Introduction
Ulcerative colitis (UC) refers to prevalent chronic,
non-specific localized inflammation of the intestine, marked by the
presence of local ulcers (1). This
condition can lead to damaged intestinal mucosa, causing symptoms
such as abdominal pain, diarrhea, and bloody or pus-filled stools,
which markedly affect patient health (2). The exact etiology of UC remains
unclear, but involves immune, genetic, environmental and dietary
factors, and treatment strategies typically encompass a combination
of pharmacological interventions, dietary modifications and
psychological support to alleviate symptoms, control disease
progression and enhance the quality of life of patients. Notably,
the incidence of colitis and colitis-related colon cancer has been
increasing in China; despite extensive research into the
influencing factors of UC, the precise pathogenesis remains unclear
(3,4). Advancements in understanding UC
pathogenesis have emphasized the role of dysregulated inflammatory
responses, particularly the extensive tissue damage caused by the
release of pro-inflammatory mediators (5,6).
Sirtuin 1 (SIRT1), a histone deacetylase III widely
present in human cells, has been implicated in regulating various
biological processes, including modulation of the tumor suppressor
p53 and NF-κB through deacetylation, thereby influencing multiple
biological activities (7).
Acetylation of p53 affects its transcriptional activity, and SIRT1
specifically deacetylates the lysine 382 of p53. Beyond
acetylation, histone glycoxidation can induce aggregation and
conformational changes in nuclei that perturb chromatin
architecture and gene regulation, intersecting with SIRT1/NF-κB/p53
crosstalk in inflamed epithelia (8). Moreover, p53 forms a negative
feedback loop by inhibiting SIRT1 transcription through binding to
the p53 response element in the SIRT1 promoter. Increasing evidence
has reported that pro-inflammatory mediators, including TNF-α and
IL-1β, contribute to intestinal inflammation in UC primarily via
NF-κB-mediated pro-inflammatory effects (7,9). Use
of natural products and their derivatives in treating UC is gaining
traction; Donglingcao (Rabdosia rubescens) is commonly used
in traditional Chinese medicine to treat inflammatory conditions,
and is often administered for chronic tonsillitis, pharyngitis,
laryngitis and stomatitis in China (6), Notably, spontaneous recovery from UC
has been reported following Donglingcao administration (10). These phenomena have demonstrated
that Donglingcao may exert anti-UC effects; however, Donglingcao
has not been formally approved for the treatment of UC in China.
Oridonin (Ori), a bioactive tetracyclic diterpenoid compound, is
derived from Donglingcao (6,11),
and our previous study indicated that SIRT1 is a primary target of
Ori, which may mitigate dextran sulfate sodium-induced UC in mice
by activating the SIRT1 signaling pathway (12). Therefore, Ori shows promise as a
future therapeutic agent for UC; however, the molecular mechanisms
underlying its efficacy require further investigation.
The present study aimed to evaluate the SIRT1
signaling pathway in an in vitro model of lipopolysaccharide
(LPS)-treated human intestinal Caco-2 cells, and to evaluate the
safe concentrations for the potential treatment of UC.
LPS-stimulated Caco-2 cells were used to establish a cell model
representing the inflamed human intestinal epithelium;
subsequently, the impact of Ori on the SIRT1/NF-κB/p53 signaling
pathway within the inflamed Caco-2 cells was assessed. Using this
approach, the current study aimed to provide insights into the
potential therapeutic mechanisms of Ori in mitigating inflammation
and preserving intestinal barrier function in vitro.
Materials and methods
Reagents
LPS (batch no. L2630) was purchased from
MilliporeSigma and Ori (batch no. 17073013) was obtained from Baoji
Chenguang Biotechnology Co., Ltd.; the purity of Ori (as determined
by high-performance liquid chromatography) was 98%. Both reagents
were dissolved in sterile distilled water to a concentration of 10
mM and stored at -20˚C for later use. The following antibodies were
purchased from Cell Signaling Technology, Inc. and were used for
western blotting: SIRT1 (1:1,000; cat. no. 8469), NF-κB (1:1,000;
cat. no. 6956), p53 (1:1,000; cat. no. 18032) and GAPDH (1:1,000;
cat. no. 51332); the horseradish peroxidase (HRP)-conjugated
secondary antibody (cat. no. 7074) was purchased from Cell
Signaling Technology, Inc. (1:5,000).
Cell culture and cell viability
assay
The Caco-2 human colon adenocarcinoma cell line was
used in the present study. The cells were cultured in 24-well
plates according to a previously published method (9). Briefly, the fully differentiated
Caco-2 cells were seeded into 96-well plates at a density of 10,000
cells/well and were cultured in DMEM (cat. no. C11995500BT; Gibco;
Thermo Fisher Scientific, Inc.) containing 10% FBS (cat. no.
10091148; Gibco; Thermo Fisher Scientific, Inc.) in an incubator
containing 5% CO2 at 37˚C. After adhering to the plate
for 24 h, the cell culture medium was discarded and fresh DMEM was
supplemented with Ori at distinct concentrations (0.625, 1.25, 2.5,
5.0, 10.0, 20.0, 40.0 and 80.0 µM). After treatment for 24 h at
37˚C, 10 µl Cell Counting Kit (CCK)-8 solution (Sigma-Aldrich;
Merck KGaA) was added to each well and the plate was incubated for
1.5 h at 37˚C. The optical density (OD) was determined at 450 nm
using a microplate reader (model 550; Bio-Rad Laboratories, Inc.).
A total of 6 parallel wells were established at each concentration.
Cell viability calculations were repeated three times.
ELISA
Differentiated, well-grown Caco-2 cells (20 days
post-confluence) in 24-well plates at a density of 10,000
cells/well were randomly divided into four groups: Normal control
(NC), 20 µg/ml LPS, and LPS (20 µg/ml) + high-dose Ori (2.5 µM,
Ori-H) and LPS (20 µg/ml) + low-dose Ori (1.25 µM, Ori-L) groups.
Briefly, both Ori-H and Ori-L groups were incubated with Ori for 1
h prior to stimulation with LPS (20 µg/ml) for 12 h at 37˚C. The NC
group was incubated with 10 µl 0.75% saline, whereas the LPS group
was incubated with LPS (20 µg/ml) for 12 h at 37˚C. At the end of
the experiment, the supernatants were collected by centrifugation
of the cell culture medium at 3,000 x g for 10 min at 4˚C. The
levels of TNF-α and IL-1β in cells were measured by ELISA using
commercially available ELISA kits according to the manufacturer's
instructions. The human TNF-α (cat. no. 430204) and IL-1β (cat. no.
437007) ELISA kits were obtained from BioLegend, Inc.
Western blot analysis
Western blot analysis was performed as previously
described (7). Briefly,
differentiated cells (20 days post-confluence) in 6-well plates
were incubated, grouped and treated with various concentrations of
Ori and LPS as aforementioned; however, they were only stimulated
with LPS for 30 min, not 12 h. Cells were harvested and lysed with
lysis buffer [50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 0.5% NP-40, 5
mM EDTA, 50 mM NaF and 1 mM PMSF; Gibco, Thermo Fisher Scientific,
Inc.]. After complete homogenization, the supernatant was
centrifuged at 10,000 x g for 10 min at 4˚C and collected, and the
protein concentration in the supernatant was determined using the
BCA quantitative method. Subsequently, the supernatant was placed
in boiling water for 10 min to denature the protein and 10 µg
proteins/lane underwent SDS-PAGE on 10% gels and were transferred
to a PVDF membrane. The membrane was blocked with 5% skimmed milk
(Procell Life Science & Technology Co., Ltd.) for 1 h at room
temperature and was then incubated overnight at 4˚C with SIRT1,
NF-κB and p53 antibodies (1:1,000); after which, an appropriate
HRPconjugated secondary antibody was added (1:5,000) and incubated
at room temperature for 2 h. The blots were visualized by enhanced
chemiluminescence imaging (Amersham; Cytiva), and proteins levels
were semi-quantified using ImageJ software (version 1.46; National
Institutes of Health) to analyze the grey value of each protein
band, with GAPDH used as an internal control.
Statistical analysis
All data were statistically analyzed and plotted
using SPSS 20.0 (IBM, Corp.) and GraphPad Prism software 8.0
(Dotmatics). The data are presented as the mean ± standard
deviation. One-way analysis of variance followed by Bonferroni post
hoc analysis was used for data analysis between the groups. All
measurements were performed in triplicate. P<0.05 was considered
to indicate a statistically significant difference.
Results
Effects of Ori on the viability of
Caco-2 cells
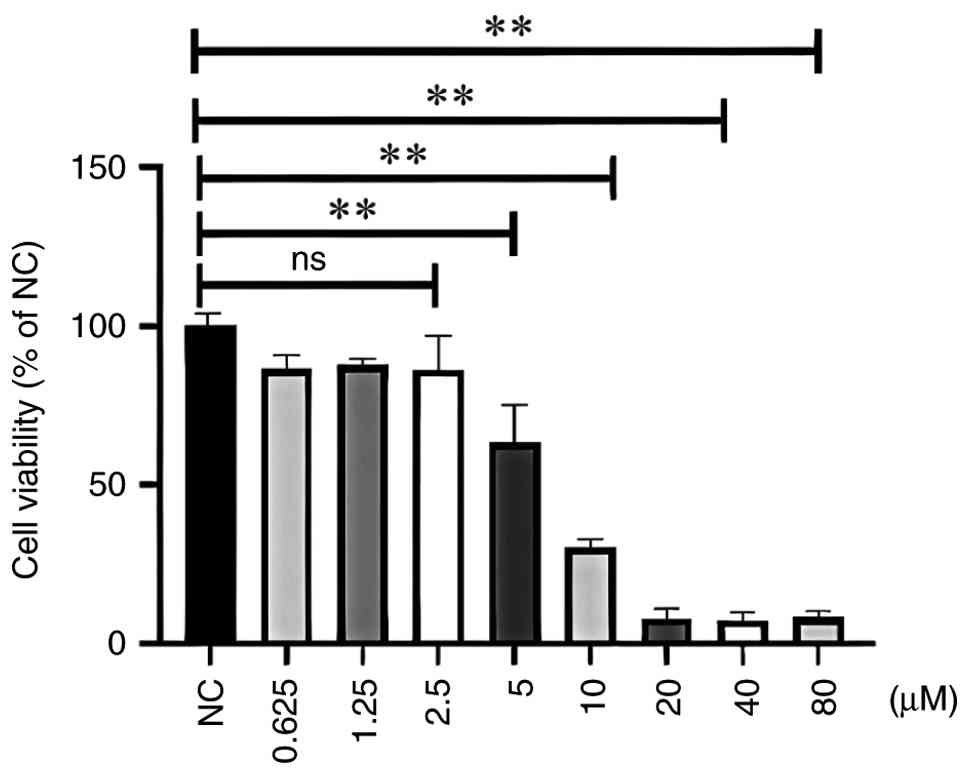
To investigate the effect of Ori on the viability of
Caco-2 cells, cell viability was assessed using the CCK-8 method
after treatment with different concentrations of Ori for 24 h. The
results demonstrated that cell viability was affected by treatment
with Ori at different doses; 5.0-80.0 µM Ori significantly
inhibited the OD values in a dose-dependent manner (P<0.01),
whereas 0.625-2.50 µM Ori did not inhibit viability (P>0.01),
compared with in the control group (Fig. 1). These findings suggested that the
inhibitory effect of Ori on intestinal epithelial cell viability
did not result from a cytotoxic action at doses of 1.25 or 2.50 µM.
Therefore, two concentrations of Ori, 1.25 and 2.50 µM, were
selected for subsequent functional and mechanistic experiments due
to their lack of toxicity. In addition, the IC50 value
of Ori on Caco-2 cells was calculated to be 7.51 µM.
Effects of Ori on LPS-induced
secretion of inflammatory factors in Caco-2 cells
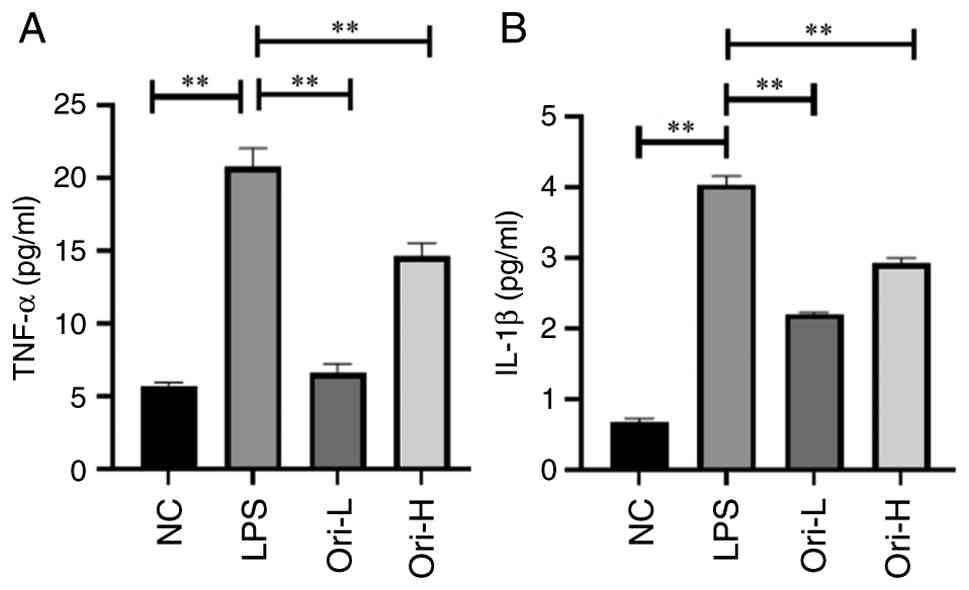
After pre-treatment of Caco-2 cells with Ori for 0.5
h and stimulation with 20 µg/ml LPS for 24 h, it was observed that
LPS stimulation significantly induced the release of TNF-α and
IL-1β in Caco-2 cells, compared with that in the NC group
(P<0.01; Fig. 2). However, Ori
treatment significantly reduced the LPS-induced secretion of TNF-α
and IL-1β in a concentration-dependent manner (P<0.01),
indicating that Ori may suppress the inflammatory response in
intestinal epithelial cells at doses of 1.25 and 2.50 µM.
Effect of Ori on LPS-induced protein
expression in Caco-2 cells
Compared with in the NC group, LPS significantly
decreased the protein expression levels of SIRT1, and increased
those of NF-κB and p53 (P<0.01; Fig. 3). Conversely, Ori significantly
increased the protein expression levels of SIRT1, and decreased
those of NF-κB and p53, compared with in the LPS group (P<0.01;
Fig. 3). These findings indicated
that Ori may reduce the LPS-induced expression of members of the
NF-κB signaling pathway and increase SIRT1 expression, which might
inactivate NF-κB/p53 acetylation and promote SIRT1 transcription
through inhibiting nuclear translocation and binding to the p53
response element in the SIRT1 promoter (9), thereby reducing intestinal
inflammatory response and improving UC.
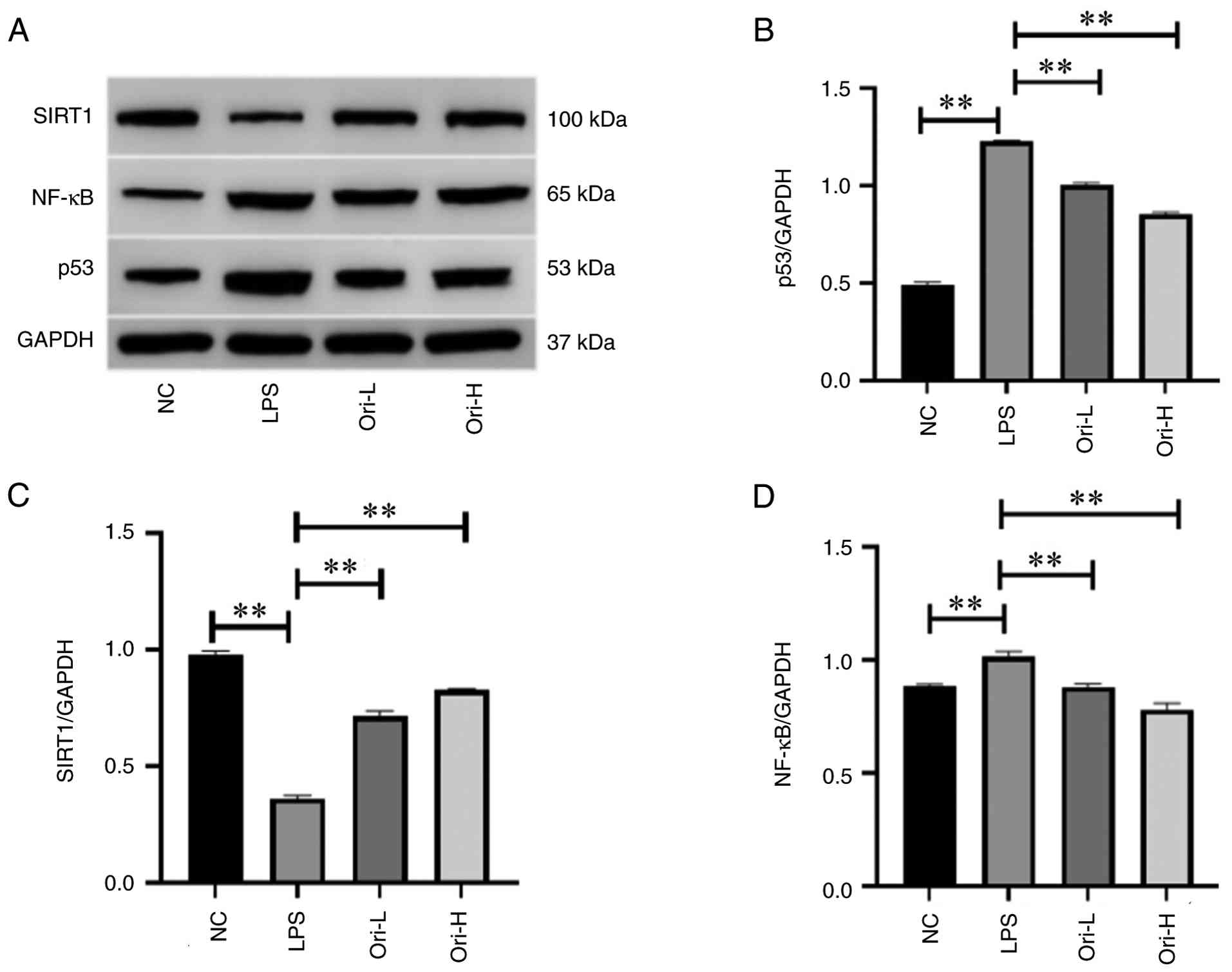 | Figure 3Effect of Ori on the protein
expression levels of SIRT1, NF-κB and p53 in LPSexposed,
differentiated Caco2 cells. (A) SIRT1, NF-κB and p53 were detected
by western blot analysis, with GAPDH as the internal control.
Densitometric analysis of (B) p53/GAPDH, (C) SIRT1/GAPDH, and (D)
NF-κB/GAPDH. **P<0.01. H, high-dose; L, low-dose;
LPS, lipopolysaccharide; NC, normal control; Ori, oridonin. |
Discussion
At present, the cause and onset of UC is not yet
fully understood; however, it is known that it is related to
immunity and is associated with recurrent attacks (4). UC is mainly treated with symptomatic
drugs, such as aminosalicylic acids, corticosteroids,
immunosuppressants and biologics (5); however, while these drugs can
alleviate disease symptoms, they have toxic side-effects (6). Therefore, research into the
pathogenesis and treatment of UC is increasingly important. Studies
have reported that the SIRT1/NF-κB/p53 signaling pathway has marked
effects on inflammation and inflammation related to cancer
development (13,14), and is therefore of interest as a
target to treat UC.
LPS is a notable pathological stimuli of intestinal
epithelial cells, which stimulates macrophages to increase the
secretion of IL-1β and TNF-α. Pro-inflammatory cytokines produced
in the intestine cause damage to intestinal epithelial cells
through various pathways, thus adversely affecting their function
(14). Furthermore, IL-1 increases
intestinal tight junction permeability by activating NF-κB and
myosin light chain kinase, thus decreasing barrier function in
experimental colitis (15). TNF
induces pro-inflammatory processes in the pathogenesis of UC,
including neoangiogenesis and activation of myofibroblasts,
macrophages and T cells; consequently, the release of cytokines is
regarded as a marker of the inflammatory response. Inhibition of
IL-1 and TNF-α function has been shown to ameliorate the
inflammatory activity of experimental colitis in animal models and
reduce inflammation-associated carcinogenesis (16). Therefore, in the current study, LPS
was used to stimulate Caco-2 cells to simulate the inflammatory
environment, with the aim of assessing the effects of Ori on
inflammatory factors in this environment. The results demonstrated
that LPS significantly activated the inflammatory markers TNF-α and
IL-1β in Caco-2 cells compared with that in the NC group. After
treatment with Ori, the levels of these markers were significantly
decreased, indicating that Ori may inhibit the secretion of
pro-inflammatory cytokines induced by LPS stimulation. In addition,
after treatment with Ori, the expression levels of SIRT1 were
increased, whereas those of NF-κB and p53 were decreased, compared
with those in the LPS-treated group. Notably, Ori has garnered
considerable attention from researchers due to its pharmacological
properties, including antibacterial, anti-inflammatory,
pro-apoptotic and antitumor effects (12). Accumulating evidence has
demonstrated that the mechanisms underlying the pharmacological
activities of Ori are primarily mediated through signaling pathways
such as c-Met, Notch and VEGF (15,16).
Consistent with previous findings, studies have reported that Ori
reduces the inflammatory response and inhibits activation of the
key SIRT1 signaling axis, which is associated with inflammation in
LPS-induced cells (13,17).
In the present study, the findings demonstrated that
Ori inhibited LPS-induced inflammatory cytokine response in
intestinal epithelial cells to alleviate the inflammatory response.
In a previous study, Ori has been reported to inhibit the
phosphorylation of SIRT1 and prevent the nuclear translocation of
NF-κB in a mouse model of colitis (13). Similarly, this finding shows that
Ori suppresses the activation of the NF-κB signaling pathway.
Mechanistically, SIRT1-mediated deacetylation notably impacts
multiple biological processes, including cellular senescence,
apoptosis, sugar and lipid metabolism, oxidative stress and
inflammation (18). A recent study
has revealed marked links between SIRT1 and inflammation, in which
alterations to SIRT1 expression and activity have been linked to
inflammatory diseases (19).
Specifically, the present study determined that Ori notably
increased the protein expression levels of SIRT1, and decreased
those of NF-κB and p53 through the NF-κB signaling pathway, which
may reduce the intestinal inflammatory response and promote UC
recovery. Another study showed that Ori can inhibit the
phosphorylation of SIRT1 and prevent the nuclear translocation of
NF-κB in a mouse model of UC (13). As such, SIRT1-targeted
anti-inflammatory therapies are attracting increasing attention for
their clinical applications in treating inflammatory diseases
(7). The present study
investigated the effect of Ori on UC-related inflammation in
LPS-induced Caco-2 cells, an in vitro model of inflamed
human intestinal epithelium. The Caco-2 cell line, derived from a
human colon carcinoma, has been proven to be a good alternative to
an intestinal epithelial cell model, and is the most commonly used
cell line for studies of the structural and functional
characteristics of human differentiated intestinal epithelial cells
(20-22).
Taken together, the present study showed that Ori may be considered
a promising drug for treating UC depending on its suitable
concentration (14,19,20).
However, the present study has some limitations; the
study mainly focused on the in vitro Caco-2 cell model and
therefore does not fully represent the intestinal epithelium in UC.
In addition, another limitation of the present study is that it
contains preliminary research, which did not systematically reveal
if Ori directly modulates SIRT1 activity or whether changes are
secondary to reduced inflammation. Furthermore, the effect of Ori
on cytokine levels in cells that were not treated with LPS was not
explored. In future studies, in vitro and in vivo
experiments will be supplemented using pharmacological inhibitors
or small interfering RNA to confirm the dependence of the effects
of Ori on SIRT1.
In conclusion, to the best of our knowledge, the
present study is the first to provide evidence of the suppressive
effects of Ori on LPS-induced inflammatory responses in intestinal
epithelial cells through inhibition of the SIRT1/NF-κB/p53
signaling pathway, and the increased expression of SIRT1.
Acknowledgements
Not applicable.
Funding
Funding: The present study was financially supported by the
Science and Technology Development Program of Jilin, China (grant
no. YDZJ202301ZYTS122).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
MNW and MCL contributed to the study design. BX, LYL
and YW conducted the investigation. MNW and LYL wrote the original
draft, and MCL reviewed and edited the manuscript. LYL and YW
confirm the authenticity of all the raw data. All authors discussed
the results and agreed to be accountable for all aspects of the
work. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cauchi S, Van Venetien F, Sciberras M and
Ellul P: Colitis trouble up high: a case of gastroduodenal
ulcerative colitis and literature review. J Gastrointestin Liver
Dis. 34:128–132. 2025.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Ruan G, Sun Y, Yu Z, Bai X, Yang H and
Qian J: Global, regional, and national burden of inflammatory bowel
disease from 1990 to 2021: Findings from the global burden of
disease 2021. Gastroenterol Rep (Oxf). 13(goaf082)2025.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Wu Y, He S, Cao M, Teng Y, Li Q, Tan N,
Wang J, Zuo T, Li T, Zheng Y, et al: Comparative analysis of cancer
statistics in China and the United States in 2024. Chin Med J
(Engl). 137:3093–3100. 2024.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Harpaz N and Polydorides AD: Upper
gastrointestinal manifestations of inflammatory bowel disease. Surg
Pathol Clin. 13:413–430. 2020.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Katsandegwaza B, Horsnell W and Smith K:
Inflammatory bowel disease:A review of pre-clinical murine models
of human disease. Int J Mol Sci. 23(9344)2022.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Chen B, Dong X, Zhang JL, Sun X, Zhou L,
Zhao K, Deng H and Sun Z: Natural compounds target programmed cell
death (PCD) signaling mechanism to treat ulcerative colitis: A
review. Front Pharmaco. l15(1333657)2024.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Shen P, Deng X, Chen Z, Ba X, Qin K, Huang
Y, Huang Y, Li T, Yan J and Tu S: SIRT1: A potential therapeutic
target in autoimmune diseases. Front Immunol.
12(779177)2021.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Mir AR, Moinuddin and Islam S:
Circulating autoantibodies in cancer patients have high specificity
for glycoxidation modified histone H2A. Clin Chim Acta. 453:48–55.
2016.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Jia X, Liu H, Ren X, Li P, Song R and Li
X, Guo Y and Li X: Nucleolar protein NOC4L inhibits tumorigenesis
and progression by attenuating SIRT1-mediated p53 deacetylation.
Oncogene. 41:4474–4484. 2022.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Barjasteh AH, Al-Asady AM, Latifi H, Al
Okla S, Al-Nazwani N, Avan A, Khazaei M, Ryzhikov M, Nadi-Yazdi H
and Hassanian SM: Maximizing treatment options for IBD through drug
repurposing. Curr Pharm Des. 30:2538–2549. 2024.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Zhang Y, Wang S, Dai M, Nai J, Zhu L and
Sheng H: Solubility and bioavailability enhancement of oridonin: A
review. Molecules. 25(332)2020.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Li X, Zhang C, Ma W, Xie X and Huang Q:
Oridonin: A review of its pharmacology, pharmacokinetics and
toxicity. Front Pharmacol. 12(645824)2021.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Wang M, Xu B, Liu L and Wang D: Oridonin
attenuates dextran sulfate sodium-induced ulcerative colitis in
mice via the Sirt1/NF-κB/p53 pathway. Mol Med Rep. 26(312)2022.
|
|
14
|
Tu J, Xu Y, Xu J, Ling Y and Cai Y:
Chitosan nanoparticles reduce LPS-induced inflammatory reaction via
inhibition of NF-κB pathway in Caco-2 cells. Int J Biol Macromol.
86:848–856. 2016.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Chen M, Chen C, Gao Y, Li D, Huang D, Chen
Z, Zhao X, Huang Q, Wu D, Lai T, et al: Bergenin-activated SIRT1
inhibits TNF-α-induced proinflammatory response by blocking the
NF-κB signaling pathway. Pulm Pharmacol Ther.
62(101921)2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Ban J, Peng X, Zhang Y, Liu Y, Wei Y, Ao
L, Tian H, He X, Zhao H and Li J: Oridonin alleviates SiNPs-induced
pulmonary fibrosis by inhibiting pyroptosis via IRE1α-XBP1s-NLRP3
pathway. Int Immunopharmacol. 164(115388)2025.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Shao YY, Guo Y, Feng XJ, Liu JJ, Chang ZP,
Deng GF, Xu D, Gao JP and Hou RG: Oridonin attenuates TNBS-induced
post-inflammatory irritable bowel syndrome via PXR/NF-κB signaling.
Inflammation. 44:645–658. 2021.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Yang Y, Liu Y, Wang Y, Chao Y, Zhang J,
Jia Y, Tie J and Hu D: Regulation of SIRT1 and its roles in
inflammation. Front Immunol. 13(831168)2022.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Jia XM, Hao H, Zhang Q, Yang MX, Wang N,
Sun SL, Yang ZN, Jin YR, Wang J and Du YF: The bioavailability
enhancement and insight into the action mechanism of poorly soluble
natural compounds from co-crystals preparation: Oridonin as an
example. Phytomedicine. 122(155179)2024.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Zhao G, Zhang T, Ma X, Jiang K, Wu H, Qiu
C, Guo M and Deng G: Oridonin attenuates the release of
pro-inflammatory cytokines in lipopolysaccharide-induced RAW264.7
cells and acute lung injury. Oncotarget. 8:68153–68164.
2017.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Artursson P, Palm K and Luthman K: Caco-2
monolayers in experimental and theoretical predictions of drug
transport. Adv Drug Deliv Rev. 46:27–43. 2001.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Sambuy Y, De Angelis I, Ranaldi G, Scarino
ML, Stammati A and Zucco F: The Caco-2 cell line as a model of the
intestinal barrier: Influence of cell and culture related factors
on Caco-2 cell functional characteristics. Cell Biol Toxicol.
21:1–26. 2005.PubMed/NCBI View Article : Google Scholar
|