Bruxism is a parafunctional condition affecting more
than two-thirds of the population at some point in their life;
however, only 25% of affected individuals are aware of this
condition and seek medical attention (1). According to an international
consensus, Bruxism is defined as repetitive jaw muscle activity
characterized by clenching or grinding of the teeth and/or bracing
or thrusting of the mandible; however, it is not regarded a
movement disorder or a sleep disorder in otherwise healthy
individuals (2). Bruxism can be
distinguished into awake and sleep bruxism based on the time of
occurrence (circadian manifestation). In a systematic review by
Manfredini et al (3), it
was shown that the prevalence of awake bruxism ranges from 22 to
31%, whereas sleep bruxism has a prevalence of 12.8%. Finally,
women are more prone to bruxism compared to men with a ratio of
5:1(4).
Although numerous factors have been implied to
contribute to the etiopathogenesis of bruxism, stress and emotional
disturbances are the most commonly accepted (5,6).
Emotional stress is associated with an increase in head and muscle
tonicity, as well as an increase in non-functional muscle activity
(7,8). In addition, the activation of the
sympathetic system in response to stressful stimuli leads to
increased muscle tone and reduced pain thresholds (8,9). In
a systematic review and meta-analysis published in 2021, Fritzen
et al (10) verified
increased levels of salivary cortisol in bruxists. These increased
levels correlate with strong circadian rhythms with peak levels
during the activation periods (11). Under experimental stress, there is
increased masseter activity, which returns to baseline levels upon
relaxation (12). In animal
studies, stress-induced muscle hyperactivity was associated with
muscle dysfunction and pain (13),
whereas humans who experience panic attacks more frequently exhibit
tooth clenching, bruxism and nail-biting (14). Overactivation of the sympathetic
system and hypothalamic-pituitary-adrenal axis (HPA) axis can make
an individual more vulnerable to new stress, as evidenced by animal
and human studies (15,16). Patients with temporomandibular
disorders (TMD) exhibit higher levels of anxiety (17), while TMD symptoms worsen whenever a
person is under stress (18,19).
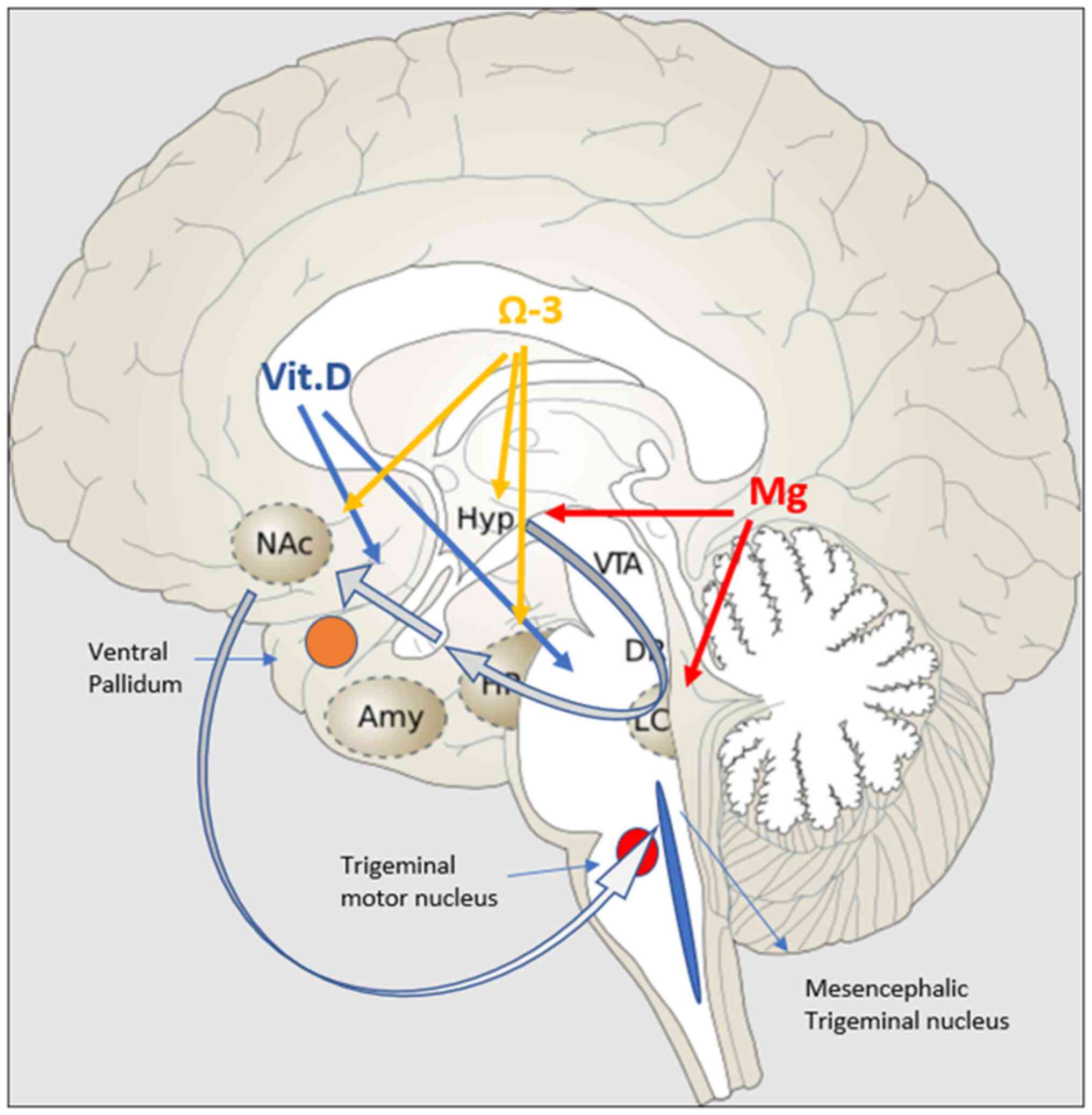
Two important neuronal areas are implicated in the
genesis of bruxism and are highly affected by stress: The
mesencephalic trigeminal nucleus (Me5) and the mesocortical
dopaminergic pathway. Me5 controls the masseter inhibitory reflex,
which, when activated, suppresses the trigeminal motor nucleus and
inhibits masseter and temporalis contraction. The mesocortical
dopaminergic tract connects the VTA with the nucleus accumbens
(N.Acc) via either the ventral subiculum (vSub) or amygdala
(36). N.Acc receives
glutamatergic projections from the hippocampus and basolateral
amygdala (BLA) and sends projections to mesencephalic motor
effector sites, and is therefore considered to be a motor-limbic
interface (37). The mesocortical
tract is equally important because it inhibits undesired muscular
movements (38). In acute mild
stressors, the tract follows the vSub-ventral pallidum (VP)-N.Acc
route (39,40). The hippocampus is essential in
counteracting overactivation of the HPA axis (41). However, in chronic stressors, the
hippocampus presents with neurodegenerative alterations that
attenuate this pathway, while at the same time, they activate the
BLA-VP-N.Acc pathway (42).
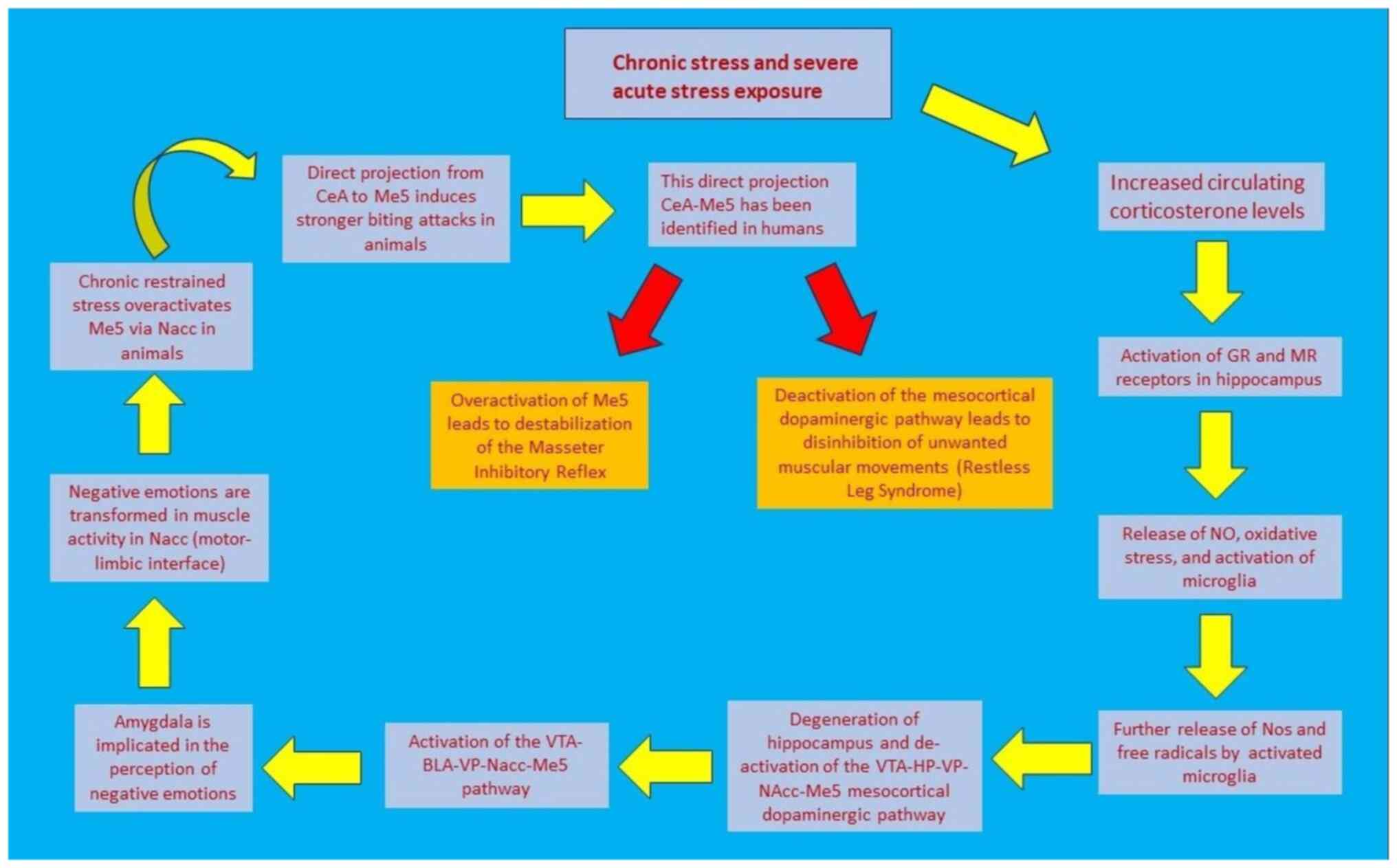
Studies in mice have revealed that a direct projection from CeA to
trigeminal motor nucleus is highly activated during hunting of prey
and that it promotes strong biting attacks (43). Of note, the same pathway has
recently been identified in humans, and in higher distribution than
BLA-Mo5 (Trigeminal Motor Nucleus) (44), further highlighting that the same
association may also apply to humans and warranting further
research in this field. Fig. 1
diagrammatically represents the effect of certain nutrients on the
mesocortical dopaminergic pathway.
Chronic stress exposure and increased circulating
levels of corticosterone impair progenitor cell proliferation,
inhibit neuronal differentiation and suppress cell survival in the
hippocampal dentate gyrus (45-49),
events that affect hippocampal neurogenesis (50-52).
This is happening as a result of oxidative stress due to
auto-oxidation of catecholamines (53). Oxidative stress due to increased
cellular activity induces nitrogen oxide (NO) production, which
activates microglia. The latter further releases NO and reactive
oxygen species (ROS) that induce neuroinflammation and
neurodegeneration (54,55). This in turn negatively modulates
the ability of the hippocampus to control hyperactivity of the HPA
axis. Neurodegeneration of the hippocampus and attenuation of the
vSub-VP-N.Acc pathway invoke alterations in the mesocortical
dopaminergic neurons that are implicated in neuropathic and chronic
pain (56-58).
The dysregulation of dopamine D2 receptor expressing indirect
pathway output neurons promotes hypersensitivity to pain and
increased impulsivity due to reduced levels of dopamine in N.Acc
(59,60). The attenuation of the vSub-VP-N.Acc
pathway and the activation of the BLA-VP-N.Acc cause further
reduction of dopamine neurons in the VTA (42). All these aforementioned events have
a strong effect on Me5. Animal electrophysiological studies have
shown that chronic restrained stress induces an increase in the
excitability of Me5 and glutamatergic neurotransmission from Me5 to
Mo5(61). This was evidenced by
increased levels of acetylcholinesterase (Ache) and creatine kinase
(CK-MM) in the masseter muscles (61). In another study by Ueno et
al (62), it was demonstrated
that activation of the ventral part of amygdala in guinea pigs may
cause rhythmic jaw movement.
Recent evidence has revealed similar degenerative
findings in bruxists. Keskinruzgar et al (63), by studying the retinas of bruxists,
found that retinal nerve fiber layer (axon) thickness, inferior
parietal lobe (dendrites) and granule cell layer (soma) volume are
significantly reduced. The retina is considered to be a
continuation of the brain; therefore, any neurodegenerative changes
in the retina are representative of those induced in the
mesocortical dopaminergic tract. In the same study, an increase in
choroidal thickness was also demonstrated. The choroid is an
important site of vascularization and is responsible for transport
of nutrients and oxygen (63).
This study, however, does not specify the cause of
neurodegeneration or an association between bruxism and
neurodegeneration. The fact that bruxism is a common comorbid
condition or symptom in other neurodegenerative disorders such as
Parkinson's disease or Parkisonism (64), as well as in rapid eye movement
sleep behavior disorder (65),
constitutes enough evidence so as to not exclude neurodegeneration
from this equation. Ozcan-Kucuk et al (66) concluded that there is a significant
increase in oxidative stress and prolidase activity in bruxists.
Increased levels of prolidase, an enzyme found in the plasma and
several organs, including the brain, are associated with higher
levels of proline, a brain neurotransmitter, which can induce
oxidative stress and subsequently neurodegeneration (67). These effects have been examined in
anxiety (68), several
musculoskeletal disorders (e.g. ankylosis, osteoarthritis)
(69,70) and neuropsychiatric diseases
(Alzheimer's, bipolar disorder) (67,71).
Fig. 2 illustrates the sequence of
events that connect stress with jaw muscle activity.
Based on these findings, particularly on the
neurodegeneration of the hippocampus, the attenuation of the
vSub-VP-N.Acc pathway and the fact that a malfunction may occur at
any given point in the mesocortical dopaminergic tract, it is
difficult to identify a therapeutic target. According to a
systematic review from 2022 by Minakuchi et al (72), none of the already used treatment
modalities is able to achieve universal positive results. As far as
pharmacologic agents are concerned, the evidence of efficacy has
low to moderate confidence, with numerous adverse effects; at the
same time, the efficacy of cognitive behavioral therapy is still
low and does not appear to improve the symptoms in <6 months
(72). Overwhelming evidence
points towards insufficiencies and deficiencies of necessary
elements for the homeostasis and normal function of the central
nervous system (CNS), with vitamin D (vit.D), magnesium and omega-3
fatty acids being the most studied ones.
Vit.D) is a very important nutrient, not only for
maintaining a healthy musculoskeletal system but also for the
positive effect on the nervous system. It has long been known that
vit.D has a vital role in calcium and phosphorous metabolism, in
three main ways: i) By enhancing intestinal absorption, ii) by
controlling the synthesis of parathyroid hormone and iii) by
promoting the maturation of pre-osteoclasts into osteoclasts
(73-75).
However, current evidence suggests that vit.D also has a major role
in neuroimmune modulation, neuroplasticity and neurological
oxidative stress reduction (76).
This property is supported by the abundance of vit.D receptors
throughout the CNS (77), as well
as by its ability to cross the blood brain barrier (78). Vit.D is also known to control ~3%
of the human genome via the vit.D receptor (VDR), which forms a
heterodimer with the retinoid X receptor (79,80).
This heterodimer complex binds to specific DNA sequences and alters
the expression of genes that are involved in glutamatergic and
GABAergic neurotransmission, calcium regulation and also the
expression of neurotrophic factors and genes involved in
neuroprotection (81,82).
The strong antioxidant capacity of vit.D helps to
improve brain enzyme activity and to reduce lipid peroxidation.
Oxidative stress has a major role in neurodegenerative diseases,
mainly through auto-oxidation of catecholamines (53,54).
Mitochondrial dysfunction leads to a rise in ROS and activated
microglia, resulting in nitric oxide (NO) and ROS production during
neuroinflammation (55). Vit.D
also protects the brain by suppressing inflammatory cytokines such
as IL-6(83). However, the most
important function of vit.D is the protection of dopaminergic
neurons. Although the mechanism has remained to be fully
elucidated, there is sufficient evidence to suggest that it can
affect calcium metabolism, apoptosis, inflammation,
immunomodulation, detoxification and neurotrophin upregulation
(84-86).
Cholecalciferol (VD3) may directly modulate tyrosine hydroxylase, a
rate-limiting enzyme in dopamine synthesis, as evidenced by
immunohistochemical staining. The glial cell-derived neurotrophic
factor (GDNF) protein also appears to be increased by these
effects, thereby supporting the neuroprotective effects of VD3 on
dopaminergic neurons (87). Orme
et al (87) demonstrated
that adding VD3 to mesencephalic cultures of dopaminergic neurons
increased their number and upregulated GDNF. Furthermore, VD3 has
also been shown to regulate factors involved in dopaminergic system
ontogeny (88), as evidenced
through studies in animals with altered dopaminergic metabolism due
to VD3 deficiency, where VD3 supplementation resulted in a
reduction of the degree of dopaminergic denervation (87,89).
Magnesium (Mg) is the body's fourth most abundant
cation and the second most abundant intracellularly; aerobic and
anaerobic metabolism, bioenergetic reactions, metabolic pathway
regulation, signal transduction, ion channel activity, cell
proliferation, differentiation, apoptosis, angiogenesis and
membrane stabilization are all biological processes that involve
magnesium (113-115).
More than 325 enzymes, many of which are specific to the nervous
system, are Mg-dependent, highlighting the importance of this
particular mineral in CNS physiological function (116).
Stress is known to promote oxidative stress through
catecholamine auto-oxidation: it aggravates lipid peroxidation,
increases DNA oxidative damage marker production and decreases
plasma anti-oxidative activity (117,118). During a stressful event, there is
activation of the HPA axis and release of corticotropin-releasing
factor (CRF) from parvocellular neurons. Mg antagonizes the
glutamate-stimulated CRF release, stabilizes CRF receptor binding
and stimulates the Na/K ATPase, which decreases CRF-receptor
activity (119-121).
Mg also reduces adrenocorticotropic hormone release by suppressing
HPA-axis activity via angiotensin II antagonism (122).
Mg has been shown to have a direct suppressive
effect on locus coeruleus (LC) activity and low Mg levels seem to
increase sensitivity to stress (123). The release of substance P, a
neuropeptide that preferentially activates tachykinin NK1
receptors, can explain many of the effects of Mg deficiency in
stress situations: Low Mg concentrations reduce Mg-gated blockade
of N-methyl-D-aspartate (NMDA) receptor channels, resulting in the
release of substance P and calcitonin gene-related peptide (CGRP)
from sensory C fibers, i.e. neuropeptides that are involved in the
production of reactive oxygen and nitrogen species and in the
induction of neurogenic inflammation that is characterized by an
increase in inflammatory cells and cytokines (124,125).
Magnesium is also essential for the synthesis of
kynureninase. Therefore, in cases of hypomagnesemia, increased
levels of kynurenines are observed in the circulation, which in
turn promotes anxiety, an increase in the release of noradrenaline,
reduced levels of serotonin, GABA receptor blockade and locomotor
hyperactivity (126). The
increased neural excitability observed in hypomagnesemia is
believed to occur from the increased activity of NMDA receptors,
which in turn induces anxiety, increased dopamine release and
modulation of serotonin receptors (127).
Magnesium deficiency reduces the activity of B6 by
inhibiting alkaline phosphatase, an enzyme necessary for the
transformation of the active form of B6, pyridoxal phosphate. This
causes increased levels of kynurenines, increased sympathetic
activation and increased sensitivity to glucorticoids (128). As B6 is known to participate in
decarboxylation of glutamic acid to GABA, of DOPA to dopamine and
of 5-hydroxytryptophan to serotonin, it is regarded as a
neuroprotective and antitoxic agent (129).
Mg deficiency symptoms include neuromuscular
irritability and weakness, headaches, hyperemotionality,
generalized anxiety, insomnia, and asthenia (116,133,134). In Mg-deficient animals, there are
increased plasma corticosterone levels, increased irritability and
aggressive behavior (123,135).
Experimentally-induced Mg deficiency in animal studies has been
associated with disrupted sleep patterns, while an increase in the
amplitude of daily variation of sleep and slow-wave sleep delta
power has also been observed (135). Chronic sleep deprivation in
humans is associated with progressively decreasing intracellular Mg
levels, reduced duration of cardiopulmonary exercise and increased
hypersensitivity to sympathetic nervous stimulation (136). In a study by Sarchielli et
al (137), patients who
suffered from migraine and tension-type headaches, very common
symptoms in patients with bruxism and TMD, presented with
significantly lower levels of serum and salivary Mg. This is
because hypomagnesaemia makes cerebral arteries more sensitive to
CO2, which promotes cerebral vasospasm and headache
(138,139). Hypomagnesaemia is also linked to
exacerbated neural excitability, migraine, orofacial tardive
dyskinesia and increased anxiety, symptoms that may be improved by
combined supplementation of Mg and B6(140).
Overall, Mg has a strong neuroprotective role. As
mentioned earlier, stress has a deleterious effect on neurons
because of the increased levels of IL-6 that initiate microglial
activation. Increased inducible NO synthase levels that are
expressed during stressful periods impair hippocampal neurogenesis
(142,143). Mg deficiency increases neuronal
calcium influx and, as a result, NO production, which has been
linked to cytotoxic effects (116). Mg counteracts the effects of
calcium and reduces ROS production via the phospholipase
lipoxygenase and cyclooxygenase pathways (144).
Iron, although not having been linked to bruxism,
has recently been associated with restless leg syndrome (RLS), a
common neurologic disorder characterized by involuntary muscle
movement, which is associated with malfunction of the mesocortical
dopaminergic pathway (145). This
pathway, as discussed earlier, may be implicated in the
pathogenesis of sleep (central) bruxism. Some consider bruxism as a
manifestation of RLS, based on the fact that ~40% of patients with
RLS report a history of bruxism (146), while others assume that bruxism
and RLS are comorbid conditions (147). Both entities share some common
features as they are both observed in non-REM sleep and are linked
to basal ganglia dysfunction (148).
Brain iron deficiency has a critical role in the
pathogenesis of RLS. Anemia and pregnancy are two conditions
characterized by systemic iron deficiency, which are highly
associated with RLS symptoms (149-151).
Similar to bruxists, females are more susceptible to RLS. In a
study by Umbreit (152), it was
found that RLS without anemia exhibits a sex-dependent preference.
Patients with RLS with and without anemia present with more
profound tiredness, poorer sleep quality and less energy during the
day (151). Depleted iron stores
are associated with decreased activity of iron-dependent enzymes,
reduced cellular oxidative capacity, as well as decreased energy
efficiency (153,154). Previous clinical studies have
shown that peripheral iron deficiency increases the risk of RLS
prevalence (151). Reduced serum
iron concentrations reduce brain iron levels, and iron therapy
benefits patients with RLS with low peripheral iron levels
(155). In a systematic review
and meta-analysis, it was shown that iron therapy, in a way similar
to that of a dopamine agonist, reduces restlessness and RLS
severity (156).
Both human and animal studies have demonstrated that
stress exposure causes a reduction in serum iron ranging from 27 to
44%, a reduction in ferritin by 24% and and a reduction in
hemoglobin by 12.5% (157,158).
Although omega-3 fatty acids have not been directly
linked with bruxism, they have been highly implicated in anxiety
and emotional mood disorders (159), as well as in neuroinflammation
and neurodegeneration (159-162).
They are esterified into phospholipids in the sn-2 position of the
cell membrane (160), playing a
key role in the structure and function of the brain cell membrane.
Overwhelming evidence signifies their involvement in hippocampal
development and synaptic function (162).
Omega-3 fatty acids may be found in numerous brain
regions, but more abundantly in the prefrontal cortex, hippocampus
and hypothalamus (163). They
have been classified into n-6 polyunsaturated fatty acids (PUFA)
and n-3 PUFA, with their main metabolites being arachidonic acid
and docosahexaenoic acid (DHA), respectively. The free forms of
PUFA are transformed into specific derivatives, such as
eicosanoids, specialized proresolving mediators (SPM) and
endocannabinoids (eCBs), which strongly regulate inflammation
(160). The eCBs are lipids
produced by the membrane's fatty acids after neuronal stimulation
and bind to cytochrome P450 carbonyl reductase 1(164). Activation of this receptor causes
an inhibition in glutamate and GABA release from presynaptic
neurons (165). Regulation of
eCBs in the brain by PUFA has a strong impact on hippocampal
synaptic plasticity and in eCB-dependent plasticity (166), while CB1-associated signaling
pathways in the prefrontal cortex (PFC) and N.Acc are also
affected. Another signaling pathway, GPR120, which functions as an
omega-3 fatty acid receptor, signals DHA activity that is highly
prominent in the hypothalamus and N.Acc (167), areas that are closely linked to
the pathogenesis of anxiety and bruxism, as discussed earlier.
Oxidative stress causes NO production, which
activates microglia. The latter are glial cells of myeloid origin,
important in maintaining brain homeostasis and in protecting nerve
cells (173). Activation of
microglia causes further production of NO and ROS, leading to
neuroinflammation (174). DHA
deficiency induces abnormal microglial activation through changes
in their membrane fluidity and the modulation of several
proinflammatory transcription factors (175). These changes are gender- and
area-dependent. Adult females have more microglia than males in the
paraventricular nucleus of the hypothalamus, amygdala and
hippocampus, particularly in the dentate gyrus (175), which has a crucial role in
controlling the HPA axis. This is probably the reason why females
appear to be more vulnerable to stress and more prone to
bruxism.
Finally, noteworthy findings have also been obtained
in individuals with morning headaches, a common symptom in
bruxists. Morning headaches usually exhibit the characteristics of
migraine (182). Numerous
hypotheses have been made regarding the mechanism of their genesis,
with oxidative stress and activation of the trigeminovascular
pathway being the two most prevalent (183). In this context, the role of CGRP
has been highlighted both as a mediator and a potential therapeutic
target (184). First of all, the
release of CGRP has also been shown to be regulated by oxylipin
receptors in the trigeminal nerve endings and central pain
processing pathways (185,186).
Furthermore, a recent study by Marchetti et al (184) showed that supplementation of
PUFAs at a ratio of 1.5 omega-6/1 omega-3 induced a marked
reduction in migraine symptoms, frequency and pain intensity; the
explanation given by the authors was that EPA, DHA and their SPM
activate innate and adaptive immune systems, remove ROS, and
inhibit COX-2 and NF-κB intracellular signaling pathways. Table I summarizes the main
characteristics of the nutrients discussed in this section and
evidence that highlights their potential implication in the
etiopathogenesis of bruxism.
This review has discussed the effects of chronic
stress exposure in the pathogenesis of bruxism. In particular,
oxidative stress, through the release of NO and the activation of
microglia, has been shown to lead to neurodegeneration of the
hippocampus and destabilization of the mesocortical dopaminergic
pathway. It has also been highlighted that the former impairs the
ability to counteract HPA axis overactivity, while the latter
results in loss of inhibition of unwanted (involuntary) muscle
movements. Based on recent evidence, it appears that attenuation of
the vSub-VP-N.Acc route leads to activation of the BLA-VP-N.Acc
pathway. The prevailing route has a strong yet opposing effect on
the Me5, a nucleus that inhibits the involuntary contraction of
masseter muscles by controlling the masseter inhibitory reflex.
Indeed, activation of the amygdala appears to induce rhythmic jaw
activity, while overactivation of Me5 under chronic stress exposure
causes overactivation of Mo5, as evidenced by increased levels of
CK-MM and pain in masseter muscles (61).
This report, instead of focusing on pharmacological
agents, has shifted the attention towards homeostatic disturbances
and deficiencies in important elements, such as vit.D, magnesium,
omega-3 fatty acids, and, to a lesser degree, iron. Their
neuroprotective properties, their capacity to reduce oxidative
stress to suppress the HPA and LC axes, and their positive action
on the neuroplasticity and neuronal growth of the hippocampus,
particularly in the case of omega-3, further support this notion.
Conversely, a decrease in the levels of these elements in the brain
is associated with neurodegeneration, increased anxiety, increased
neuromuscular excitability, bruxism, muscle spasms and pain. Of
note, appropriate and individualized supplementation of these
nutrients appears to reduce or alleviate the neurological and
musculoskeletal symptoms (108,109). Future studies should focus on
whether these concentration alterations have a causal effect or
whether they constitute the aftermath of chronic stress exposure,
and if re-establishment of the nutrient balance has a significant
impact on the pathogenesis and chronicity of bruxism.
The present review has certain limitations.
Regarding the pathogenesis of bruxism, the articles included
constitute a small number of cases, with self-reporting subjective
symptoms, which makes it difficult to provide a definite diagnosis.
On the other hand, the actual levels of Mg and omega-3 fatty acids
in the patients' circulation (plasma) and brain may not be easily
assessed. In addition, patient variability is probably the reason
behind the inconsistency in achieving the desired outcome in
affected individuals, while there is also a lack of data regarding
the optimum duration of nutrient supplementation to achieve the
desired outcome. However, despite the small number of studies
addressing these issues and their associated limitations, the
existing evidence supports the rationale that subsequent research
should be conducted in this field, with the aim to further enhance
the current understanding of this complex, multifactorial condition
and hopefully to provide the affected individuals with alternative
and more effective therapeutic modalities.
This article is a part of and constitutes a
(partial) requirement for the fellowship program on
temporomandibular joint (TMJ) disorders of the TMJ Foundation, TMJ
Consultancy Services (Bhopal, India) and DARSN Academy for
Maxillofacial Education and Research (Damer, India).
Funding: No funding was received.
Not applicable.
Conceptualization: IAP. Writing and original draft
preparation: IAP and MA. Review and editing: MA, VZ and
DAS;.Visualization: IAP and MA. Supervision: VZ and MA. All authors
have read and approved the final version of the manuscript. Data
authentication is not applicable.
Not applicable.
Not applicable.
IAP, MA and VZ declare that they have no competing
interests. DAS is the Editor-in-Chief for the journal, but had no
personal involvement in the reviewing process, or any influence in
terms of adjudicating on the final decision for this article.
|
1
|
Simoes WA: Occlusal plane: A clinical
evaluation. J Clin Pediatr Dent. 19:75–81. 1995.PubMed/NCBI
|
|
2
|
Lobbezoo F, Ahlberg J, Raphael KG,
Wetselaar P, Glaros AG, Kato T, Santiago V, Winocur E, De Laat A,
De Leeuw R, et al: International consensus on the assessment of
bruxism: Report of a work in progress. J Oral Rehabil. 45:837–844.
2018.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Manfredini D, Winocur E, Guarda-Nardini L,
Paesani D and Lobbezoo F: Epidemiology of bruxism in adults: A
systematic review of the literature. J Orofac Pain. 27:99–110.
2013.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Manfredini D, Piccotti F, Ferronato G and
Guarda-Nardini L: Age peaks of different RDC/TMD diagnoses in a
patient population. J Dent. 38:392–399. 2010.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Segall SK, Maixner W, Belfer I, Wiltshire
T, Seltzer Z and Diatchenko L: Janus molecule I: Dichotomous
effects of COMT in neuropathic vs. nociceptive pain modalities. CNS
Neurol Disord Drug Targets. 11:222–235. 2012.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Cruz-Fierro N, Martinez-Fierro M,
Cerda-Flores RM, Gómez-Govea MA, Delgado-Enciso I,
Martínez-De-Villarreal LE, González-Ramírez MT and
Rodríguez-Sánchez IP: The phenotype, psychotype and genotype of
bruxism. Biomed Rep. 8:264–268. 2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Carlson CR, Okeson JP, Falace DA, Nitz AJ,
Curran SL and Anderson D: Comparison of psychologic and physiologic
functioning between patients with masticatory muscle pain and
matched controls. J Orofac Pain. 7:15–22. 1993.PubMed/NCBI
|
|
8
|
Fillingim RB, Ohrbach R, Greenspan JD,
Knott C, Diatchenko L, Dubner R, Bair E, Baraian C, Mack N, Slade
GD and Maixner W: Psychological factors associated with development
of TMD: The OPPERA prospective cohort study. J Pain. 14 (12
Suppl):T75–T90. 2013.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Bertoli E, de Leeuw R, Schmidt JE, Okeson
JP and Carlson CR: Prevalence and impact of post-traumatic stress
disorder symptoms in patients with masticatory muscle or
temporomandibular joint pain: Differences and similarities. J
Orofac Pain. 21:107–119. 2007.PubMed/NCBI
|
|
10
|
Fritzen VM, Colonetti T, Cruz MVB, Ferraz
SD, Ceretta L, Tuon L, DA Rosa MI and Ceretta RA: Levels of
salivary cortisol in adults and children with bruxism diagnosis: A
systematic review and meta-analysis. J Evid Based Dent Pract.
22(101634)2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chung S, Son GH and Kim K: Circadian
rhythm of adrenal glucocorticoid: Its regulation and clinical
implications. Biochim Biophys Acta. 1812:581–591. 2011.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Tsai CM, Chou SL, Gale EN and McCall WD
Jr: Human masticatory muscle activity and jaw position under
experimental stress. J Oral Rehabil. 29:44–51. 2002.PubMed/NCBI View Article : Google Scholar
|
|
13
|
de Leeuw JR, Steenks MH, Ros WJ,
Lobbezoo-Scholte AM, Bosman F and Winnubst JA: Multidimensional
evaluation of craniomandibular dysfunction. I: Symptoms and
correlates. J Oral Rehabil. 21:501–514. 1994.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Manfredini D and Lobbezoo F: Role of
psychosocial factors in the etiology of bruxism. J Orofac Pain.
23:153–166. 2009.PubMed/NCBI
|
|
15
|
Miyake H, Mori D, Katayama T, Fujiwara S,
Sato Y, Azuma K and Kubo KY: Novel stress increases
hypothalamic-pituitary-adrenal activity in mice with a raised bite.
Arch Oral Biol. 68:55–60. 2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Aizawa H, Cui W, Tanaka K and Okamoto H:
Hyperactivation of the habenula as a link between depression and
sleep disturbance. Front Hum Neurosci. 7(826)2013.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Gameiro GH, da Silva Andrade A, Nouer DF
and Ferraz de Arruda Veiga MC: How may stressful experiences
contribute to the development of temporomandibular disorders? Clin
Oral Investig. 10:261–268. 2006.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Liu X, Zhou KX, Yin NN, Zhang CK, Shi MH,
Zhang HY, Wang DM, Xu ZJ, Zhang JD, Li JL and Wang MQ: Malocclusion
generates anxiety-like behavior through a putative lateral
habenula-mesencephalic trigeminal nucleus pathway. Front Mol
Neurosci. 12(174)2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Suvinen TI, Hanes KR, Gerschman JA and
Reade PC: Psychophysical subtypes of temporomandibular disorders. J
Orofac Pain. 11:200–205. 1997.PubMed/NCBI
|
|
20
|
Lavigne GJ, Kato T, Kolta A and Sessle BJ:
Neurobiological mechanisms involved in sleep bruxism. Crit Rev Oral
Biol Med. 14:30–46. 2003.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Blanchet PJ, Rompre PH, Lavigne GJ and
Lamarche C: Oral dyskinesia: A clinical overview. Int J
Prosthodont. 18:10–19. 2005.PubMed/NCBI
|
|
22
|
Clark GT and Ram S: Four oral motor
disorders: Bruxism, dystonia, dyskinesia and drug-induced dystonic
extrapyramidal reactions. Dent Clin North Am. 51:225–243, viii-ix.
2007.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Kwak YT, Han IW, Lee PH, Yoon JK and Suk
SH: Associated conditions and clinical significance of awake
bruxism. Geriatr Gerontol Int. 9:382–390. 2009.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Winocur E, Gavish A, Voikovitch M,
Emodi-Perlman A and Eli I: Drugs and bruxism: A critical review. J
Orofac Pain. 17:99–111. 2003.PubMed/NCBI
|
|
25
|
Garrett AR and Hawley JS: SSRI-associated
bruxism: A systematic review of published case reports. Neurol Clin
Pract. 8:135–141. 2018.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Bendtsen L, Jensen R and Olesen J:
Amitriptyline, a combined serotonin and noradrenaline re-uptake
inhibitor, reduces exteroceptive suppression of temporal muscle
activity in patients with chronic tension-type headache.
Electroencephalogr Clin Neurophysiol. 101:418–422. 1996.PubMed/NCBI
|
|
27
|
Falisi G, Rastelli C, Panti F, Maglione H
and Quezada Arcega R: Psychotropic drugs and bruxism. Expert Opin
Drug Saf. 13:1319–1326. 2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Okamoto K, Imbe H, Tashiro A, Kimura A,
Donishi T, Tamai Y and Senba E: The role of peripheral 5HT2A and
5HT1A receptors on the orofacial formalin test in rats with
persistent temporomandibular joint inflammation. Neuroscience.
130:465–474. 2005.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Nakanishi O and Ishikawa T: Involvement of
peripheral 5-HT2A receptor activation in inflammatory pain. Nihon
Rinsho. 59:1675–1680. 2001.PubMed/NCBI(In Japanese).
|
|
30
|
Chaouloff F: Regulation of 5-HT receptors
by corticosteroids: Where do we stand? Fundam Clin Pharmacol.
9:219–233. 1995.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Lopez JF, Vazquez DM, Chalmers DT and
Watson SJ: Regulation of 5-HT receptors and the
hypothalamic-pituitary-adrenal axis. Implications for the
neurobiology of suicide. Ann N Y Acad Sci. 836:106–134.
1997.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Yeung LY, Kung HF and Yew DT: Localization
of 5-HT1A and 5-HT2A positive cells in the brainstems of control
age-matched and Alzheimer individuals. Age (Dordr). 32:483–495.
2010.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Inan R, Senel GB, Yavlal F, Karadeniz D,
Gunduz A and Kiziltan ME: Sleep bruxism is related to decreased
inhibitory control of trigeminal motoneurons, but not with
reticulobulbar system. Neurol Sci. 38:75–81. 2017.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Lauria G, Majorana A, Borgna M, Lombardi
R, Penza P, Padovani A and Sapelli P: Trigeminal small-fiber
sensory neuropathy causes burning mouth syndrome. Pain.
115:332–337. 2005.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Bostwick JM and Jaffee MS: Buspirone as an
antidote to SSRI-induced bruxism in 4 cases. J Clin Psychiatry.
60:857–860. 1999.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Belujon P and Grace AA: Regulation of
dopamine system responsivity and its adaptive and pathological
response to stress. Proc Biol Sci. 282(20142516)2015.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Floresco SB: Dopaminergic regulation of
limbic-striatal interplay. J Psychiatry Neurosci. 32:400–411.
2007.PubMed/NCBI
|
|
38
|
Dunlop BW and Nemeroff CB: The role of
dopamine in the pathophysiology of depression. Arch Gen Psychiatry.
64:327–337. 2007.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Belujon P and Grace AA: Critical role of
the prefrontal cortex in the regulation of hippocampus-accumbens
information flow. J Neurosci. 28:9797–9805. 2008.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Floresco SB, Blaha CD, Yang CR and
Phillips AG: Modulation of hippocampal and amygdalar-evoked
activity of nucleus accumbens neurons by dopamine: Cellular
mechanisms of input selection. J Neurosci. 21:2851–2860.
2001.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Herman JP and Mueller NK: Role of the
ventral subiculum in stress integration. Behav Brain Res.
174:215–224. 2006.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Valenti O, Gill KM and Grace AA: Different
stressors produce excitation or inhibition of mesolimbic dopamine
neuron activity: Response alteration by stress pre-exposure. Eur J
Neurosci. 35:1312–1321. 2012.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Han W, Tellez LA, Rangel MJ Jr, Motta SC,
Zhang X, Perez IO, Canteras NS, Shammah-Lagnado SJ, van den Pol AN
and de Araujo IE: Integrated control of predatory hunting by the
central nucleus of the amygdala. Cell. 168:311–324 e318.
2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Kaya B, Geha P, de Araujo I, Cioffi I and
Moayedi M: Identification of central amygdala and trigeminal motor
nucleus connectivity in humans: An ultra-high field diffusion MRI
study. Hum Brain Mapp. 44:1309–1319. 2023.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Kubo KY, Iinuma M and Chen H: Mastication
as a stress-coping behavior. Biomed Res Int.
2015(876409)2015.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Chen H, Iinuma M, Onozuka M and Kubo KY:
Chewing maintains hippocampus-dependent cognitive function. Int J
Med Sci. 12:502–509. 2015.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Muramoto T, Takano Y and Soma K:
Time-related changes in periodontal mechanoreceptors in rat molars
after the loss of occlusal stimuli. Arch Histol Cytol. 63:369–380.
2000.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Mori D, Katayama T, Miyake H, Fujiwara S
and Kubo KY: Occlusal disharmony leads to learning deficits
associated with decreased cellular proliferation in the hippocampal
dentate gyrus of SAMP8 mice. Neurosci Lett. 534:228–232.
2013.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Mori D, Miyake H, Mizutani K, Shimpo K,
Sonoda S, Yamamoto T, Fujiwara S and Kubo KY: Effects of occlusal
disharmony on the hippocampal dentate gyrus in aged
senescence-accelerated mouse prone 8 (SAMP8). Arch Oral Biol.
65:95–101. 2016.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Azuma K, Ogura M, Kondo H, Suzuki A,
Hayashi S, Iinuma M, Onozuka M and Kubo KY: Maternal active
mastication during prenatal stress ameliorates prenatal
stress-induced lower bone mass in adult mouse offspring. Int J Med
Sci. 14:348–355. 2017.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Suzuki A, Iinuma M, Hayashi S, Sato Y,
Azuma K and Kubo KY: Maternal chewing during prenatal stress
ameliorates stress-induced hypomyelination, synaptic alterations,
and learning impairment in mouse offspring. Brain Res. 1651:36–43.
2016.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Onishi M, Iinuma M, Tamura Y and Kubo KY:
Learning deficits and suppression of the cell proliferation in the
hippocampal dentate gyrus of offspring are attenuated by maternal
chewing during prenatal stress. Neurosci Lett. 560:77–80.
2014.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Blesa J, Trigo-Damas I, Quiroga-Varela A
and Jackson-Lewis VR: Oxidative stress and Parkinson's disease.
Front Neuroanat. 9(91)2015.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Berridge MJ: Vitamin D and Depression:
Cellular and regulatory mechanisms. Pharmacol Rev. 69:80–92.
2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Tohari AM, Zhou X and Shu X: Protection
against oxidative stress by vitamin D in cone cells. Cell Biochem
Funct. 34:82–94. 2016.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Watanabe M, Narita M, Hamada Y, Yamashita
A, Tamura H, Ikegami D, Kondo T, Shinzato T, Shimizu T, Fukuchi Y,
et al: Activation of ventral tegmental area dopaminergic neurons
reverses pathological allodynia resulting from nerve injury or bone
cancer. Mol Pain. 14(1744806918756406)2018.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Baliki MN, Geha PY, Fields HL and Apkarian
AV: Predicting value of pain and analgesia: Nucleus accumbens
response to noxious stimuli changes in the presence of chronic
pain. Neuron. 66:149–160. 2010.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Martikainen IK, Nuechterlein EB, Pecina M,
Love TM, Cummiford CM, Green CR, Stohler CS and Zubieta JK: Chronic
back pain is associated with alterations in dopamine
neurotransmission in the ventral striatum. J Neurosci.
35:9957–9965. 2015.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Borsook D, Linnman C, Faria V, Strassman
AM, Becerra L and Elman I: Reward deficiency and anti-reward in
pain chronification. Neurosci Biobehav Rev. 68:282–297.
2016.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Ramdani C, Carbonnell L, Vidal F, Beranger
C, Dagher A and Hasbroucq T: Dopamine precursors depletion impairs
impulse control in healthy volunteers. Psychopharmacology (Berl).
232:477–487. 2015.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Zhao YJ, Liu Y, Wang J, Li Q, Zhang ZM, Tu
T, Lei R, Zhang M and Chen YJ: Activation of the mesencephalic
trigeminal nucleus contributes to masseter hyperactivity induced by
chronic restraint stress. Front Cell Neurosci.
16(841133)2022.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Ueno Y, Higashiyama M, Haque T, Masuda Y,
Katagiri A, Toyoda H, Uzawa N, Yoshida A and Kato T: Motor
representation of rhythmic jaw movements in the amygdala of guinea
pigs. Arch Oral Biol. 135(105362)2022.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Keskinruzgar A DDS, PhD Kalenderoglu A MD,
Yapici Yavuz G DDS, PhD Koparal M DDS, PhD Simsek A MD, Karadag AS
MD and Utkun M DDS: Investigation of neurodegenerative and
inflammatory processes in sleep bruxism. Cranio. 38:358–364.
2020.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Verhoeff MC, Koutris M, Berendse HW, van
Dijk KD and Lobbezoo F: Parkinson's disease, temporomandibular
disorder pain and bruxism and its clinical consequences: A protocol
of a single-centre observational outpatient study. BMJ Open.
12(e052329)2022.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Abe S, Gagnon JF, Montplaisir JY, Postuma
RB, Rompré PH, Huynh NT, Kato T, Kawano F and Lavigne GJ: Sleep
bruxism and oromandibular myoclonus in rapid eye movement sleep
behavior disorder: A preliminary report. Sleep Med. 14:1024–1030.
2013.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Ozcan-Kucuk A, Ege B, Koparal M, Gonel A
and Koyuncu I: Evaluation of the oxidative stress level and serum
prolidase activity in patients with sleep bruxism. Comb Chem High
Throughput Screen. 24:286–293. 2021.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Selek S, Altindag A, Saracoglu G, Celik H
and Aksoy N: Prolidase activity and its diagnostic performance in
bipolar disorder. J Affect Disord. 129:84–86. 2011.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Ercan AC, Bahceci B, Polat S, Cenker OC,
Bahceci I, Koroglu A, Sahin K and Hocaoglu C: Oxidative status and
prolidase activities in generalized anxiety disorder. Asian J
Psychiatr. 25:118–122. 2017.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Altay MA, Erturk C, Bilge A, Yapti M,
Levent A and Aksoy N: Evaluation of prolidase activity and
oxidative status in patients with knee osteoarthritis:
Relationships with radiographic severity and clinical parameters.
Rheumatol Int. 35:1725–1731. 2015.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Baspinar S, Kirnap M, Baspinar O, Dizdar
OS and Kocer D: Serum prolidase level in ankylosing spondylitis:
Low serum levels as a new potential gold standard biomarker for
disease activity. Rheumatol Int. 36:1609–1616. 2016.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Arikanoglu A, Akil E, Varol S, Yucel Y,
Yuksel H, Cevik MU, Palanci Y and Unan F: Relationship of cognitive
performance with prolidase and oxidative stress in Alzheimer
disease. Neurol Sci. 34:2117–2121. 2013.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Minakuchi H, Fujisawa M, Abe Y, Iida T,
Oki K, Okura K, Tanabe N and Nishiyama A: Managements of sleep
bruxism in adult: A systematic review. Jpn Dent Sci Rev.
58:124–136. 2022.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Holick MF: The vitamin D deficiency
pandemic: Approaches for diagnosis, treatment and prevention. Rev
Endocr Metab Disord. 18:153–165. 2017.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Brown AJ, Dusso AS and Slatopolsky E:
Vitamin D analogues for secondary hyperparathyroidism. Nephrol Dial
Transplant. 17 (Suppl 10):S10–S19. 2002.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Cranney A, Horsley T, O'Donnell S, Weiler
H, Puil L, Ooi D, Atkinson S, Ward L, Moher D, Hanley D, et al:
Effectiveness and safety of vitamin D in relation to bone health.
Evid Rep Technol Assess (Full Rep). 1–235. 2007.PubMed/NCBI
|
|
76
|
Berridge MJ: Vitamin D, reactive oxygen
species and calcium signalling in ageing and disease. Philos Trans
R Soc Lond B Biol Sci. 371(20150434)2016.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Koundourakis NE, Avgoustinaki PD,
Malliaraki N and Margioris AN: Muscular effects of vitamin D in
young athletes and non-athletes and in the elderly. Hormones
(Athens). 15:471–488. 2016.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Gascon-Barre M and Huet PM: Apparent
[3H]1,25-dihydroxyvitamin D3 uptake by canine and rodent brain. Am
J Physiol. 244:E266–E271. 1983.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Chau YY and Kumar J: Vitamin D in chronic
kidney disease. Indian J Pediatr. 79:1062–1068. 2012.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Bouillon R, Carmeliet G, Verlinden L, van
Etten E, Verstuyf A, Luderer HF, Lieben L, Mathieu C and Demay M:
Vitamin D and human health: Lessons from vitamin D receptor null
mice. Endocr Rev. 29:726–776. 2008.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Groves NJ, McGrath JJ and Burne TH:
Vitamin D as a neurosteroid affecting the developing and adult
brain. Annu Rev Nutr. 34:117–141. 2014.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Dimitrakis E, Katsarou MS, Lagiou M,
Papastefanopoulou V, Spandidos DA, Tsatsakis A, Papageorgiou S,
Moutsatsou P, Antoniou K, Kroupis C and Drakoulis N: Association of
vitamin D receptor gene haplotypes with late-onset Alzheimer's
disease in a Southeastern European Caucasian population. Exp Ther
Med. 24(584)2022.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Ikonen H, Palaniswamy S, Nordstrom T,
Järvelin MR, Herzig KH, Jääskeläinen E, Seppälä J, Miettunen J and
Sebert S: Vitamin D status and correlates of low vitamin D in
schizophrenia, other psychoses and non-psychotic depression-The
Northern Finland Birth Cohort 1966 study. Psychiatry Res.
279:186–194. 2019.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Wang JY, Wu JN, Cherng TL, Hoffer BJ, Chen
HH, Borlongan CV and Wang Y: Vitamin D(3) attenuates
6-hydroxydopamine-induced neurotoxicity in rats. Brain Res.
904:67–75. 2001.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Klotz B, Mentrup B, Regensburger M, Zeck
S, Schneidereit J, Schupp N, Linden C, Merz C, Ebert R and Jakob F:
1,25-dihydroxyvitamin D3 treatment delays cellular aging in human
mesenchymal stem cells while maintaining their multipotent
capacity. PLoS One. 7(e29959)2012.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Cass WA, Peters LE, Fletcher AM and Yurek
DM: Calcitriol promotes augmented dopamine release in the lesioned
striatum of 6-hydroxydopamine treated rats. Neurochem Res.
39:1467–1476. 2014.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Orme RP, Middleditch C, Waite L and
Fricker RA: The role of vitamin D(3) in the development and
neuroprotection of midbrain dopamine neurons. Vitam Horm.
100:273–297. 2016.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Pertile RA, Cui X and Eyles DW: Vitamin D
signaling and the differentiation of developing dopamine systems.
Neuroscience. 333:193–203. 2016.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Kesby JP, Cui X, O'Loan J, McGrath JJ,
Burne TH and Eyles DW: Developmental vitamin D deficiency alters
dopamine-mediated behaviors and dopamine transporter function in
adult female rats. Psychopharmacology (Berl). 208:159–168.
2010.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Huang JY, Arnold D, Qiu CF, Miller RS,
Williams MA and Enquobahrie DA: Association of serum vitamin D with
symptoms of depression and anxiety in early pregnancy. J Womens
Health (Larchmt). 23:588–595. 2014.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Altınbas A: The effect of vitamin D levels
on the mood disorders of the operating room and intensive care unit
staff. J Clin Anal Med. 10:156–161. 2019.
|
|
92
|
Al-Atram A, Raghunath G and Kannan SK: The
Relationship between Vitamin D and Mental Health among Dental
Students in Saudi Arabia: A Descriptive Cross-Sectional Study. J
Clin Diag Res. 14:VC07–VC10. 2020.
|
|
93
|
Silva MRM, Barros WMA, Silva MLD, Silva
JMLD, Souza APDS, Silva ABJD, Fernandes MSS, Souza SL and Souza
VON: Relationship between vitamin D deficiency and
psychophysiological variables: A systematic review of the
literature. Clinics (Sao Paulo). 76(e3155)2021.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Croll PH, Boelens M, Vernooij MW, van de
Rest O, Zillikens MC, Ikram MA and Voortman T: Associations of
vitamin D deficiency with MRI markers of brain health in a
community sample. Clin Nutr. 40:72–78. 2021.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Johnson MM, Patel S and Williams J: Don't
Take It ‘Lytely’: A case of acute tetany. Cureus.
11(e5845)2019.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Straube S, Derry S, Moore RA and McQuay
HJ: Vitamin D for the treatment of chronic painful conditions in
adults. Cochrane Database Syst Rev. 2010(CD007771)2010.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Alkhatatbeh MJ, Hmoud ZL, Abdul-Razzak KK
and Alem EM: Self-reported sleep bruxism is associated with vitamin
D deficiency and low dietary calcium intake: A case-control study.
BMC Oral Health. 21(21)2021.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Anjum I, Jaffery SS, Fayyaz M, Samoo Z and
Anjum S: The role of vitamin D in brain health: A mini literature
review. Cureus. 10(e2960)2018.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Allaf BAW and Abdul-Hak M: Association
between bruxism severity and serum concentrations of
25-hydroxyvitamin D levels. Clin Exp Dent Res. 8:827–835.
2022.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Garfinkel RJ, Dilisio MF and Agrawal DK:
Vitamin D and its effects on articular cartilage and
osteoarthritis. Orthop J Sports Med.
5(2325967117711376)2017.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Nowaczewska M, Wicinski M, Osinski S and
Kazmierczak H: The role of vitamin D in primary headache-from
potential mechanism to treatment. Nutrients. 12(243)2020.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Sohn JH, Chu MK, Park KY, Ahn HY and Cho
SJ: Vitamin D deficiency in patients with cluster headache: A
preliminary study. J Headache Pain. 19(54)2018.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Yildiz S, Tümer MK, Yigit S, Nursal AF,
Rustemoglu A and Balel Y: Relationship of Vitamin D and Bsml
variant with temporomandibular diseases in Turkish population. Br J
Oral Maxillofac Surg. 59:555–560. 2021.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Wu Z, Malihi Z, Stewart AW, Lawes CM and
Scragg R: The association between vitamin D concentration and pain:
A systematic review and meta-analysis. Public Health Nutr.
21:2022–2037. 2018.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Demir CY and Ersoz ME: Biochemical changes
associated with temporomandibular disorders. J Int Med Res.
47:765–771. 2019.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Lima LAR, Lopes MJP, Costa RO, Lima FAV,
Neves KRT, Calou IBF, Andrade GM and Viana GSB: Vitamin D protects
dopaminergic neurons against neuroinflammation and oxidative stress
in hemiparkinsonian rats. J Neuroinflammation.
15(249)2018.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Dias V, Junn E and Mouradian MM: The role
of oxidative stress in Parkinson's disease. J Parkinsons Dis.
3:461–491. 2013.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Jin X, Antony B, Wang X, Persson MS,
McAlindon T, Arden NK, Srivastava S, Srivastava R, Van Middelkoop
M, Bierma-Zeinstra SM, et al: Effect of vitamin D supplementation
on pain and physical function in patients with knee osteoarthritis
(OA): An OA Trial Bank protocol for a systematic review and
individual patient data (IPD) meta-analysis. BMJ Open.
10(e035302)2020.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Manoy P, Yuktanandana P, Tanavalee A,
Anomasiri W, Ngarmukos S, Tanpowpong T and Honsawek S: Vitamin D
supplementation improves quality of life and physical performance
in osteoarthritis patients. Nutrients. 9(799)2017.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Nykjaer A, Dragun D, Walther D, Vorum H,
Jacobsen C, Herz J, Melsen F, Christensen EI and Willnow TE: An
endocytic pathway essential for renal uptake and activation of the
steroid 25-(OH) vitamin D3. Cell. 96:507–515. 1999.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Quraishi SA and Camargo CA Jr: Vitamin D
in acute stress and critical illness. Curr Opin Clin Nutr Metab
Care. 15:625–634. 2012.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Lee P: Vitamin D metabolism and deficiency
in critical illness. Best Pract Res Clin Endocrinol Metab.
25:769–781. 2011.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Szewczyk B, Poleszak E, Sowa-Kucma M,
Siwek M, Dudek D, Ryszewska-Pokraśniewicz B, Radziwoń-Zaleska M,
Opoka W, Czekaj J, Pilc A and Nowak G: Antidepressant activity of
zinc and magnesium in view of the current hypotheses of
antidepressant action. Pharmacol Rep. 60:588–589. 2008.PubMed/NCBI
|
|
114
|
Vink R and Nechifor M (eds): Magnesium in
the Central Nervous System. University of Adelaide Press, Adelaide,
AU, 2011.
|
|
115
|
Wolf FI, Maier JA, Nasulewicz A,
Feillet-Coudray C, Simonacci M, Mazur A and Cittadini A: Magnesium
and neoplasia: From carcinogenesis to tumor growth and progression
or treatment. Arch Biochem Biophys. 458:24–32. 2007.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Eby GA and Eby KL: Rapid recovery from
major depression using magnesium treatment. Med Hypotheses.
67:362–370. 2006.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Seelig MS: Consequences of magnesium
deficiency on the enhancement of stress reactions; preventive and
therapeutic implications (a review). J Am Coll Nutr. 13:429–446.
1994.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Sivoňová M, Žitňanová I, Hlinčíková L,
Škodáček I, Trebatická J and Ďuračková Z: Oxidative stress in
university students during examinations. Stress. 7:183–188.
2004.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Cratty MS and Birkle DL:
N-methyl-D-aspartate (NMDA)-mediated corticotropin-releasing factor
(CRF) release in cultured rat amygdala neurons. Peptides.
20:93–100. 1999.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Perrin MH, Haas Y, Rivier JE and Vale WW:
Corticotropin-releasing factor binding to the anterior pituitary
receptor is modulated by divalent cations and guanyl nucleotides.
Endocrinology. 118:1171–1179. 1986.PubMed/NCBI View Article : Google Scholar
|
|
121
|
De Souza EB: Corticotropin-releasing
factor receptors: Physiology, pharmacology, biochemistry and role
in central nervous system and immune disorders.
Psychoneuroendocrinology. 20:789–819. 1995.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Murck H: Magnesium and affective
disorders. Nutr Neurosci. 5:375–389. 2002.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Henrotte JG, Franck G, Santarromana M,
Frances H, Mouton D and Motta R: Mice selected for low and high
blood magnesium levels: A new model for stress studies. Physiol
Behav. 61:653–658. 1997.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Kramer JH, Spurney C, Iantorno M, Tziros
C, Mak IT, Tejero-Taldo MI, Chmielinska JJ, Komarov AM and Weglicki
WB: Neurogenic inflammation and cardiac dysfunction due to
hypomagnesemia. Am J Med Sci. 338:22–27. 2009.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Tejero-Taldo MI, Kramer JH, Mak IuT,
Komarov AM and Weglicki WB: The nerve-heart connection in the
pro-oxidant response to Mg-deficiency. Heart Fail Rev. 11:35–44.
2006.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Rickards H, Dursun SM, Farrar G, Betts T,
Corbett JA and Handley SL: Increased plasma kynurenine and its
relationship to neopterin and tryptophan in Tourette's syndrome.
Psychol Med. 26:857–862. 1996.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Nakamura M, Abe S, Goto Y, Chishaki A,
Akazawa K and Kato M: In vivo assessment of prevention of
white-noise-induced seizure in magnesium-deficient rats by
N-methyl-D-aspartate receptor blockers. Epilepsy Res. 17:249–256.
1994.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Planells E, Lerma A, Sanchez-Morito N,
Aranda P and LLopis J: Effect of magnesium deficiency on vitamin B2
and B6 status in the rat. J Am Coll Nutr. 16:352–356.
1997.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Fowler B: Recent advances in the mechanism
of pyridoxine-responsive disorders. J Inherit Metab Dis. 8 (Suppl
1):S76–S83. 1985.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Henkin RI, Gouliouk V and Fordyce A:
Distinguishing patients with glossopyrosis from those with
oropyrosis based upon clinical differences and differences in
saliva and erythrocyte magnesium. Arch Oral Biol. 57:205–210.
2012.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Abaamrane L, Raffin F, Gal M, Avan P and
Sendowski I: Long-term administration of magnesium after acoustic
trauma caused by gunshot noise in guinea pigs. Hear Res.
247:137–145. 2009.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Muneyyirci-Delale O, Nacharaju VL, Dalloul
M, Altura BM and Altura BT: Serum ionized magnesium and calcium in
women after menopause: Inverse relation of estrogen with ionized
magnesium. Fertil Steril. 71:869–872. 1999.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Touyz RM: Magnesium in clinical medicine.
Front Biosci. 9:1278–1293. 2004.PubMed/NCBI View
Article : Google Scholar
|
|
134
|
Durlach J, Bac P, Durlach V, Bara M and
Guiet-Bara A: Neurotic, neuromuscular and autonomic nervous form of
magnesium imbalance. Magnes Res. 10:169–195. 1997.PubMed/NCBI(In English, French).
|
|
135
|
Caddell JL: A triple-risk model for the
sudden infant death syndrome (SIDS) and the apparent
life-threatening episode (ALTE): The stressed magnesium deficient
weanling rat. Magnes Res. 14:227–238. 2001.PubMed/NCBI
|
|
136
|
Omiya K, Akashi YJ, Yoneyama K, Osada N,
Tanabe K and Miyake F: Heart-rate response to sympathetic nervous
stimulation, exercise, and magnesium concentration in various sleep
conditions. Int J Sport Nutr Exerc Metab. 19:127–135.
2009.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Sarchielli P, Coata G, Firenze C, Morucci
P, Abbritti G and Gallai V: Serum and salivary magnesium levels in
migraine and tension-type headache. Results in a group of adult
patients. Cephalalgia. 12:21–27. 1992.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Thomas J, Tomb E, Thomas E and Faure G:
Migraine treatment by oral magnesium intake and correction of the
irritation of buccofacial and cervical muscles as a side effect of
mandibular imbalance. Magnes Res. 7:123–127. 1994.PubMed/NCBI
|
|
139
|
Bagheri G, Rezaee R, Tsarouhas K, Docea
AO, Shahraki J, Shahriari M, Wilks MF, Jahantigh H, Tabrizian K,
Moghadam AA, et al: Magnesium sulfate ameliorates carbon
monoxide-induced cerebral injury in male rats. Mol Med Rep.
19:1032–1039. 2019.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Garcia-Lopez R, Perea-Milla E, Garcia CR,
Rivas-Ruiz F, Romero-Gonzalez J, Moreno JL, Faus V, Aguas Gdel C
and Diaz JC: New therapeutic approach to Tourette Syndrome in
children based on a randomized placebo-controlled double-blind
phase IV study of the effectiveness and safety of magnesium and
vitamin B6. Trials. 10(16)2009.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Cernak I, Savic V, Kotur J, Prokic V,
Kuljic B, Grbovic D and Veljovic M: Alterations in magnesium and
oxidative status during chronic emotional stress. Magnes Res.
13:29–36. 2000.PubMed/NCBI
|
|
142
|
Esch T, Stefano GB, Fricchione GL and
Benson H: The role of stress in neurodegenerative diseases and
mental disorders. Neuro Endocrinol Lett. 23:199–208.
2002.PubMed/NCBI
|
|
143
|
Esch T, Stefano GB, Fricchione GL and
Benson H: Stress-related diseases-a potential role for nitric
oxide. Med Sci Monit. 8:RA103–RA118. 2002.PubMed/NCBI
|
|
144
|
Truelsen T, Nielsen N, Boysen G and
Gronbaek M: Copenhagen City Heart Study. Self-reported stress and
risk of stroke: The Copenhagen City Heart Study. Stroke.
34:856–862. 2003.PubMed/NCBI View Article : Google Scholar
|
|
145
|
Ella B, Ghorayeb I, Burbaud P and Guehl D:
Bruxism in movement disorders: A comprehensive review. J
Prosthodont. 26:599–605. 2017.PubMed/NCBI View Article : Google Scholar
|
|
146
|
Dickoff D: Bruxism Associated with
Restless Limbs Syndrome (RLS): A Dopamine Responsive Movement
Disorder (P02.063). Neurology. 80 (Suppl 7)(P02.063)2013.
|
|
147
|
Grimaldi BL: The central role of magnesium
deficiency in Tourette's syndrome: Causal relationships between
magnesium deficiency, altered biochemical pathways and symptoms
relating to Tourette's syndrome and several reported comorbid
conditions. Med Hypotheses. 58:47–60. 2002.PubMed/NCBI View Article : Google Scholar
|
|
148
|
Rizzo G, Li X, Galantucci S, Filippi M and
Cho YW: Brain imaging and networks in restless legs syndrome. Sleep
Med. 31:39–48. 2017.PubMed/NCBI View Article : Google Scholar
|
|
149
|
Connor JR, Boyer PJ, Menzies SL, Dellinger
B, Allen RP, Ondo WG and Earley CJ: Neuropathological examination
suggests impaired brain iron acquisition in restless legs syndrome.
Neurology. 61:304–309. 2003.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Allen RP and Earley CJ: The role of iron
in restless legs syndrome. Mov Disord. 22 (Suppl 18):S440–S448.
2007.PubMed/NCBI View Article : Google Scholar
|
|
151
|
Allen RP, Auerbach S, Bahrain H, Auerbach
M and Earley CJ: The prevalence and impact of restless legs
syndrome on patients with iron deficiency anemia. Am J Hematol.
88:261–264. 2013.PubMed/NCBI View Article : Google Scholar
|
|
152
|
Umbreit J: Iron deficiency: A concise
review. Am J Hematol. 78:225–231. 2005.PubMed/NCBI View Article : Google Scholar
|
|
153
|
Camaschella C: Iron-deficiency anemia. N
Engl J Med. 372:1832–1843. 2015.PubMed/NCBI View Article : Google Scholar
|
|
154
|
Lopez A, Cacoub P, Macdougall IC and
Peyrin-Biroulet L: Iron deficiency anaemia. Lancet. 387:907–916.
2016.PubMed/NCBI View Article : Google Scholar
|
|
155
|
Allen RP, Picchietti DL, Auerbach M, Cho
YW, Connor JR, Earley CJ, Garcia-Borreguero D, Kotagal S, Manconi
M, Ondo W, et al: Evidence-based and consensus clinical practice
guidelines for the iron treatment of restless legs
syndrome/Willis-Ekbom disease in adults and children: An IRLSSG
task force report. Sleep Med. 41:27–44. 2018.PubMed/NCBI View Article : Google Scholar
|
|
156
|
Trotti LM and Becker LA: Iron for the
treatment of restless legs syndrome. Cochrane Database Syst Rev.
1(CD007834)2019.PubMed/NCBI View Article : Google Scholar
|
|
157
|
Wei C, Zhou J, Huang X and Li M: Effects
of psychological stress on serum iron and erythropoiesis. Int J
Hematol. 88:52–56. 2008.PubMed/NCBI View Article : Google Scholar
|
|
158
|
Singh A, Smoak BL, Patterson KY, LeMay LG,
Veillon C and Deuster PA: Biochemical indices of selected trace
minerals in men: Effect of stress. Am J Clin Nutr. 53:126–131.
1991.PubMed/NCBI View Article : Google Scholar
|
|
159
|
Ferraz AC, Delattre AM, Almendra RG,
Sonagli M, Borges C, Araujo P, Andersen ML, Tufik S and Lima MM:
Chronic ω-3 fatty acids supplementation promotes beneficial effects
on anxiety, cognitive and depressive-like behaviors in rats
subjected to a restraint stress protocol. Behav Brain Res.
219:116–122. 2011.PubMed/NCBI View Article : Google Scholar
|
|
160
|
Laye S, Nadjar A, Joffre C and Bazinet RP:
Anti-Inflammatory effects of omega-3 fatty acids in the brain:
Physiological mechanisms and relevance to pharmacology. Pharmacol
Rev. 70:12–38. 2018.PubMed/NCBI View Article : Google Scholar
|
|
161
|
Larrieu T, Hilal ML, Fourrier C, De
Smedt-Peyrusse V, Sans N, Capuron L and Layé S: Nutritional omega-3
modulates neuronal morphology in the prefrontal cortex along with
depression-related behaviour through corticosterone secretion.
Transl Psychiatry. 4(e437)2014.PubMed/NCBI View Article : Google Scholar
|
|
162
|
Cao D, Kevala K, Kim J, Moon HS, Jun SB,
Lovinger D and Kim HY: Docosahexaenoic acid promotes hippocampal
neuronal development and synaptic function. J Neurochem.
111:510–521. 2009.PubMed/NCBI View Article : Google Scholar
|
|
163
|
Joffre C, Gregoire S, De Smedt V, Acar N,
Bretillon L, Nadjar A and Layé S: Modulation of brain PUFA content
in different experimental models of mice. Prostaglandins Leukot
Essent Fatty Acids. 114:1–10. 2016.PubMed/NCBI View Article : Google Scholar
|
|
164
|
Mackie K: Signaling via CNS cannabinoid
receptors. Mol Cell Endocrinol. 286 (1-2 Suppl 1):S60–S65.
2008.PubMed/NCBI View Article : Google Scholar
|
|
165
|
Freund TF, Katona I and Piomelli D: Role
of endogenous cannabinoids in synaptic signaling. Physiol Rev.
83:1017–1066. 2003.PubMed/NCBI View Article : Google Scholar
|
|
166
|
Larrieu T and Laye S: Food for mood:
Relevance of nutritional omega-3 fatty acids for depression and
anxiety. Front Physiol. 9(1047)2018.PubMed/NCBI View Article : Google Scholar
|
|
167
|
Auguste S, Fisette A, Fernandes MF,
Hryhorczuk C, Poitout V, Alquier T and Fulton S: Central Agonism of
GPR120 acutely inhibits food intake and food reward and chronically
suppresses anxiety-like behavior in mice. Int J
Neuropsychopharmacol. 19(pyw014)2016.PubMed/NCBI View Article : Google Scholar
|
|
168
|
Kidd PM: Omega-3 DHA and EPA for
cognition, behavior, and mood: Clinical findings and
structural-functional synergies with cell membrane phospholipids.
Altern Med Rev. 12:207–227. 2007.PubMed/NCBI
|
|
169
|
Simopoulos AP: Evolutionary aspects of
diet: The omega-6/omega-3 ratio and the brain. Mol Neurobiol.
44:203–215. 2011.PubMed/NCBI View Article : Google Scholar
|
|
170
|
Carrie I, Clement M, de Javel D, Frances H
and Bourre JM: Phospholipid supplementation reverses behavioral and
biochemical alterations induced by n-3 polyunsaturated fatty acid
deficiency in mice. J Lipid Res. 41:473–480. 2000.PubMed/NCBI
|
|
171
|
Bondi CO, Taha AY, Tock JL, Totah NK,
Cheon Y, Torres GE, Rapoport SI and Moghaddam B: Adolescent
behavior and dopamine availability are uniquely sensitive to
dietary omega-3 fatty acid deficiency. Biol Psychiatry. 75:38–46.
2014.PubMed/NCBI View Article : Google Scholar
|
|
172
|
Suri D and Vaidya VA: Glucocorticoid
regulation of brain-derived neurotrophic factor: Relevance to
hippocampal structural and functional plasticity. Neuroscience.
239:196–213. 2013.PubMed/NCBI View Article : Google Scholar
|
|
173
|
Tay TL, Savage JC, Hui CW, Bisht K and
Tremblay ME: Microglia across the lifespan: From origin to function
in brain development, plasticity and cognition. J Physiol.
595:1929–1945. 2017.PubMed/NCBI View Article : Google Scholar
|
|
174
|
Moon DO, Kim KC, Jin CY, Han MH, Park C,
Lee KJ, Park YM, Choi YH and Kim GY: Inhibitory effects of
eicosapentaenoic acid on lipopolysaccharide-induced activation in
BV2 microglia. Int Immunopharmacol. 7:222–229. 2007.PubMed/NCBI View Article : Google Scholar
|
|
175
|
De Smedt-Peyrusse V, Sargueil F, Moranis
A, Harizi H, Mongrand S and Laye S: Docosahexaenoic acid prevents
lipopolysaccharide-induced cytokine production in microglial cells
by inhibiting lipopolysaccharide receptor presentation but not its
membrane subdomain localization. J Neurochem. 105:296–307.
2008.PubMed/NCBI View Article : Google Scholar
|
|
176
|
Levine B, Mizushima N and Virgin HW:
Autophagy in immunity and inflammation. Nature. 469:323–335.
2011.PubMed/NCBI View Article : Google Scholar
|
|
177
|
Orr SK, Palumbo S, Bosetti F, Mount HT,
Kang JX, Greenwood CE, Ma DW, Serhan CN and Bazinet RP:
Unesterified docosahexaenoic acid is protective in
neuroinflammation. J Neurochem. 127:378–393. 2013.PubMed/NCBI View Article : Google Scholar
|
|
178
|
Lynch AM, Loane DJ, Minogue AM, Clarke RM,
Kilroy D, Nally RE, Roche OJ, O'Connell F and Lynch MA:
Eicosapentaenoic acid confers neuroprotection in the amyloid-beta
challenged aged hippocampus. Neurobiol Aging. 28:845–855.
2007.PubMed/NCBI View Article : Google Scholar
|
|
179
|
Huang CT and Tsai YJ: Docosahexaenoic acid
confers analgesic effects after median nerve injury via inhibition
of c-Jun N-terminal kinase activation in microglia. J Nutr Biochem.
29:97–106. 2016.PubMed/NCBI View Article : Google Scholar
|
|
180
|
Mirza M, Volz C, Karlstetter M, Langiu M,
Somogyi A, Ruonala MO, Tamm ER, Jägle H and Langmann T: Progressive
retinal degeneration and glial activation in the CLN6 (nclf) mouse
model of neuronal ceroid lipofuscinosis: A beneficial effect of DHA
and curcumin supplementation. PLoS One. 8(e75963)2013.PubMed/NCBI View Article : Google Scholar
|
|
181
|
Kiecolt-Glaser JK, Belury MA, Andridge R,
Malarkey WB and Glaser R: Omega-3 supplementation lowers
inflammation and anxiety in medical students: A randomized
controlled trial. Brain Behav Immun. 25:1725–1734. 2011.PubMed/NCBI View Article : Google Scholar
|
|
182
|
Headache Classification Committee of the
International Headache Society (IHS). The International
Classification of Headache Disorders, 3rd edition (beta version).
Cephalalgia. 33:629–808. 2013.PubMed/NCBI View Article : Google Scholar
|
|
183
|
Ashina M: Migraine. N Engl J Med.
383:1866–1876. 2020.PubMed/NCBI View Article : Google Scholar
|
|
184
|
Marchetti M, Gualtieri P, De Lorenzo A,
Trombetta D, Smeriglio A, Ingegneri M, Cianci R, Frank G, Schifano
G, Bigioni G and Di Renzo L: Dietary ω-3 intake for the treatment
of morning headache: A randomized controlled trial. Front Neurol.
13(987958)2022.PubMed/NCBI View Article : Google Scholar
|
|
185
|
Ramsden CE, Zamora D, Faurot KR, MacIntosh
B, Horowitz M, Keyes GS, Yuan ZX, Miller V, Lynch C, Honvoh G, et
al: Dietary alteration of n-3 and n-6 fatty acids for headache
reduction in adults with migraine: Randomized controlled trial.
BMJ. 374(n1448)2021.PubMed/NCBI View Article : Google Scholar
|
|
186
|
Ramsden CE, Faurot KR, Zamora D,
Suchindran CM, MacIntosh BA, Gaylord S, Ringel A, Hibbeln JR,
Feldstein AE, Mori TA, et al: Targeted alteration of dietary n-3
and n-6 fatty acids for the treatment of chronic headaches: A
randomized trial. Pain. 154:2441–2451. 2013.PubMed/NCBI View Article : Google Scholar
|