Introduction
Stroke is the second leading cause of death and a
significant cause of disability globally. In 2019, the worldwide
prevalence of stroke was 101.5 million individuals, with ischemic
stroke accounting for 77.2 million of these cases (1). Severe and/or prolonged reductions in
cerebral blood flow result in a lack of oxygen and energy supply to
brain tissues, as well as an accumulation of potentially harmful
substances. The substantial damage to brain tissues can be
attributed to energy failure, loss of cellular homeostasis,
acidosis, increased intracellular calcium, excitotoxicity and
toxicity mediated by free radicals.
Currently, the primary therapeutic strategy against
cerebral ischemia is thrombolysis, which involves dissolving blood
clots to restore blood flow before damage occurs. However,
thrombolysis often leads to reperfusion of the infarcted brain
tissue, thereby causing reperfusion injury (2). Furthermore, numerous studies have
established a time window of 4.5 h for systemic thrombolysis and
early recanalization and reperfusion in ischemic stroke (3). Beyond this window, the risk of
ischemia/reperfusion (I/R) injury significantly escalates and could
lead to catastrophic outcomes, such as fatal edema. Indeed,
oxidative stress is a crucial mechanism involved in the
pathogenesis and disease progression of both cerebral ischemia and
I/R injury. In the pathology of cerebral ischemia and I/R injury,
oxygen and energy deprivation, along with post-translational
modification of oxidative phosphorylation proteins, could elevate
the mitochondrial membrane potential. This, in turn, results in the
excessive generation of reactive oxygen species (ROS) (4). In a clinical study involving stroke
patients, Domínguez et al (5) found that oxidative stress markers,
including malondialdehyde (MDA) and myeloperoxidase, increased when
a stroke occurred. In rat brains with cerebral I/R, an enhanced ROS
production that overwhelmed antioxidant capacity was detected
(6). ROS directly damages all
cellular components, including proteins, DNA, RNA and lipids,
subsequently inducing apoptosis of neuronal cells (7). Therefore, anti-oxidative stress
during cerebral ischemia may be a strategy to rescue the neurons of
the penumbra and ensure their survival. In previous studies,
anti-oxidative stress therapy was shown to improve the prognosis of
cerebral ischemia (8-10).
Galectin-1 is a carbohydrate-binding protein that
belongs to the galectin family. Galectin-1 (Uniprot ID: P09382) is
a small protein consisting of ~135 amino acids. The expression of
galectin-1 is widespread across different tissues and cell types,
including immune cells, endothelial cells, epithelial cells and
neurons. It contains a conserved carbohydrate recognition domain
that allows it to bind specifically to beta-galactoside-containing
glycoconjugates (11). It plays a
crucial role in a wide range of biological processes, including
cell adhesion, immune modulation, angiogenesis and tumor
progression. Galectin-1 has also been implicated in neural
development, tissue remodeling and various disease states.
Galectin-1 influences neural development, axon guidance, and
synapse formation, making it relevant in the context of
neuro-regeneration and repair. Therefore, galectin-1 may contribute
to the survival of neurons in the pathology of cerebral ischemia.
In the present study, the anti-oxidative stress effect of
galectin-1 was investigated, and it was identified that galectin-1
decreased the ischemic area by an antioxidant action in a cerebral
I/R injury mouse model.
Materials and methods
Animals and model of middle cerebral
artery occlusion-reperfusion (MCAO/R)
All the procedures were strictly in line with the
regulations of the National Institutes for Animal Research. All the
animal-related experiments were approved (approval. no. 2022023) by
the Animal Care and Management Committee of Beijing Geriatric
Hospital (Beijing, China). A total of 90 male C57BL/6 mice (25-30
g, 10 weeks-old) were obtained from the Beijing Vital River
Laboratory Animal Technology Co., Ltd. The experimental animals
were housed in conditions with 12/12-h light/dark cycles, a
temperature of 22±3˚C and 60±5% humidity. All the mice had access
to a standard rodent diet and tap water.
A MCAO/R mouse model was created as previously
described (12,13). The mice were anesthetized with
isoflurane (induction dosage 3%; maintenance dosage 1.5%). Through
a midline incision on the ventral side of the neck, the common
carotid arteries were exposed, the external carotid artery (ECA)
and the internal carotid artery (ICA) were carefully isolated.
Subsequently, a 6-0 silicone suture was introduced through the ECA
and advanced into the ICA to occlude the origin of the middle
cerebral artery, ~10-11 mm from the common carotid artery
bifurcation. A laser Doppler flow probe was used to confirm the
development of MCAO. After 60 min of ischemia, the suture was
withdrawn to allow reperfusion. Concurrently, a sham operation was
conducted using the same procedure, except for MCA occlusion. After
72 h of the experiment, the mice were euthanized via CO2
inhalation (50% of the chamber volume per minute), and their brain
tissues were collected.
The protocol of the present study, which involves
treating with galectin-1 for 3 days, was developed based on
previous studies and experimental data of the present study.
According to previous data by the authors on zebrafish (14), a 3-day treatment with galectin-1
significantly reduced the generation of oxidative stress in
vivo. Additionally, the study by Vallecillo-Zúniga et al
(15) observed the anti-ROS effect
of galectin-1 in myogenic cells after 2-3 days of treatment
(15). Cheng et al
(16) found that a 3-day treatment
with galectin-1 could promote vascular remodeling, while
Arda-Pirincci and Aykol-Celik (17) demonstrated that galectin-1 can
reduce the severity of dextran sulfate sodium-induced ulcerative
colitis after at least 3 days of treatment. The preliminary data of
the present study demonstrated the antioxidative stress effects of
galectin-1 in cerebral I/R. Therefore, in the present study, a
treatment duration of 3 days was selected.
The dose and administration route for the present
study were selected based on previous studies and the preliminary
experimental data of the present study. In addition, in the study
by Carlos et al (18), rats
were treated with recombinant human galectin-1 at a dose of 100 µg
per animal, administered intravenously. Similarly, the study by Ye
et al (19) involved
injecting mice with recombinant human galectin-1 at doses ranging
from 250-500 µg per animal, administered intraperitoneally. The
preliminary data of the present study indicated that an injection
of 500 µg of galectin-1 significantly reduced ROS levels in the
mouse brain.
All the male C57BL/6 mice were randomly allocated to
three groups (30/group): i) galectin-1 group, the mice received the
MCAO/R operation and were treated with recombinant human galectin-1
(500 µg/mouse) (18,19) intraperitoneally starting 1 h after
the operation, and this treatment continued for 3 days; ii) MCAO/R
group, the mice received the MCAO/R operation and were treated with
saline (same amount as galectin-1 intraperitoneally from 1 h after
operation, and this treatment continued for 3 days; and iii) sham
surgery group, the mice underwent the sham surgery and received
saline (same amount as galectin-1) intraperitoneally from 1 h after
operation, and this treatment continued for 3 days.
After the brain tissues were collected, 10 mice
brain tissues from each group were subjected to examine cerebral
infarct volume and microglial cell infiltration, 10 were used for
immunohistochemistry (IHC) analysis of allograft inflammatory
factor 1 (IBA-1) expression, and the brain tissues of the remaining
10 mice brain tissues from each group were homogenized and stored
for further tests (ROS and cytokines levels). Galectin-1 (cat. no.
P00388) was purchased from Solarbio Science & Technology Co.,
Ltd. and dissolved in saline for the experiments based on the
previous published study (16).
Cerebral infarct volume
measurement
After the mice were euthanized, the brain tissues
were collected and then sliced to a thickness of 2 mm. The slices
were stained with 2, 3, 5-triphenyl tetrazolium chloride (2%) for
20 min at 37˚C. Thereafter, the slices were fixed using 4%
paraformaldehyde overnight at room temperature. Images of all the
stained slices were captured and quantified using ImageJ software
(version 1.54; National Institutes of Health). Infarct volumes were
calculated using the following formula: Percentage hemisphere
lesion volume (% HLV)=[total infarct volume-(volume of intact
ipsilateral hemisphere-volume of intact contralateral
hemisphere)]/contralateral hemisphere volume x100%.
Rotarod test
The neurological deficits of the mice were evaluated
using the rotarod test. On 4 consecutive days (before surgery, and
24, 48 and 72 h after surgery), the mice were placed on rotating
rods and practiced three times a day. The rods accelerated in speed
from 4 to 40 rpm over a 5-min period. The results were expressed as
the latency to fall from the rod.
Neurological deficit scores
A neurological test was performed to investigate
neurological deficit after 72 h of surgery. The degree of
neurological impairment was evaluated based on the following
features: i) absence of deficits; ii) body bending towards the
opposite side; iii) involuntary contralateral circling; iv)
tendency to fall to the opposite side; and v) lack of spontaneous
movement or a state of unconsciousness (20).
In the present study, the data for the neurological
deficit score were presented, exclusively at the 72-h mark,
following the precedent set by several other studies in the field
(21-25).
These studies, focusing on cerebral ischemia in mouse models,
typically report neurological deficit scores only at the
experiment's endpoint, which is often at 72 h. Consequently, the
reporting methodology of the present study was aligned with this
standard practice.
ROS level measurement
ROS generation was estimated using the
oxidation-sensitive 2',7'-dichlorodihydrofluorescein diacetate
(CM-H2DCFDA) dye (cat. no. KGAF018, Nanjing Jiancheng
Bioengineering Institute). CM-H2DCFDA becomes fluorescent when
oxidized by ROS. In brief, the cerebral cortex tissues of the
ischemic hemisphere were collected and gently homogenized in 500 µl
of PBS. The concentration of proteins was determined by the BCA
method. Thereafter, the brain homogenate was incubated with 25 µl
CM-H2DCFDA (10 mM) for 30 min at 37˚C in dark conditions.
Subsequently, 500 µl of each sample was loaded into the wells of a
96-well plate. The fluorescence intensities of ROS were measured
using a fluorescent microplate reader (excitation: 485 nm,
emission: 525 nm). The activity is expressed as U/mg protein
(26).
Enzyme-linked immunosorbent assay
(ELISA)
The hemispheres containing the ischemic zones of
mice were collected, and then gently homogenized. Tissue
homogenates were diluted in the buffer provided by each kit at a
concentration of 10 mg tissue/ml. The quantities of cytokines,
including interleukin (IL)-6, IL-1 and tumor necrosis factor alpha
(TNF-α), and the activity of superoxide dismutase (SOD), catalase
(CAT), and glutathione peroxidation enzyme (GSH-Px), were measured
using ELISA kits (IL-1, cat. no. PI301; IL-6, cat. no. PI326;
TNF-α, cat. no. PT513; SOD, cat. no. S0101S; CAT, cat. no. S0051;
GSH-Px, cat. no. S0057S) from Beyotime Institute of Biotechnology.
Briefly, 96-well plates were first coated with primary antibodies
(provided within the kits; ready-to-use) and were filled with 100
µl of either the samples or standards. 100 µl of biotinylated
detection antibodies (provided within the kits; ready-to-use) were
then added to these wells and left to incubate at room temperature
for 1 h. Following this, the wells were emptied, rinsed with PBS,
and then filled with 100 µl of the streptavidin complex reagent,
for another h of room temperature incubation. A total of 90 µl of
3,3',5,5'-tetramethylbenzidine substrate solution was next added to
the wells and left to incubate for 20 min at room temperature. Then
50 µl of stop solution was added to the wells. Absorbance was
evaluated using a multimode plate reader at 450 nm. Standard curves
were produced using the optical density (OD) values of standard
reagents, and these were used to ascertain the cytokine levels. The
activity of SOD, CAT and GSH-Px were expressed as U/mg protein.
Assay of MDA levels
The lipid peroxidation product MDA is used as an
indicator of oxidation. The tissue homogenate was analyzed using an
MDA kit following the manufacturers' instructions via the
thiobarbituric acid reaction (cat. no. A003-4-1; Nanjing Jiancheng
Bioengineering Institute). The samples were measured by using a
microplate reader at 532 nm and the values were expressed as
nmol/mg of tissue protein.
IHC
Brain tissues were collected from euthanized mice.
These tissues were first fixed with 10% formalin at room
temperature for 24 h, then embedded in paraffin, sliced into
5-µm-thick sections, and arranged on glass slides. The tissue
sections were subsequently deparaffinized with xylene at 55˚C and
rehydrated in a descending alcohol series (ethanol; 100, 95, 75 and
50%; 3 min each). Thereafter, antigen retrieval was performed by
boiling in citrate-EDTA buffer, followed by a 20-min cooling period
at room temperature. After a blocking step with 5% goat serum (cat.
no. C0265; Beyotime Institute of Biotechnology) at room temperature
for 1 h, the sections were incubated with primary antibodies
diluted against IBA-1 rabbit monoclonal (1:200; cat. no. MA5-36257;
Thermo Fisher Scientific, Inc.) and left overnight at 4˚C.
Following this, the slides were rinsed with TBS with 0.1% Tween-20,
and treated with a horseradish peroxidase-labelled dextran polymer
bound with an anti-rabbit antibody (1:1,000; cat. no. A0279;
Beyotime Institute of Biotechnology) for 1 h at room temperature.
Subsequently, the slides were incubated with DAB solution and
inspected under a light microscope (Olympus Corporation). The
expression of IBA-1 was assessed under a light microscope at a x40
magnification. The expression level was quantified using a standard
IHC staining grade system by measuring positive expression in 5
random fields within the brain tissues, received from three
separate non-adjacent sections per mouse.
In addition, a preliminary experiment was conducted
to validate the protocol. This included optimizing blocking
reagents and antigen retrieval, as well as determining the
appropriate concentrations for primary and secondary antibodies.
Additionally, the DAB staining step was carefully monitored under a
light microscope to prevent overreaction. Furthermore, both
positive and negative controls were employed in the staining
process. The positive control tissue exhibited strong and specific
staining for the target antigen, while no positive staining was
observed in the negative control slides.
Statistical analysis
All the data were analyzed in SPSS 26 (IBM Corp.)
for statistical computing and graphics. The quantitative data were
expressed as the mean ± standard deviation (SD). Statistical
comparisons were performed by one-way ANOVA followed by the Tukey's
post hoc test or unpaired Student's t-test as described in figure
legends. For data that were not normally distributed in the
analyses of neurological scores, The Mann-Whitney U test was used
for comparisons between 2 groups. Two-way mixed ANOVA followed by
the Bonferroni's post hoc test was used to compare the difference
in rotarod test among different groups at 24, 48 and 72 h after
surgery. P<0.05 was considered to indicate a statistically
significant difference.
Results
Galectin-1 ameliorates the
neurological deficit and infarct size in MCAO/R mice
In the present study, the results revealed that
cerebral ischemia developed in mice after MCAO/R surgery. The
neurological outcome was investigated after 3 days of galectin-1
treatment in MCAO/R mice. Compared with the control group that
underwent sham surgery, both the neurological deficit and the
cerebral infarct area were observed in the MCAO/R mice group
(Fig. 1A-D). Moreover, the results
demonstrated that treatment with galectin-1 significantly improved
the outcome of cerebral ischemia, as evidenced by longer latency on
the rotarod, lower neurological deficit scores, and smaller infarct
volumes. Data on the neurological deficit score for mice that
underwent sham surgery were not included, as no deficits were
detected in these mice.
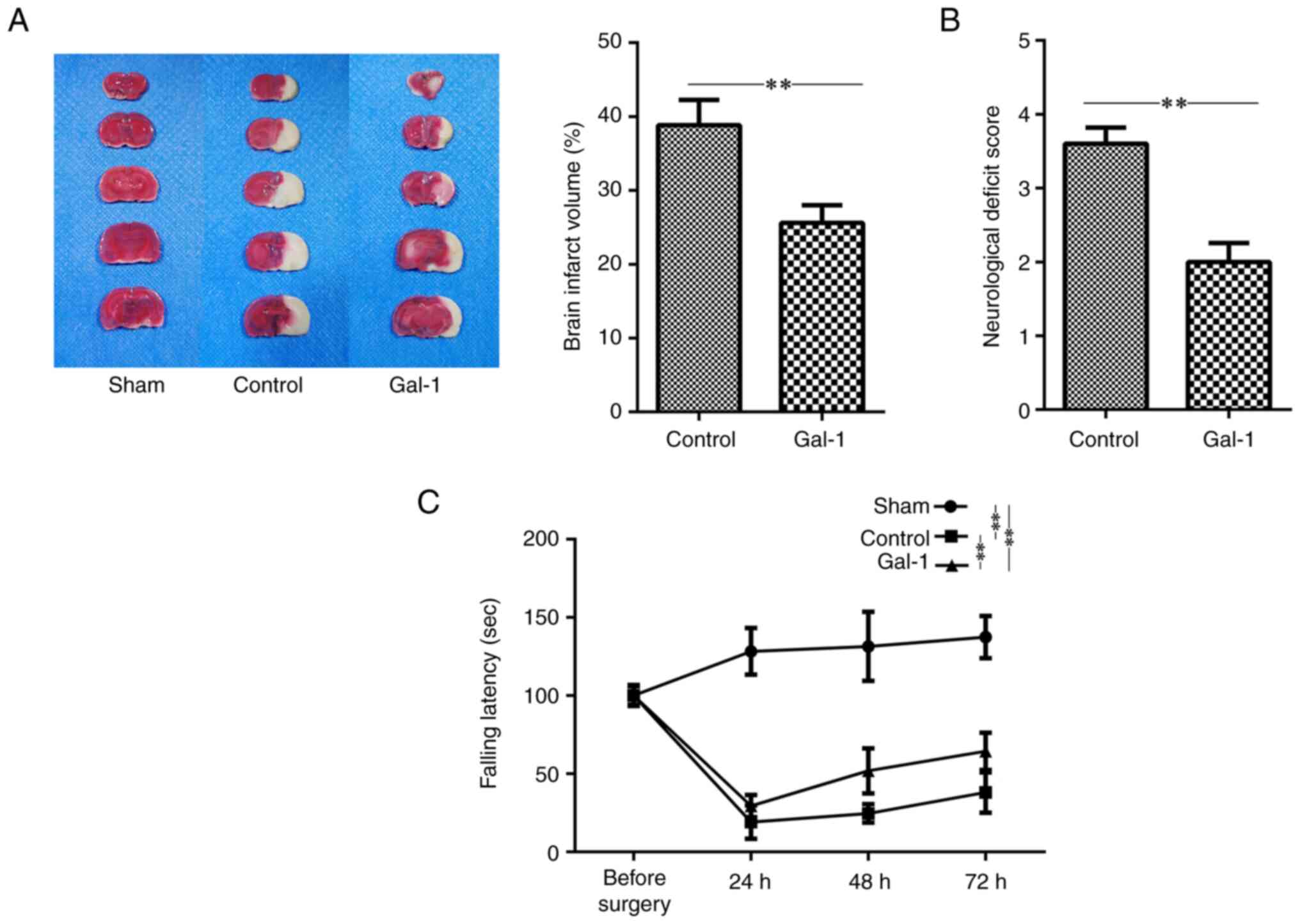 | Figure 1Galectin-1 ameliorates neurological
deficits and reduces infarct volumes in MCAO/R mice. The mice
received MCAO/R surgery, MCAO/R surgery + galectin-1 treatment, or
sham surgery (n=10 per group). (A) Representative figures of brain
sections and statistical results. The data revealed that the
infarct volumes were significantly reduced in the MCAO/R +
galectin-1 group compared with the non-galectin-1 MCAO/R group. (B
and C) Neurological deficit scores were assessed at 72 h after
surgery in MCAO/R mice, while rotarod tests were performed on 4
consecutive days (before surgery, and 24, 48 and 72 h after
surgery). Galectin-1 treatment improved motor ability. The tests
showed that mice which received galectin-1 treatment exhibited
improved motor function, with (B) lower neurological deficit scores
and (C) longer duration on the rotarod. Analyses were performed to
compare the differences in the latency to fall from the rod among
various groups at 24, 48 and 72 h after surgery. These groups
included those undergoing MCAO/R surgery, MCAO/R surgery with
galectin-1 treatment, and sham surgery. Compared with the sham
surgery group at 24, 48 and 72 h after surgery, respectively, the
latencies to fall from the rod were significantly decreased in both
the MCAO/R mice with and without Gal-1 treatment. However, the
latency to fall from the rod in the galectin-1 + MCAO/R group was
significantly higher than that in the non-galectin-1 MCAO/R group
at 24, 48 and 72 h after surgery, respectively (Sham vs. control at
24, 48 and 72 h after surgery, P<0.01; Sham vs. Gal-1 at 24, 48
and 72 h after surgery, P<0.01; Gal-1 vs. control at 24, 48 and
72 h after surgery, P<0.01). The data are presented as the mean
± SD. **P<0.01 by Student's t-test or two-way mixed
ANOVA followed by Bonferroni's post hoc test. MCAO/R, middle
cerebral artery occlusion-reperfusion; Gal-1, galectin-1
treatment. |
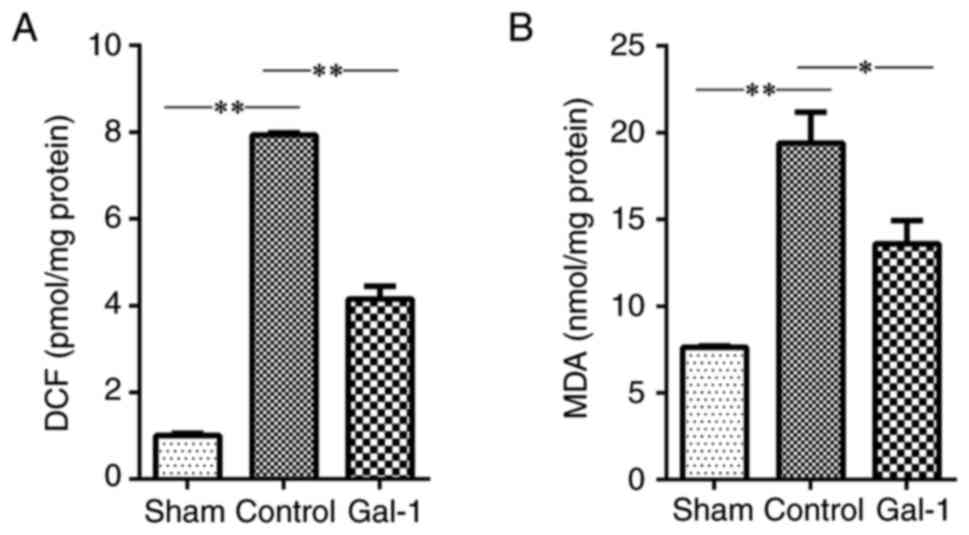
Galectin-1 reduces ROS production and
lipid peroxidation
ROS and MDA are both biomarkers for the status of
oxidative stress. The data of the present study identified an
increased level of both ROS (Fig.
2A) and MDA (Fig. 2B) in the
ischemic hemisphere of MCAO/R mice, compared with that in the sham
surgery. In addition, treatment with galectin-1 significantly
decreased the levels of ROS and MDA in the ischemic hemisphere of
MCAO/R mice, compared with that in MCAO/R mice without galectin-1
treatment.
Galectin-1 treatment improves
antioxidant defenses
The data of the present study revealed a
significantly decreased level of antioxidant defense molecules,
including SOD, CAT and GSH-Px, in the ischemic hemisphere of MCAO/R
mice, compared with that in the sham surgery. However, compared
with the non-galectin-1 treatment group, treatment with galectin-1
significantly increased the levels of SOD, CAT and GSH-Px in the
ischemic hemisphere of MCAO/R mice (Fig. 3).
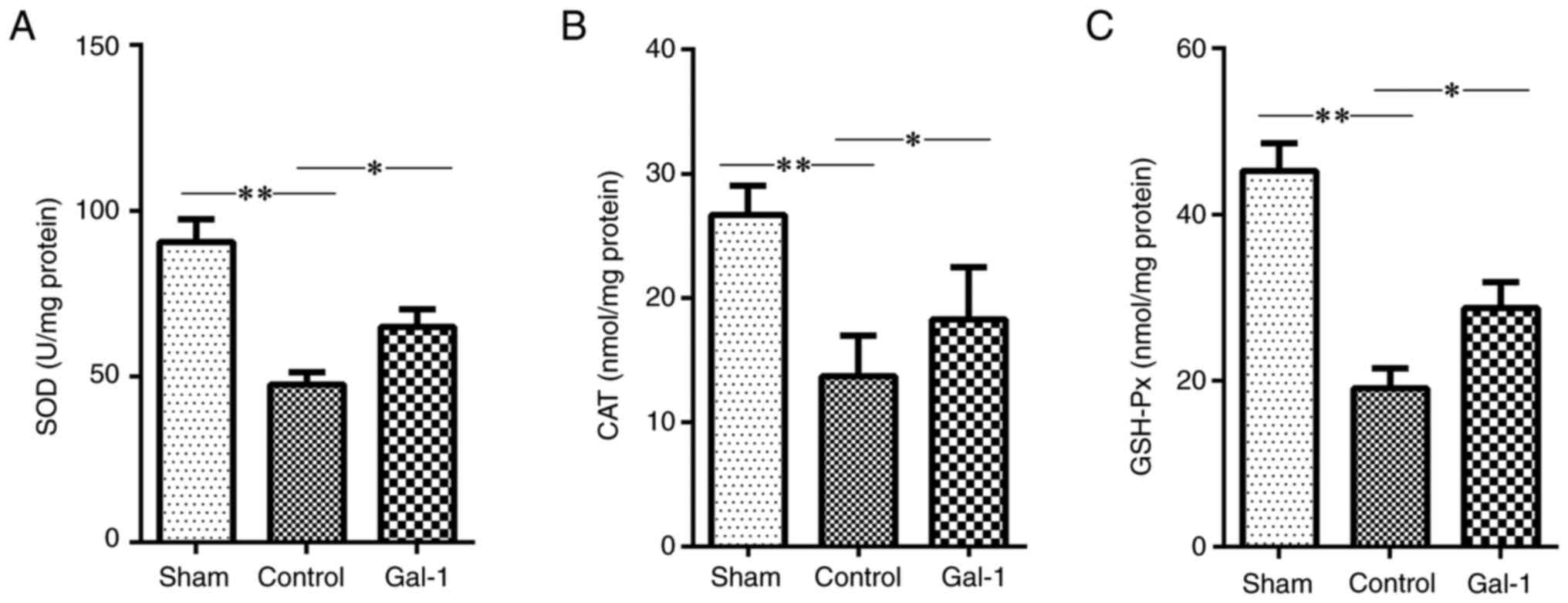 | Figure 3Levels of SOD, CAT and GSH-Px in the
ischemic hemisphere of MCAO/R mice. The mice received MCAO/R
surgery, MCAO/R surgery + galectin-1 treatment, or sham surgery
(n=10 per group). Subsequently, the levels of (A) SOD, (B) CAT and
(C) GSH-Px in the ischemic hemisphere were measured. Compared with
the sham surgery group, the levels of SOD, CAT and GSH-Px were
significantly decreased in the ischemic hemisphere of MCAO/R mice.
However, galectin-1 treatment promoted the levels of anti-oxidative
stress molecules, including SOD, CAT and GSH-Px. All of these were
increased in the galectin-1 + MCAO/R group compared with the
non-galectin-1 MCAO/R group. The data are presented as the mean ±
SD. *P<0.05 and **P<0.01 by one-way
ANOVA followed by Tukey's post hoc test. SOD, superoxide dismutase;
CAT, catalase; GSH-Px, glutathione peroxidation enzyme; MCAO/R,
middle cerebral artery occlusion-reperfusion; Gal-1, galectin-1
treatment. |
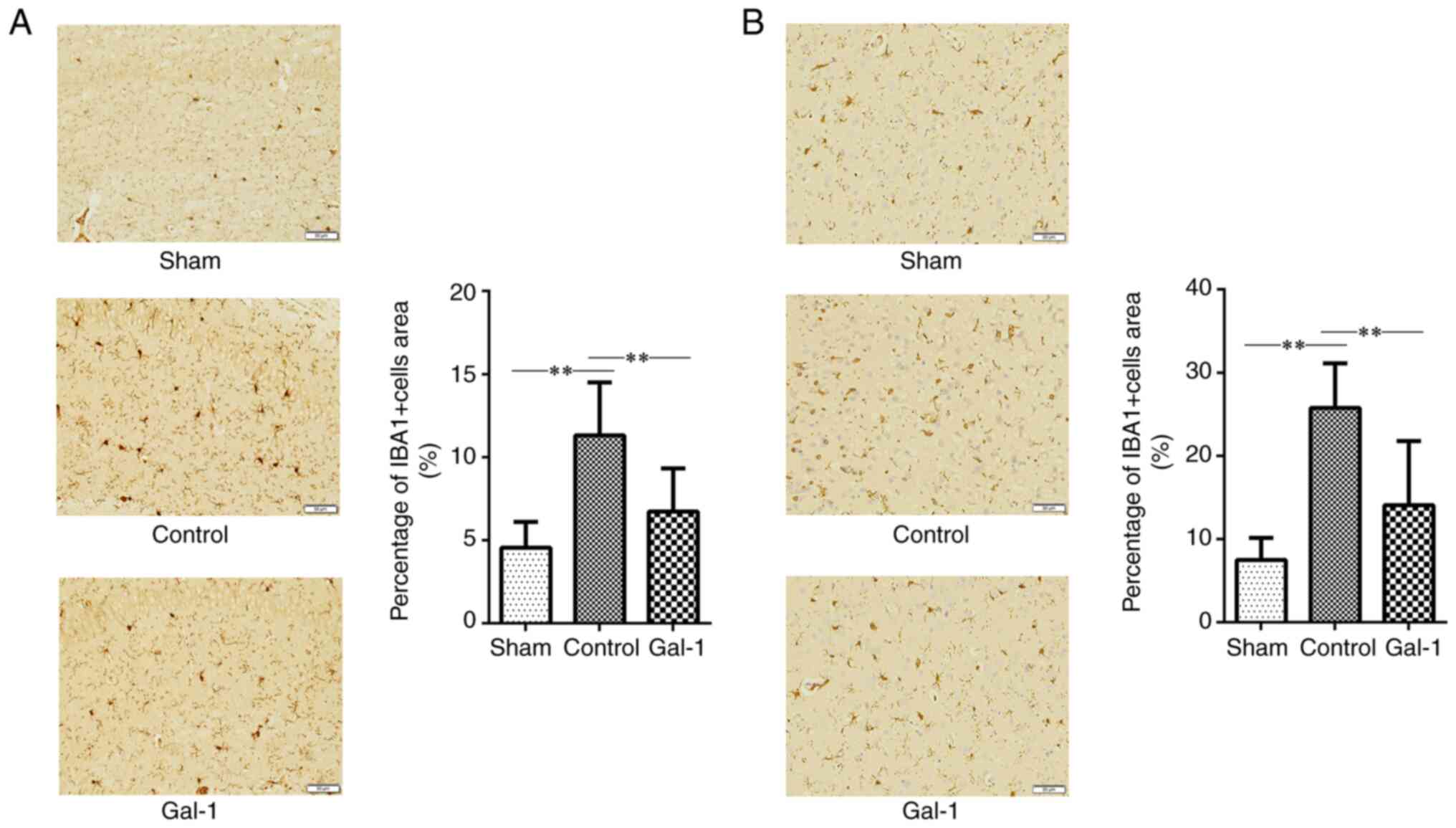
Administration of galectin-1 decreases
microglial activation and inflammatory level in the brain of MCAO/R
mice
IHC staining was performed to evaluate the
activation of microglial cells in the ischemic hemisphere of mouse
brain cortex and hippocampus. Galectin-1 treatment significantly
decreased the number of IBA-1-positive microglial cells in the
ischemic hemisphere of MCAO/R mice, compared with mice without
galectin-1 treatment (Fig. 4A and
B). In addition, cytokine levels
in the ischemic hemisphere tissues of MCAO/R mice were assessed.
The results revealed that mice treated with galectin-1 had lower
inflammatory cytokines, including IL-6, IL-1 and TNF-α, compared
with those in the brains of mice without galectin-1 treatment
(Fig. 5A-C).
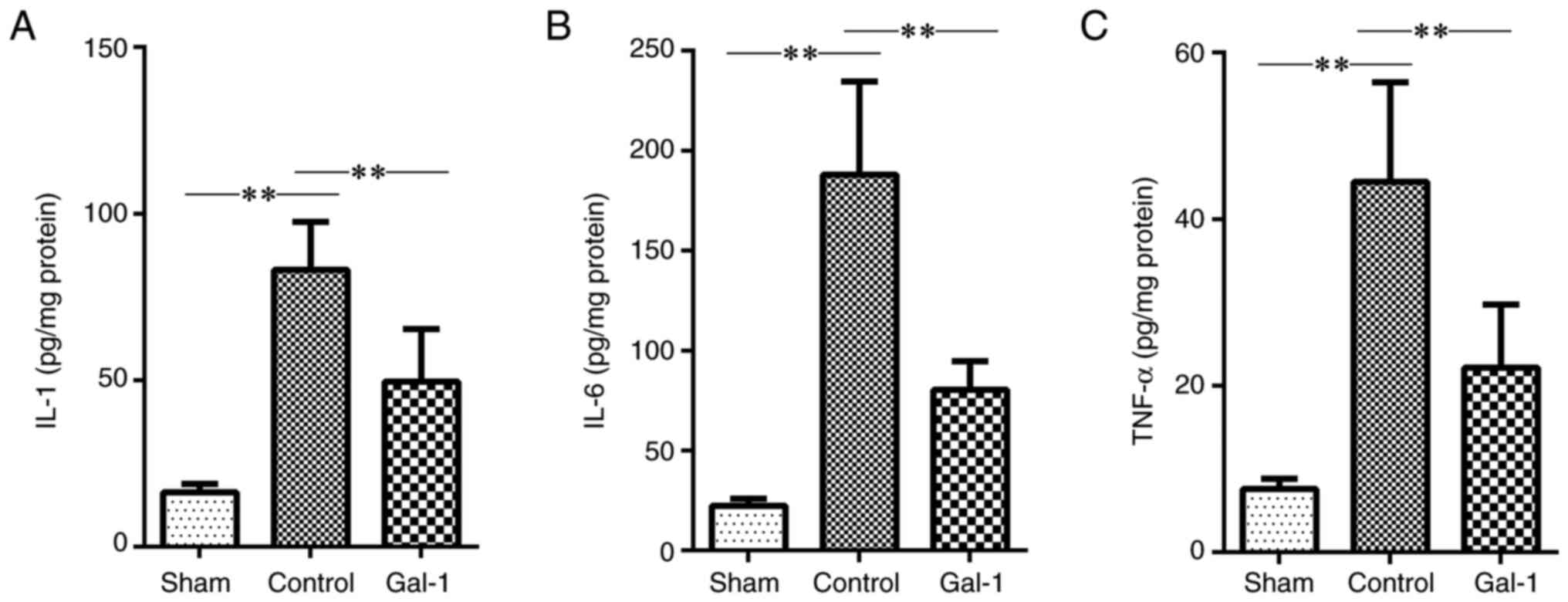 | Figure 5Inflammatory cytokine levels in the
brains of MCAO/R mice. The mice received MCAO/R surgery, MCAO/R
surgery + galectin-1 treatment, or sham surgery (n=10 per group).
Thereafter, ischemic hemisphere tissues were collected. The levels
of inflammatory cytokines (IL-1, IL-6 and TNF-α) were examined by
ELISA. (A-C) Compared with the sham surgery group, the levels of
IL-1, IL-6 and TNF-α were all significantly increased in the
ischemic hemisphere of MCAO/R mice. However, treatment with
galectin-1 decreased the levels of IL-1, IL-6 and TNF-α, compared
with the non-galectin-1 MCAO/R group. Data are expressed as the
mean ± SD. **P<0.01 by one-way ANOVA followed by
Tukey's post hoc test. MCAO/R, middle cerebral artery
occlusion-reperfusion; IL, interleukin; TNF-α, tumor necrosis
factor alpha; Gal-1, galectin-1 treatment. |
Discussion
In the present study, the neuroprotective effect of
galectin-1 on MCAO/R mouse model was investigated and it was
demonstrated that galectin-1 treatment improved cerebral I/R
outcomes, as evidenced by the lower neurological deficit and lower
infarct volumes.
Oxidative stress is closely associated with cerebral
I/R injury. When an ischemic event occurs, oxygen and nutrient
supply to brain tissues is halted, leading to energy depletion and
cell death. However, paradoxically, restoration of blood flow,
termed reperfusion, could exacerbate injury due to the sudden burst
of ROS. ROS are a group of small molecules that include free
radicals and peroxides. including stable oxidants, such as
H2O2, and unstable free radicals, such as
superoxide anion, nitric oxide, hydroxyl moiety and hypochlorite.
Under normal condition, intracellular ROS can be effectively
eliminated by the combined action of the antioxidant systems,
including SOD, CAT, GSH-Px and other endogenous antioxidants,
providing a repair mechanism for oxidized membrane components
(27). However, excess ROS
production can overwhelm the effect of antioxidant molecules,
leading to protein denaturation, lipid peroxidation and DNA
damage.
Moreover, ROS production induced by I/R may
exacerbate the inflammatory response following transient focal
ischemia. Nishi et al (28)
demonstrated that reducing ROS production by enhancing the
expression of SOD-1 in mice can decrease levels of pro-inflammatory
cytokines, including TNF-α, IL-1, IL-6, monocyte chemoattractant
protein-1 and macrophage inflammatory protein-1. Similarly, Bowler
et al (29) found that the
administration of the antioxidant AEOL 10150 attenuated
pro-inflammatory genes and modulated the immune response after
transient focal ischemia in mice. Furthermore, ROS are implicated
in the activation of immune cells and endothelial cell functions
through the activation of oxidative stress-sensitive nuclear
transcriptional factors, such as NF-κB (30). Oxidative stress may also mobilize
pools of preformed adhesion molecules in leukocytes and endothelial
cells (31). The activation of
microglia has been identified to parallel the induction of cellular
apoptosis and correlates with the severity of the ischemic insult
(32), suggesting a significant
role for inflammation in the progression of cerebral ischemic
injury. The second critical process involves the mobilization and
infiltration of inflammatory cells, such as leukocytes, from the
periphery and the activation of the endothelium. This leads to
excessive ROS production, causing oxidative damage to endothelial
cells and tissues (33-35).
In the present study, ROS production in the brain
tissues of MCAO/R mice was examined and the ROS induction by MCAO/R
was observed. The excess ROS production could affect
neurofunctional recovery from MCAO/R. Hsieh et al (36) demonstrated that urine 8-OHdG levels
were negatively correlated with functional outcomes after a stroke
and significantly decreased after rehabilitation. By contrast,
antioxidant molecules play a critical role in cell defense against
the toxic effects of oxygen radicals, reducing superoxide anions to
hydrogen peroxide. A short-term increase in ROS could elevate the
level of antioxidant molecules as an adaptation and defense
response against ROS production. However, long-term excess
production of ROS could exhaust the antioxidant system, leading to
a decrease in antioxidant levels. This is especially true in the
neuronal system, which is more sensitive to oxygen deprivation than
other organs. Consistent with this, the present study observed a
decrease in antioxidant molecules in ischemic brain tissues.
Therefore, antioxidants, compounds that neutralize ROS, and
strategies targeting the reduction of ROS generation have been
investigated as potential therapies for I/R injury in recent years.
In the present study, treatment with galectin-1 increased the
levels of antioxidant molecules in the ischemic hemisphere. These
molecules then scavenged ROS and other free radicals, including SOD
and CAT, removing hydrogen and lipid peroxides. This process
ultimately prevented the oxidation of biomolecules, thereby
reducing oxidative stress in the brains of mice with reperfusion
injury. Consequently, galectin-1 treatment improved the outcome of
ischemia in MCAO/R mice.
Previous studies have investigated the
anti-oxidative stress effect of galectin-1 in various other
pathological conditions (37). Liu
et al (14) reported that
galectin-1 inhibited the production of ROS in neuronal cells
induced by 1-methyl-4-phenyl pyridine ion. Rodrigues et al
(38) found that galectin-1 could
regulate the production of ROS in neutrophils. Moreover,
Arda-Pirincci et al (39)
reported that galectin-1 decreased the level of ROS formation in
livers of mice. Ou et al (40) found that it protected the
myocardial tissue after I/R, while Huang et al (41) suggested that galectin-1 ameliorated
acute lung injury induced by lipopolysaccharide in mice. Moreover,
galectin-1 has been revealed to protect the liver (19) and kidney (18) from I/R injury. Thus galectin-1
administration could attenuate the cell injury induced by oxidative
stress. Researchers have identified various potential pathways
involving galectin-1 in the regulation of oxidative stress. In a
recent study by the authors, it was found that galectin-1 can
promote Nrf-1 activity to increase the production of anti-ROS
molecules (14), while Huang et
al (41) demonstrated that
galectin-1 can target the AMPK-Nrf2 pathway in mice to inhibit the
level of oxidative stress. Li et al (42) discovered that galectin-1 had an
effect on NF-κB activation through the regulation of the MAPK
pathway.
Galectin-1 is expressed in immune cells. Aalinkeel
et al (43) identified that
galectin-1 can suppress the inflammation response of microglial
cells. Castillo-González et al (44) found that deficient of galectin-1
resulted in increased infiltration of CD8+ T cells and
neutrophils in the skin of mice. Thus galectin-1 can attenuate the
activation of immune cells, regulate the oxidative stress and
inflammation, and eventually ameliorate the I/R injury. In the
present study, it was observed that administration of galectin-1
alleviated the infiltration of microglial cells in the ischemic
hemisphere of MCAO/R mice. Furthermore, it was found that
galectin-1 can inhibit the generation of pro-inflammatory cytokines
in pathology of I/R injury. Similarly, Ye et al (19) suggested that administration of
galectin-1 significantly reduced proinflammatory cytokines
including TNF-α, IL-6, IL-1β, IL-12, IFN-γ and IL-17 in mice with
hepatic ischemia reperfusion injury. Meanwhile, Ou et al
(40) suggested that
administration of galectin-1 reduced the proinflammatory cytokines,
such as IL-6 and IL-1β in myocardial tissue of rats with myocardial
I/R injury. Moreover, galectin-1 can suppress the expression of
pro-inflammatory cytokines, further contributing to its
anti-oxidative stress effects through inhibition of the pathway of
NF-κB (45). Besides, galectin-1
has been found to promote angiogenesis (46) and neurogenesis (47), which are crucial for the recovery
of brain function after ischemic stroke. It also inhibits
apoptosis, further contributing to its neuroprotective effects
(45).
In addition, the data of the present study indicated
that galectin-1 treatment resulted in limited improvement in the
rotarod test; it was proposed that this may be due to the inherent
limitations of the rotarod test itself. While the rotarod test is a
widely accepted method in neuroscience research for assessing motor
coordination and balance in mice, it has certain drawbacks.
Specifically, the test can be stressful and fatiguing for the
animals, potentially affecting their performance in ways that are
not directly related to their motor coordination abilities.
Furthermore, the rotarod test primarily evaluates gross motor
coordination. As such, it may not be sufficiently sensitive to
detect subtle changes in fine motor skills, which are also known to
be affected in ischemic conditions. This limitation could account
for the observed minimal improvements in mice treated with
galectin-1, as the test may not fully capture the nuances of their
motor skill enhancements.
Therefore, galectin-1 might represent a promising
strategy in cerebral I/R injury, although to the best of the
authors' knowledge, the data on galectin-1 treatment is still
preliminary. Further research is needed to validate the outcomes of
anti-oxidative stress and anti-inflammation treatments.
In conclusion, the present study demonstrated that
galectin-1 can alleviate oxidative stress in the brains of the
MCAO/R mouse model. Moreover, galectin-1 treatment improved
outcomes in cerebral I/R, suggesting that it might be a promising
therapeutic strategy for stroke.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ, XM and XL designed the study. JZ, RZ and FH
acquired the data. JZ, MW and YW analyzed and interpreted the data.
JZ, XM and XL wrote, reviewed and edited the manuscript. JZ and XL
confirm the authenticity of all the raw data. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
All the animal-related experiments were approved
(approval no. 2022023) by the Animal Care and Management Committee
of Beijing Geriatric Hospital (Beijing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hasan TF, Hasan H and Kelley RE: Overview
of acute ischemic stroke evaluation and management. Biomedicines.
9(1486)2021.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Pan J, Konstas AA, Bateman B, Ortolano GA
and Pile-Spellman J: Reperfusion injury following cerebral
ischemia: Pathophysiology, MR imaging, and potential therapies.
Neuroradiology. 49:93–102. 2007.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Trendelenburg G: Molecular regulation of
cell fate in cerebral ischemia: Role of the inflammasome and
connected pathways. J Cereb Blood Flow Metab. 34:1857–1867.
2014.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Orellana-Urzúa S, Rojas I, Libano L and
Rodrigo R: Pathophysiology of ischemic stroke: Role of oxidative
stress. Curr Pharm Des. 26:4246–4260. 2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Domínguez C, Delgado P, Vilches A,
Martín-Gallán P, Ribó M, Santamarina E, Molina C, Corbeto N,
Rodríguez-Sureda V, Rosell A, et al: Oxidative stress after
thrombolysis-induced reperfusion in human stroke. Stroke.
41:653–660. 2010.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Zhou T, Prather ER, Garrison DE and Zuo L:
Interplay between ROS and antioxidants during ischemia-reperfusion
injuries in cardiac and skeletal muscle. Int J Mol Sci.
19(417)2018.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Li P, Stetler RA, Leak RK, Shi Y, Li Y, Yu
W, Bennett MVL and Chen J: Oxidative stress and DNA damage after
cerebral ischemia: Potential therapeutic targets to repair the
genome and improve stroke recovery. Neuropharmacology. 134:208–217.
2018.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Yang Q, Huang Q, Hu Z and Tang X:
Potential Neuroprotective treatment of stroke: Targeting
excitotoxicity, oxidative stress, and inflammation. Front Neurosci.
13(1036)2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Li W and Yang S: Targeting oxidative
stress for the treatment of ischemic stroke: Upstream and
downstream therapeutic strategies. Brain Circ. 2:153–163.
2016.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Shirley R, Ord EN and Work LM: Oxidative
stress and the use of antioxidants in stroke. Antioxidants (Basel).
3:472–501. 2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Olie RA, Hall J, Natt F, Stahel RA and
Zangemeister-Wittke U: Analysis of ribosyl-modified, mixed backbone
analogs of a bcl-2/bcl-xL antisense oligonucleotide. Biochim
Biophys Acta. 1576:101–109. 2002.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Dimitrijevic OB, Stamatovic SM, Keep RF
and Andjelkovic AV: Absence of the chemokine receptor CCR2 protects
against cerebral ischemia/reperfusion injury in mice. Stroke.
38:1345–1353. 2007.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Sun R, Peng M, Xu P, Huang F, Xie Y, Li J,
Hong Y, Guo H, Liu Q and Zhu W: Low-density lipoprotein receptor
(LDLR) regulates NLRP3-mediated neuronal pyroptosis following
cerebral ischemia/reperfusion injury. J Neuroinflammation.
17(330)2020.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Liu HB, Li QY, Zhang XD, Shi Y and Li JY:
The neuroprotective effects of galectin-1 on Parkinson's disease
via regulation of Nrf2 expression. Eur Rev Med Pharmacol Sci.
26:623–636. 2022.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Vallecillo-Zúniga ML, Rathgeber MF,
Poulson PD, Hayes S, Luddington JS, Gill HN, Teynor M, Kartchner
BC, Valdoz J, Stowell C, et al: Treatment with galectin-1 improves
myogenic potential and membrane repair in dysferlin-deficient
models. PLoS One. 15(e0238441)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Cheng YH, Jiang YF, Qin C, Shang K, Yuan
Y, Wei XJ, Xu Z, Luo X, Wang W and Qu WS: Galectin-1 contributes to
vascular remodeling and blood flow recovery after cerebral ischemia
in mice. Transl Stroke Res. 13:160–170. 2022.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Arda-Pirincci P and Aykol-Celik G:
Galectin-1 reduces the severity of dextran sulfate sodium
(DSS)-induced ulcerative colitis by suppressing inflammatory and
oxidative stress response. Bosn J Basic Med Sci. 20:319–328.
2020.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Carlos CP, Silva AA, Gil CD and Oliani SM:
Pharmacological treatment with galectin-1 protects against renal
ischaemia-reperfusion injury. Sci Rep. 8(9568)2018.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Ye Y, Wang W, Zhang W, Peng Y, Liu Y, Yu
S, Chen Q, Geng L, Zhou L, Xie H, et al: Galectin-1 attenuates
hepatic ischemia reperfusion injury in mice. Int Immunopharmacol.
77(105997)2019.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Xia Q, Li X, Zhou H, Zheng L and Shi J:
S100A11 protects against neuronal cell apoptosis induced by
cerebral ischemia via inhibiting the nuclear translocation of
annexin A1. Cell Death Dis. 9(657)2018.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Chen C, Ma Q, Jiang J, Wang T, Qiu L and
Liu A: Protective effects of nuciferine in middle cerebral artery
occlusion rats based on transcriptomics. Brain Sci.
12(572)2022.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Wang J, Lin X, Mu Z, Shen F, Zhang L, Xie
Q, Tang Y, Wang Y, Zhang Z and Yang GY: Rapamycin Increases
collateral circulation in rodent brain after focal ischemia as
detected by multiple modality dynamic imaging. Theranostics.
9:4923–4934. 2019.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Tian W, Zhu M, Zhou Y, Mao C, Zou R, Cui
Y, Li S, Zhu J and Hu C: Electroacupuncture pretreatment alleviates
cerebral ischemia-reperfusion injury by regulating mitophagy via
mTOR-ULK1/FUNDC1 axis in rats. J Stroke Cerebrovasc Dis.
31(106202)2022.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Wang C, Ma Z, Wang Z, Ming S, Ding Y, Zhou
S and Qian H: Eriodictyol attenuates MCAO-induced brain injury and
neurological deficits via reversing the autophagy dysfunction.
Front Syst Neurosci. 15(655125)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Otsuka S, Itashiki Y, Tani A, Matsuoka T,
Takada S, Matsuzaki R, Nakanishi K, Norimatsu K, Tachibe Y,
Kitazato R, et al: Effects of different remote ischemia
perconditioning methods on cerebral infarct volume and neurological
impairment in rats. Sci Rep. 13(2158)2023.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Paris L, Roussel M, Pereira B, Delbac F
and Diogon M: Disruption of oxidative balance in the gut of the
western honeybee Apis mellifera exposed to the intracellular
parasite Nosema ceranae and to the insecticide fipronil. Microb
Biotechnol. 10:1702–1717. 2017.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Bhattacharyya A, Chattopadhyay R, Mitra S
and Crowe SE: Oxidative stress: An essential factor in the
pathogenesis of gastrointestinal mucosal diseases. Physiol Rev.
94:329–354. 2014.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Nishi T, Maier CM, Hayashi T, Saito A and
Chan PH: Superoxide dismutase 1 overexpression reduces MCP-1 and
MIP-1 alpha expression after transient focal cerebral ischemia. J
Cereb Blood Flow Metab. 25:1312–1324. 2005.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Bowler RP, Sheng H, Enghild JJ, Pearlstein
RD, Warner DS and Crapo JD: A catalytic antioxidant (AEOL 10150)
attenuates expression of inflammatory genes in stroke. Free Radic
Biol Med. 33:1141–1152. 2002.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Taylor JM, Crack PJ, Gould JA, Ali U,
Hertzog PJ and Iannello RC: Akt phosphorylation and NFkappaB
activation are counterregulated under conditions of oxidative
stress. Exp Cell Res. 300:463–475. 2004.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Ochoa CD, Wu RF and Terada LS: ROS
signaling and ER stress in cardiovascular disease. Mol Aspects Med.
63:18–29. 2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Zhao SC, Ma LS, Chu ZH, Xu H, Wu WQ and
Liu F: Regulation of microglial activation in stroke. Acta
Pharmacol Sin. 38:445–458. 2017.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Wong CHY and Crack PJ: Modulation of
neuro-inflammation and vascular response by oxidative stress
following cerebral ischemia-reperfusion injury. Curr Med Chem.
15:1–14. 2008.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Fisher M: Injuries to the vascular
endothelium: Vascular wall and endothelial dysfunction. Rev Neurol
Dis. 5 (Suppl 1):S4–S11. 2008.PubMed/NCBI
|
|
35
|
Kataoka H, Kim SW and Plesnila N:
Leukocyte-endothelium interactions during permanent focal cerebral
ischemia in mice. J Cereb Blood Flow Metab. 24:668–676.
2004.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Hsieh YW, Lin KC, Korivi M, Lee TH, Wu CY
and Wu KY: The reliability and predictive ability of a biomarker of
oxidative DNA damage on functional outcomes after stroke
rehabilitation. Int J Mol Sci. 15:6504–6516. 2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Liu FT and Rabinovich GA: Galectins:
Regulators of acute and chronic inflammation. Ann NY Acad Sci.
1183:158–182. 2010.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Rodrigues LC, Kabeya LM, Azzolini AECS,
Cerri DG, Stowell SR, Cummings RD, Lucisano-Valim YM and
Dias-Baruffi M: Galectin-1 modulation of neutrophil reactive oxygen
species production depends on the cell activation state. Mol
Immunol. 116:80–89. 2019.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Arda-Pirincci P, Sacan O, Ozal-Coskun C,
Aykol-Celik G, Karabulut-Bulan O, Yanardag R and Bolkent S:
Galectin-1 exhibits a protective effect against hepatotoxicity
induced by dextran sulfate sodium in mice. Hum Exp Toxicol.
39:423–432. 2020.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Ou D, Ni D, Li R, Jiang X, Chen X and Li
H: Galectin-1 alleviates myocardial ischemia-reperfusion injury by
reducing the inflammation and apoptosis of cardiomyocytes. Exp Ther
Med. 23(143)2022.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Huang XT, Liu W, Zhou Y, Sun M, Yang HH,
Zhang CY and Tang SY: Galectin-1 ameliorates
lipopolysaccharide-induced acute lung injury via AMPK-Nrf2 pathway
in mice. Free Radic Biol Med. 146:222–233. 2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Li Y, Chen N, Wu C, Lu Y, Gao G, Duan C,
Yang H and Lu L: Galectin-1 attenuates neurodegeneration in
Parkinson's disease model by modulating microglial MAPK/IκB/NFκB
axis through its carbohydrate-recognition domain. Brain Behav
Immun. 83:214–225. 2020.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Aalinkeel R, Mangum CS, Abou-Jaoude E,
Reynolds JL, Liu M, Sundquist K, Parikh NU, Chaves LD, Mammen MJ,
Schwartz SA and Mahajan SD: Galectin-1 reduces neuroinflammation
via modulation of nitric oxide-arginase signaling in HIV-1
transfected microglia: A gold nanoparticle-galectin-1 ‘nanoplex’ a
possible neurotherapeutic? J Neuroimmune Pharmacol. 12:133–151.
2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Castillo-González R, Cibrian D,
Fernandez-Gallego N, Ramírez-Huesca M, Saiz ML, Navarro MN, Fresno
M, de la Fuente H and Sánchez-Madrid F: Galectin-1 expression in
CD8+ T lymphocytes controls inflammation in contact
hypersensitivity. J Invest Dermatol. 141:1522–1532.e3.
2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Starossom SC, Mascanfroni ID, Imitola J,
Cao L, Raddassi K, Hernandez SF, Bassil R, Croci DO, Cerliani JP,
Delacour D, et al: Galectin-1 deactivates classically activated
microglia and protects from inflammation-induced neurodegeneration.
Immunity. 37:249–263. 2012.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Li X, Shi W, Wei G, Lv J, Wang D, Xing B,
Zhou J, Zhao J and Sun H: Galectin-1 promotes angiogenesis and
chondrogenesis during antler regeneration. Cell Mol Biol Lett.
28(40)2023.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Ishibashi S, Kuroiwa T, Sakaguchi M, Sun
L, Kadoya T, Okano H and Mizusawa H: Galectin-1 regulates
neurogenesis in the subventricular zone and promotes functional
recovery after stroke. Exp Neurol. 207:302–313. 2007.PubMed/NCBI View Article : Google Scholar
|