Introduction
Allergic rhinitis (AR) is one of the most common
nasal diseases in the world, and is usually endured throughout life
(1). It is estimated that the
prevalence of AR accounts for 10-30% of the global population
(2). AR is defined as an
immunoglobulin E (IgE)-mediated inflammatory disease of the nasal
mucosa caused by allergen exposure, and is characterized by
sneezing, nasal congestion, itching and rhinorrhea (3).
In classical AR pathophysiology, elevated allergen
specific IgE levels and T-helper (Th) 1/Th2 imbalance are
considered to be major deviant immune factors (4). Epithelial cells play a key protective
role, since they are the first physiological barrier against
allergen infiltration (5). Th2
cytokines also induce the breakdown of the epithelial barrier in
allergic diseases, and epithelial cells release cytokines to
promote Th2 immune response (6).
Therefore, preservation of the epithelial barrier function can
reduce the type 2 immune response caused by the influx of harmful
environmental molecules, including allergens, which is considered
as a promising therapeutic strategy for AR therapy (7).
Hyperforin, a bicyclic tetraketone with four
isoprenoid chains, is a major active constituent of Hypericum
perforatum L. (8). It is also
known as St. John's wort, and is widely used for the treatment of
depressive disorders (9). In
recent years, it has been revealed that hyperforin has various
pharmacological activities, such as anti-diabetic, antitumor and
anti-dementia activities (10-12).
In addition, hyperforin has been revealed to have anti-inflammatory
properties and is resistant to barrier damage. For instance,
hyperforin can improve imiquimod-induced psoriasis-like skin
inflammation in mice by regulating γδ T cells that produce IL-17A
(13). Moreover, hyperforin
promotes the expression of tight junction proteins in endothelial
cells after blood-brain barrier injury (14). However, whether hyperforin is
involved in nasal epithelial cell damage caused by AR is unknown.
Thus, the present study aimed to explore the functional roles of
hyperforin on IL-13-stimulated nasal epithelial cells and discuss
the mechanism by which hyperforin influences nasal epithelial cell
viability, inflammation and barrier damage.
Materials and methods
Bioinformatics analysis
SWISS TargetPrediction (http://www.swisstargetprediction.ch/) was used to
identify the potential target genes of hyperforin, while DisGeNET
(https://www.disgenet.org/) was used to
identify AR target genes. Intersection genes were obtained through
Venn diagrams (https://bioinfogp.cnb.csic.es/), and the network was
presented using the STRING database (https://cn.string-db.org/). The intersection genes
were analyzed by Gene Ontology (GO) (http://geneontology.org/) and Pathway
(genome.jp/kegg/) databases. AutoDockTools 1.5.6 software
(https://autodock.scripps.edu/) was
applied to process proteins, while Discovery Studio 4.5 software
(BIOVIA) was used to visualize the results from the molecular
docking analyses to obtain 2- and 3D images.
Cell culture and treatment
The human nasal epithelial cell line JME/CF15 was
obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences and cultured in DMEM/F12 (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% fetal bovine serum
(HyClone; Cytiva) and 1% penicillin/streptomycin in a humidified
atmosphere with 5% CO2 at 37˚C. JME/CF15 cells were
treated with 10 ng/ml IL-13 (cat. no. HY-P7033; MedChemExpress) at
37˚C for 30 min (15), with or
without treatment with hyperforin at doses of 0.1, 1 and 10 µM for
24 h [the doses of hyperforin were selected referring to previous
study (13)].
Cell transfection
Specific small interfering RNA (siRNA) targeting
B-cell lymphoma 6 (BCL6) (siRNA-BCL6-1 forward,
5'-GCGUCAUGCUUGUGUUAUAAC-3', and reverse,
5'-UAUAACACAAGCAUGACGCAG-3'; siRNA-BCL6-2 forward,
5'-GGUGGUUGAAGCUGGUUAAAG-3', and reverse:
5'-UUAACCAGCUUCAACCACCAG-3') and the corresponding scrambled
negative control (NC) siRNA (siRNA-NC forward,
5'-UUCUCCGAACGUGUCACGU-3' and reverse, 5'-ACGUGACACGUUCGGAGAA-3')
were synthesized by Shanghai GenePharma Co., Ltd. A total of 100 nM
siRNAs were transfected into JME/CF15 cells using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
After 48 h of transfection, cells were harvested for subsequent
experiments.
Cell Counting Kit-8 (CCK-8) assay
JME/CF15 cells were seeded into 96-well plates at a
density of 5x104 cells/ml, treated with different doses
of hyperforin (0.1, 1 and 10 µM) and cultured at 37˚C for 24 h.
Next, 10 µl of WST-8 (Beyotime Institute of Biotechnology) was
added to each well and the plates were incubated at 37˚C for 2 h.
The absorbance value at 450 nm was measured with a microplate
reader (Bio-Rad Laboratories, Inc.).
ELISA
The concentrations of TNFα, IL-1β, IL-6 and CCL11 in
JME/CF15 cells were determined by ELISA assay kits related to TNFα
(cat. no. F3768), IL-6 (cat. no. F3743) and IL-1β (cat. no. F3739;
all from Shanghai Westang Biotechnology Co., Ltd.) according to the
manufacturer's protocols. The absorbance was measured at 450 nm
with a microplate reader (BioTek Instruments, Inc.).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from JME/CF15 cells by
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) in accordance with the manufacturer's protocol. NanoDrop 3000
spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific,
Inc.) was used according to the manufacturer's protocol to confirm
the quality of total RNA. cDNA Synthesis Kit (Takara Bio, Inc.) was
used to reverse transcribe 2 µg RNA into cDNA at 37˚C for 15 min
and 85˚C for 5 sec. Next, amplification of cDNA was performed by
real-time qPCR using the SYBR Premix Ex Taq™ II kit
(Takara Bio, Inc.). The primer sequences for PCR were as follows:
TNF-α, 5'-GACAAGCCTGTAGCCCATGT-3' (forward) and
5'-GGAGGTTGACCTTGGTCTGG-3' (reverse); IL-1β,
5'-AACCTCTTCGAGGCACAAGG-3' (forward) and 5'-AGATTCGTAGCTGGATGCCG-3'
(reverse); IL-6, 5'-CCTTCGGTCCAGTTGCCTT-3' (forward) and
5'-ATTTGTGGTTGGGTCAGGGG-3' (reverse); BCL6,
5'-CCCACTCCCATGTGTCTTCA-3' (forward) and 5'-CGGCTCCAAGGTTAGTGTGT-3'
(reverse); GAPDH, 5'-GGGAAACTGTGGCGTGAT-3' (forward) and
5'-GAGTGGGTGTCGCTGTTGA-3' (reverse). The thermocycling conditions
were as follows: Initial denaturation at 95˚C for 3 min; followed
by 40 cycles of denaturation at 95˚C for 30 sec, annealing at 60˚C
for 30 sec and extension at 72˚C for 30 sec. The relative mRNA
level was normalized with GAPDH using the 2-ΔΔCq method
(16).
Immunofluorescence staining
Following hyperforin treatment and indicated
transfection, IL-13-induced JME/CF15 cells were fixed in 4%
polyoxymethylene for 30 min at room temperature and permeabilized
with 0.5% Triton-X100 for 7 min at room temperature. After blocking
with 2% BSA for 1 h at room temperature, cells were incubated with
a primary antibody against zonula occludens-1 (ZO-1; 1:100; cat.
no. ab221547; Abcam) at 4˚C overnight. Next, goat anti-rabbit IgG
H&L (Alexa Fluor® 488) secondary antibody (1:400;
cat. no. ab150077; Abcam) was added and incubated for 1 h at room
temperature. The nuclei were stained using a DAPI solution for 5
min at room temperature, and the cells were subsequently visualized
under a fluorescence microscope (Eclipse80i; Nikon
Corporation).
Western blot analysis
Total proteins were extracted from cells using RIPA
buffer (Beyotime Institute of Biotechnology) and quantified using
the bicinchoninic acid method (Thermo Fisher Scientific, Inc.).
Proteins (20-30 µg per lane) were separated using 10% SDS-PAGE
(Bio-Rad Laboratories, Inc.), transferred onto PVDF membranes
(MilliporeSigma), and blocked for 1 h in 5% non-fat milk in 0.1%
TBS-Tween-20 at room temperature. The membranes were incubated at
4˚C overnight with primary antibodies against ZO-1 (cat. no.
ab276131), occludin (cat. no. ab216327), claudin 1 (cat. no.
ab211737), BCL6 (cat. no. ab33901), phosphorylated (p)-p38 (cat.
no. ab45381) (all from Abcam), p38 (cat. no. AF7668; Beyotime
Institute of Biotechnology) and β-actin (1:1,000; cat. no. ab8227;
Abcam). Subsequently, the membranes were incubated with
HRP-conjugated secondary antibodies (1:5,000; cat. no. ab6721;
Abcam) for 1 h at room temperature. Finally, the protein bands were
visualized using an ECL detection system (Beyotime Institute of
Biotechnology) and quantified using densitometry with Q uantityOne
4.5.0 software (Bio-Rad Laboratories, Inc.).
Statistical analysis
All statistical analyses were conducted using SPSS
version 18.0 software (SPSS, Inc.). The results were analyzed by
one-way ANOVA followed by Bonferroni's post-hoc test for multiple
comparisons. Data are presented as the mean ± standard deviation of
three independent experiments. P<0.05 was considered to indicate
a statistically significant difference.
Results
Analysis of intersection genes between
hyperforin and AR
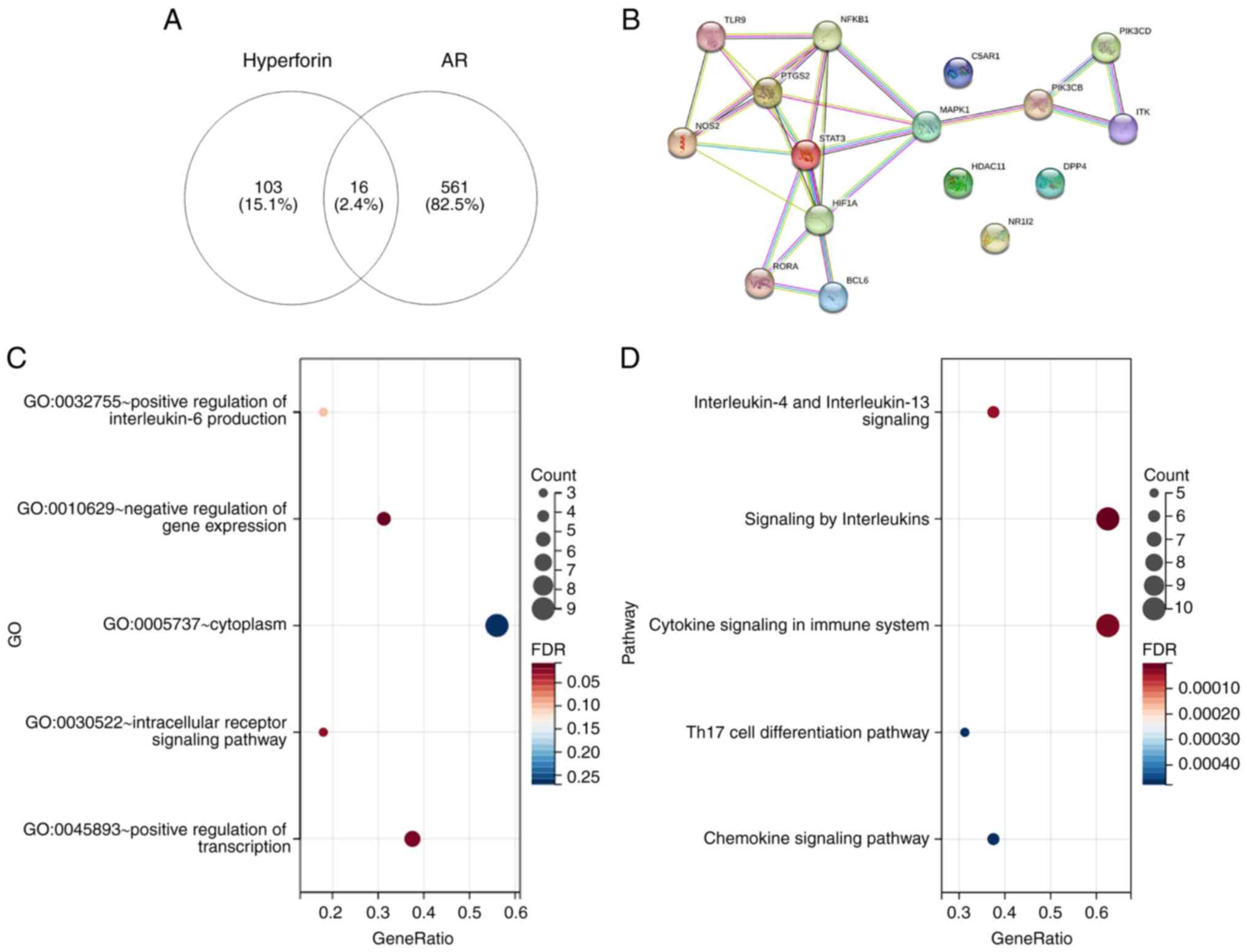
Using Venn diagrams (https://bioinfogp.cnb.csic.es/), 16 intersection genes
of hyperforin and AR were obtained, and the network was presented
using the STRING database (https://cn.string-db.org/) (Fig. 1A and B). The intersection genes were then
analyzed using the GO and Pathway databases, which demonstrated
that the 16 intersection genes were mainly located in the
‘cytoplasm’, were involved in ‘(...)regulation of transcription’
and were enriched in ‘signaling by interleukins’/‘cytokine
signaling in immune system’/‘chemokine signaling pathway’/‘IL-4 and
IL-13 signaling’ (Fig. 1C and
D).
Hyperforin suppresses the release of
nasal epithelial inflammatory cytokines induced by IL-13
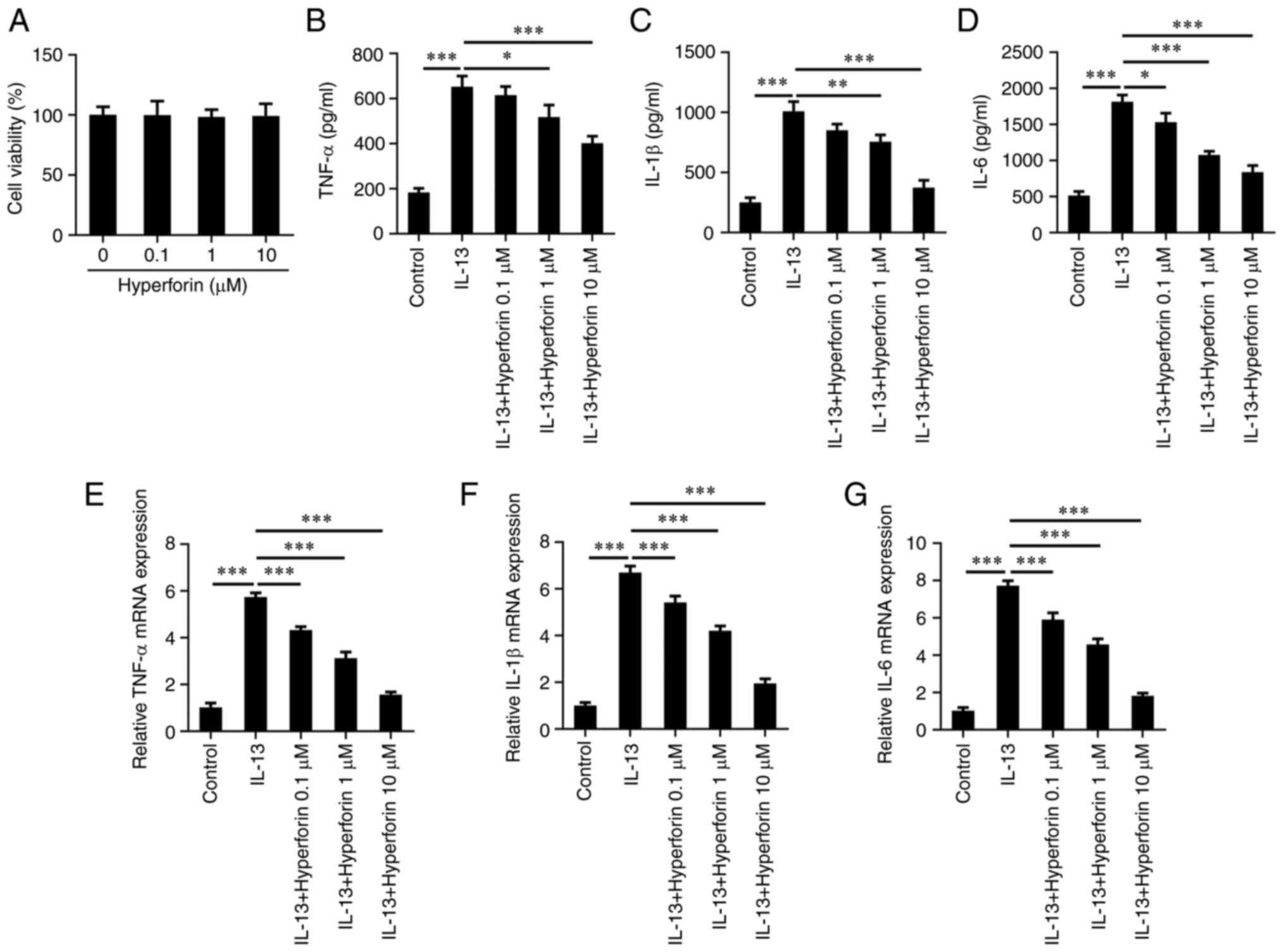
To investigate the effects of hyperforin in AR, the
effects of hyperforin on the viability of JME/CF15 cells were first
detected. As presented in Fig. 2A,
0.1-100 µM hyperforin had no obvious effects on JME/CF15 cell
viability. In addition, ELISA revealed that the protein levels of
TNFα, IL-1β and IL-6 in cells treated with IL-13 were significantly
increased compared with those of control cells, but treatment with
hyperforin reduced the protein levels of TNFα, IL-1β and IL-6
induced by IL-13 in a dose-dependent manner (Fig. 2B-D). Moreover, RT-qPCR also
revealed that IL-13 induced the production of TNFα, IL-1β and IL-6,
which was reversed by hyperforin treatment (Fig. 2E-G).
Hyperforin restrains IL-13-induced
JME/CF15 cell barrier damage
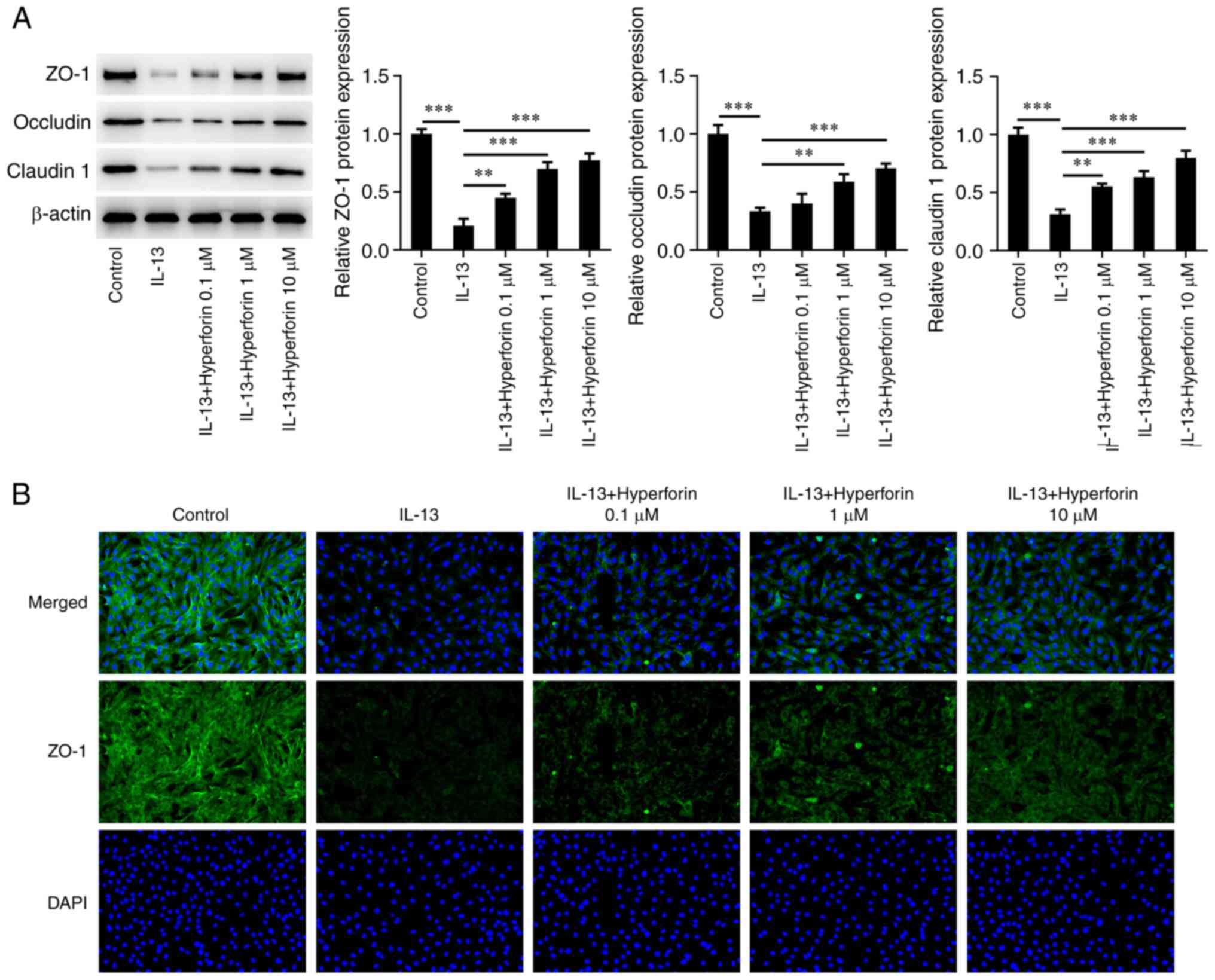
The present study next explored the effects of
hyperforin on IL-13-treated JME/CF15 cells and the cell barrier. As
presented in Fig. 3A, western
blotting revealed that IL-13 significantly reduced the levels of
ZO-1, occludin and claudin 1 in JME/CF15 cells compared with the
control, while addition of hyperforin increased the IL-13-reduced
levels of these three proteins in a dose-dependent manner. In
addition, immunofluorescence staining revealed that the number of
positive cells in the IL-13 group was decreased, but increased
after hyperforin treatment (Fig.
3B).
Hyperforin increases BCL6 and inhibits
the p38MAPK/C-C motif chemokine 11 (CCL11) pathway in IL-13-induced
JME/CF15 cells
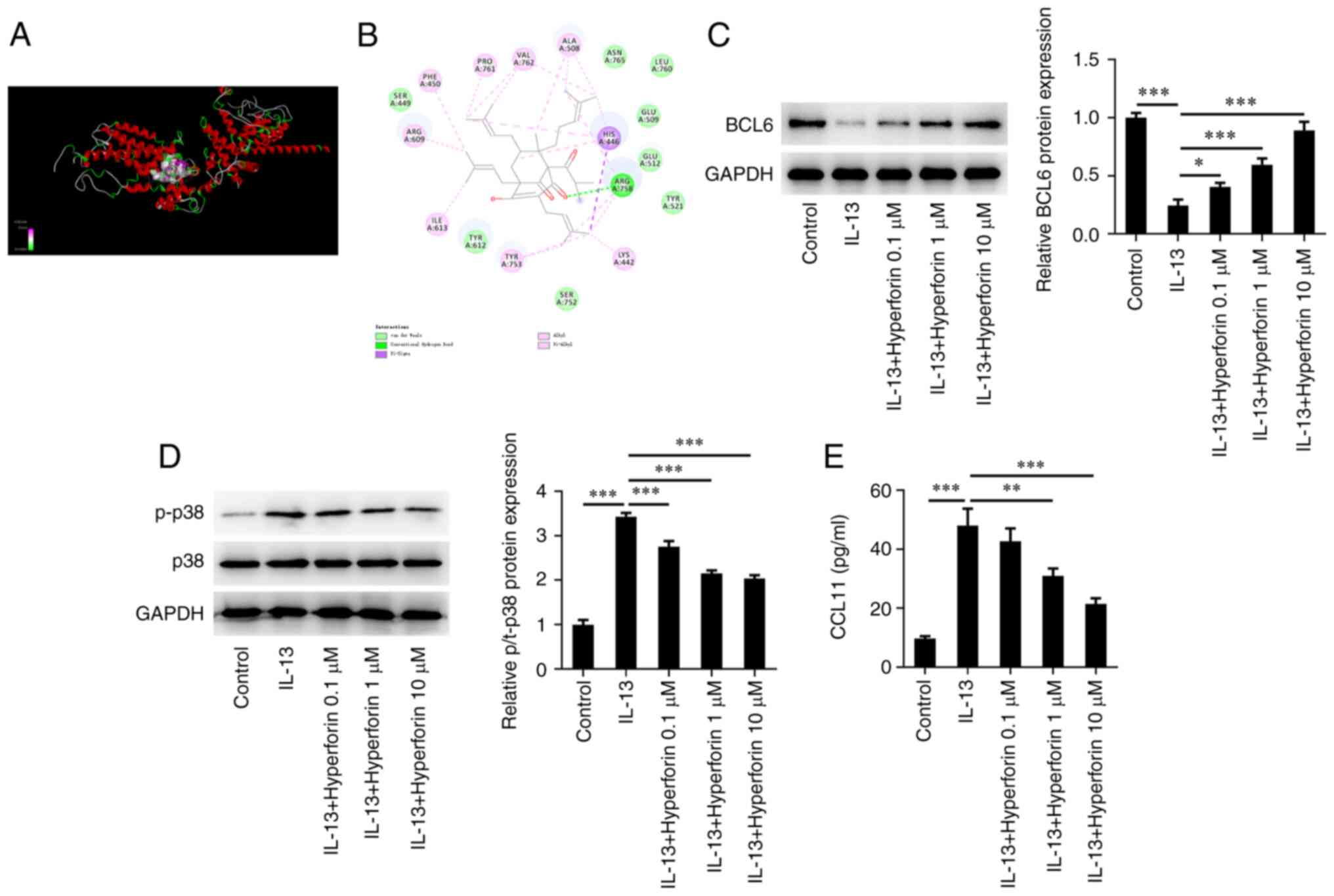
AutoDock software was used to dock hyperforin with
the BCL6 protein at the molecular level, and the result revealed
that the binding free energy of this active ingredient and BCL6 was
-5.8 kcal/mol (Fig. 4A). The
molecular docking diagram is presented in Fig. 4B. The expression levels of
molecules associated with the BCL6/p38MAPK/CCL11 pathway were then
detected. Western blotting revealed that treatment with IL-13
significantly reduced the protein level of BCL6, which was
significantly reversed by treatment with hyperforin in a
dose-dependent manner (Fig. 4C).
Moreover, IL-13 significantly elevated the levels of p-p38 and
CCL11, whereas hyperforin suppressed the expression levels of p-p38
and CCL11 in IL-13-induced cells (Fig.
4D and E).
Hyperforin inhibits IL-13-induced
nasal epithelial inflammation and cell barrier damage by activating
BCL6
To explore the biological roles of BCL6 in
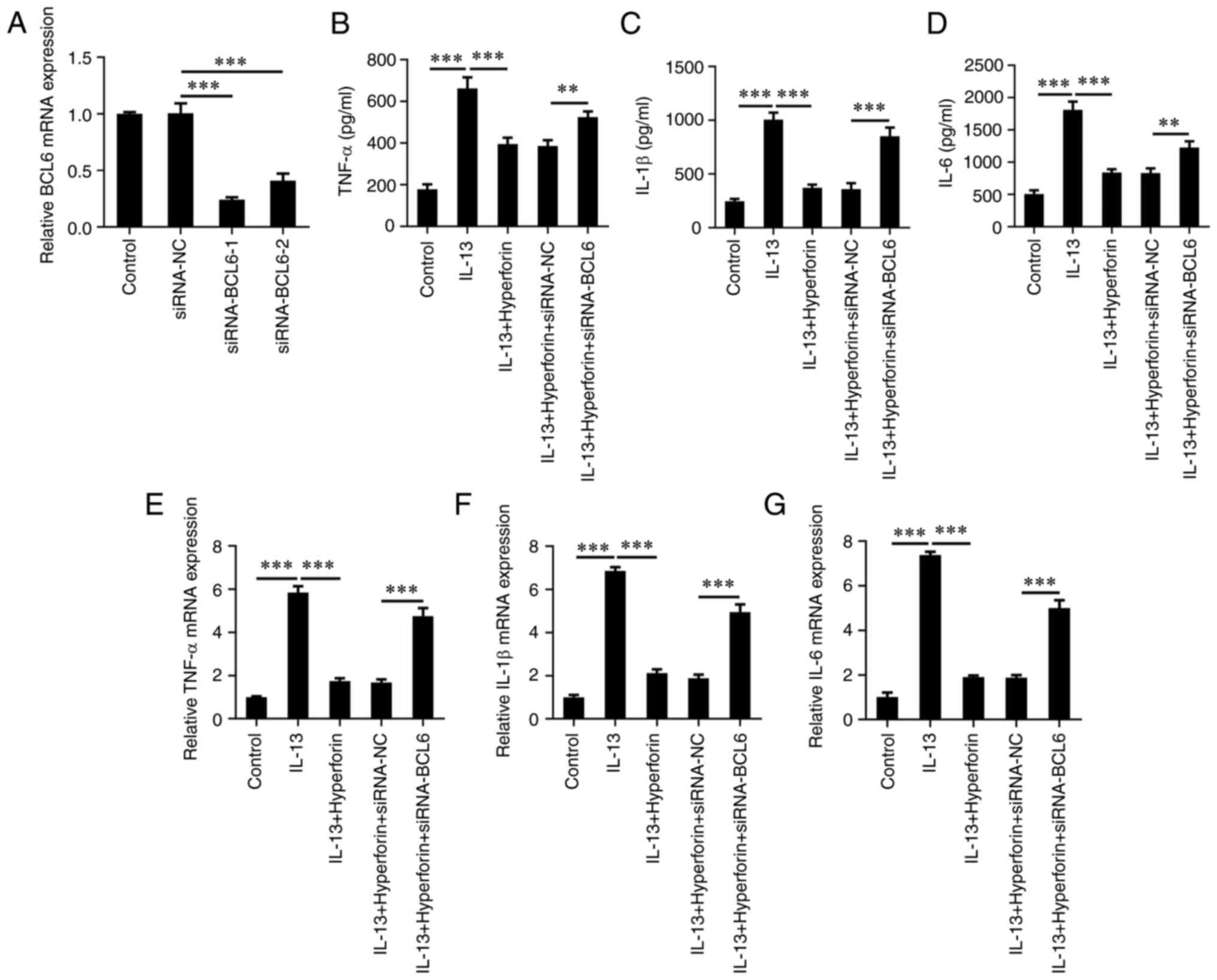
hyperforin-treated AR, siRNA-BCL6-1/2 was transfected to silence
BCL6 in JME/CF15 cells. The results demonstrated that siRNA-BCL6-1
had the greater transfection efficiency; thus, siRNA-BCL6-1 (named
as siRNA-BCL6) was selected for subsequent assays (Fig. 5A). The results of ELISA
demonstrated that co-treatment of hyperforin and BCL6-silencing
significantly increased the levels of TNFα, IL-1β and IL-6 in
JME/CF15 cells compared with the hyperforin-treated cells alone
(Fig. 5B-D). In addition, RT-qPCR
also demonstrated the results that BCL6-silencing enhanced the
levels of these inflammatory factors compared with the
hyperforin-treated cells (Fig.
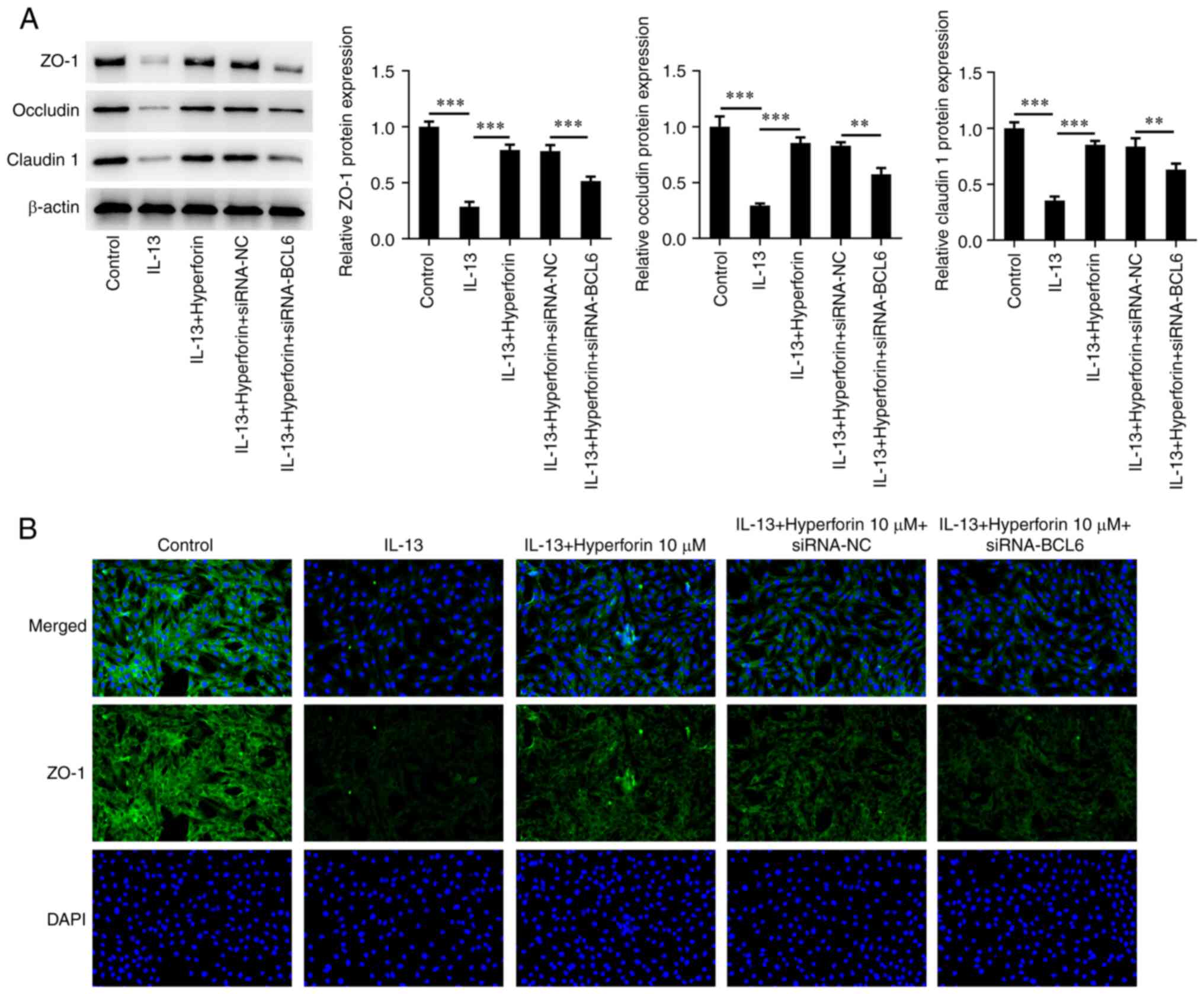
5E-G). Additionally, siRNA-BCL6 significantly inhibited the
production of ZO-1, occludin and claudin 1 in IL-13-induced cells
treated with hyperforin (Fig. 6A).
Immunofluorescence assay demonstrated the number of positive cells
after silencing of BCL6 was notably decreased compared with that in
hyperforin-treated cells (Fig.
6B).
Discussion
AR is a growing public health, medical and economic
problem worldwide (17). The risk
factors for AR include environmental exposure, climate change and
lifestyle (18). Although the
pathological mechanisms and treatment of AR have been widely
researched, numerous aspects of AR remain unclear. At present,
management of AR includes patient education, pharmacotherapy,
allergen-specific immunotherapy and biologics (19).
Despite the variety of medications for AR, such as
glucocorticosteroids, decongestants, histamine receptor and
leukotriene blockers and immunotherapy, multiple patients still
experience treatment failures or unsatisfactory results (20). Emerging evidence had demonstrated
that allergic diseases, including AR, are often caused by numerous
inflammatory mediators such as histamine, chemokines and cytokines
from immune cells (21,22). IL-13 has been demonstrated to
promote mucus production and the secretion of inflammatory
cytokines in AR (23). The present
study aimed to determine the therapeutic potential of hyperforin in
AR cell inflammation and barrier function, and the molecular
mechanisms underlying its function. AR is mostly affected by
genetics and immunity; however, it is currently hypothesized that
rhinitis can be treated with medication or surgery to relieve
symptoms (24). The present study
revealed the protective effects of hyperforin on the inflammatory
response and barrier injury in IL-13-treated nasal mucosal
epithelial cells, which revealed that hypericin may be a novel
alternative drug used to treat AR.
Hyperforin is a pharmacologically active component
of the medicinal plant Hypericum perforatum, and has been
reported to exhibit anti-inflammatory activity and resistance to
barrier damage (25-27).
Novelli et al (23)
reported that hyperforin protects pancreatic β cells in
vitro against the deleterious effects of immune and
inflammatory cytokines by inhibiting multiple phosphorylation steps
of cytokine signaling. In addition, hyperforin reduces inflammatory
cell infiltration in the spinal cord, and promotes the
differentiation of regulatory T cells and Th2 cells by adjusting
their master transcription factors to reduce the severity of
autoimmune encephalomyelitis (9).
Moreover, hyperforin has been reported to promote wound healing and
to function as a topical medication for atopic dermatitis (28,29).
Takada et al (26) also
revealed that hyperforin/2-hydroxypropyl-β-cyclodextrin recovers
impaired Ca2+ responses in the atopic epidermis, which
may facilitate wound closure. The current study revealed that
hyperforin (0.1-100 µM) had no obvious effects on human nasal
epithelial JME/CF15 cell viability, and hyperforin significantly
reduced the release of inflammatory cytokines induced by IL-13 and
withstood the barrier damage.
According to bioinformatics analysis, 16 genes were
revealed to be associated with hyperforin and AR. BCL6 was the one
of these genes, and enrichment analysis demonstrated that BCL6 was
enriched in ‘interleukin-4 and interleukin-13 signaling’,
‘signaling by interleukins’ and ‘cytokine signaling in immune
system’. BCL6 is a sequence-specific transcriptional regulator that
inhibits the transcription of target genes by binding to a specific
DNA sequence in the promoter region (30). It has been reported that BCL6 is
expressed in human nasal mucosal epithelial cells, and the
expression of BCL6 is decreased in the nasal mucosa of patients
with AR (31). Chen et al
(29) reported that BCL6
suppresses the inflammatory response and attenuates renal
inflammation by negatively regulating the transcription of NLRP3 in
human renal tubular epithelial cells, indicating the importance of
BCL6 in the regulation of human inflammation diseases. The present
study used AutoDock software to dock hyperforin with BCL6 protein
successfully. It was revealed that treatment with hyperforin
increased the expression of BCL6, while silencing of BCL6 reversed
the inhibitory effects of hyperforin on IL-13-induced nasal
epithelial cell inflammation and barrier injury.
BCL6 can negatively regulate the expression of
p38MAPK signaling molecules (32).
In addition, p38 has been reported to be involved in AR injury and
to activate the downstream expression of CCL11 (33-35).
Therefore, it was hypothesized whether hyperforin could inhibit
p38MAPK/CCL11 signaling by activating BCL6 to participate in
IL-13-induced AR injury. The present results demonstrated that
IL-13 enhanced the expression of p-p38 and CCL11 in JME/CF15 cells,
and hyperforin inhibited the production of p-p38 and CCL11 induced
by IL-13, which was consistent with the aforementioned hypothesis.
However, there are several limitations in the present study. The
association between BCL6 and AR was demonstrated and rescue
experiments were also performed. However, no animal experiments
were performed in the current study, and, therefore, relevant
animal experiments will be performed in the future. Moreover, the
present study did not explore the separate effects of hyperforin on
AR inflammation and barrier injury, and this will be explored in
the future.
In conclusion, the present results demonstrated that
hyperforin reduced inflammatory response and barrier injury in
IL-13-treated nasal mucosal epithelial cells by downregulating p38
MAPK/CCL11 signaling by targeting BCL6, which may provide a novel
insight into prospective novel strategies for AR therapy.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by the Wuhan Medical
Research Project (grant nos. WZ21Q05 and WZ20M01).
Availability of data and materials
All data generated and/or analyzed during this study
are included in this published article.
Authors' contributions
CX and WS designed the study, performed the
experiments and drafted and revised the manuscript. CX analyzed the
data and searched the literature. CX and WS confirm the
authenticity of all the raw data. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Schuler Iv CF and Montejo JM: Allergic
rhinitis in children and adolescents. Pediatr Clin North Am.
66:981–993. 2019.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Schuler Iv CF and Montejo JM: Allergic
rhinitis in children and adolescents. Immunol Allergy Clin North
Am. 41:613–625. 2021.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Hoyte FCL and Nelson HS: Recent advances
in allergic rhinitis. F1000Res. 7(1333)2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Siddiqui ZA, Walker A, Pirwani MM, Tahiri
M and Syed I: Allergic rhinitis: Diagnosis and management. Br J
Hosp Med (Lond). 83:1–9. 2022.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Incorvaia C, Cavaliere C, Frati F and
Masieri S: Allergic rhinitis. J Biol Regul Homeost Agents. 32 (1
Suppl 1):S61–S66. 2018.PubMed/NCBI
|
|
6
|
Meng Y, Wang C and Zhang L: Advances and
novel developments in allergic rhinitis. Allergy. 75:3069–3076.
2020.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Ghezzi M, Pozzi E, Abbattista L, Lonoce L,
Zuccotti GV and D'Auria E: Barrier impairment and Type 2
inflammation in allergic diseases: The pediatric perspective.
Children (Basel). 8(1165)2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Liu S, Yu B, Dai J and Chen R: Targeting
the biological activity and biosynthesis of hyperforin: A
mini-review. Chin J Nat Med. 20:721–728. 2022.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Li XX, Yan Y, Zhang J, Ding K, Xia CY, Pan
XG, Shi YJ, Xu JK, He J and Zhang WK: Hyperforin: A natural lead
compound with multiple pharmacological activities. Phytochemistry.
206(113526)2023.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Wu S, Malaco Morotti AL, Wang S, Wang Y,
Xu X, Chen J, Wang G and Tatsis EC: Convergent gene clusters
underpin hyperforin biosynthesis in St John's wort. New Phytol.
235:646–661. 2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Novelli M, Masiello P, Beffy P and
Menegazzi M: Protective role of St. John's wort and its components
hyperforin and hypericin against diabetes through inhibition of
inflammatory signaling: Evidence from in vitro and in vivo studies.
Int J Mol Sci. 21(8108)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Nawrot J, Gornowicz-Porowska J,
Budzianowski J, Nowak G, Schroeder G and Kurczewska J: Medicinal
herbs in the relief of neurological, cardiovascular, and
respiratory symptoms after COVID-19 infection a literature review.
Cells. 11(1897)2022.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Zhang S, Zhang J, Yu J, Chen X, Zhang F,
Wei W, Zhang L, Chen W, Lin N and Wu Y: Hyperforin ameliorates
imiquimod-induced psoriasis-like murine skin inflammation by
modulating IL-17A-producing γδ T cells. Front Immunol.
12(635076)2021.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Lemmen J, Tozakidis IE and Galla HJ:
Pregnane X receptor upregulates ABC-transporter Abcg2 and Abcb1 at
the blood-brain barrier. Brain Res. 1491:1–13. 2013.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Wang B, Gao Y, Zheng G, Ren X, Sun B, Zhu
K, Luo H, Wang Z and Xu M: Platycodin D inhibits
interleukin-13-induced the expression of inflammatory cytokines and
mucus in nasal epithelial cells. Biomed Pharmacother. 84:1108–1112.
2016.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Kakli HA and Riley TD: Allergic rhinitis.
Prim Care. 43:465–475. 2016.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Campo P, Eguiluz-Gracia I, Bogas G, Salas
M, Plaza Serón C, Pérez N, Mayorga C, Torres MJ, Shamji MH and
Rondon C: Local allergic rhinitis: Implications for management.
Clin Exp Allergy. 49:6–16. 2019.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Zhang Y, Lan F and Zhang L: Advances and
highlights in allergic rhinitis. Allergy. 76:3383–3389.
2021.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Beken B, Eguiluz-Gracia I, Yazıcıoğlu M
and Campo P: Local allergic rhinitis: A pediatric perspective. Turk
J Pediatr. 62:701–710. 2020.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Wang L, Lv Q, Song X, Jiang K and Zhang J:
ADRB2 suppresses IL-13-induced allergic rhinitis inflammatory
cytokine regulated by miR-15a-5p. Hum Cell. 32:306–315.
2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Teng Y, Zhang R, Liu C, Zhou L, Wang H,
Zhuang W, Huang Y and Hong Z: miR-143 inhibits
interleukin-13-induced inflammatory cytokine and mucus production
in nasal epithelial cells from allergic rhinitis patients by
targeting IL13Rα1. Biochem Biophys Res Commun. 457:58–64.
2015.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Novelli M, Menegazzi M, Beffy P, Porozov
S, Gregorelli A, Giacopelli D, De Tata V and Masiello P: St. John's
wort extract and hyperforin inhibit multiple phosphorylation steps
of cytokine signaling and prevent inflammatory and apoptotic gene
induction in pancreatic β cells. Int J Biochem Cell Biol.
81:92–104. 2016.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Pavón-Romero GF, Parra-Vargas MI,
Ramírez-Jiménez F, Melgoza-Ruiz E, Serrano-Pérez NH and Teran LM:
Allergen immunotherapy: Current and future trends. Cells.
11(212)2022.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Meinke MC, Schanzer S, Haag SF, Casetti F,
Müller ML, Wölfle U, Kleemann A, Lademann J and Schempp CM: In vivo
photoprotective and anti-inflammatory effect of hyperforin is
associated with high antioxidant activity in vitro and ex vivo. Eur
J Pharm Biopharm. 81:346–350. 2012.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Takada H, Yonekawa J, Matsumoto M, Furuya
K and Sokabe M: Hyperforin/HP-β-cyclodextrin enhances
mechanosensitive Ca2+ signaling in HaCaT keratinocytes
and in atopic skin ex vivo which accelerates wound healing. Biomed
Res Int. 2017(8701801)2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Brondz I: Super antibiotics, part II.
Hyperforin, mass spectroscopy (MS) and gas chromatography-mass
spectrometry (GC-MS), evidence of permeability of the blood-testis
barrier (BTB) and the blood-brain barrier (BBB) to hyperforin. Int
J Anal Mass Spectrom Chromatogr. 4:66–73. 2016.
|
|
28
|
Marrelli M, Statti G, Conforti F and
Menichini F: New potential pharmaceutical applications of hypericum
species. Mini Rev Med Chem. 16:710–720. 2016.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Chen D, Xiong XQ, Zang YH, Tong Y, Zhou B,
Chen Q, Li YH, Gao XY, Kang YM and Zhu GQ: BCL6 attenuates renal
inflammation via negative regulation of NLRP3 transcription. Cell
Death Dis. 8(e3156)2017.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Béguelin W, Teater M, Gearhart MD, Calvo
Fernández MT, Goldstein RL, Cárdenas MG, Hatzi K, Rosen M, Shen H,
Corcoran CM, et al: EZH2 and BCL6 cooperate to assemble CBX8-BCOR
complex to repress bivalent promoters, mediate germinal center
formation and lymphomagenesis. Cancer Cell. 30:197–213.
2016.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Peng Y, Li XQ and Qiu QH: Detection of
differentially expressed gene of allergic rhinitis based on RT²
profiler PCR array. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za
Zhi. 31:869–872. 2017.PubMed/NCBI View Article : Google Scholar : (In Chinese).
|
|
32
|
Gu Y, Luo M, Li Y, Su Z, Wang Y, Chen X,
Zhang S, Sun W and Kong X: Bcl6 knockdown aggravates hypoxia injury
in cardiomyocytes via the P38 pathway. Cell Biol Int. 43:108–116.
2019.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Long S and Zhang H: MIR-181A-5P attenuates
ovalbumin-induced allergic inflammation in nasal epithelial cells
by targeting IL-33/P38 MAPK pathway. Clin Invest Med. 44:E31–E38.
2021.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Roh KB, Jung E, Park D and Lee J: Fumaric
acid attenuates the eotaxin-1 expression in TNF-α-stimulated
fibroblasts by suppressing p38 MAPK-dependent NF-κB signaling. Food
Chem Toxicol. 58:423–431. 2013.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Park JY, Choi JH, Lee SN, Cho HJ, Ahn JS,
Kim YB, Park DY, Park SC, Kim SI, Kang MJ, et al: Protein arginine
methyltransferase 1 contributes to the development of allergic
rhinitis by promoting the production of epithelial-derived
cytokines. J Allergy Clin Immunol. 147:1720–1731. 2021.PubMed/NCBI View Article : Google Scholar
|