Introduction
Pneumonia is a common inflammation-related disease
with high morbidity and mortality rates among infectious diseases
and is an important health problem worldwide. Persistent
inflammatory infection with pneumonia can induce lung tissue damage
(1,2). The drugs commonly administered in the
clinical treatment of pneumonia include antibiotics and
adrenocorticoid hormones (3).
However, after treatment with these drugs, the body will develop
resistance and significant side effects including gastrointestinal
infections and metabolic disturbance (4,5).
Therefore, it has become an urgent problem for researchers to
improve the treatment efficacy of pneumonia and to understand the
underlying mechanism of the disease.
Under pathological conditions, the environment in
the endoplasmic reticulum (ER) changes and misfolded and unfolded
proteins accumulate in the ER cavity until the ER homeostasis is
disrupted. Following this, a series of stress reactions in the ER
are triggered to process these misfolded or unfolded proteins and
maintain the normal function of cells. This process is called
endoplasmic reticulum stress (ERS) (6). Hypoxia, inflammatory response,
oxidative stress and other interfering factors can lead to the
increase of protein misfolding rate and protein load in the ER,
inducing ERS (7). A previous study
showed that alleviating inflammation and ERS in pediatric pneumonia
can significantly inhibit the development of pediatric pneumonia
(8). Therefore, it is beneficial
to improve the symptoms of pneumonia by alleviating ERS and
inflammation. Previous research has shown that Stromal interaction
molecule 1 (STIM1)-Orai1 interaction exacerbates
LPS-induced inflammation and ERS in bovine hepatocytes through
store-operated calcium entry (9).
STIM1 is a recently discovered vasoactive protein that can
participate in the regulation of inflammatory processes (10-13).
A previous study has shown that STIM1 is closely related to
activation of the inflammatory response and platelet aggregation
after stent implantation in percutaneous coronary intervention
(14). In addition, overexpression
of SOX9 can alleviate the inflammatory damage of bronchial
epithelial cells induced by cigarette smoke extract by inhibiting
STIM1 (15). Furthermore,
silencing STIM1 expression inactivates NLR pyrin domain
containing 3 by promoting the expression of microRNA-223, thereby
reducing the inflammatory damage of lung epithelial cells induced
by influenza A virus (16).
STIM1 has also been shown to reduce lipopolysaccharide
(LPS)-induced inflammation by inhibiting NF-κB signaling in bovine
mammary epithelial cells (17).
However, there are few studies investigating the regulatory role of
STIM1 on ERS and inflammation in pneumonia.
The present study predicted the potential
interaction of the RNA-binding protein insulin-like growth factor 2
mRNA binding protein 2 (IGF2BP2) with STIM1 via RBPmap
website. IGF2BP2 belongs to a highly conserved family of
RNA-binding proteins whose function is to regulate mRNA
localization, stability and translation and fine-tune the
physiological function of the encoded protein (18). IGF2BP2 knockdown inhibited
LPS-induced inflammation of lung epithelial cells by targeting
caspase 4, thus inhibiting the non-standard scorching pathway
(19). So it was hypothesized that
IGF2BP2 can regulate STIM1 and thus play a role in
LPS-induced pneumonia.
In the present study, the role of STIM1 in an
LPS-induced pneumonia cell model and its regulatory mechanism were
investigated, to provide a useful theoretical basis for the future
clinical treatment of pneumonia.
Materials and methods
Database
The RNA-binding protein IGF2BP2 has a potential
interaction with STIM1, which was investigated using the
web-tool RBPmap (http://rbpmap.technion.ac.il/) that enables prediction
of RBP binding on genome sequences from a huge list of
experimentally validated motifs of RBPmap database (Table SI) (20).
Cell culture
The human lung adenocarcinoma epithelial cells A549
purchased from the BeNa Culture Collection (cat. no. BNCC337696)
and was cultured in Dulbecco's Modified Eagle's Medium (DMEM;
Thermo Fisher Scientific, Inc.) containing 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) in a 37˚C incubator with 5%
carbon dioxide. For the injury model, the cells were placed in
6-well plates (3x105 cells/well) and incubated for 12 h,
before incubation with LPS (10 µg/ml) for a further 24 h (21). LPS induces the human pulmonary
epithelial cell line A549 to create a pneumonia model (22,23).
Reverse transcription quantitative PCR
(RT-qPCR)
The A549 cells in a 6-well plate at a density of
2x105 cells/well were lysed using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) and total RNA was purified
using a RNeasy Plus Mini Kit (Qiagen, Inc.). Then, cDNA was
generated using 1 µg total RNA and a High-Capacity cDNA Reverse
Transcription Kit (Applied Biosystems; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. The cDNA was then
amplified by IQTM SYBR Green Master Mix (Bio-Rad Laboratories,
Inc.), which was then quantified using a SYBR-Green detection
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
following thermocycling conditions were used for the qPCR: Initial
denaturation at 95˚C for 10 min; 50 cycles of 95˚C for 15 sec and
60˚C for 60 sec. Each reaction was performed three times. Finally,
the 2-ΔΔCq quantification method was used to calculate
the expression levels of target mRNAs (24). The GAPDH mRNA level was used
as the normalized standard. The PCR primers were as follows:
STIM1 forward: 5'-GCCTAGGAGGCCCAGGAT-3', reverse:
5'-ACAGCCAAAGGTCAAGTGCT-3'; IGF2BP2 forward:
5'-GGAACAAGTCAACACAGACACA-3', reverse: 5'-CGCAGCGGGAAATCAATCTG-3';
GAPDH forward: 5'-AATGGGCAGCCGTTAGGAAA-3', reverse:
5'-GCGCCCAATACGACCAAATC-3'.
Western blotting
The A549 cells were lysed in RIPA buffer and protein
contents were determined by the BCA method. In total, 20 µg of
proteins was separated by 12% SDS-PAGE and then transferred to PVDF
membranes. The membranes blocked with 5% BSA (Sigma-Aldrich; Merck
KGaA) for 1 h at room temperature were incubated with primary
antibodies STIM1 (1:1,000; cat. no. ab108994; Abcam), Bcl-2
(1:1,000; cat. no. ab182858; Abcam), Bax (1:1,000; cat. no.
ab32503; Abcam), glucose-regulated protein 78 (GRP78; 1:1,000; cat.
no. ab21685; Abcam), activating transcription factor 6 (ATF6;
1:1,000; cat. no. ab227830; Abcam), C/EBP homologous protein (CHOP)
(1:1,000; cat. no. 5554; CST), caspase 12 (1:1,000; cat. no.
ab62484; Abcam), PKR-like ER kinase (PERK; 1:1,000; cat. no.
ab229912; Abcam), insulin-like growth factor 2 mRNA binding protein
3 (IGF2BP3; 1:1,000; cat. no. ab177477; Abcam), phosphorylated
(p-)PERK (1:1,000; cat. no. 3179; CST), GAPDH (1:1,000; cat. no.
ab9485; Abcam) overnight at 4˚C. Then, the appropriate horseradish
peroxidase-conjugated secondary antibody (1:5,000; cat. no.
ab150077; Abcam) was incubated with the membranes for 1 h at 37˚C.
GAPDH was used as the internal control. Protein bands were then
detected by enhanced chemiluminescence (MilliporeSigma). The
western blot images were analyzed using Image J software (V1.8.0,
National Institutes of Health).
Cell transfection
Insulin-like growth factor 2 mRNA binding protein 2
(IGF2BP2) overexpression lentivirus (Oe-IGF2BP2) and
the corresponding negative control (Oe-NC; pcDNA3.1) and small
interfering (si)RNAs against STIM1 (si-STIM1#1 and
si-STIM1#2) and their corresponding scrambled sequence
negative control (si-NC) were obtained from Shanghai GenePharma
Co., Ltd. Transfection was conducted at 37˚C for 48 h using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
transfection efficiency was detected by RT-qPCR or western blotting
as aforementioned 48 h after transfection. The sequences were as
follows: si-STIM1#1 sense, 5'-UCAAUUCGGCAAAACUCUGCU-3' and
antisense, 5'-CAGAGUUUUGCCGAAUUGACA-3'; si-STIM1#2 sense,
5'-UCAGUUUGUGGAUGUUACGGA-3' and antisense,
5'-CGUAACAUCCACAAACUGAUG-3'; si-IGF2BP2#1 sense,
5'-AGUAGUUCUCAAACUGAUGCC-3' and antisense,
5'-CAUCAGUUUGAGAACUACUCC-3'; si-IGF2BP2#2 sense,
5'-UCUUGAAGGAGUAGUUCUCAA-3' and antisense,
5'-GAGAACUACUCCUUCAAGAUU-3'; and si-NC sense,
5'-UUCUCCGAACGUGUCACGUTT-3' and antisense,
5'-ACGUGACACGUUCGGAGAATT-3'.
Cell Counting Kit (CCK)-8 assay
A549 cells were seeded into 96-well plates at a
density of 5x103 cells/well and were treated accordingly
before incubation with 10 µl CCK-8 working solution
(MedChemExpress) for 4 h. Absorbance was measured at 450 nm using a
microplate reader.
Flow cytometry
A549 cells were seeded into 96-well plates at a
density of 2x106 cells/well. After the corresponding
incubation, cells were stained with FITC-conjugated annexin V and
propidium iodide using an annexin V-FITC apoptosis kit (Beyotime
Institute of Biotechnology) for 10 min in the dark at room
temperature. Apoptotic cells were quantified by loading the cell
mixture onto a CytoFLEX flow cytometry system (Beckman Coulter,
Inc.) and FlowJo software (Version 10; FlowJo LLC) was used to
analyze the data. Apoptosis rate was calculated as the sum of the
early apoptosis rate (the lower right quadrant) and the late
apoptosis rate (the upper right quadrant).
Enzyme-linked immunosorbent assay
(ELISA)
To detect the levels of IL-6 (cat. no. ab178013;
Abcam), IL-1β (cat. no. ab214025; Abcam) and TNF-α (cat. no.
ab181421; Abcam) in the cell supernatants, related ELISA assay kits
were used according to the manufacturer's recommendations.
Immunofluorescence (IF)
After treatment, A549 cells were fixed in 4%
paraformaldehyde (300 µl/well; Sigma-Aldrich; Merck KGaA),
incubated in 0.3% Triton X-100 (500 µl/well; Sigma-Aldrich; Merck
KGaA) for 15 min and then blocked with 5% goat serum
(Sigma-Aldrich; Merck KGaA). The slides were then stained with CHOP
primary antibody (1:300; cat. no. 5554; CST) overnight at 4˚C and
the fluorescent secondary antibody (1:500; cat. no. ab150077;
Abcam) at 37˚C for 30 min, before being stained with DAPI
(MilliporeSigma) for 10 min at room temperature in mounting medium.
The cells were observed using an LSM 710 confocal laser microscope
system (Carl Zeiss AG).
RNA-binding protein
immunoprecipitation (RIP) assay
RIP assays were performed according to the EZ-Magna
RIP RNA-binding Protein Immunoprecipitation Kit (MilliporeSigma).
The cells were collected and added with 10 ml PBS and 0.01%
formaldehyde to crosslink for 15 min. Then 1.4 ml 2 mol/l glycine
was added and mixed for 5 min before being centrifuged for 5 min at
1,000 x g at room temperature. After discarding the supernatant,
the cells were lysed with RIPA lysis buffer with 20 µl of protein A
resin addition and incubated for 1 h at 4˚C. The supernatant was
discarded and 50 µl PBS was added for suspension. Total RNA was
extracted from suspension with TRIzol® reagent and
reverse synthesis of cDNA. Finally, STIM1 mRNA abundance was
detected by RT-qPCR as aforementioned.
RNA stability analysis
After treatment, A549 cells were exposed to 2 µg/ml
actinomycin D (Cayman Chemical Company) for 0, 4, 8, 12 or 24 h to
block transcription. DMSO (MilliporeSigma) was used as the control
reagent. Cell samples were collected at the indicated time points
and RNA was extracted. Finally, STIM1 mRNA expression levels
were detected by RT-qPCR as aforementioned (25).
Statistical analysis
The data are presented as the mean ± SD and were
analyzed using GraphPad Prism 5 (Dotmatics). The comparisons were
assessed using one-way ANOVA followed by Tukey's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference. Each experiment was repeated at least three times.
Results
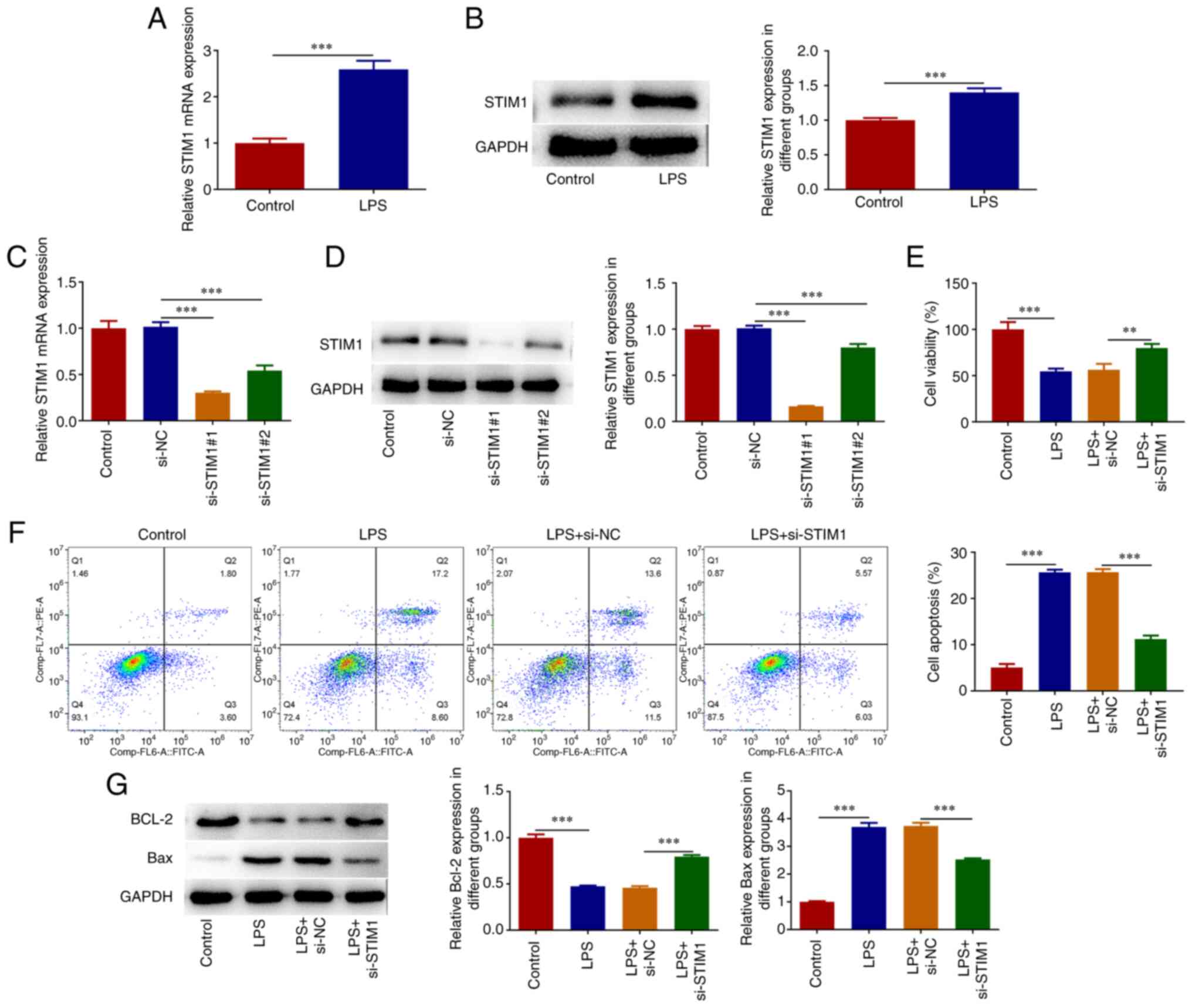
STIM1 is upregulated in LPS-induced
A549 cells and STIM1 knockdown inhibits LPS-induced A549 cell
apoptosis
After A549 cells were induced with LPS, the
expression of STIM1 was detected by RT-qPCR and western
blotting. The results demonstrated that the expression of
STIM1 was significantly increased after LPS induction,
compared with the untreated control cells (Fig. 1A and B). Following this, STIM1
interference plasmids were constructed, A549 cells were transfected
with these plasmids and the transfection efficacy was detected by
RT-qPCR and western blotting (Fig.
1C and D). si-STIM1#1
was selected for follow-up experiments due to its more prominent
interference efficacy. Cells were then divided into the control,
LPS, LPS + si-NC and LPS + si-STIM1 groups. Subsequently,
cell viability was measured by CCK-8 and the results demonstrated
that cell viability was significantly decreased in the LPS group
compared with the control group. Furthermore, compared with the LPS
+ si-NC group, cell viability was increased in the LPS +
si-STIM1 group (Fig. 1E).
Flow cytometry demonstrated that LPS treatment induced cell
apoptosis, which was significantly reduced after inhibition of
STIM1 expression (Fig. 1F).
Western blotting analysis of apoptosis-related proteins
demonstrated that Bcl-2 expression was decreased and Bax expression
was increased in the LPS group, compared with the control group.
Furthermore, compared with the LPS + si-NC group, Bcl-2 expression
was increased and Bax expression was decreased in the LPS +
si-STIM1 group (Fig.
1G).
STIM1 knockdown alleviates LPS-induced
inflammation and ERS in A549 cells
ELISA kits were used to detect the levels of
inflammatory cytokines and the results indicated that LPS could
significantly increase the levels of IL-6, IL-1β and TNF-α.
Following inhibition of STIM1 expression, the levels of the
inflammatory cytokines were reversed (Fig. 2A). An IF assay was used to detect
the expression of CHOP and it was found that the fluorescence
intensity of CHOP was significantly increased after LPS induction,
while expression was significantly decreased after STIM1
inhibition (Fig. 2B). Western
blotting analysis demonstrated that GRP78, ATF6, CHOP, caspase 12
and p-PERK expression levels were increased in the LPS group
compared with the control group. Furthermore, compared with the LPS
+ si-NC group, the expression of the aforementioned proteins in the
LPS + si-STIM1 group were decreased (Fig. 2C).
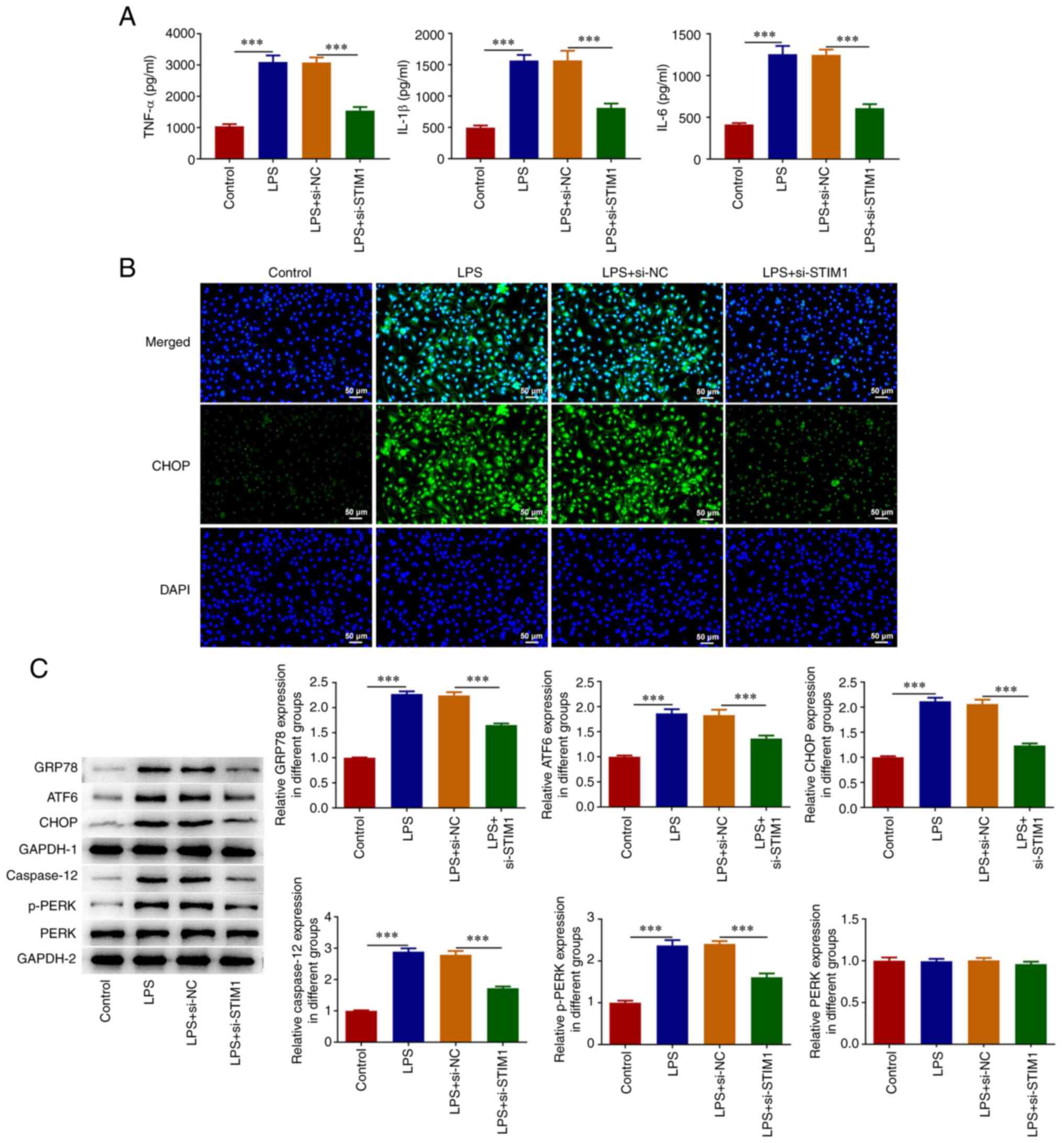 | Figure 2STIM1 knockdown alleviates
LPS-induced A549 cell inflammation and ER stress. (A) Enzyme-linked
immunosorbent assay kits were used to detect the levels of
inflammatory cytokines. (B) Immunofluorescence assay was used to
detect the expression of CHOP. (C) Western blotting was used to
detect the expression of GRP78, ATF6, CHOP, caspase 12 and p-PERK.
***P<0.001. STIM1, stromal interaction
molecule 1; LPS, lipopolysaccharide; ATF6, activating transcription
factor 6; ER, endoplasmic reticulum; CHOP, C/EBP homologous
protein; GRP78, glucose-regulated protein 78; NC, negative control;
p-PERK, phosphorylated PKR-like ER kinase; si, small interfering
RNA. |
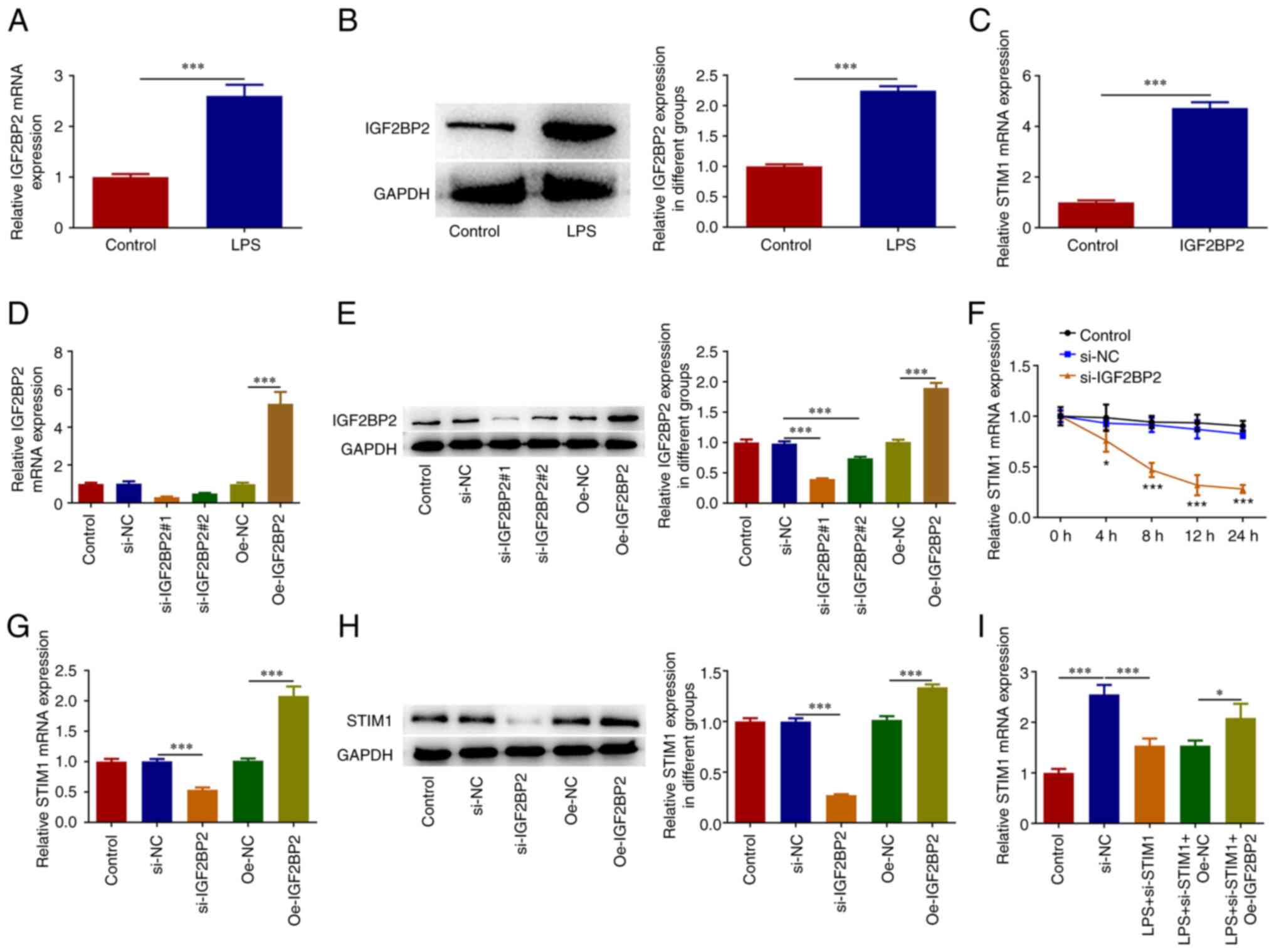
IGF2BP2 enhances the stability of
STIM1 mRNA
The results of RT-qPCR and western blotting assays
demonstrated that IGF2BP2 levels were also significantly
increased in LPS-induced A549 cells (Fig. 3A and B). The binding ability of IGF2BP2 to
STIM1 mRNA was detected using a RIP assay; as expected,
STIM1 mRNA was enriched in RNA pulled down by anti-IGF2BP2
in cells (Fig. 3C). IGF2BP2
interference and overexpression plasmids were constructed and
transfected into A549 cells and the transfection efficacy was
detected by RT-qPCR and western blotting (Fig. 3D and E). Following this, si-IGF2BP2#1
was chosen for the follow-up experiments. After actinomycin D
treatment, the stability of STIM1 mRNA decreased upon
IGF2BP2 inhibition (Fig.
3F). Next, cells were divided into the control, si-NC,
si-IGF2BP2, Oe-NC and Oe-IGF2BP2 groups and the
STIM1 expression level in these groups was detected by
RT-qPCR and western blotting. The results demonstrated that the
expression of STIM1 decreased significantly after
IGF2BP2 expression was inhibited. Furthermore,
overexpression of IGF2BP2 significantly increased the
expression of STIM1 in cells (Fig. 3G and H). Next, cells were divided into the
control, LPS, LPS + si-STIM1, LPS + si-STIM1 + Oe-NC
and LPS + si-STIM1 + Oe-IGF2BP2 groups. RT-qPCR
results showed that STIM1 expression was significantly
inhibited in LPS + si-STIM1 group compared with LPS group.
Compared with LPS + si-STIM1 + Oe-NC group, the expression
of STIM1 was increased in LPS + si-STIM1 +
Oe-IGF2BP2 group (Fig.
3I).
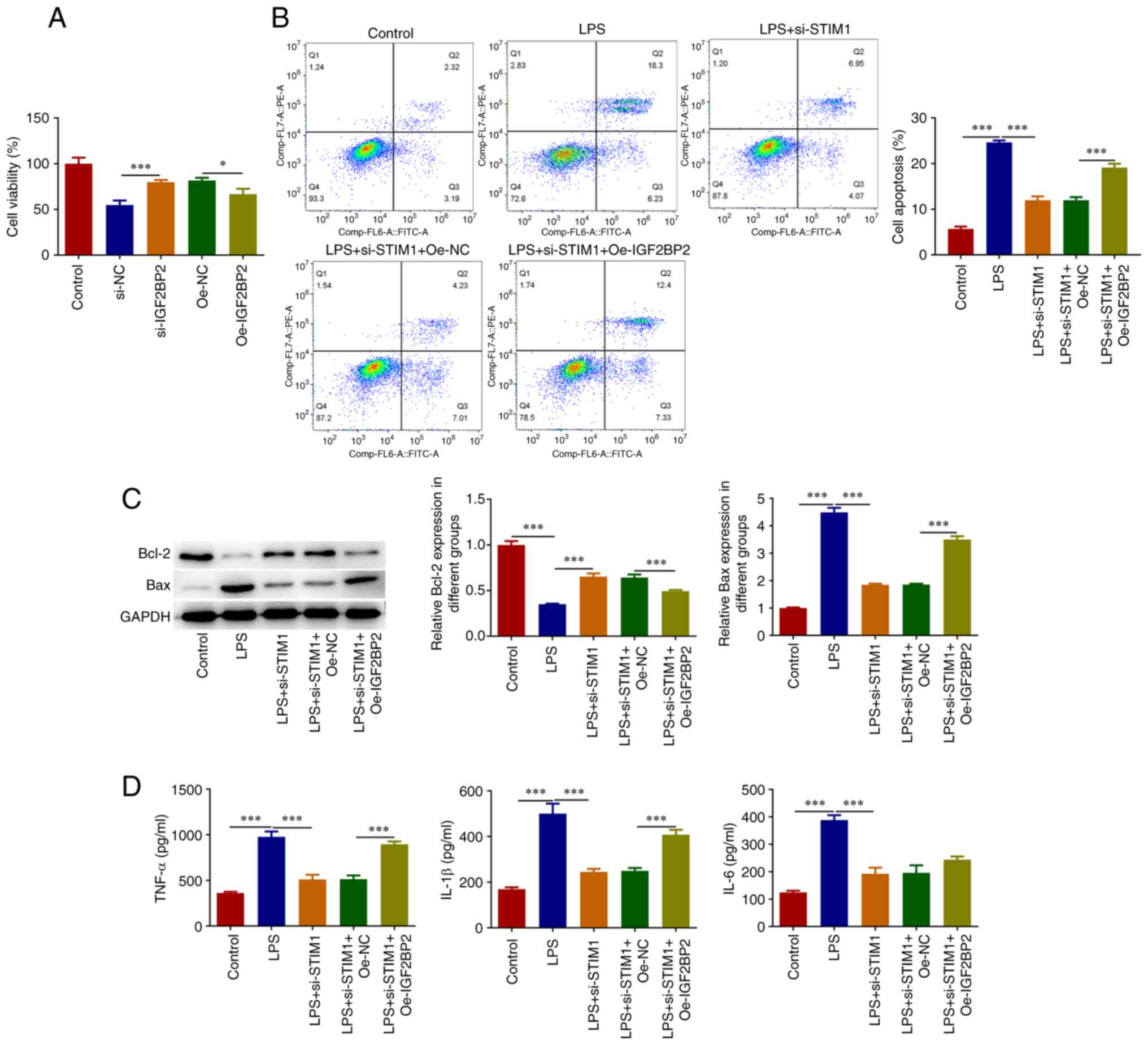
Knockdown of IGF2BP2-regulated STIM1
expression alleviates LPS-induced ERS and inflammatory responses in
A549 cells
The results demonstrated that overexpression of
IGF2BP2 reversed the inhibitory effect of STIM1
knockdown on the LPS-induced apoptosis of A549 cells (Fig. 4A-C). The ELISA results demonstrated
that, compared with the LPS + si-STIM1 + Oe-NC group, the
levels of IL-1β and TNF-α in the LPS + si-STIM1 +
Oe-IGF2BP2 group were significantly increased, IL-6 showed
an increasing trend, but it was not marked (Fig. 4D). Furthermore, the results of the
IF assay showed that overexpression of IGF2BP2 reversed the
inhibitory effect of STIM1 knockdown on CHOP expression in
LPS-induced A549 cells (Fig. 5A).
In addition, western blotting analysis demonstrated that, compared
with the LPS + si-STIM1 + Oe-NC group, the levels of GRP78,
ATF6, CHOP, caspase 12 and p-PERK in the LPS + si-STIM1 +
Oe-NC group were significantly increased (Fig. 5B).
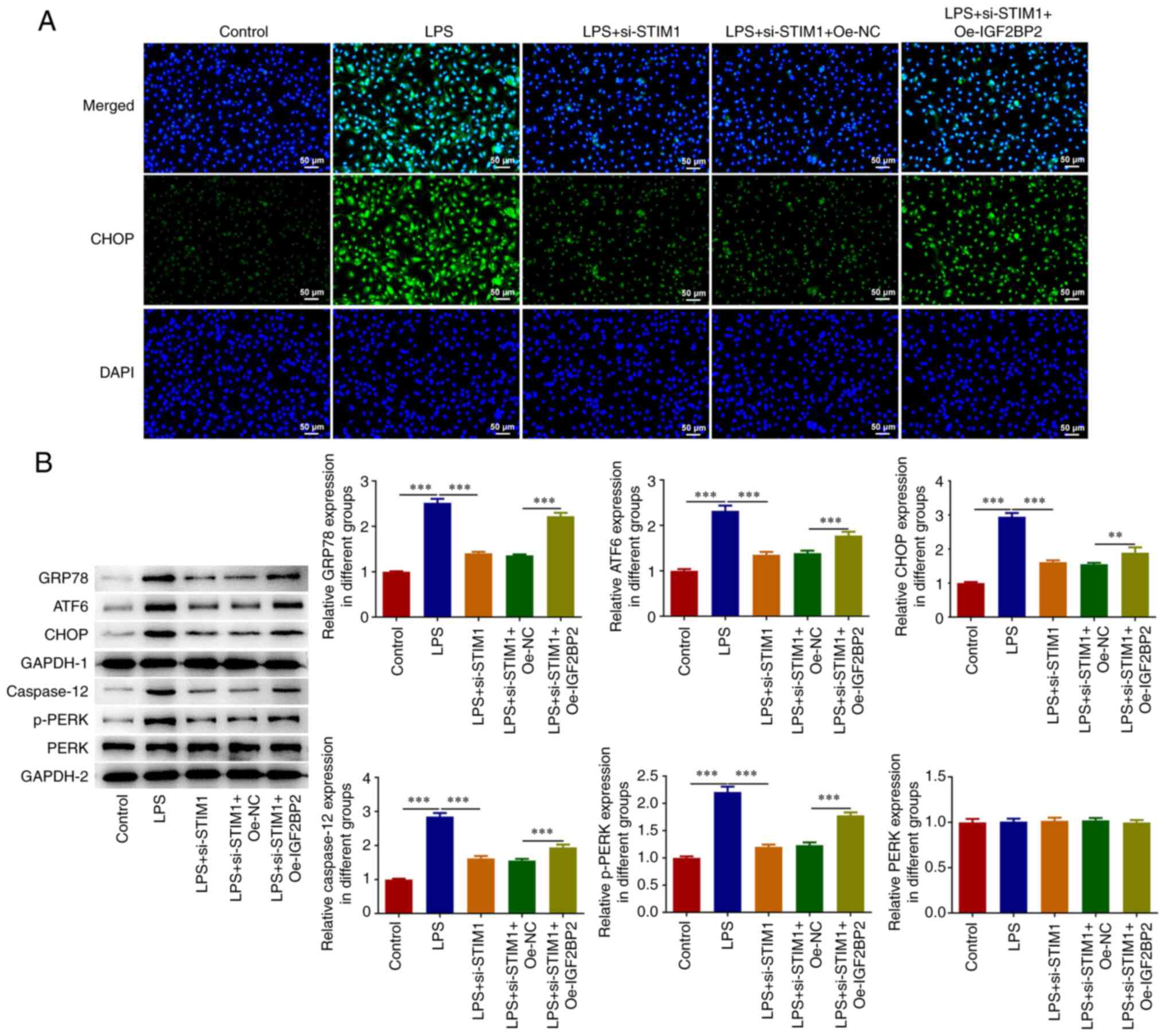 | Figure 5Knockdown of IGF2BP2-regulated
STIM1 expression alleviates LPS-induced ER stress in A549
cells. (A) An immunofluorescence assay was used to detect the
expression of CHOP. (B) Western blotting was used to detect the
expression of GRP78, ATF6, CHOP, caspase 12 and p-PERK.
**P<0.01, ***P<0.001. IGF2BP2,
insulin-like growth factor 2 mRNA binding protein 2; STIM1,
stromal interaction molecule 1; LPS, lipopolysaccharide; ER,
endoplasmic reticulum; CHOP, C/EBP homologous protein; GRP78,
glucose-regulated protein 78; ATF6, activating transcription factor
6; NC, negative control; Oe, overexpression; p-PERK, phosphorylated
PKR-like ER kinase; si, small interfering (RNA). |
Discussion
Pneumonia is a severe infectious disease attributed
to a number of pathogenic factors, among which bacterial pneumonia
is the most common. LPS, a major component of the gram-negative
bacterial cell wall, is a common inducer of the inflammatory
response in tissues and cells (26,27).
After gram-negative bacteria enters the body, LPS can induce
chemotaxis, migration and eventually infiltration of the lung
tissues and, accompanied by the upregulation of inflammatory
mediators, ultimately lead to the occurrence of acute lung injury
(28). Hence, LPS is often used as
an inducer for acute lung injury models. In the present study, LPS
was used to construct a lung injury model using A549 cells. The
results demonstrated that LPS led to significantly decreased cell
viability and increased apoptosis and inflammatory response, which
indicated the successful construction of the model.
Classical ERS receptors are ER membrane proteins,
PERK and ATF6. Typically, when these receptor proteins are bound to
the GRP78 molecular chaperone, they are inactive and become
activated upon dissociation from GRP78 during ERS, triggering
downstream events through signal transduction (29). In addition, upregulation of CHOP
transcription factor may induce the inflammatory response and
apoptosis and promote the occurrence and development of diseases
(30,31). Inhibition of ERS-mediated apoptosis
has been reported to reduce susceptibility to Streptococcus
pneumoniae co-infection after influenza infection (32). Furthermore, ginsenoside Rg1
regulates sirtuin 1 and improves lung inflammation and injury
caused by sepsis by inhibiting ERS and inflammation (33). In addition, caspase12 is a
pro-apoptotic molecule specific to the outer membrane of the ER and
activation of caspase12 is one of the core links in the development
of ERS phase decay (34). In the
present study, LPS was administered to A549 cells to induce ERS
injury. The expression of ERS-related proteins, GRP78, ATF6, CHOP,
caspase 12 and p-PERK, were significantly increased after LPS
induction. These results indicated that LPS induced ERS injury in
A549 cells.
In the present study, it was found that STIM1
expression was significantly elevated in LPS-induced A549 cells.
STIM1 is a newly discovered human gene located in a specific
region of human chromosome 11p15.5(35). STIM1 is involved in the
regulation of the inflammatory processes. Regulation of
STIM1/Orai1 signaling in bovine mammary epithelial
cells has been shown to alleviate LPS-induced inflammation
(17). In addition, the expression
of STIM1 and Orai1 is upregulated when exposed to a high
water volume or large cyclic stretching, which further activates
calcium-sensitive protein kinase Cα, leading to calcium overload,
excessive endothelial permeability and ultimately
ventilator-induced lung injury (36). STIM1 activation also
mediates S-phase stagnation and cell death in acute pulmonary
poisoning induced by paraquat (37). In addition, STIM1-Orai1
interaction exacerbates LPS-induced bovine hepatocyte inflammation
and ERS through store-operated calcium entry (9). However, to the best of the authors'
knowledge, the role of STIM1 in LPS-induced A549 cell injury
has not yet been reported. In the present study, it was found that
inhibition of STIM1 expression in LPS-induced A549 cells
significantly inhibited LPS-induced apoptosis, inflammation and
ERS.
In the present study, through RIP and other
experiments, it was demonstrated that IGF2BP2 could enhance the
stability of STIM1 mRNA. A previous study demonstrated that
IGF2BP2 knockdown inhibited LPS-induced inflammation of lung
epithelial cells by targeting caspase 4, thus inhibiting the
non-standard scorching pathway (19). In addition, ameliorating
TGF-β-activated kinase 1 binding protein 3
N6-methyladenine modification by
IGF2BP2-dependent mechanisms reduces kidney injury and
inflammation (38). In the present
study, it was found that overexpression of IGF2BP2 reversed
the inhibitory effect of STIM1 knockdown on LPS-induced
apoptosis, inflammation and ERS in A549 cells.
The present study also has certain limitations.
First, it only investigated IGF2BP2-dependent STIM1
inhibition in cells and did not verify it in animals. The
conclusions will be verified in animals in future experiments.
Second, the present study showed that the abnormality of IGF2BP2
can affect the immune response of the body (39). Will the subsequent downregulation
of IGF2BP2 indirectly affect the expression of STIM1,
rather than because of the influence on STIM1? This will be
explored further in future experiments. In addition, as for the
experiments for STIM1 overexpression, it is still necessary
to further verify the experiment and this will be discussed in
future studies.
In conclusion, knockdown of IGF2BP2-regulated STIM1
expression protects against LPS-induced pneumonia in vitro
by alleviating ERS and the inflammatory response.
Supplementary Material
Supplementary Data
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by National K&D
Program of China (grant no. 2022YFC2304800), Guangzhou Medical Key
Discipline (grant no. 2021-2023) and Guangzhou Science and
Technology Planning Project (grant no. 2023A03J0534).
Availability of data and materials
The data sets analyzed and/or generated during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
JH conceived the present study. WZ, QD, NS and ZL
performed the experiments. WZ wrote the manuscript. QD, NS and ZL
processed the experimental data and JH ensured the accuracy of the
experimental data. WZ and JH confirm the authenticity of all the
raw data. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bordon J, Aliberti S, Fernandez-Botran R,
Uriarte SM, Rane MJ, Duvvuri P, Peyrani P, Morlacchi LC, Blasi F
and Ramirez JA: Understanding the roles of cytokines and neutrophil
activity and neutrophil apoptosis in the protective versus
deleterious inflammatory response in pneumonia. Int J Infect Dis.
17:e76–83. 2013.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Hespanhol V and Barbara C: Pneumonia
mortality, comorbidities matter? Pulmonology. 26:123–129.
2020.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Bartoletti M, Azap O, Barac A, Bussini L,
Ergonul O, Krause R, Paño-Pardo JR, Power NR, Sibani M, Szabo BG,
et al: ESCMID COVID-19 living guidelines: Drug treatment and
clinical management. Clin Microbiol Infect. 28:222–238.
2022.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Torres A, Cilloniz C, Niederman MS,
Menendez R, Chalmers JD, Wunderink RG and van der Poll T:
Pneumonia. Nat Rev Dis Primers. 7(25)2021.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Prina E, Ranzani OT and Torres A:
Community-acquired pneumonia. Lancet. 386:1097–1108.
2015.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Khan MM, Yang WL and Wang P: Endoplasmic
reticulum stress in sepsis. Shock. 44:294–304. 2015.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Liu Q, Korner H, Wu H and Wei W:
Endoplasmic reticulum stress in autoimmune diseases. Immunobiology.
225(151881)2020.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Cao X and Wan H and Wan H: Urolithin A
induces protective autophagy to alleviate inflammation, oxidative
stress and endoplasmic reticulum stress in pediatric pneumonia.
Allergol Immunopathol (Madr). 50:147–153. 2022.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Xue Y, Zhou S, Xie W, Meng M, Ma N, Zhang
H, Wang Y, Chang G and Shen X: STIM1-Orai1 interaction exacerbates
LPS-induced inflammation and endoplasmic reticulum stress in bovine
hepatocytes through store-operated calcium entry. Genes (Basel).
13(874)2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Li Y, Feng YF, Liu XT, Li YC, Zhu HM, Sun
MR, Li P, Liu B and Yang H: Songorine promotes cardiac
mitochondrial biogenesis via Nrf2 induction during sepsis. Redox
Biol. 38(101771)2021.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Pan S, Zhao X, Shao C, Fu B, Huang Y,
Zhang N, Dou X, Zhang Z, Qiu Y, Wang R, et al: STIM1 promotes
angiogenesis by reducing exosomal miR-145 in breast cancer
MDA-MB-231 cells. Cell Death Dis. 12(38)2021.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Garrud TAC and Jaggar JH: STIMulating
blood pressure. Elife. 11(e77978)2022.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bolotina VM: Orai1, STIM1, and iPLA2beta
determine arterial vasoconstriction. Arterioscler Thromb Vasc Biol.
32:1066–1067. 2012.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Li H, Jiang Z, Liu X and Yang Z: Higher
plasma level of STIM1, OPG are correlated with stent restenosis
after PCI. Int J Clin Exp Med. 8:21089–21097. 2015.PubMed/NCBI
|
|
15
|
Zhu X, Huang H, Zong Y and Zhang L:
SRY-related high-mobility group box 9 (SOX9) alleviates cigarette
smoke extract (CSE)-induced inflammatory injury in human bronchial
epithelial cells by suppressing stromal interaction molecule 1
(STIM1) expression. Inflamm Res. 71:565–576. 2022.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Liu CC, Miao Y, Chen RL, Zhang YQ, Wu H,
Yang SM and Shang LQ: STIM1 mediates IAV-induced inflammation of
lung epithelial cells by regulating NLRP3 and inflammasome
activation via targeting miR-223. Life Sci.
266(118845)2021.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Meng M, Huo R, Ma N, Chang G and Shen X:
beta-carotene alleviates LPS-induced inflammation through
regulating STIM1/ORAI1 expression in bovine mammary epithelial
cells. Int Immunopharmacol. 113(109377)2022.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Xu X, Shen HR, Zhang JR and Li XL: The
role of insulin-like growth factor 2 mRNA binding proteins in
female reproductive pathophysiology. Reprod Biol Endocrinol.
20(89)2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Wang J, Yuan X and Ding N: IGF2BP2
knockdown inhibits LPS-induced pyroptosis in BEAS-2B cells by
targeting caspase 4, a crucial molecule of the non-canonical
pyroptosis pathway. Exp Ther Med. 21(593)2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Paz I, Kosti I, Ares M Jr, Cline M and
Mandel-Gutfreund Y: RBPmap: A web server for mapping binding sites
of RNA-binding proteins. Nucleic Acids Res. 42:W361–W367.
2014.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen W, Xu S, Xiang L, Zhang Y, Wang C,
Fan T, Huang W and Lu Z: The silencing of SAAL1 suppresses
pneumonia progression via modulating the NLR signaling pathway. Ann
Transl Med. 10(1128)2022.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Fei S, Cao L and Pan L: microRNA-3941
targets IGF2 to control LPS-induced acute pneumonia in A549 cells.
Mol Med Rep. 17:4019–4026. 2018.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Shi J, Wang H, Liu J, Zhang Y, Luo J, Li
Y, Yang C and Jiang J: Ganoderic acid B attenuates LPS-induced lung
injury. Int Immunopharmacol. 88(106990)2020.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Wu H, Xu J, Gong G, Zhang Y and Wu S:
CircARL8B contributes to the development of breast cancer via
regulating miR-653-5p/HMGA2 axis. Biochem Genet. 59:1648–1665.
2021.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Yang R, Liu H, Bai C, Wang Y, Zhang X, Guo
R, Wu S, Wang J, Leung E, Chang H, et al: Chemical composition and
pharmacological mechanism of Qingfei Paidu Decoction and Ma Xing
Shi Gan Decoction against Coronavirus Disease 2019 (COVID-19): In
silico and experimental study. Pharmacol Res.
157(104820)2020.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Gao P, Wang J, Jiang M, Li Z, Xu D, Jing J
and Yihepaer and Hu T: LncRNA SNHG16 is downregulated in Pneumonia
and Downregulates miR-210 to promote LPS-induced lung cell
apoptosis. Mol Biotechnol. 65:446–452. 2023.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Zhang Y, Zhu Y, Gao G and Zhou Z:
Knockdown XIST alleviates LPS-induced WI-38 cell apoptosis and
inflammation injury via targeting miR-370-3p/TLR4 in acute
pneumonia. Cell Biochem Funct. 37:348–358. 2019.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Yang YF, Wang H, Song N, Jiang YH, Zhang
J, Meng XW, Feng XM, Liu H, Peng K and Ji FH: Dexmedetomidine
Attenuates Ischemia/Reperfusion-Induced Myocardial inflammation and
apoptosis through inhibiting endoplasmic reticulum stress
Signaling. J Inflamm Res. 14:1217–1233. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Keestra-Gounder AM, Byndloss MX, Seyffert
N, Young BM, Chavez-Arroyo A, Tsai AY, Cevallos SA, Winter MG, Pham
OH, Tiffany CR, et al: NOD1 and NOD2 signalling links ER stress
with inflammation. Nature. 532:394–397. 2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Kim I, Xu W and Reed JC: Cell death and
endoplasmic reticulum stress: Disease relevance and therapeutic
opportunities. Nat Rev Drug Discov. 7:1013–1030. 2008.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Wang X, Yuan J, Wang H, Gan N, Zhang Q,
Liu B, Wang J, Shu Z, Rao L, Gou X, et al: Progranulin decreases
susceptibility to streptococcus pneumoniae in influenza and
protects against lethal coinfection. J Immunol. 203:2171–2182.
2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wang QL, Yang L, Peng Y, Gao M, Yang MS,
Xing W and Xiao XZ: Ginsenoside Rg1 regulates SIRT1 to ameliorate
sepsis-induced lung inflammation and injury via inhibiting
endoplasmic reticulum stress and inflammation. Mediators Inflamm.
2019(6453296)2019.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Wu JP, Li XZ, Wang Y, Ma L, Yao TW, Zhang
YY and Long F: Effects of electroacupuncture and intracerebral
injection of VEGF on Caspase12, Caspase3, and GRP78 genes in rats
with cerebral ischemia-reperfusion injury. Sichuan Da Xue Xue Bao
Yi Xue Ban. 50:34–39. 2019.PubMed/NCBI(In Chinese).
|
|
35
|
Parker NJ, Begley CG, Smith PJ and Fox RM:
Molecular cloning of a novel human gene (D11S4896E) at chromosomal
region 11p15.5. Genomics. 37:253–256. 1996.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Song X, Liu Y, Dong L and Wang Y:
Stromal-Interacting Molecule 1 (Stim1)/Orai1 modulates endothelial
permeability in ventilator-induced lung injury. Med Sci Monit.
24:9413–9423. 2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Fan H, Huang H, Hu L, Zhu W, Yu Y, Lou J,
Hu L and Chen F: The activation of STIM1 mediates S-phase arrest
and cell death in paraquat induced acute lung intoxication. Toxicol
Lett. 292:123–135. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Wang JN, Wang F, Ke J, Li Z, Xu CH, Yang
Q, Chen X, He XY, He Y, Suo XG, et al: Inhibition of METTL3
attenuates renal injury and inflammation by alleviating TAB3 m6A
modifications via IGF2BP2-dependent mechanisms. Sci Transl Med.
14(eabk2709)2022.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Zhou L, Li H, Cai H, Liu W, Pan E, Yu D
and He S: Upregulation of IGF2BP2 promotes oral squamous cell
carcinoma progression that is related to cell proliferation,
metastasis and tumor-infiltrating immune cells. Front Oncol.
12(809589)2022.PubMed/NCBI View Article : Google Scholar
|