1. Introduction
Digestive tumors are a series of malignancies
affecting the gastrointestinal tract and associated tissues,
including the esophagus, stomach, liver, pancreas, colon, rectum
and biliary ducts (1). These
malignancies, especially colorectal, gastric and hepatocellular
cancers (HCC), account for a substantial proportion of
cancer-related mortality worldwide. According to the Global Cancer
Statistics, ~4.7 million new cases and >3.2 million reported
mortalities were associated with digestive tumors in 2022. Among
them, the most frequent types of digestive malignancies were
colorectal cancer (CRC), which accounted for 1.9 million new cases
with >900,000 mortalities annually, and gastric cancer, which
accounted for ~1.1 million new cases and >800,000 mortalities
annually (2). Gastric cancer is
more frequent in Eastern Asian countries, such as Japan, South
Korea and China (3). Genetic
predisposition, environmental risk factors, infections, metabolic
changes and microbial factors are markedly involved in the
pathogenesis of these malignancies (1). Over the years, researchers have
focused on gut-microbial interactions, particularly regarding how
dysbiosis influences host metabolic alterations and participates in
tissue microenvironment changes, tumor development, progression and
response to treatment (4-7).
Within this framework, manipulation of the gut microbial metabolism
axis represents a promising avenue for developing innovative
therapeutic strategies for cancer management.
The human gastrointestinal tract harbors a complex
and diverse microbial ecosystem composed of bacteria, archaea,
fungi and viruses, known as the gut microbiota (8). This ecosystem has emerged as a
fundamental regulator of human physiology in health and disease,
particularly affecting metabolism, immunity and tissue homeostasis
(9). It serves as a repository for
numerous microorganisms that encode a complex pool of enzymes and
facilitate the digestion of complex dietary components of the gut.
In addition, gut microbiota can orchestrate the host metabolic
functions, including fermentation of the dietary fibers, modulation
of protein and amino acid metabolism, vitamin synthesis and bile
acid transformation (10).
Disruption of the equilibrium between the host and
microbiota, known as dysbiosis, is characterized by a decreased
proportion of beneficial commensals and overrepresentation of
pathogenic or proinflammatory species (6,7).
This imbalance has been associated with a range of pathological
consequences, including metabolic dysregulation, chronic
inflammatory diseases, digestive tumors and even neurodegenerative
complications (7). The gut
microbiota-host metabolism axis produces various bioactive
metabolites, such as amino acid derivatives, short-chain fatty
acids (SCFAs), secondary bile acids and other microbial products.
These compounds serve notable roles in immune cell activation,
inflammation, cellular signaling and epigenetic modification
(8,11). These microbial metabolites can
potentially influence the tumor microenvironment (TME) by
activating or suppressing tumor immune cells, checkpoint inhibitors
and contributing to tumor immune escape, therapeutic response or
resistance (12-14).
Due to the rapid expansion of research in this
field, a comprehensive focus on how the gut microbiota-host
interplay affects metabolic changes and their therapeutic
implications for digestive tumors is essential. Through a
multidisciplinary search approach, the present review aimed to
integrate current knowledge, summarizing evidence on compositional
changes in the gut microbiota across various digestive malignancies
and providing detailed mechanistic insights into microbiota-based
applications in oncology and targeted therapeutic strategies. The
challenges in translating microbiome research into clinical
practice are also addressed and emerging microbial-targeted
interventions are discussed, focusing on the prospects of
microbiome-based personalized medicine.
2. Search strategy, inclusion and exclusion
criteria
The present narrative review was performed based on
targeted literature searches spanning January 2000 to September
2025 across major online medical academic databases, including
MEDLINE/PubMed (https://www.nlm.nih.gov/medline/medline_home.html;
https://pubmed.ncbi.nlm.nih.gov/),
Scopus (https://www.scopus.com/), and Web of
Science (https://www.webofscience.com/), supplemented with
manual screening of references from the included studies. The
primary search syntax was combined using the Boolean operators
(AND, OR and NOT), and included the following terms as keywords:
‘Gut microbiota’, ‘microbiome’, ‘digestive tumors’,
‘gastrointestinal cancer’, ‘hepatocellular carcinoma’, ‘pancreatic
cancer’, ‘colorectal cancer’, ‘microbial metabolites’, ‘short-chain
fatty acids’, ‘bile acids’, ‘polyamines’ and ‘tumor
microenvironment’.
The included studies were English-written original
research articles, review articles and meta-analyses in the field
of the study. Studies were excluded if they addressed non-digestive
malignancies or lacked substantial information on
microbiota-metabolism related mechanisms. Due to the narrative
design of the present review, formal systematic review procedures
(such as protocol registration or risk-of-bias scoring) were not
applied. Instead, emphasis was placed on mechanistic and clinically
relevant studies to provide an integrative prospective.
3. Gut microbiota in digestive tumors:
Current understanding
Distinct microbial signatures have been observed in
various digestive tumors, and are often associated with disease
stage, progression and treatment outcomes (15-17).
These patterns suggest that the gut microbiota is not only
effective in digestive biology but also in pathogenic status,
serving as a driver, modulator and notable therapeutic target for
personalized medicine (15,18).
The following section details the specific composition of
microbiota and their alterations in dysbiosis observed across major
digestive malignancies.
4. Composition and dysbiosis of the gut
microbiota in digestive tumors
Understanding the alterations in microbial
composition across different digestive tumors can provide insights
into cancer etiology and offer novel biomarkers and therapeutic
targets (Fig. 1). Common features
of dysbiosis in digestive tumors, such as pancreatic, liver,
gastric and CRCs, substantially contribute to tumorigenesis through
mechanisms such as genotoxicity, metabolic reprogramming, chronic
inflammation and immune evasion (19,20).
CRC
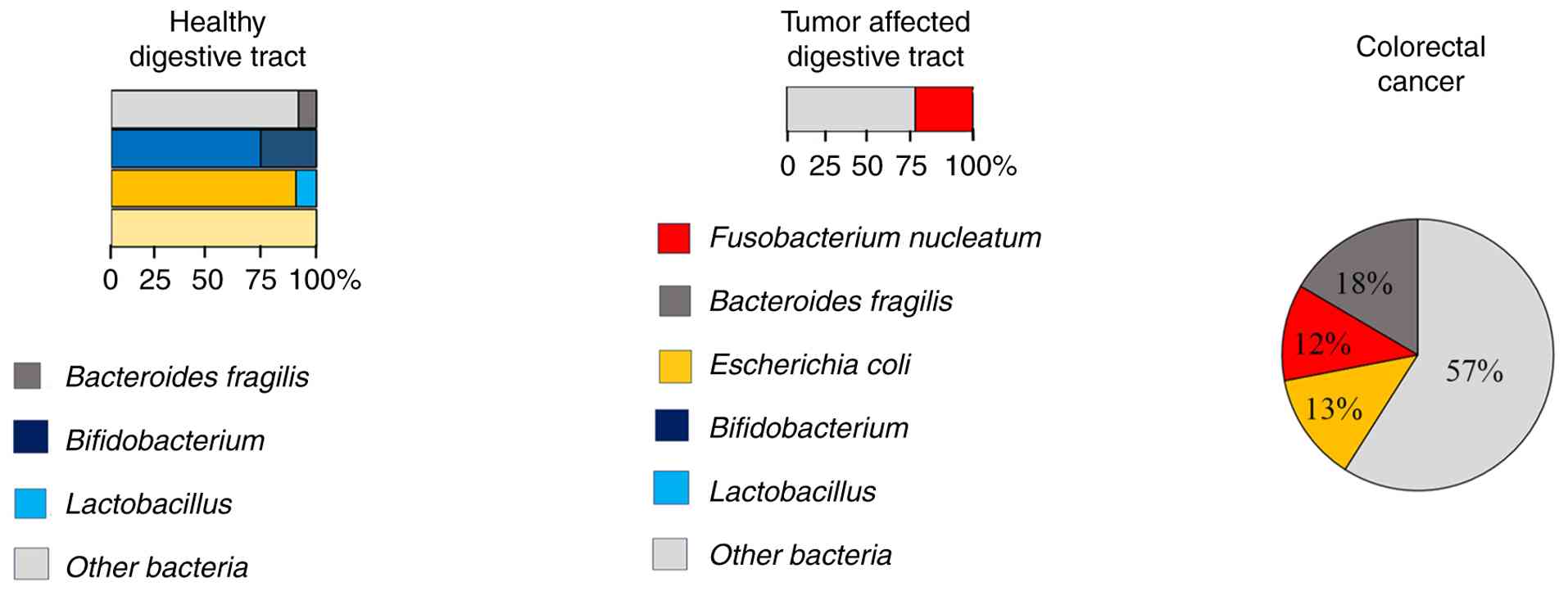
The gut bacterial composition of colon tissue in
healthy individuals is dominated by Firmicutes and Bacteroidetes,
with a lower proportion of Proteobacteria, Fusobacteria and
Acinetobacter (21,22). Dysbiosis in CRC is marked by the
abundance of pathogenic species in the colon tissue, including
Bacteroides fragilis, Fusobacterium nucleatum
(F. nucleatum) and some strains of Escherichia
coli. (E. coli), such as phylogroups B2 and A (23-26).
These bacteria contribute to tumorigenesis by inducing chronic
inflammation, promoting DNA damage and suppressing the antitumor
immune responses in cancerous tissues. Additionally, the
overrepresentation of species, such as Peptostreptococcus
and Parvimonas, is frequently reported in the advanced
stages of CRC (18,27,28).
Furthermore, depletion of beneficial commensals, such as
Faecalibacterium prausnitzii (F. prausnitzii),
Lachnospiraceae and Akkermansia muciniphila (A.
muciniphila), serving as the key regulators of intestinal
barrier function, was associated with decreased production of
anti-inflammatory SCFAs (26,29,30).
Decreased production of SCFAs, such as butyrate, impedes the
inhibitory effects of histone deacetylase (HDAC), thereby improving
survival and increasing the proliferation of CRC cells through the
Warburg effect (31-33).
In addition, carcinogenic metabolites, such as secondary bile
acids, N-nitroso compounds and hydrogen sulfide, induce genotoxic
stress (caused by production of colibactin by pathogenic strains of
E. coli that results in DNA double-strand breakage and
somatic mutations) and epithelial transformation (34-37).
Gastric cancer
Compared with the intestine, the gastric microbiota
has less diversity, owing to the acidic environment of the gastric
tissue (38). Nevertheless,
distinct microbial shifts have been reported in terms of gastric
complications. Helicobacter pylori (H. pylori)
induces chronic gastritis, epithelial damage and carcinogenesis in
the gastric tissues (4,39). The bacterial load of H.
pylori is markedly increased in >70% of patients with
gastric adenocarcinoma (40).
Beyond H. pylori, the other changes in the gastric
microbiota, including increased Lactobacillus reuteri
(25) and Streptococcus,
have been identified in gastric atrophy and intestinal metaplasia
(41).
In gastric cancer, H. pylori induces
oncogenic signaling pathways through its virulence factors,
including cytotoxin-associated gene A (CagA) and vacuolating
cytotoxin A gene. CagA activates nuclear factor κB (NF-κB) and
STAT3, resulting in the overexpression of IL-8/cyclooxygenase-2 and
gastric epithelial apoptosis (35,42,43).
H. pylori also increases the production of reactive oxygen
species, causes mutations in the tumor protein p53 gene and
chromosomal instability (44). By
contrast, decreased proportions of beneficial bacteria such as
Lactobacillus gasseri and Bifidobacterium have been
observed in patients with gastric cancer (45,46).
Lactobacillus gasseri is needed for lactic acid production,
gastric mucosa protection and maintenance of gastric acidity.
Gastric bacteria dysbiosis results in overgrowth of opportunistic
bacteria such as Streptococcus, which contributes to
alteration in arginine metabolism and increased mucosal
inflammation, activation of oncogenic signaling pathways, and
increase in the production of N-nitroso compounds in the gastric
tissue, thereby promoting DNA damage and mutagenesis in gastric
epithelial cells (4,47,48).
Pancreatic cancer
Contrary to traditional beliefs, the pancreas is not
a completely sterile organ (49).
Previous studies have demonstrated that pancreatic ductal
adenocarcinoma is influenced by microbial translocation from the
oval cavity or gut, indicating a notable link between dysbiosis and
pancreatic tumors (50-52).
Genomic studies have demonstrated the presence of Proteobacteria,
Acinetobacter and Enterobacteriaceae within the pancreatic
tumor environment, where they may contribute to immune responses
modulation and chemoresistance (20,53-55).
In addition, the decreased abundance of beneficial bacteria,
particularly Bacteroides and Clostridium in the gut
reduces the production of SCFAs which may support cancer
progression by disturbing mucosal defense and providing an
inflammatory microbial environment (20). Periodontal pathogens, such as
Porphyromonas gingivalis and Aggregatibacter
actinomycetemcomitans, have also been identified as risk
factors for pancreatic cancer, possibly through the oral-gut axis,
systemic inflammation and immune modulation (50). Pancreatic dysbiosis may contribute
to the regulation of bile acid metabolism, tumorigenic
microenvironment development and tumor progression (20).
HCC
The development of HCC is notably associated with
chronic liver diseases, such as viral hepatitis and non-alcoholic
steatohepatitis (1); both
conditions are affected by disturbances in the gut microbiota-liver
axis. Dysbiosis in HCC is characterized by increased abundance of
pathogenic bacteria (Enterococcus faecalis, Clostridium
scindens and Escherichia species) and depletion of
beneficial bacteria (Lactobacillus and Bifidobacterium
adolescentis), thus reducing the production of SCFAs and
impairing epithelial repair and anti-inflammatory signaling
(19). Pathogenic bacteria in HCC
produce endotoxins, such as lipopolysaccharides (LPS) and other
microbial products, that infiltrate the gut, activate hepatic
Kupffer cells and promote chronic inflammation and fibrosis in the
liver. For instance, Clostridium scindens exerts bile acid
toxicity by converting primary bile acids to deoxycholic acid and
activating hepatic stellate cells through farnesoid X receptor
(FXR)/G protein-coupled bile acid receptor (TGR5) signaling
(56,57).
Esophageal cancer
The esophageal microbiota is markedly influenced by
diet, reflux and oral hygiene. Dysbiotic alterations in esophageal
adenocarcinoma include an increase in the abundance of
Veillonella, Campylobacter and Proteobacteria
(1). These bacteria can produce
proinflammatory molecules and LPS in the esophageal tissue
(24). Continuous exposure of
esophageal cells to LPS results in the activation of NF-κB and
STAT3 signaling pathways and increasing the production of TNF-α and
IL-6, which pro-tumorigenesis events, supporting cell survival,
proliferation, migration and invasion associated with malignant
phenotypes (58,59).
5. Key microbial metabolites in digestive
tumors
The gut microbiota produces various metabolites that
directly or indirectly influence the immune response, inflammation,
signaling pathways and cellular microenvironment in digestive
tumors. These bioactive compounds serve as signaling mediators,
epigenetic modifiers, inflammatory modulators and TME modifiers.
The major microbial metabolites of the gut microbiota in patients
with digestive tumors are presented in Table I and discussed in the present
section.
 | Table IMajor microbiota shift across
digestive tumors. |
Table I
Major microbiota shift across
digestive tumors.
| Taxon | Primary tumor
sites | Functional
category | Principal
mechanisms | Microbiota effect
on tumor biology | (Refs.) |
|---|
|
Fusobacterium nucleatum | Colorectal,
gastric, esophagus | Pathogenic
bacteria | Induce
inflammation, promotes M2-macrophage polarization | Interfere with
chemotherapeutic agents, such as 5-FU | (59,133,134) |
| Bacteroides
fragilis | Colorectal,
hepatocellular | Pathogenic
bacteria | Produce
enterotoxins, disrupts E-cadherin, activates Wnt/β-catenin
signaling and promotes inflammation | Promote epithelial
mesenchymal transition, increase genomic instability, promote tumor
progression | (135-137) |
| Escherichia
coli (colibactin-positive species) | Colorectal,
hepatocellular | Pathogenic
bacteria | Produce colibactin,
promote double stranded DNA breakage and induce KRAS mutations | Causes genotoxicity
injuries, incorporates in tumor initiation and progression | (105,106,138) |
| Helicobacter
pylori | Gastric cancer | Pathogenic
bacteria | Produce CagA and
VacA factors, activates NF-κB and STAT3 signaling and promotes
ROS-mediated DNA damage | Induce gastritis,
gastric atrophy, metaplasia and gastric adenocarcinoma | (40,44) |
|
Streptococcus | Gastric,
esophagus | Pathogenic
bacteria | Increase lactate
production, activates VEGF/MMP9 signaling and promotes
pro-inflammatory signaling | Induces
angiogenesis in tumor microenvironment, enhance tumor invasion and
metastasis | (39,48,59) |
|
Faecalibacterium
prausnitzii | Colorectal,
hepatocellular | Beneficial
commensal | Produces butyrate
and other SCFAs, inhibits histone deacetylases and maintains
epithelial barrier integrity | Tumor suppressive
milieu, reduces inflammation, improves epithelial integrity | (24,139) |
|
Akkermansia | Colorectal,
metabolic diseases | Beneficial
commensal | Maintaining mucosal
integrity | Exert
anti-inflammation and improves response to checkpoint
inhibitors. | (23,27,123) |
|
Lachnospiraceae | Colorectal,
hepatocellular | Beneficial
commensal | Produces butyrate
and other SCFAs, inhibits histone deacetylases and maintains
epithelial barrier integrity | Tumor suppressive
milieu, reduces inflammation and improves epithelial integrity | (140,141) |
|
Bifidobacterium | Gastric,
colorectal | Beneficial
commensal | Produces SCFA and
indole derivatives, promotes dendritic cell maturation | Improves antitumor
immunity, reduces colitis and associated tumorigenesis | (30,46) |
SCFAs
Beneficial commensals, such as F. prausnitzii
and Lachnospiraceae, are gut microbiota that produce SCFAs
(acetate, butyrate and propionate) through bacterial fermentation
of dietary fibers in the colon tissue (31,60).
Butyrate serves a bifunctional role in digestive tumors, acting as
an epigenetic regulator (inhibition of HDAC) and modulating cell
cycle checkpoints, NF-κB and Wnt/β-catenin pathways, which are
context-dependent (33,61). This means that in healthy
colonocytes, butyrate helps improve differentiation, increases
apoptosis and promotes anti-inflammatory signaling primarily by
inhibiting HDAC and binding to G-protein-coupled receptors (such as
free fatty acid receptor 2/3 and hydroxycarboxylic acid receptor 2)
(62-64).
However, in advanced colorectal tumors, particularly under hypoxia
which results in increased glycolytic effects (Warburg phenotype),
the uptake and oxidation of butyrate are altered as its
intracellular accumulation suppresses HDAC and arrests cellular
growth and differentiation. In addition, butyrate interacts with
immune and stromal components within the TME, thereby promoting the
differentiation of T lymphocytes and enhancing the epithelial
barrier integrity (32). Acetate
and propionate affect lipid metabolism and glucose hemostasis in
CRC cell lines and exhibit anti-inflammatory properties by
activating G-protein-coupled receptors (GPR41 and GPR43) that
regulate cytokine production and immune cell function (64-66).
In addition, propionate inhibits the activation of the NLR family
pyrin domain containing 3 inflammasome and exerts protective
effects by reducing colitis-associated CRC (67).
Bile acids
Within the gut, bile acids originate from two
distinct sources as follows: Primary bile acids are naturally
synthesized in the liver and secreted in conjugated form to the
intestine, whereas secondary bile acids, such as deoxycholic acid
(DCA) and lithocholic acid (LCA) are produced through
transformation by gut microbiota, such as Clostridium
species (37). DCA and LCA
subsequently contribute to tumor biology and metabolic
reprogramming by activating cell-surface receptors (such as FXR and
TGR5) in epithelial, immune and hepatic cells, inducing
pro-inflammatory cytokines, as well as oxidative and nitrosative
stress, thereby modulating DNA damage through the activation of
NADPH oxidase. In addition, they can activate oncogenic signaling
pathways by affecting the expression of gene involved in the
Wnt/β-catenin, NF-κB and EGFR/MAPK pathways (36). Although LCA can induce apoptosis in
some types of cancer (such as nephroblastoma, acute myeloid
leukemia, and breast cancer), it exhibits inflammatory properties
and induces epithelial injury under dysbiotic conditions (68-72).
Furthermore, LCA promotes metastasis in gastric cancer cells by
activating TGR5, modulating the EGFR/MAPK signaling pathway and
inducing cell migration (11,37,61,73,74).
Amino acid-derived metabolites
The gut microbiota can metabolize amino acids into
different compounds, such as polyamines, hydrogen sulfide
(H2S), kynurenine, indole-3-propionic acid (IPA) and
indole-3-acetic acid (IAA), which influence tumor biology (75). Kynurenine, produced by
Lactobacillus, serves as a biomarker of poor prognosis in
digestive tumors (76); it
suppresses the activation of T-lymphocytes, facilitates immune
evasion and mediates tryptophan catabolism in the gut (77). Tryptophan can be metabolized in the
gut by both kynurenine-dependent pathways and the gut
microbiota-host metabolism axis. IPA, a byproduct of tryptophan
catabolism, can impair T-cytotoxic immune function, improve
epithelial barrier function and modulate the immune response in
pancreatic cancer (78,79). In a CRC model, IPA demonstrated
anti-inflammatory effects and inhibited tumor growth (80). Bifidobacterium produces IAA,
which binding to and activating aryl hydrocarbon receptors on
immune cell surface, therefore, contributes to downregulating the
expression of interleukin-22 binding protein (IL-22BP), This
results in decreasing the intercellular IL-22, a potent oncogenic
cytokine, levels that indicates its protective effects in HCC
(81-83).
Polyamines (putrescine, spermine and spermidine) are
the other amino acid-derived metabolites, which can be produced by
Bacteroides species and are derived from ornithine and
arginine metabolism (84,85). Aberrant production of polyamines
promote tumor proliferation, angiogenesis and metastasis (85). In gastric cancer, elevated
polyamine levels are associated with angiogenesis through increased
production of hypoxia inducible factor 1-α and VEGF (86). Increased polyamine levels have also
been associated with the proliferation of CRC cell lines (87). Sulfate-reducing bacteria, such as
Desulfovibrio and Bilophila, metabolize dietary
cysteine and produce H2S, which has dual functions under
different conditions. At low, physiological concentrations,
H2S exerts a protective role by supporting mucosal
barrier integrity, redox hemostasis and mitigating excessive
inflammation in the gut (88,89),
thereby contributing to epithelial defense and immune balance. In
addition. H2S serves as a signaling molecule that
supports intestinal homeostasis, prevents chronic injury
development and preserves epithelial cell survival and repair. By
contrast, increasing the abundance of sulfidogenic bacteria during
dysbiosis results in locally increased levels of H2S;
therefore, a disequilibrium between H2S levels and
detoxification pathways occurs in colonocytes and shifts its
effects toward promoting tumor development (90,91).
However, in dysbiosis and at high concentrations, H2S
modulates DNA hypermethylation (inhibits DNA methyltransferase),
silences the secreted frizzled related protein 2 tumor suppressor
gene and activates NF-κB signaling (92).
Other microbial metabolites
The gut microbiota can produce other metabolites,
including phenolic compounds (such as flavonoids), lactate and
ammonia, which are functional in reshaping the TME and immune
responses (93). LPS is a
structural component of the outer membrane of gram-negative
bacteria (45); LPS is a potent
bacterial endotoxin that activates the innate immune system via
Toll-like receptor 4 (TLR4) signaling, resulting in chronic
inflammation by activating NF-κB signaling and increasing the
production of tumor-promoting cytokines, such as IL-6 and IL-8, in
gastric and pancreatic cancers (94). Increased LPS levels have also been
identified in hepatocellular and gastric cancers (94).
6. Metabolic interaction between gut
microbiota and digestive tumors
Beyond specific metabolites, the gut microbiota is
involved in numerous metabolic interactions that contribute to the
pathogenesis of digestive tumors. The gut microbiota is a key
regulator of host physiology, particularly in digestive tumor
biology, immune responses, cellular signaling and TME remodeling
(7,26,79).
A comprehensive list of microbial alterations in digestive tumors
and their corresponding mechanisms is presented in Table II.
| Table IIKey microbial-derived metabolites and
their mechanisms in digestive tumors. |
Table II
Key microbial-derived metabolites and
their mechanisms in digestive tumors.
| Metabolite | Source
microbes | Mechanisms of
action | Clinical
implications | (Refs.) |
|---|
| SCFAs-butyrate,
acetate, propionate |
Faecalibacterium
prausnitzii, Lachnospiraceae, Roseburia | Inhibit histone
deacetylases, modulate NF-κB pathway, activate G-protein coupled
receptors (FFAR2/3 and GPR109A); regulate T regulatory cells | Potential adjuvant
therapy; dietary fiber and probiotic supplementation to boost
SCFAs | (26,63,142) |
| Secondary bile
acids (deoxycholic acid, lithocholic acid) | A wide range of gut
microbiota | Gut microbiota
enzymatically converts primary bile acids, induce oxidative stress,
stimulate oncogenic Wnt/β-catenin and NF-κB pathways; DNA damage
induction | Target bile acid
receptors (FXR and TGR5); bile acid metabolism modulators;
antioxidants | (11,34,74) |
| Kynurenine |
Lactobacillus, other gut
microbes | Suppresses
cytotoxic T cell activation by modulating tryptophan metabolism;
facilitates immune evasion | Target immune
checkpoints (PD1/PD-L1); therapeutic tryptophan pathway
modulation | (77,86) |
| Indole compounds
(indole-3-propionic acid, indole-3-acetic acid) |
Bifidobacterium, other gut
commensals | Modulate epithelial
barrier function; regulate immune signaling and inflammatory
responses | Prognostic
biomarkers; therapeutic metabolites supplementation;
microbiome-targeted therapies | (75,81,100) |
| Polyamines
(putrescine, spermine, spermidine) | Bacteroides
species | Stimulate
angiogenesis, tumor cell proliferation, and metastasis through
modulation of gene expression and signaling pathways | Target polyamine
biosynthesis or uptake pathways; potential chemotherapy
adjuncts | (85) |
| Hydrogen sulfide
(H2S) |
Desulfovibrio,
Bilophila | Dual role: supports
mucosal integrity at physiological levels but induces DNA
hypermethylation, inhibits DNA repair, and promotes inflammation at
high concentrations | Potential biomarker
for stromal activation; therapeutic target for modulating H2S
levels | (88,90) |
| ROS | Multiple gut
microbial species | Produce by multiple
gut microbial species and metabolic reactions and induces DNA
damage, lipid peroxidation, and oxidative stress; promotes
inflammatory cytokine production | Antioxidant therapy
adjunct; microbiome modulation to reduce ROS-producing
bacteria | (60,84) |
| Trimethylamine
N-oxide | Various
species | Gut microbiota
metabolizes dietary choline and carnitine, promotes inflammatory
responses and immune cell activation; induces pyroptosis in tumor
cells | Potential
immunotherapeutic adjuvant; dietary modulation of precursors | (85,143) |
|
Lipopolysaccharides | Klebsiella,
Escherichia coli | Activates TLR4
signaling, promotes NF-κB-mediated proinflammatory cytokine
production | Target TLR4
pathway; anti-inflammatory agents; microbiome-directed therapy | (63) |
Remodeling of the TME
The gut microbiota contributes to TME remodeling
within cancer cells by regulating cellular interactions and
cytokine production to support cellular proliferation and survival
(14). Butyrate acts as an
epigenetic regulator in normal colonocytes, inhibits HDAC, induces
transcriptional changes to induce apoptosis and supports epithelial
barrier integrity in normal colonocytes; however, within the
hypoxic TME of CRC, it serves as an energy source, promoting the
survival and chemoresistance of tumor cells (33). Lactate is another metabolite
produced within the TME, owing to the increased demand for energy
sources or by the in-house microbiota (75). By binding to glycoprotein
receptors, such as GPR81, it serves as a metabolic substrate for
signaling molecules and therefore promotes immunosuppression and
angiogenesis in the TME (95).
Streptococcus species acidify the gastric TME by producing
lactate, resulting in VEGF/MMP9 activation and immune evasion
(96,97). Immune evasion is a unique
characteristic of the cancer cells within the TME; this phenomenon
is facilitated by the activation of inflammatory pathways, such as
NF-κB, Wnt/β-catenin signaling and STAT3, which are mediated by
secondary bile acids and the production of reactive oxygen species
(98). In HCC, the overgrowth of
Clostridium scindens was observed under dysbiosis, which is
potentially associated with increased conversion of primary bile
acids to secondary ones such as DCA (36). As aforementioned, the increased
levels of DCA within the TME is mechanistically associated with DNA
damage and inflammation resulting in cancerous cell migration,
invasion and extracellular matrix remodeling (99,100).
Metabolic reprogramming and immune
evasion of microbiota-host immune cells
Microbial metabolites, such as SCFA, kynurenine and
indole derivatives, influence cancer cell proliferation and tumor
growth (8). In dysbiosis, changes
in amino acid metabolism by the gut microbiota (such as
Lactobacillus) alter the levels of amino acid catabolites
such as indole-3-acetic acid and kynurenine, Tryptophan derived
catabolites, which can activate aryl hydrocarbon receptors on
immune cell surface, promote immunosuppressive phenotypes, reduce
T-cytotoxic lymphocytes and mediate immune tolerance in pancreatic
ductal adenocarcinoma (93,101-103).
In addition, increased ammonia levels, originating from proteolytic
bacteria, result in T cell exhaustion and impair antitumor
activity. Microbial metabolites also provide nutrients and immune
cell energy sources within the TME. The microbiota can also act as
proinflammatory mediators. For instance, LPS from gram-negative
bacteria (such as Klebsiella) induce epithelial mesenchymal
transition in pancreatic cancer by activating TLR4 signaling,
resulting in the activation of NF-κB and the release of
proinflammatory cytokines such as TNF-α and IL-6(104).
Genotoxicity and epigenetic
alteration
In CRC, some E. coli strains (such as
phylogroups B1 and B2) can produce colibactin, a genotoxin that
induces chromosomal instability and breakage in double-stranded DNA
and exerts somatic mutations in the KRAS gene, resulting in
mutagenesis (105-107).
In dysbiosis, microbial compounds can also promote the accumulation
of reactive oxygen and nitrogen species, thereby intensify
oxidative DNA damage and promote lipid peroxidation (44). In addition to directed DNA damage,
microbial metabolites such as butyrate (HDAC inhibitor) can induce
epigenetic alterations and affect cellular DNA condensation within
the TME (33,79).
Role of gut microbiota in metabolism
of xenobiotic compounds
Microbial biotransformation is a process by which
the gut microbiota processes chemical compounds, including dietary
substances and xenobiotics (108). Xenobiotics are chemical
substances that do not originate intrinsically, such as
medications, environmental pollutants, dietary compounds and other
chemical substances. The gut microbiota serves a pivotal role in
biotransformation and markedly affects the risk of cancer
development in the digestive tissues. In dysbiosis, microbial
biotransformation may activate carcinogenic compounds, induce DNA
damage and affect host detoxification pathways (108).
The microbial metabolism of xenobiotic compounds
typically involves two main phases (108). In the first phase, the oxidation,
reduction and hydrolysis of xenobiotics are often mediated by
analog compounds of cytochrome P450 (cytP450). For instance,
aflatoxin B1 (AFB-1), a mycotoxin produced by Aspergillus
species; AFB-1, is typically converted by cytP450 to a carcinogenic
epoxide. Gut microbiota, such as Lactobacillus, produce
enzymes that detoxify AFB-1. Under dysbiosis, overproduction of
AFB-1 disrupts DNA structure and leads to HCC (109). The second phase of xenobiotic
metabolism involves increasing the solubility and facilitating the
excretion of xenobiotic compounds (110). This phase includes conjugation,
sulfation and methylation reactions. For example,
Bacteroides and Clostridium produce enzymes similar
to β-glucuronidase and azoreductase, which metabolize heterocyclic
amines, such as polycyclic aromatic hydrocarbons (111). Heterocyclic amines potentially
induce mutations in essential genes, such as adenomatous polyposis
coli (APC) and KRAS, which are risk factors for CRC. Under
dysbiosis, these transformation processes are probably disturbed,
resulting in carcinogenesis (112,113).
In dysbiosis, the gut microbiota can reduce dietary
nitrates to nitrites, and oral Veillonella and gastric H.
pylori convert nitrites to nitrosamines, which are potent
mutagens that contribute to gastric carcinoma (114,115). Environmental pollutants, such as
heavy metals, can also be detoxified by some microbiota strains,
such as Lactobacillus strains, which bind to and reduce the
bioavailability and nephrotoxicity of cadmium and arsenic (116,117). Another notable role of the
microbiota in the metabolism of xenobiotic compounds is
detoxification and drug metabolism (117,118); for instance, Eggerthella
lenta inactivates digoxin, a chemotherapeutic agent, and
Enterococcus species can detoxify and mitigate the
production of cisplatin-induced reactive oxygen species (119).
Gut microbiota modulates immune
response and TME remodeling
The intricate association between the gut microbiota
and immune response in digestive tumors supports their pivotal role
in regulating inflammatory signals, immune cell differentiation,
cytokine production and immune checkpoint alterations within the
TME of digestive tumors (Fig. 2).
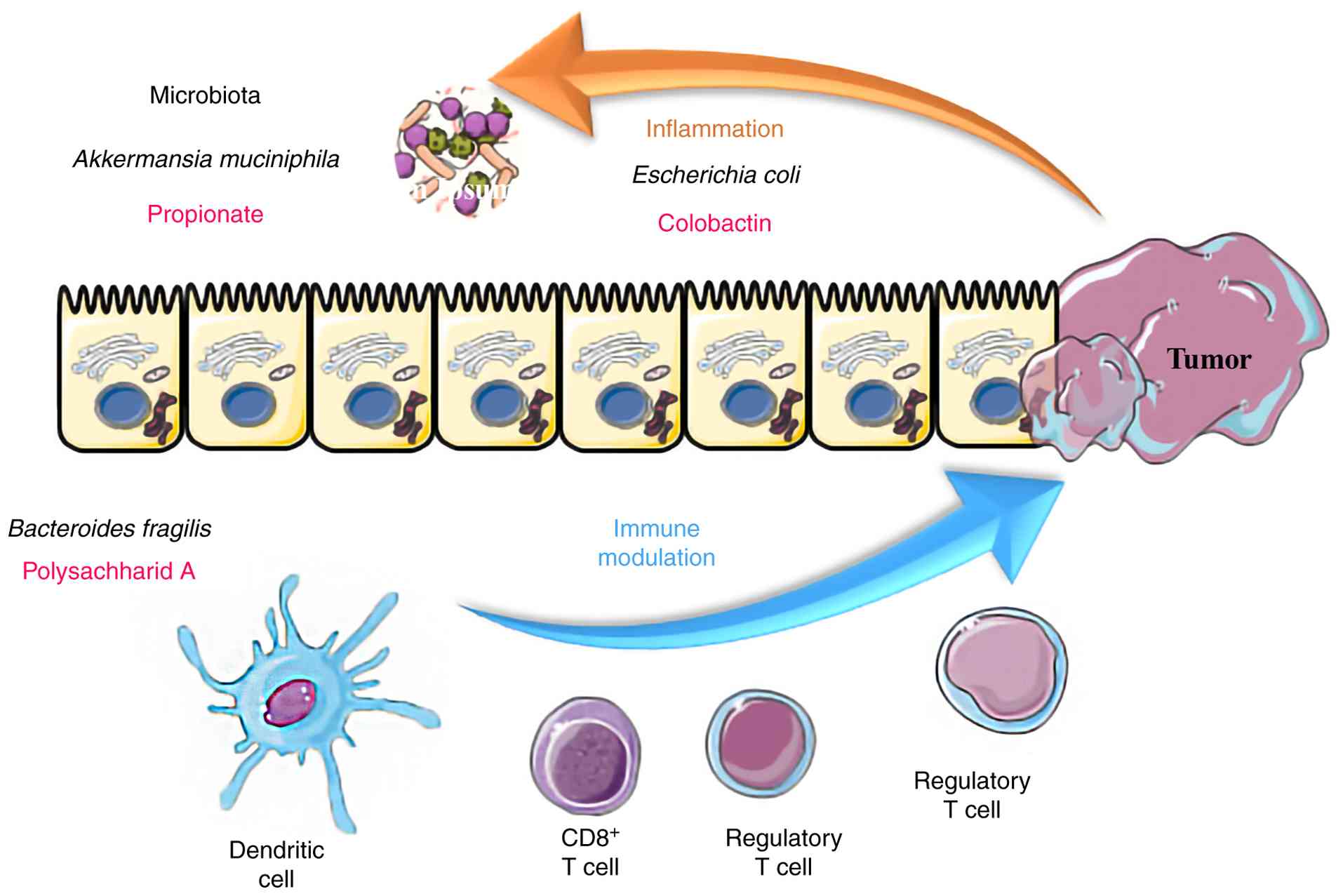
The gut microbiota can directly affect the local immunity of the
TME within digestive tumors, such as gastric and CRC. Within the
gastrointestinal tract, a consistent interplay between the gut
microbiota and mucosal immune system is evident. In dysbiosis,
pathogenic bacteria such as F. nucleatum suppress local
immunity, and in CRC, F. nucleatum inhibits tumor cell
cytotoxicity by binding to the T cell immunoreceptor with Ig and
ITIM domains receptor on natural killer cells and cytotoxic T
lymphocytes (Fig. 3). Dysbiotic
microbiota, such as F. nucleatum induce M2-polarization of
tumor-associated macrophages and block the production of
tumor-suppressor cytokines, such as IL-10 and TGF-β (78,86).
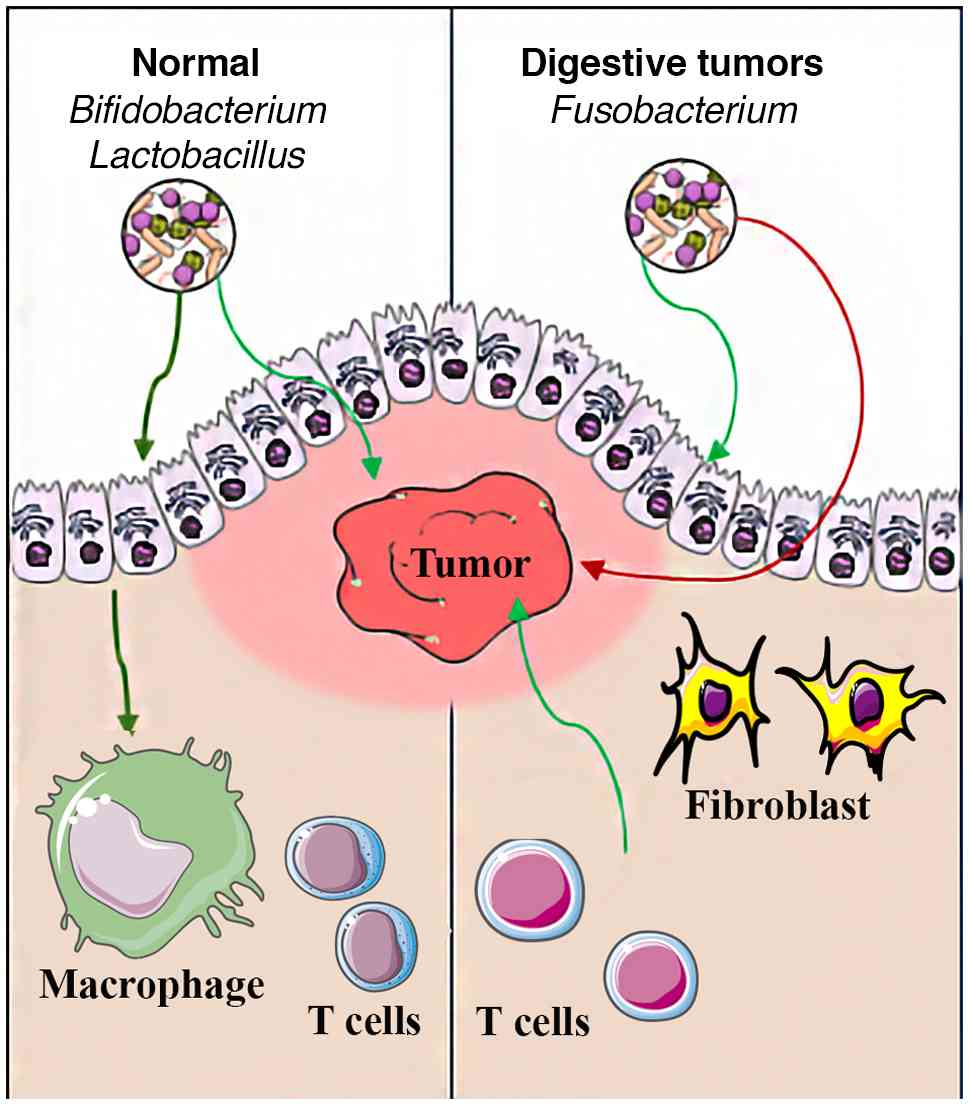
Commensal bacteria are essential for maintaining
epithelial integrity and equilibrium between pro- and
anti-inflammatory signals. A balanced intestinal microbiota is
essential for the host to support immune homeostasis by inducing
the maturation of circulating immune cells, including dendritic
cells, macrophages and different subtypes of T lymphocytes within
secondary lymphoid organs (7,45).
Bifidobacterium and F. prausnitzii produce SCFAs,
tryptophan derivatives and secondary bile acids that can enter the
bloodstream, interact with immune cells and promote
anti-inflammatory responses and cytotoxicity of tumor-infiltrating
lymphocytes (23,24,29).
In addition, microbial translocation has been proposed as a risk
factor for premetastatic niches, mainly because of the immune
suppression and inflammation induced by microbial metabolites. Some
Escherichia strains and Proteus mirabilis have been
found to be incorporated into liver metastases from colorectal
tumors. In dysbiosis, the translocation of gut microbiota, such as
Enterobacteriaceae, results in locally elevated LPS levels and the
activation of TLR4 in hepatic Kupffer cells. These events induce
the activation of IL-6/STAT3 signaling and the progression of tumor
cells in HCC (35,43).
7. Application of gut microbiota in
therapeutic approaches
Mechanistic insights into the interactions of the
gut microbiota-metabolism axis are beneficial for developing
targeted therapeutic strategies, such as manipulating microbiome
composition to induce favorable metabolic or immunological
responses, which is possible through microbiota profiling.
Integrating knowledge of microbiota profile on regulation of the
host-metabolic pathways provides a roadmap to understand how
microbiota-based intervention can be translated in personalized
therapeutic approaches.
Gut microbiota determines response to
treatment
Gut microbiota profiling is useful for predicting
therapeutic outcomes, particularly in digestive tumors. Beyond its
influence on tumor biology, microbial composition can also affect
drug metabolism and determine the efficiency of checkpoint
inhibitors and chemotherapeutic agents (7,118,120). Therefore, microbial diversity can
be used as a determining factor for whether a patient is resistant
or responsive to treatment. For instance, Heshiki et al
(121) investigated the
intestinal microbiota profile in patients with different cancers
and identified a significant microbiota diversity in patients who
were responded successfully to treatment than non-responder
patients. Based on microbiota profile, the authors developed a
machine leaning model to predict treatment outcome indicating a
positive correlation between Bacteroides and positive
response to treatment, and validated the accuracy of model in a
different cohort.
Microbiota modulates response to
immune checkpoint inhibitors
The use of antibodies against immune checkpoint
inhibitors, including anti-programmed cell death protein 1,
anti-programmed death-ligand 1 and anti-cytotoxic T-lymphocyte
associated protein 4, is an emerging therapy that affects T
cell-mediated antitumor activity. Currently, gut microbiota
composition has been introduced as a predictor of response to
treatment with checkpoint inhibitors (12,122). Patients with a microbiota profile
enriched in beneficial commensals, such as A. muciniphila
and Bifidobacterium, usually respond favorably to treatment
(123). These bacteria have the
potential to increase antigen presentation, increase effector
T-cells levels and promote dendritic cell maturation. Transferring
beneficial microbiota to non-responders to immune checkpoint
inhibitors is a promising hypothesis for restoring immunomodulatory
and therapeutic outcomes in a personalized medicine approach.
Microbiota interacts with
chemotherapeutic agents
Multiple studies on the influence of gut microbiota
on treatment outcomes with chemotherapeutic agents have indicated
that microbial enzymes can induce the activation or degradation of
certain medications (124,125).
For example, Mycoplasma hyorhinis produces cytidine
deaminase, which inactivates gemcitabine in patients with
pancreatic cancer. In addition, Lactobacillus species exert
a synergistic effect on 5-FU cytotoxicity by producing thymidylate
synthase inhibitors (126).
8. Challenges in clinical translation of
microbiota-targeted therapies
Although the use of beneficial microbiota in the
real world seems promising through the administration of
probiotics, prebiotics, fecal microbiota transplantation or
medications targeting the gut microbiota, several challenges
confine its clinical translation. First, inter-individual
variability exists in the microbial composition; these
variabilities, which arise from geography, genetics, age and
lifestyle, can complicate patient stratification and prediction of
the response to treatment (127).
The possibility of confounding factors, such as the use of
medications targeting proton pump inhibitors, antibiotics or
dietary habits, are other challenging factors that limit the
reproducibility of microbiota in different populations (128). For instance, antibiotics can
eradicate beneficial taxa, such as A. muciniphila (129); another example is the variability
in the use of dietary fibers, as they enhance the production of
SCFAs. Therefore, it is necessary to control dietary intake when
designing clinical trials (130).
Beyond the inter-individual variabilities, the long-term safety of
sustained microbiota-targeted interventions remains incompletely
characterized; in addition, the standardization of sampling,
sequencing and functional validation, along with its regulatory
framework is essential for more robust, guideline-based clinical
validation.
9. Discussion and future directions
In the present review, the intricate and
multifaceted importance of the gut microbiota in digestive tumors
was comprehensively explored. The microbiota-metabolism axis exerts
substantial influence on digestive tumorigenesis through mainly
regulating cellular processes such as metabolic reprogramming,
epigenetic remodeling and immunomodulating within the TME. In
particular, microbial dysbiosis and its related key metabolites,
including polyamines, SCFA, secondary bile acids and indole
derivatives, were found to be critical mediators of epithelial
integrity and tumor immunity by promoting chronic inflammation, DNA
damage, genotoxic injury and metabolic reprogramming.
In addition, the bridging of immunology, oncology,
microbiology and pharmacology is a unique aspect, highlighting the
bidirectional TME-microbial metabolites in the digestive system,
which have often been overlooked in research. As previously
discussed, microbial compounds and metabolites can be incorporated
into the host metabolism-gut microbiome to regulate signaling
pathways (such as Wnt/β-catenin, STAT3 and NF-κB pathways),
epigenetics, inflammation and immune evasion of CRC cells. The
present review discussed the influence of microbial metabolites,
such as SCFA and bile acids, on immunomodulation, inflammation and
regulation of cellular signaling. For instance, beneficial
commensals such as A. muciniphila and F. praussnitzii
were identified to have anti-inflammatory and antitumor properties
owing to the production of SCFA and maintenance of epithelial
barrier integrity. However, pathogenic bacteria such as H.
pylori, F. nucleatum, and some strains of E. coli
produce destructive metabolites and toxins that result in
inflammation, impaired immune activity, DNA damage and
oncogenesis.
Another unique aspect of the present review is the
consideration of the gut microbiota as both a modulator and
predictor in the counterpart aspects of health and pathogenesis.
According to this concept, the microbiota is valid in research on
precision oncology and personalized medicine. Future studies may
identify these findings not only as biomarkers for early diagnosis
but also to elucidate the microbial metabolic pathways for future
interventions to block tumor-promoting pathways.
In clinical practice, microbiota modulation can be
achieved using antibiotics, probiotics, prebiotics, dietary
approaches and fecal microbiota transplantation to restore a
healthy microbiome that can enhance antitumor immunity or even
remove inflammation. New emerging data have demonstrated the role
of microbiota composition in influencing the efficacy of immune
checkpoint inhibitors or chemotherapy, emphasizing the role of this
axis in personalized treatment strategies.
Despite the unique aspects of the present review,
it has some limitations that should be considered. Although it was
attempted to explore the functional role of the microbiota in the
digestive system, it was not possible to completely address the
complexity of the host microbiota-metabolism axis across different
conditions. Through increasing knowledge of the mechanistically
influence of microbiome aiming for biomarker discovery and
therapeutic targeting across digestive tumors, future research can
utilize them as determinant biomarkers for risk stratification,
predict the response to treatments or consider them as checkpoint
inhibitors. Future research should explore the functional
validation of microbiota-host interplay by designing observational
studies utilizing dietary, probiotic or prebiotic interventions.
Future studies should investigate gut microbiome alterations under
different pathological and physiological conditions across
different populations. By systematically increasing knowledge of
microbiome-metabolite complexities in health and disease, they
could be considered promising therapeutic candidates for routine
clinical cancer treatments. Although some studies have investigated
the short-term influence of gut microbiota in clinical trials
(131,132); studies investigating the
long-term effects of probiotic administration on digestive diseases
could not be found. Future research should focus on the dynamic
changes in the gut microbiota during cancer management of clinical
interventions.
In addition, crosstalk studies on the integration
of multi-omics data, combining emerging genomics, metabolomics and
immune profiling as part of longitudinal cohorts, will enhance the
understanding of causal relationships and identify potential
predictive biomarkers. Future research should be designed to
address several specific multi-omics aspects to develop
personalized microbiota information for precision medicine. First,
prospective cohort studies would be beneficial to integrate
metagenomics and meta-transcriptomics profiling of the gut
microbiota, which is associated with host multi-omics alterations
across pathogenic conditions. Improving knowledge in this field
(from premalignant lesions to advanced digestive tumors) will be
applied in personalized medicine, both in biomarker discovery and
personalized therapeutic strategies. Second, studies on multilayer
datasets, combining microbiome composition and host interactions,
will enhance knowledge of the mechanisms of pathogenesis, immune
escape, pharmacokinetic aspects of anticancer agents and clarifying
how microbiota taxa influence drug efficiency, dose optimization
and treatment selection. Third, integrative studies on detailed
dietary patterns with microbiome signatures and host-specific
signaling pathways could be promising, bridging lifestyle
modifications in high-risk populations.
Clinical trials in the field of microbiota
modulation should be designed to assess the integrity of
microbiome. Future studies should investigate the interaction of
dietary components with gut microbiota dynamics and their influence
on gut microbiota function to develop new nutritional strategies
for the prevention of tumorigenesis or to improve treatment
response. Future studies should elucidate and characterize how gut
microbes alter the pharmacokinetics and pharmacodynamics of
anticancer drugs to optimize dosing and limit their toxicity.
In conclusion, the gut microbiota-metabolism axis
represents a disruptive paradigm in the field of digestive tumors,
with potentially important implications for novel diagnostics and
therapeutics. Highlighting the dual function of gut microbial
metabolites in tumor progression and repression, the present review
emphasizes the importance of microbiome analysis in personalized
medicine. The integration of microbiology, oncology, immunology and
systems biology will enable interdisciplinary discovery of
actionable targets. Ultimately, these combinatorial strategies and
personalized microbiome-based therapies can improve patient
outcomes.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
ZY as the first author was contributed to the study
conceptualization and design, data acquisition and interpretation,
and providing the primary draft of the manuscript. SW and XY were
the co-authors who contributed to the study design, literature
searching, data collection and interpretation, and assistance for
writing the manuscript. The project administration was equally
supported by SW and XY. BS as the correspondence was involved in
all the study process including conceptualization and the study
design, supervision, consensus and discussion and review and
editing of the manuscript. All co-authors participated in writing
and evaluating the manuscript. All authors read and approved the
final version of the manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhan T, Betge J, Schulte N, Dreikhausen L,
Hirth M, Li M, Weidner P, Leipertz A, Teufel A and Ebert MP:
Digestive cancers: Mechanisms, therapeutics and management. Signal
Transduct Target Ther. 10(24)2025.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Bray F, Laversanne M, Sung H, Ferlay J,
Siegel RL, Soerjomataram I and Jemal A: Global cancer statistics
2022: GLOBOCAN estimates of incidence and mortality worldwide for
36 cancers in 185 countries. CA Cancer J Clin. 74:229–263.
2024.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Mousavi SE, Ilaghi M, Elahi Vahed I and
Nejadghaderi SA: Epidemiology and socioeconomic correlates of
gastric cancer in Asia: Results from the GLOBOCAN 2020 data and
projections from 2020 to 2040. Sci Rep. 15(6529)2025.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Ferreira RM, Pereira-Marques J,
Pinto-Ribeiro I, Costa JL, Carneiro F, Machado JC and Figueiredo C:
Gastric microbial community profiling reveals a dysbiotic
cancer-associated microbiota. Gut. 67:226–236. 2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Buchta Rosean C, Feng TY, Azar FN and
Rutkowski MR: Impact of the microbiome on cancer progression and
response to anti-cancer therapies. Adv Cancer Res. 143:255–294.
2019.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Singh R, Zogg H, Wei L, Bartlett A,
Ghoshal UC, Rajender S and Ro S: Gut microbial dysbiosis in the
pathogenesis of gastrointestinal dysmotility and metabolic
disorders. J Neurogastroenterol Motil. 27:19–34. 2021.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Asseri AH, Bakhsh T, Abuzahrah SS, Ali S
and Rather IA: The gut dysbiosis-cancer axis: Illuminating novel
insights and implications for clinical practice. Front Pharmacol.
14(1208044)2023.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Rooks MG and Garrett WS: Gut microbiota,
metabolites and host immunity. Nat Rev Immunol. 16:341–352.
2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Wang Z, Li L, Wang S, Wei J, Qu L, Pan L
and Xu K: The role of the gut microbiota and probiotics associated
with microbial metabolisms in cancer prevention and therapy. Front
Pharmacol. 13(1025860)2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Feng W, Liu J, Cheng H, Zhang D, Tan Y and
Peng C: Dietary compounds in modulation of gut microbiota-derived
metabolites. Front Nutr. 9(939571)2022.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Molinaro A, Wahlström A and Marschall HU:
Role of bile acids in metabolic control. Trends Endocrinol Metab.
29:31–41. 2018.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Barrueto L, Caminero F, Cash L, Makris C,
Lamichhane P and Deshmukh RR: Resistance to checkpoint inhibition
in cancer immunotherapy. Transl Oncol. 13(100738)2020.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Bae J, Park K and Kim YM: Commensal
microbiota and cancer immunotherapy: Harnessing commensal bacteria
for cancer therapy. Immune Netw. 22(e3)2022.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Feng P, Xue X, Bukhari I, Qiu C, Li Y,
Zheng P and Mi Y: Gut microbiota and its therapeutic implications
in tumor microenvironment interactions. Front Microbiol.
15(1287077)2024.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Yang Z, Wang X, Zhou H, Jiang M, Wang J
and Sui B: Molecular complexity of colorectal cancer: Pathways,
biomarkers, and therapeutic strategies. Cancer Manag Res.
16:1389–1403. 2024.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Yang Z, Chang Y, Hu Y, An W, Xing C and
Sui B: A concise review on advancement in molecular targeted
therapy for lung cancer. Period Biol. 126:15–31. 2024.
|
|
17
|
Ryu MH, Lee JL, Chang HM, Kim TW, Kang HJ,
Sohn HJ, Lee JS and Kang YK: Patterns of progression in
gastrointestinal stromal tumor treated with imatinib mesylate. Jpn
J Clin Oncol. 36:17–24. 2006.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Gao X, Yin P, Ren Y, Yu L, Tian F, Zhao J,
Chen W, Xue Y and Zhai Q: Predicting personalized diets based on
microbial characteristics between patients with superficial
gastritis and atrophic gastritis. Nutrients.
15(4738)2023.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Mei S, Deng Z, Chen Y, Ning D, Guo Y, Fan
X, Wang R, Meng Y, Zhou Q and Tian X: Dysbiosis: The first hit for
digestive system cancer. Front Physiol. 13(1040991)2022.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Cruz MS, Tintelnot J and Gagliani N: Roles
of microbiota in pancreatic cancer development and treatment. Gut
Microbes. 16(2320280)2024.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Rinninella E, Raoul P, Cintoni M,
Franceschi F, Miggiano GAD, Gasbarrini A and Mele MC: What is the
healthy gut microbiota composition? A changing ecosystem across
age, environment, diet, and diseases. Microorganisms.
7(14)2019.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Magne F, Gotteland M, Gauthier L, Zazueta
A, Pesoa S, Navarrete P and Balamurugan R: The
firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis
in obese patients? Nutrients. 12(1474)2020.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Colombo F, Illescas O, Noci S, Minnai F,
Pintarelli G, Pettinicchio A, Vannelli A, Sorrentino L, Battaglia
L, Cosimelli M, et al: Gut microbiota composition in colorectal
cancer patients is genetically regulated. Sci Rep.
12(11424)2022.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Li N, Bai C, Zhao L, Sun Z, Ge Y and Li X:
The relationship between gut microbiome features and chemotherapy
response in gastrointestinal cancer. Front Oncol.
11(781697)2021.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Kulmambetova G, Kurentay B, Gusmaulemova
A, Utupov T, Auganova D, Tarlykov P, Mamlin M, Khamzina S,
Shalekenov S and Kozhakhmetov A: Association of Fusobacterium
nucleatum infection with colorectal cancer in Kazakhstani
patients. Front Oncol. 14(1473575)2024.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Abdulla MH, Agarwal D, Singh JK, Traiki
TB, Pandey MK, Ahmad R and Srivastava SK: Association of the
microbiome with colorectal cancer development (review). Int J
Oncol. 58(17)2021.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Osman MA, Neoh HM, Ab Mutalib NS, Chin SF,
Mazlan L, Raja Ali RA, Zakaria AD, Ngiu CS, Ang MY and Jamal R:
Parvimonas micra, Peptostreptococcus stomatis,
Fusobacterium nucleatum and Akkermansia muciniphila as a
four-bacteria biomarker panel of colorectal cancer. Sci Rep.
11(2925)2021.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Conde-Pérez K, Buetas E, Aja-Macaya P,
Martin-De Arribas E, Iglesias-Corrás I, Trigo-Tasende N, Nasser-Ali
M, Estévez LS, Rumbo-Feal S, Otero-Alén B, et al: Parvimonas
micra can translocate from the subgingival sulcus of the human oral
cavity to colorectal adenocarcinoma. Mol Oncol. 18:1143–1173.
2024.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Effendi RMRA, Anshory M, Kalim H, Dwiyana
RF, Suwarsa O, Pardo LM, Nijsten TEC and Thio HB: Akkermansia
muciniphila and Faecalibacterium prausnitzii in
immune-related diseases. Microorganisms. 10(2382)2022.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Montgomery TL, Toppen LC, Eckstrom K,
Heney ER, Kennedy JJ, Scarborough MJ and Krementsov DN:
Lactobacillaceae differentially impact butyrate-producing gut
microbiota to drive CNS autoimmunity. Gut Microbes.
16(2418415)2024.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Gomes S, Baltazar F, Silva E and Preto A:
Microbiota-derived short-chain fatty acids: New road in colorectal
cancer therapy. Pharmaceutics. 14(2359)2022.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Liu G, Tang J, Zhou J and Dong M:
Short-chain fatty acids play a positive role in colorectal cancer.
Discov Oncol. 15(425)2024.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Han A, Bennett N, Ahmed B, Whelan J and
Donohoe DR: Butyrate decreases its own oxidation in colorectal
cancer cells through inhibition of histone deacetylases.
Oncotarget. 9:27280–27292. 2018.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Xu Z, Xiao L, Wang S, Cheng Y, Wu J, Meng
Y, Bao K, Zhang J and Cheng C: Alteration of gastric microbiota and
transcriptome in a rat with gastric intestinal metaplasia induced
by deoxycholic acid. Front Microbiol. 14(1160821)2023.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Wang S, Kuang J, Zhang H, Chen W, Zheng X,
Wang J, Huang F, Ge K, Li M, Zhao M, et al: Bile acid-microbiome
interaction promotes gastric carcinogenesis. Adv Sci (Weinh).
9(e2200263)2022.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Ma C, Han M, Heinrich B, Fu Q, Zhang Q,
Sandhu M, Agdashian D, Terabe M, Berzofsky JA, Fako V, et al: Gut
microbiome-mediated bile acid metabolism regulates liver cancer via
NKT cells. Science. 360(eaan5931)2018.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Phelan JP, Reen FJ, Caparros-Martin JA,
O'Connor R and O'Gara F: Rethinking the bile acid/gut microbiome
axis in cancer. Oncotarget. 8:115736–115747. 2017.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chen C, Shen J, Du Y, Shi X, Niu Y, Jin G,
Liu Y, Shi Y, Lyu J and Lin L: Characteristics of gut microbiota in
patients with gastric cancer by surgery, chemotherapy and lymph
node metastasis. Clin Transl Oncol. 24:2181–2190. 2022.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Dai D, Yang Y, Yu J, Dang T, Qin W, Teng
L, Ye J and Jiang H: Interactions between gastric microbiota and
metabolites in gastric cancer. Cell Death Dis.
12(1104)2021.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Correa P and Piazuelo MB: Helicobacter
pylori infection and gastric adenocarcinoma. US Gastroenterol
Hepatol Rev. 7:59–64. 2011.PubMed/NCBI
|
|
41
|
Yu D, Yang J, Jin M, Zhou B, Shi L, Zhao
L, Zhang J, Lin Z, Ren J, Liu L, et al: Fecal Streptococcus
alteration is associated with gastric cancer occurrence and liver
metastasis. mBio. 12(e0299421)2021.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Jang S, Hansen LM, Su H, Solnick JV and
Cha JH: Host immune response mediates changes in cagA copy number
and virulence potential of Helicobacter pylori. Gut
Microbes. 14(2044721)2022.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Jin D, Huang K, Xu M, Hua H, Ye F, Yan J,
Zhang G and Wang Y: Deoxycholic acid induces gastric intestinal
metaplasia by activating STAT3 signaling and disturbing gastric
bile acids metabolism and microbiota. Gut Microbes.
14(2120744)2022.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Sah DK, Arjunan A, Lee B and Jung YD:
Reactive oxygen species and H. pylori infection: A
comprehensive review of their roles in gastric cancer development.
Antioxidants (Basel). 12(1712)2023.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Blackwood BP, Yuan CY, Wood DR, Nicolas
JD, Grothaus JS and Hunter CJ: Probiotic Lactobacillus
species strengthen intestinal barrier function and tight junction
integrity in experimental necrotizing enterocolitis. J Probiotics
Health. 5(159)2017.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Devi TB, Devadas K, George M, Gandhimathi
A, Chouhan D, Retnakumar RJ, Alexander SM, Varghese J, Dharmaseelan
S, Chandrika SK, et al: Low Bifidobacterium abundance in the
lower gut microbiota is associated with Helicobacter
pylori-related gastric ulcer and gastric cancer. Front
Microbiol. 12(631140)2021.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Park JY, Seo H, Kang CS, Shin TS, Kim JW,
Park JM, Kim JG and Kim YK: Dysbiotic change in gastric microbiome
and its functional implication in gastric carcinogenesis. Sci Rep.
12(4285)2022.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Yuan L, Pan L, Wang Y, Zhao J, Fang L,
Zhou Y, Xia R, Ma Y, Jiang Z, Xu Z, et al: Characterization of the
landscape of the intratumoral microbiota reveals that
Streptococcus anginosus increases the risk of gastric cancer
initiation and progression. Cell Discov. 10(117)2024.PubMed/NCBI View Article : Google Scholar
|
|
49
|
May MS, Park H, Moallem DH, Seeram D,
Dajiang S, Hibshoosh H, Jamison JK, Uhlemann AC and Manji GA: Low
bacterial biomass in human pancreatic cancer and adjacent normal
tissue. Int J Mol Sci. 26(140)2024.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Pushalkar S, Hundeyin M, Daley D,
Zambirinis CP, Kurz E, Mishra A, Mohan N, Aykut B, Usyk M, Torres
LE, et al: The pancreatic cancer microbiome promotes oncogenesis by
induction of innate and adaptive immune suppression. Cancer Discov.
8:403–416. 2018.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Pust MM, Rocha Castellanos DM, Rzasa K,
Dame A, Pishchany G, Assawasirisin C, Liss A, Fernandez-Del
Castillo C and Xavier RJ: Absence of a pancreatic microbiome in
intraductal papillary mucinous neoplasm. Gut. 73:1131–1141.
2024.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Bellotti R, Speth C, Adolph TE, Lass-Flörl
C, Effenberger M, Öfner D and Maglione M: Micro- and mycobiota
dysbiosis in pancreatic ductal adenocarcinoma development. Cancers
(Basel). 13(3431)2021.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Wei X, Mei C, Li X and Xie Y: The unique
microbiome and immunity in pancreatic cancer. Pancreas. 50:119–129.
2021.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Wang S, Li Y, Xing C, Ding C, Zhang H,
Chen L, You L, Dai M and Zhao Y: Tumor microenvironment in
chemoresistance, metastasis and immunotherapy of pancreatic cancer.
Am J Cancer Res. 10:1937–1953. 2020.PubMed/NCBI
|
|
55
|
Nista EC, Del Gaudio A, Del Vecchio LE,
Mezza T, Pignataro G, Piccioni A, Gasbarrini A, Franceschi F and
Candelli M: Pancreatic cancer resistance to treatment: The role of
microbiota. Biomedicines. 11(157)2023.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Saenz C, Fang Q, Gnanasekaran T, Trammell
SAJ, Buijink JDA, Pisano P, Wierer M, Moens F, Lengger B, Brejnrod
A and Arumugam M: Clostridium scindens secretome suppresses
virulence gene expression of Clostridioides difficile in a bile
acid-independent manner. Microbiol Spectr.
11(e0393322)2023.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Li W, Chen H and Tang J: Interplay between
bile acids and intestinal microbiota: Regulatory mechanisms and
therapeutic potential for infections. Pathogens.
13(702)2024.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Wu Z, Guo J, Zhang Z, Gao S, Huang M, Wang
Y, Zhang Y, Li Q and Li J: Bacteroidetes promotes esophageal
squamous carcinoma invasion and metastasis through LPS-mediated
TLR4/Myd88/NF-κB pathway and inflammatory changes. Sci Rep.
14(12827)2024.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Greathouse KL, Stone JK, Vargas AJ,
Choudhury A, Padgett RN, White JR, Jung A and Harris CC:
Co-enrichment of cancer-associated bacterial taxa is correlated
with immune cell infiltrates in esophageal tumor tissue. Sci Rep.
14(2574)2024.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Mostafavi Abdolmaleky H and Zhou JR: Gut
microbiota dysbiosis, oxidative stress, inflammation, and
epigenetic alterations in metabolic diseases. Antioxidants (Basel).
13(985)2024.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Nguyen TT, Ung TT, Li S, Lian S, Xia Y,
Park SY and Do Jung Y: Metformin inhibits lithocholic acid-induced
interleukin 8 upregulation in colorectal cancer cells by
suppressing ROS production and NF-kB activity. Sci Rep.
9(2003)2019.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Sun J, Chen S, Zang D, Sun H, Sun Y and
Chen J: Butyrate as a promising therapeutic target in cancer: From
pathogenesis to clinic (review). Int J Oncol. 64(44)2024.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Xiao T, Wu S, Yan C, Zhao C, Jin H, Yan N,
Xu J, Wu Y, Li C, Shao Q and Xia S: Butyrate upregulates the TLR4
expression and the phosphorylation of MAPKs and NK-κB in colon
cancer cell in vitro. Oncol Lett. 16:4439–4447.
2018.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Kaźmierczak-Siedlecka K, Marano L, Merola
E, Roviello F and Połom K: Sodium butyrate in both prevention and
supportive treatment of colorectal cancer. Front Cell Infect
Microbiol. 12(1023806)2022.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Rodríguez-Enríquez S, Robledo-Cadena DX,
Gallardo-Pérez JC, Pacheco-Velázquez SC, Vázquez C, Saavedra E,
Vargas-Navarro JL, Blanco-Carpintero BA, Marín-Hernández Á,
Jasso-Chávez R, et al: Acetate promotes a differential energy
metabolic response in human HCT 116 and COLO 205 colon cancer cells
impacting cancer cell growth and invasiveness. Front Oncol.
11(697408)2021.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Cornall LM, Mathai ML, Hryciw DH and
McAinch AJ: The therapeutic potential of GPR43: A novel role in
modulating metabolic health. Cell Mol Life Sci. 70:4759–4770.
2013.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Yao T, Dong X, Lv J, Fu L and Li L:
Propionate alleviated colitis by modulating iron homeostasis to
inhibit ferroptosis and macrophage polarization. Int
Immunopharmacol. 162(115151)2025.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Li Y, Zhao M, Lin Y, Jiang X, Jin L, Ye P,
Lu Y, Pei R and Jiang L: Licochalcone A induces
mitochondria-dependent apoptosis and interacts with venetoclax in
acute myeloid leukemia. Eur J Pharmacol. 968(176418)2024.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Kozoni V, Tsioulias G, Shiff S and Rigas
B: The effect of lithocholic acid on proliferation and apoptosis
during the early stages of colon carcinogenesis: differential
effect on apoptosis in the presence of a colon carcinogen.
Carcinogenesis. 21:999–1005. 2000.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Luu TH, Bard JM, Carbonnelle D, Chaillou
C, Huvelin JM, Bobin-Dubigeon C and Nazih H: Lithocholic bile acid
inhibits lipogenesis and induces apoptosis in breast cancer cells.
Cell Oncol (Dordr). 41:13–24. 2018.PubMed/NCBI View Article : Google Scholar
|
|
71
|
Trah J, Arand J, Oh J, Pagerols-Raluy L,
Trochimiuk M, Appl B, Heidelbach H, Vincent D, Saleem MA,
Reinshagen K, et al: Lithocholic bile acid induces apoptosis in
human nephroblastoma cells: A non-selective treatment option. Sci
Rep. 10(20349)2020.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Schwarcz S, Kovács P, Nyerges P, Ujlaki G,
Sipos A, Uray K, Bai P and Mikó E: The bacterial metabolite,
lithocholic acid, has antineoplastic effects in pancreatic
adenocarcinoma. Cell Death Discov. 10(248)2024.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Nguyen TT, Ung TT, Kim NH and Jung YD:
Role of bile acids in colon carcinogenesis. World J Clin Cases.
6:577–588. 2018.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Carino A, Graziosi L, D'Amore C, Cipriani
S, Marchianò S, Marino E, Zampella A, Rende M, Mosci P, Distrutti
E, et al: The bile acid receptor GPBAR1 (TGR5) is expressed in
human gastric cancers and promotes epithelial-mesenchymal
transition in gastric cancer cell lines. Oncotarget. 7:61021–61035.
2016.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Su X, Gao Y and Yang R: Gut
microbiota-derived tryptophan metabolites maintain gut and systemic
homeostasis. Cells. 11(2296)2022.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Zhao P, Chen Y, Zhou S and Li F: Microbial
modulation of tryptophan metabolism links gut microbiota to disease
and its treatment. Pharmacol Res. 219(107896)2025.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Wang Z, Yin M, Zhou R, Li M, Peng J and
Wang Z: Kynurenine promotes the immune escape of colorectal cancer
cells via NAT10-mediated ac4C acetylation of PD-L1.
Clinics (Sao Paulo). 80(100658)2025.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Dehhaghi M, Kazemi Shariat Panahi H, Heng
B and Guillemin GJ: The gut microbiota, kynurenine pathway, and
immune system interaction in the development of brain cancer. Front
Cell Dev Biol. 8(562812)2020.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Ren B, Fang Y, Gu M, You L, Zhang T and
Zhao Y: Microbiota-metabolism-epigenetics-immunity axis in cancer.
Front Immunol. 15(1449912)2024.
|
|
80
|
Wilton J, de Mendonça FL, Pereira-Castro
I, Tellier M, Nojima T, Costa AM, Freitas J, Murphy S, Oliveira MJ,
Proudfoot NJ and Moreira A: Pro-inflammatory polarization and
colorectal cancer modulate alternative and intronic polyadenylation
in primary human macrophages. Front Immunol.
14(1182525)2023.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Li Y, Li Q, Yuan R, Wang Y, Guo C and Wang
L: Bifidobacterium breve-derived indole-3-lactic acid
ameliorates colitis-associated tumorigenesis by directing the
differentiation of immature colonic macrophages. Theranostics.
14:2719–2735. 2024.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Hunzeker ZE, Zhao L, Kim AM, Parker JM,
Zhu Z, Xiao H, Bai Q, Wakefield MR and Fang Y: The role of IL-22 in
cancer. Med Oncol. 41(240)2024.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Liu X, Li S, Wang L and Ma K:
Microecological regulation in HCC therapy: Gut microbiome enhances
ICI treatment. Biochim Biophys Acta Mol Basis Dis.
1870(167230)2024.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Goodwin AC, Destefano Shields CE, Wu S,
Huso DL, Wu X, Murray-Stewart TR, Hacker-Prietz A, Rabizadeh S,
Woster PM, Sears CL and Casero RA Jr: Polyamine catabolism
contributes to enterotoxigenic Bacteroides fragilis-induced
colon tumorigenesis. Proc Natl Acad Sci USA. 108:15354–15359.
2011.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Holbert CE, Cullen MT, Casero RA Jr and
Stewart TM: Polyamines in cancer: Integrating organismal metabolism
and antitumour immunity. Nat Rev Cancer. 22:467–480.
2022.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Schlichtner S, Yasinska IM, Klenova E,
Abooali M, Lall GS, Berger SM, Ruggiero S, Cholewa D, Milošević M,
Gibbs BF, et al: L-Kynurenine participates in cancer immune evasion
by downregulating hypoxic signaling in T lymphocytes.
Oncoimmunology. 12(2244330)2023.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Snezhkina AV, Krasnov GS, Lipatova AV,
Sadritdinova AF, Kardymon OL, Fedorova MS, Melnikova NV, Stepanov
OA, Zaretsky AR, Kaprin AD, et al: The dysregulation of polyamine
metabolism in colorectal cancer is associated with overexpression
of c-Myc and C/EBPβ rather than enterotoxigenic Bacteroides
fragilis infection. Oxid Med Cell Longev.
2016(2353560)2016.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Braccia DJ, Jiang X, Pop M and Hall AB:
The capacity to produce hydrogen sulfide (H2S) via cysteine
degradation is ubiquitous in the human gut microbiome. Front
Microbiol. 12(705583)2021.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Kumar A, Ali A, Kapardar RK, Dar GM,
Nimisha Apurva, Sharma AK, Verma R, Sattar RSA, Ahmad E, et al:
Implication of gut microbes and its metabolites in colorectal
cancer. J Cancer Res Clin Oncol. 149:441–465. 2023.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Blachier F, Andriamihaja M, Larraufie P,
Ahn E, Lan A and Kim E: Production of hydrogen sulfide by the
intestinal microbiota and epithelial cells and consequences for the
colonic and rectal mucosa. Am J Physiol Gastrointest Liver Physiol.
320:G125–G135. 2021.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Wallace JL, Motta JP and Buret AG:
Hydrogen sulfide: An agent of stability at the microbiome-mucosa
interface. Am J Physiol Gastrointest Liver Physiol. 314:G143–G149.
2018.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Lu W and Wen J: Anti-inflammatory effects
of hydrogen sulfide in axes between gut and other organs. Antioxid
Redox Signal. 42:341–360. 2025.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Luu M, Schütz B, Lauth M and Visekruna A:
The impact of gut microbiota-derived metabolites on the tumor
immune microenvironment. Cancers (Basel). 15(1588)2023.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Arya P, Sharma V, Singh P, Thapliyal S and
Sharma M: Bacterial endotoxin-lipopolysaccharide role in
inflammatory diseases: An overview. Iran J Basic Med Sci.
28:553–564. 2025.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Li X, Yang Y, Zhang B, Lin X, Fu X, An Y,
Zou Y, Wang JX, Wang Z and Yu T: Lactate metabolism in human health
and disease. Signal Transduct Target Ther. 7(305)2022.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Zi M, Zhang Y, Hu C, Zhang S, Chen J, Yuan
L and Cheng X: A literature review on the potential clinical
implications of streptococci in gastric cancer. Front Microbiol.
13(1010465)2022.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Fu K, Cheung AHK, Wong CC, Liu W, Zhou Y,
Wang F, Huang P, Yuan K, Coker OO, Pan Y, et al:
Streptococcus anginosus promotes gastric inflammation,
atrophy, and tumorigenesis in mice. Cell. 187:882–896.e17.
2024.
|
|
98
|
Liu H, Pan M, Liu M, Zeng L, Li Y, Huang
Z, Guo C and Wang H: Lactate: A rising star in tumors and
inflammation. Front Immunol. 15(1496390)2024.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Li L, Wang M, Jiang Z, Yi C and Yi Y:
Intratumoral microbial heterogeneity in HCC reveals a potential
therapeutic target. JHEP Rep. 7(101538)2025.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Bi C, Xiao G, Liu C, Yan J, Chen J, Si W,
Zhang J and Liu Z: Molecular immune mechanism of intestinal
microbiota and their metabolites in the occurrence and development
of liver cancer. Front Cell Dev Biol. 9(702414)2021.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Chen Z, Guan D, Wang Z, Li X, Dong S,
Huang J and Zhou W: Microbiota in cancer: Molecular mechanisms and
therapeutic interventions. MedComm (2020). 4(e417)2023.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Liu H, Xiong X, Zhu W, Wang S, Huang W,
Zhu G, Xu H and Yang L: Gut microbial metabolites in cancer
immunomodulation. Mol Cancer. 25(8)2025.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Chandra V and McAllister F: Therapeutic
potential of microbial modulation in pancreatic cancer. Gut.
70:1419–1425. 2021.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Cao W, Zheng C, Xu X, Jin R, Huang F, Shi
M, He Z, Luo Y, Liu L, Liu Z, et al: Clostridium butyricum
potentially improves inflammation and immunity through alteration
of the microbiota and metabolism of gastric cancer patients after
gastrectomy. Front Immunol. 13(1076245)2022.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Zhang G and Sun D: The synthesis of the
novel Escherichia coli toxin-colibactin and its mechanisms
of tumorigenesis of colorectal cancer. Front Microbiol.
15(1501973)2024.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Iftekhar A, Berger H, Bouznad N, Heuberger
J, Boccellato F, Dobrindt U, Hermeking H, Sigal M and Meyer TF:
Genomic aberrations after short-term exposure to
colibactin-producing E. coli transform primary colon
epithelial cells. Nat Commun. 12(1003)2021.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Guragain M, Schmidt JW, Kalchayanand N,
Dickey AM and Bosilevac JM: Characterization of Escherichia
coli harboring colibactin genes (clb) isolated from beef
production and processing systems. Sci Rep. 12(5305)2022.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Koppel N, Maini Rekdal V and Balskus EP:
Chemical transformation of xenobiotics by the human gut microbiota.
Science. 356(eaag2770)2017.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Katati B, Kovacs S, Njapau H, Kachapulula
PW, Zwaan BJ, van Diepeningen AD and Schoustra SE: Aflatoxigenic
Aspergillus modulates aflatoxin-B1 levels through an
antioxidative mechanism. J Fungi (Basel). 9(690)2023.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Bojić M, Debeljak Ž and Guengerich FP:
Principles of xenobiotic metabolism (biotransformation). In:
Pharmacogenomics in Clinical Practice. Primorac D, Höppner W and
Bach-Rojecky L (eds). Springer International Publishing, Cham,
pp13-33, 2024.
|
|
111
|
Pollet RM, D'Agostino EH, Walton WG, Xu Y,
Little MS, Biernat KA, Pellock SJ, Patterson LM, Creekmore BC,
Isenberg HN, et al: An atlas of β-glucuronidases in the human
intestinal microbiome. Structure. 25:967–977.e5. 2017.PubMed/NCBI View Article : Google Scholar
|
|
112
|
El Asri A, Zarrouq B, El Kinany K,
Bouguenouch L, Ouldim K and El Rhazi K: Associations between
nutritional factors and KRAS mutations in colorectal cancer: A
systematic review. BMC Cancer. 20(696)2020.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Zhang M, Yang D and Gold B: The
adenomatous polyposis coli (APC) mutation spectra in different
anatomical regions of the large intestine in colorectal cancer.
Mutat Res. 810:1–5. 2018.PubMed/NCBI View Article : Google Scholar
|
|
114
|
González-Soltero R, Bailén M, de Lucas B,
Ramírez-Goercke MI, Pareja-Galeano H and Larrosa M: Role of oral
and gut microbiota in dietary nitrate metabolism and its impact on
sports performance. Nutrients. 12(3611)2020.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Vermeer ITM, Gerrits MM, Moonen EJC,
Engels LGJB, Dallinga JW, Kleinjans JCS, van Maanen JMS, Kuipers EJ
and Kusters JG: Helicobacter pylori does not mediate the
formation of carcinogenic N-nitrosamines. Helicobacter. 7:163–169.
2002.
|
|
116
|
Elsanhoty RM, Al-Turki IA and Ramadan MF:
Application of lactic acid bacteria in removing heavy metals and
aflatoxin B1 from contaminated water. Water Sci Technol.
74:625–638. 2016.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Assefa S and Köhler G: Intestinal
microbiome and metal toxicity. Curr Opin Toxicol. 19:21–27.
2020.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Darbandi A, Navidifar T, Koupaei M,
Afifirad R, Nezhad RA, Emamie A, Talebi M and Kakanj M: The effect
of the combination of probiotics and heavy metals from various
aspects in humans: A systematic review of clinical trial studies.
Health Sci Rep. 8(e70521)2025.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Haiser HJ, Gootenberg DB, Chatman K,
Sirasani G, Balskus EP and Turnbaugh PJ: Predicting and
manipulating cardiac drug inactivation by the human gut bacterium
Eggerthella lenta. Science. 341:295–298. 2013.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Ting NLN, Lau HCH and Yu J: Cancer
pharmacomicrobiomics: Targeting microbiota to optimise cancer
therapy outcomes. Gut. 71:1412–1425. 2022.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Heshiki Y, Vazquez-Uribe R, Li J, Ni Y,
Quainoo S, Imamovic L, Li J, Sørensen M, Chow BKC, Weiss GJ, et al:
Predictable modulation of cancer treatment outcomes by the gut
microbiota. Microbiome. 8(28)2020.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Weber R, Fleming V, Hu X, Nagibin V, Groth
C, Altevogt P, Utikal J and Umansky V: Myeloid-derived suppressor
cells hinder the anti-cancer activity of immune checkpoint
inhibitors. Front Immunol. 9(1310)2018.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Niu H, Zhou M, Zogona D, Xing Z, Wu T,
Chen R, Cui D, Liang F and Xu X: Akkermansia muciniphila: A
potential candidate for ameliorating metabolic diseases. Front
Immunol. 15(1370658)2024.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Zhang S, Yang Y, Weng W, Guo B, Cai G, Ma
Y and Cai S: Fusobacterium nucleatum promotes chemoresistance to
5-fluorouracil by upregulation of BIRC3 expression in colorectal
cancer. J Exp Clin Cancer Res. 38(14)2019.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Hussain S: On a new proposed mechanism of
5-fluorouracil-mediated cytotoxicity. Trends Cancer. 6:365–368.
2020.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Vande Voorde J, Vervaeke P, Liekens S and
Balzarini J: Mycoplasma hyorhinis-encoded cytidine deaminase
efficiently inactivates cytosine-based anticancer drugs. FEBS Open
Bio. 5:634–639. 2015.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Ratiner K, Ciocan D, Abdeen SK and Elinav
E: Utilization of the microbiome in personalized medicine. Nat Rev
Microbiol. 22:291–308. 2024.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Nakatsu G, Andreeva N, MacDonald MH and
Garrett WS: Interactions between diet and gut microbiota in cancer.
Nat Microbiol. 9:1644–1654. 2024.PubMed/NCBI View Article : Google Scholar
|
|
129
|
Cani PD, Depommier C, Derrien M, Everard A
and de Vos WM: Akkermansia muciniphila: Paradigm for
next-generation beneficial microorganisms. Nat Rev Gastroenterol
Hepatol. 19:625–637. 2022.
|
|
130
|
Myhrstad MCW, Tunsjø H, Charnock C and
Telle-Hansen VH: Dietary fiber, gut microbiota, and metabolic
regulation-current status in human randomized trials. Nutrients.
12(859)2020.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Yang Y, Xia Y, Chen H, Hong L, Feng J,
Yang J, Yang Z, Shi C, Wu W, Gao R, et al: The effect of
perioperative probiotics treatment for colorectal cancer:
Short-term outcomes of a randomized controlled trial. Oncotarget.
7:8432–8440. 2016.PubMed/NCBI View Article : Google Scholar
|
|
132
|
Miller LE, Zimmermann AK and Ouwehand AC:
Contemporary meta-analysis of short-term probiotic consumption on
gastrointestinal transit. World J Gastroenterol. 22:5122–5131.
2016.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Feng K, Ren F and Wang X: Association
between oral microbiome and seven types of cancers in East Asian
population: A two-sample Mendelian randomization analysis. Front
Mol Biosci. 10(1327893)2023.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Hsieh YY, Tung SY, Pan HY, Yen CW, Xu HW,
Lin YJ, Deng YF, Hsu WT, Wu CS and Li C: Increased abundance of
Clostridium and Fusobacterium in gastric microbiota
of patients with gastric cancer in Taiwan. Sci Rep.
8(158)2018.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Silva-García O, Valdez-Alarcón JJ and
Baizabal-Aguirre VM: Wnt/β-catenin signaling as a molecular target
by pathogenic bacteria. Front Immunol. 10(2135)2019.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Dadgar-Zankbar L, Shariati A,
Bostanghadiri N, Elahi Z, Mirkalantari S, Razavi S, Kamali F and
Darban-Sarokhalil D: Evaluation of enterotoxigenic Bacteroides
fragilis correlation with the expression of cellular signaling
pathway genes in Iranian patients with colorectal cancer. Infect
Agent Cancer. 18(48)2023.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Yu Z, Li Y and Wang J: Mesalazine
suppresses colorectal cancer liver metastasis via the
HDAC3/Wnt/β-catenin axis through downregulating Bacteroides
fragilis abundance. APMIS. 133(e70078)2025.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Grąt M, Wronka KM, Krasnodębski M, Masior
Ł, Lewandowski Z, Kosińska I, Grąt K, Stypułkowski J, Rejowski S,
Wasilewicz M, et al: Profile of gut microbiota associated with the
presence of hepatocellular cancer in patients with liver cirrhosis.
Transplant Proc. 48:1687–1691. 2016.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Ferreira-Halder CV, Faria AVDS and Andrade
SS: Action and function of Faecalibacterium prausnitzii in
health and disease. Best Pract Res Clin Gastroenterol. 31:643–648.
2017.
|
|
140
|
Zhang W, Xu X, Cai L and Cai X: Dysbiosis
of the gut microbiome in elderly patients with hepatocellular
carcinoma. Sci Rep. 13(7797)2023.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Hexun Z, Miyake T, Maekawa T, Mori H,
Yasukawa D, Ohno M, Nishida A, Andoh A and Tani M: High abundance
of Lachnospiraceae in the human gut microbiome is related to high
immunoscores in advanced colorectal cancer. Cancer Immunol
Immunother. 72:315–326. 2023.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Castro PR, Bittencourt LFF, Larochelle S,
Andrade SP, Mackay CR, Slevin M, Moulin VJ and Barcelos LS: GPR43
regulates sodium butyrate-induced angiogenesis and matrix
remodeling. Am J Physiol Heart Circ Physiol. 320:H1066–H1079.
2021.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Tacconi E, Palma G, De Biase D, Luciano A,
Barbieri M, de Nigris F and Bruzzese F: Microbiota effect on
trimethylamine N-oxide production: From cancer to fitness-A
practical preventing recommendation and therapies. Nutrients.
15(563)2023.PubMed/NCBI View Article : Google Scholar
|