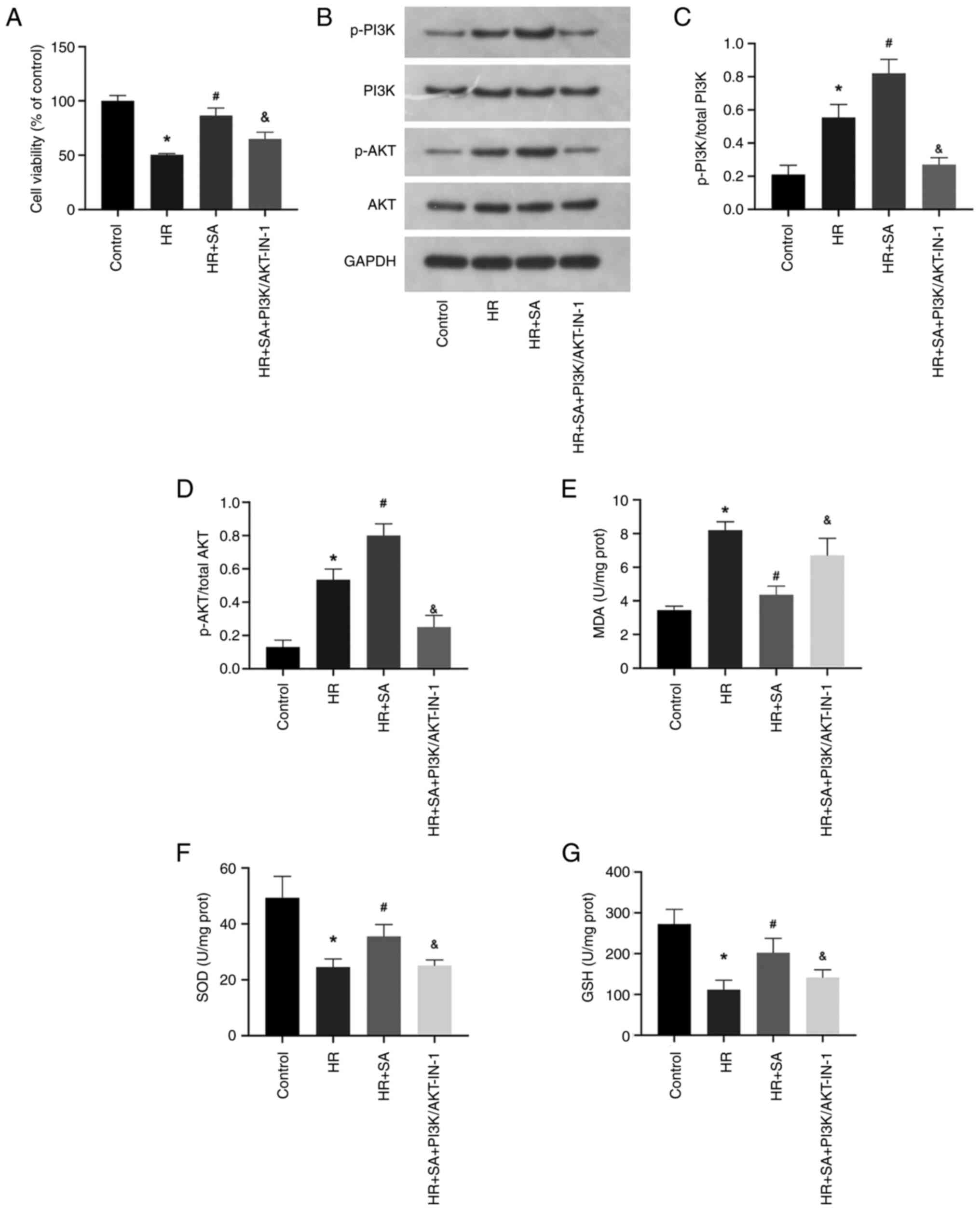
PI3K/AKT signaling pathway is involved
in the protective effects of SA on oxidative stress against HR
injury
The PI3K/AKT signaling pathway is a critical player
in the modulation of oxidative stress. Therefore, to investigate
whether SA exerts a protective effect on HR injury by targeting the
PI3K/AKT signaling pathway, a PI3K/AKT pathway inhibitor,
PI3K/AKT-IN-1 (2.62 µM), was employed. The cell viability assay
demonstrated that PI3K/AKT-IN-1 partially abrogated the protective
effect of SA against HR-induced NRK cell injury (Fig. 4A). Moreover, compared to the sham
group, there was an increase in the phosphorylation levels of PI3K
and AKT during HR injury, and SA pretreatment further augmented the
phosphorylation of PI3K and AKT. However, in the presence of
PI3K/AKT-IN-1, the increased phosphorylation of PI3K and AKT was
suppressed (Fig. 4B-D).
Additionally, PI3K/AKT-IN-1 increased MDA levels while decreasing
the levels of SOD and GSH (Fig.
4E-G). Accordingly, SA exerts its antioxidant effect on NRK
cells by activating the PI3K/AKT signaling pathway.
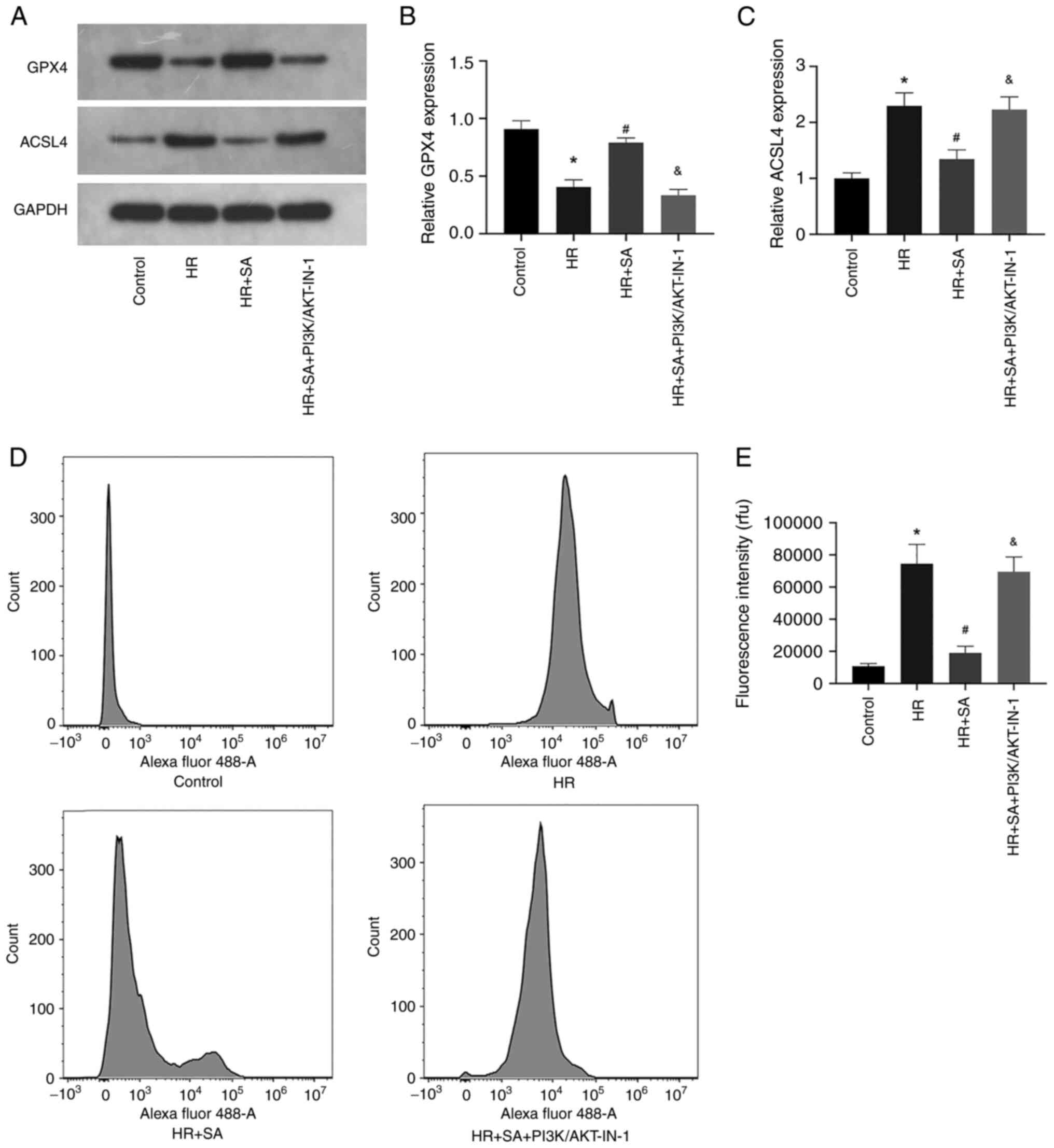
HR-induced ferroptosis can be
aggravated by blocking the PI3K/AKT signaling pathway
Finally, the effects of PI3K/AKT signaling pathway
inhibition on HR-induced ferroptosis of NRK cells were explored.
The results of western blotting showed that PI3K/AKT-IN-1 prevented
the increased expression of GPX4 and the decreased expression of
ACSL4 induced by SA (Fig. 5A-C).
Additionally, the levels of LPO were assessed using the lipid
peroxidation probe C11-BODIPY581/591. The data confirmed
that PI3K/AKT-IN-1 reversed the inhibitory effect of SA on LPO
(Fig. 2D and E). Collectively, the outcomes indicated
that the protective effects of SA pretreatment on renal tubular
epithelial cells were brought about through an anti-ferroptotic
effect; however, this effect could be counteracted by inhibition of
the PI3K/AKT signaling pathway.
Discussion
The results of the present study showed that SA
treatment is an effective method of reducing oxidative stress and
inhibiting ferroptosis during RIRI, leading to the alleviation of
this condition. Therefore, the use of natural products can be
considered a reference point for managing and preventing RIRI.
Natural products have demonstrated good efficacy in preventing and
treating AKI with minimal side effects. Among these,
Rhodiola, a perennial herb belonging to the sedum genus in
the Sedum family (22), is
noteworthy due to the presence of SA, tyrosol, flavonoid, amino
acid, trace volatile oil, and other constituents, which underlie
its beneficial effects (23,24).
Recent studies have demonstrated the potential of Rhodiola
rosea to affect various biological pathways, including
anti-hypoxic, anti-fatigue, anti-viral, immunity-enhancing,
anti-aging, and anti-radiation pathways, and bidirectional
regulation of bodily functions (25,26).
Rhodiola can significantly increase the endurance of
hypoxia-exposed human bodies, promote aerobic metabolism, reduce
the concentration of lactic acid in the blood, muscles, and brain,
increase the aerobic metabolism of the heart, brain, lung, and
other critical organs, assist the body in adapting to an hypoxic
environment, and deferring the onset of further fatigue (27-29).
SA, the primary constituent of Rhodiola, has been found by previous
studies to safeguard organs from multiple IRI-related pathway
dysfunctions (30-32).
The process of IRI involves various factors and
mechanisms, including reduced nitric oxide production,
inflammation-induced cell damage, oxygen-derived oxidative stress
formation, and lipid peroxidation (33). Prior work has identified oxidative
stress as a key potential mechanism of tissue damage induced by IRI
(34). Oxidative-free radicals and
ROS scavengers are vital for maintaining the oxidant-antioxidant
balance in vivo during IRI (35). Excessive generation of oxygen-free
radicals by ischemia-reperfusion can surpass local tissue
scavenging capacity and lead to damage (36). MDA serves as a stable metabolic
intermediate indicative of lipid peroxidation levels and
oxygen-free radical content in tissues. Prior research has shown
that IRI elevates ROS and MDA levels (35). The present study also confirmed
that RIRI led to a significant increase in oxidative stress and
that pretreatment with SA mitigated this, demonstrating the
protective effect of SA by inhibiting oxidative stress and
ferroptosis.
SA is a widely studied natural antioxidant with
potent antioxidant effects. AKT, which is mediated by
phosphoinositol-dependent kinase-1 (PDK-1) (37), acts as a downstream factor of PI3K.
Studies have suggested that natural antioxidants can activate PI3K
by binding to GPCRs and EGFR (38,39).
Therefore, it was hypothesized that SA, like other natural
antioxidants, may activate the PI3K/AKT signaling pathway; prior
work confirmed that SA indeed activates this pathway (11,40).
In the present study, SOD1/2 expression and activity in IRI kidneys
were initially evaluated, followed by verification of these
findings using NRK cells. SA pretreatment increased SOD activity by
further activating the PI3K/AKT signaling pathway during HR
treatment. These results confirmed that the PI3K/AKT signaling
pathway underlies the anti-oxidative stress effect of SA.
Ferroptosis is a form of cell death resulting from
lethal lipid peroxidation and iron-dependent damage of membrane
lipids. Ferroptosis is involved in various organ injuries induced
by IRI, including the heart, liver, kidney, and brain (41-44).
Nonetheless, it remains unclear whether SA can exert a protective
effect by inhibiting ferroptosis during RIR. Inactivation of the
lipid repair enzyme GPX4 leads to the accumulation of lipid
peroxides, including lipid hydrogen peroxide, as GPX4 is the sole
enzyme capable of reducing lipid peroxidation in biofilms, and GSH
is a necessary co-factor for GPX4 enzyme activity. GPX4
inactivation results in the accumulation of high levels of ROS,
ultimately leading to cell damage. The results of the present study
revealed that SA enhanced the antioxidant capacity of the kidney by
activating the PI3K/AKT signaling pathway, thereby inhibiting
ferroptosis induced by high levels of oxidative stress. In short,
SA exhibits a potent anti-ferroptotic effect on NRK cells, and this
effect can be reversed by inhibiting PI3K/AKT. Consequently,
ferroptosis induced by ROS is also suppressed. Additionally,
treatment with the PI3K/AKT pathway inhibitor, PI3K/AKT-IN-1,
inhibited the activation of the PI3K/AKT pathway and its downstream
regulation of oxidative stress and ferroptosis. These findings
support the notion that SA pretreatment can alleviate RIRI by
activating the PI3K/AKT signaling pathway.
However, the present study has some limitations.
First, SA may be used as a clinical agent to treat RIRI, thus the
efficacy of SA in patients who already have RIRI should be
explored. Secondly, it was confirmed that 10-100 mg/kg SA had a
strong renal protective effect; however, the optimal dose of SA was
determined. These factors should be considered in further
studies.
In conclusion, the present study showed that SA
increased the activity of SODs by activating the PI3K/AKT signaling
pathway, thereby eliminating ROS, and inhibiting oxidative stress
injury and ferroptosis, thus protecting renal function in RIRI.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZT and CL conceived and designed the study, analyzed
the data, and wrote the manuscript. YW collected and analyzed the
data. YL analyzed the data and wrote the manuscript. CL and ZT
confirm the authenticity of all the raw data. All authors have read
and approved the final manuscript.
Ethics approval and consent to
participate
This study was performed in accordance with the
Helsinki Declaration II and was approved by the Institutional
Review Boards of Renmin Hospital of Wuhan University (approval no.
K2021-08-012).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
References
Tian H, Xiong Y, Zhang Y, Leng
Y, Tao J, Li L, Qiu Z and Xia Z: Activation of NRF2/FPN1 pathway
attenuates myocardial ischemia-reperfusion injury in diabetic rats
by regulating iron homeostasis and ferroptosis. Cell Stress
Chaperones. 27:149–164. 2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Wei X, Deng W, Dong Z, Xie Z, Zhang J,
Wang R, Zhang R, Na N and Zhou Y: Identification of subtypes and a
delayed graft function predictive signature based on ferroptosis in
renal ischemia-reperfusion injury. Front Cell Dev Biol.
10(800650)2022.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bardallo RG, Panisello-Roselló A,
Sanchez-Nuno S, Alva N, Roselló-Catafau J and Carbonell T: Nrf2 and
oxidative stress in liver ischemia/reperfusion injury. FEBS J.
289:5463–5479. 2022.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Patel PM, Connolly MR, Coe TM, Calhoun A,
Pollok F, Markmann JF, Burdorf L, Azimzadeh A, Madsen JC and
Pierson RN*III: Minimizing ischemia reperfusion injury in
xenotransplantation. Front Immunol. 12(681504)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Chatauret N, Badet L, Barrou B and Hauet
T: Ischemia-reperfusion: From cell biology to acute kidney injury.
Prog Urol. 24 (Suppl 1):S4–S12. 2014.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhao H, Alam A, Soo AP, George A and Ma D:
Ischemia-reperfusion injury reduces long term renal graft survival:
Mechanism and beyond. EBioMedicine. 28:31–42. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Bonventre JV and Yang L: Cellular
pathophysiology of ischemic acute kidney injury. J Clin Invest.
121:4210–4221. 2011.PubMed/NCBI View
Article : Google Scholar
|
|
8
|
Shen Y, Qiu T, Liu XH, Zhang L, Wang ZS
and Zhou JQ: Renal ischemia-reperfusion injury attenuated by
splenic ischemic preconditioning. Eur Rev Med Pharmacol Sci.
22:2134–2142. 2018.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Kinra M, Mudgal J, Arora D and Nampoothiri
M: An insight into the role of cyclooxygenase and lipooxygenase
pathway in renal ischemia. Eur Rev Med Pharmacol Sci. 21:5017–5020.
2017.PubMed/NCBI
|
|
10
|
Rodriguez F, Bonacasa B, Fenoy FJ and
Salom MG: Reactive oxygen and nitrogen species in the renal
ischemia/reperfusion injury. Curr Pharm Design. 19:2776–2794.
2013.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Rong L, Li Z, Leng X, Li H, Ma Y, Chen Y
and Song F: Salidroside induces apoptosis and protective autophagy
in human gastric cancer AGS cells through the PI3K/Akt/mTOR
pathway. Biomed Pharmacother. 122(109726)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Yuan Y, Wang Z, Nan B, Yang C, Wang M, Ye
H, Xi C, Zhang Y and Yan H: Salidroside alleviates liver
inflammation in furan-induced mice by regulating oxidative stress
and endoplasmic reticulum stress. Toxicology.
461(152905)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang P, Xu J, Cui Q, Lin G, Wang F, Ding
X, You S, Sang N, Tan J, Xu W, et al: Multi-pathway neuroprotective
effects of a novel salidroside derivative SHPL-49 against acute
cerebral ischemic injury. Eur J Pharmacol.
949(175716)2023.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Jiang S, Fan F, Yang L, Chen K, Sun Z,
Zhang Y, Cairang N, Wang X and Meng X: Salidroside attenuates high
altitude hypobaric hypoxia-induced brain injury in mice via
inhibiting NF-κB/NLRP3 pathway. Eur J Pharmacol.
925(175015)2022.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang X, Tang Y, Xie N, Bai J, Jiang S,
Zhang Y, Hou Y and Meng X: Salidroside, a phenyl ethanol glycoside
from Rhodiola crenulata, orchestrates hypoxic mitochondrial
dynamics homeostasis by stimulating Sirt1/p53/Drp1 signaling. J
Ethnopharmacol. 293(115278)2022.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Qi C, Zhang J, Chen X, Zhang J, Yang P,
Jiao Q, Zhang P, Lu HX and Liu Y: Salidroside protects cultured rat
subventricular zone neural stem cells against hypoxia injury by
inhibiting Bax, Bcl-2 and caspase-3 expressions. Nan Fang Yi Ke Da
Xue Xue Bao. 33:962–966. 2013.PubMed/NCBI(In Chinese).
|
|
17
|
Issue Information-Declaration of Helsinki.
J Bone Miner Res. 34:BMi–BMii. 2019.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Paller MS: Free radical-mediated
postischemic injury in renal transplantation. Ren Fail. 14:257–260.
1992.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Mancardi D, Mezzanotte M, Arrigo E,
Barinotti A and Roetto A: Iron overload, oxidative stress, and
ferroptosis in the failing heart and liver. Antioxidants (Basel).
10(1864)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Ren JX, Li C, Yan XL, Qu Y, Yang Y and Guo
ZN: Crosstalk between oxidative stress and ferroptosis/oxytosis in
ischemic stroke: Possible targets and molecular mechanisms. Oxid
Med Cell Longev. 2021(6643382)2021.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Yu Y, Yan Y, Niu F, Wang Y, Chen X, Su G,
Liu Y, Zhao X, Qian L, Liu P and Xiong Y: Ferroptosis: A cell death
connecting oxidative stress, inflammation and cardiovascular
diseases. Cell Death Discov. 7(193)2021.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Brinckmann JA, Cunningham AB and Harter D:
Running out of time to smell the roseroots: Reviewing threats and
trade in wild Rhodiola rosea L. J Ethnopharmacol.
269(113710)2021.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Langeder J, Grienke U, Döring K, Jafari M,
Ehrhardt C, Schmidtke M and Rollinger JM: High-performance
countercurrent chromatography to access Rhodiola rosea
influenza virus inhibiting constituents. Planta Med. 87:818–826.
2021.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Rattan S, Kumar A, Kumar D and Warghat AR:
Enhanced production of phenylethanoids mediated through synergistic
approach of precursor feeding and light regime in cell suspension
culture of Rhodiola imbricata (Edgew.). Appl Biochem Biotech.
194:3242–3260. 2022.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Li Y, Pham V, Bui M, Song L, Wu C, Walia
A, Uchio E, Smith-Liu F and Zi X: Rhodiola rosea L.: An herb
with anti-stress, anti-aging, and immunostimulating properties for
cancer chemoprevention. Curr Pharmacol Rep. 3:384–395.
2017.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Labachyan KE, Kiani D, Sevrioukov EA,
Schriner SE and Jafari M: The impact of Rhodiola rosea on
the gut microbial community of Drosophila melanogaster. Gut Pathog.
10(12)2018.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Ma Y, Wu Y, Xia Z, Li J, Li X, Xu P, Zhou
X and Xue M: Anti-hypoxic molecular mechanisms of Rhodiola
crenulata extract in zebrafish as revealed by metabonomics.
Front Pharmacol. 10(1356)2019.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Wang X, Hou Y, Li Q, Li X, Wang W, Ai X,
Kuang T, Chen X, Zhang Y, Zhang J, et al: Rhodiola crenulata
attenuates apoptosis and mitochondrial energy metabolism disorder
in rats with hypobaric hypoxia-induced brain injury by regulating
the HIF-1α/microRNA 210/ISCU1/2(COX10) signaling pathway. J
Ethnopharmacol. 241(111801)2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Xie N, Fan F, Jiang S, Hou Y, Zhang Y,
Cairang N, Wang X and Meng X: Rhodiola crenulate alleviates
hypobaric hypoxia-induced brain injury via adjusting
NF-κB/NLRP3-mediated inflammation. Phytomedicine.
103(154240)2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Dong C, Wen S, Zhao S, Sun S, Zhao S, Dong
W, Han P, Chen Q, Gong T, Chen W, et al: Salidroside inhibits
reactive astrogliosis and glial scar formation in late cerebral
ischemia via the Akt/GSK-3β pathway. Neurochem Res. 46:755–769.
2021.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Tian X, Huang Y, Zhang X, Fang R, Feng Y,
Zhang W, Li L and Li T: Salidroside attenuates myocardial
ischemia/reperfusion injury via AMPK-induced suppression of
endoplasmic reticulum stress and mitochondrial fission. Toxicol
Appl Pharm. 448(116093)2022.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yin L, Ouyang D, Lin L, Xin X and Ji Y:
Salidroside regulates imbalance of Th17/Treg and promotes ischemic
tolerance by targeting STAT-3 in cerebral ischemia-reperfusion
injury. Arch Med Sci. 17:523–534. 2021.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Han SJ and Lee HT: Mechanisms and
therapeutic targets of ischemic acute kidney injury. Kidney Res
Clin Prac. 38:427–440. 2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Baltaci AK, Gokbudak H, Baltaci SB,
Mogulkoc R and Avunduk MC: The effects of resveratrol
administration on lipid oxidation in experimental renal
ischemia-reperfusion injury in rats. Biotech Histochem. 94:592–599.
2019.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Li Y, Zhong D, Lei L, Jia Y, Zhou H and
Yang B: Propofol prevents renal ischemia-reperfusion injury via
inhibiting the oxidative stress pathways. Cell Physiol Biochem.
37:14–26. 2015.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Qiao X, Li RS, Li H, Zhu GZ, Huang XG,
Shao S and Bai B: Intermedin protects against renal
ischemia-reperfusion injury by inhibition of oxidative stress. Am J
Physiol Renal Physiol. 304:F112–F119. 2013.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Manning BD and Toker A: AKT/PKB signaling:
Navigating the network. Cell. 169:381–405. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Singh CK, Chhabra G, Ndiaye MA, Siddiqui
IA, Panackal JE, Mintie CA and Ahmad N: Quercetin-resveratrol
combination for prostate cancer management in TRAMP mice. Cancers
(Basel). 12(2141)2020.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Ahsan A, Liu M, Zheng Y, Yan W, Pan L, Li
Y, Ma S, Zhang X, Cao M, Wu Z, et al: Natural compounds modulate
the autophagy with potential implication of stroke. Acta Pharm Sin
B. 11:1708–1720. 2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Zhu L, Liu Z, Ren Y, Wu X, Liu Y, Wang T,
Li Y, Cong Y and Guo Y: Neuroprotective effects of salidroside on
ageing hippocampal neurons and naturally ageing mice via the
PI3K/Akt/TERT pathway. Phytother Res. 35:5767–5780. 2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Wu Y, Jiao H, Yue Y, He K, Jin Y, Zhang J,
Zhang J, Wei Y, Luo H, Hao Z, et al: Ubiquitin ligase E3 HUWE1/MULE
targets transferrin receptor for degradation and suppresses
ferroptosis in acute liver injury. Cell Death Differ. 29:1705–1718.
2022.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Su L, Jiang X, Yang C, Zhang J, Chen B, Li
Y, Yao S, Xie Q, Gomez H, Murugan R and Peng Z: Pannexin 1 mediates
ferroptosis that contributes to renal ischemia/reperfusion injury.
J Biol Chem. 294:19395–19404. 2019.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Li C, Sun G, Chen B, Xu L, Ye Y, He J, Bao
Z, Zhao P, Miao Z, Zhao L, et al: Nuclear receptor coactivator
4-mediated ferritinophagy contributes to cerebral ischemia-induced
ferroptosis in ischemic stroke. Pharmacol Res.
174(105933)2021.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Fang X, Wang H, Han D, Xie E, Yang X, Wei
J, Gu S, Gao F, Zhu N, Yin X, et al: Ferroptosis as a target for
protection against cardiomyopathy. Proc Natl Acad Sci USA.
116:2672–2680. 2019.PubMed/NCBI View Article : Google Scholar
|