1. Introduction
Food is essential for survival and maintaining the
overall well-being of all living organisms, and the method of food
consumption has evolved throughout history. In order to survive,
the diet of an individual requires a balance of both nutritious and
antinutritional factors. The diet provides nutrients that serve as
raw materials for metabolic processes (1,2), and
health is greatly influenced by genetics, diet, lifestyle and
nutrition. The gastrointestinal system of the human body processes
carbohydrates, dietary fats and proteins that are consumed into
monosaccharides, fatty acids and amino acids, respectively, fueling
organ function and supporting cellular growth (3,4).
In the 21st century, factors such as climate change,
water scarcity, rising food prices and population growth challenge
agriculture and food security, particularly in arid and semi-arid
regions. Scientists and nutritionists need to explore alternative
food sources to address hunger and poverty. Cereal grains are the
global primary food source and are central to the human diet
(5).
Carbohydrates are a major macronutrient, vital for
energy production and organ function. The majority of carbohydrates
are biodegradable and biocompatible, thus rendering them suitable
for both drug delivery and functional foods. Nanostructured
carbohydrates, with self-assembling abilities, provide diverse
applications in food and therapeutics. Different saccharide
structures enable various self-assembly methods, thereby expanding
their functions in food (6).
Carbohydrates are unique among macronutrients in not having a
minimum dietary requirement.
Increase in global health risks, such as type 2
diabetes, obesity, non-alcoholic fatty liver disease (NAFLD) and
other cardiovascular health issues, have compelled the scientific
community to re-evaluate carbohydrate intake (7). Large-scale epidemiological studies
and controlled human trials are increasingly demonstrating that,
rather than quantity alone, carbohydrate quality may be the cause
of health risks (8). Rapidly
digestible starches and heavily processed foods containing refined
sugar increase the risk of developing hyperinsulinemia, hepatic
de novo lipogenesis (DNL) and postprandial glycemic
excursions, whereas fiber-rich sugars reduce the risks of
developing these disorders and support metabolic flexibility
(9-11).
This has opened views on carbohydrate toxicity, which depicts
excessive intake of certain digestible carbohydrates, particularly
fructose-rich sugars, as a metabolic stressor capable of disrupting
hepatic and systemic homeostasis (12,13).
Non-digestible carbohydrates, such as resistant starches and
oligosaccharides selectively nourish beneficial gut microbes,
leading to the production of bioactive metabolites, such as
short-chain fatty acids, that help regulate glucose and support
immune function (14,15).
Carbohydrates also contribute greatly to medicine
development. Both natural and synthetic sugar-based polymers and
glycoconjugates exhibit excellent biocompatibility, molecular and
biological recognition, and structural tunability, facilitating a
wide range of applications in biological evaluations, such as
anticancer and antiviral therapies (16). Technical utilities, such as
automated glycan assembly, have further improved the access to
complex sugar structure (17),
speeding up translational research connecting chemistry, biology
and medicine.
The present narrative review presents a
comprehensive perspective on carbohydrates, focusing on their
physicochemical characteristics, nutritional values and chemical
and metabolic functions. Furthermore, contemporary topics, such as
carbotoxicity and fructose-induced lipogenesis are also discussed.
Unlike the highly specialized reviews, the present review aimed to
connect carbohydrate chemistry with food science and nutritional
values, providing an interdisciplinary overview relevant to
students, researchers, health professionals and nutritionists.
2. Physicochemical properties of
carbohydrates and their biological significance in food
matrices
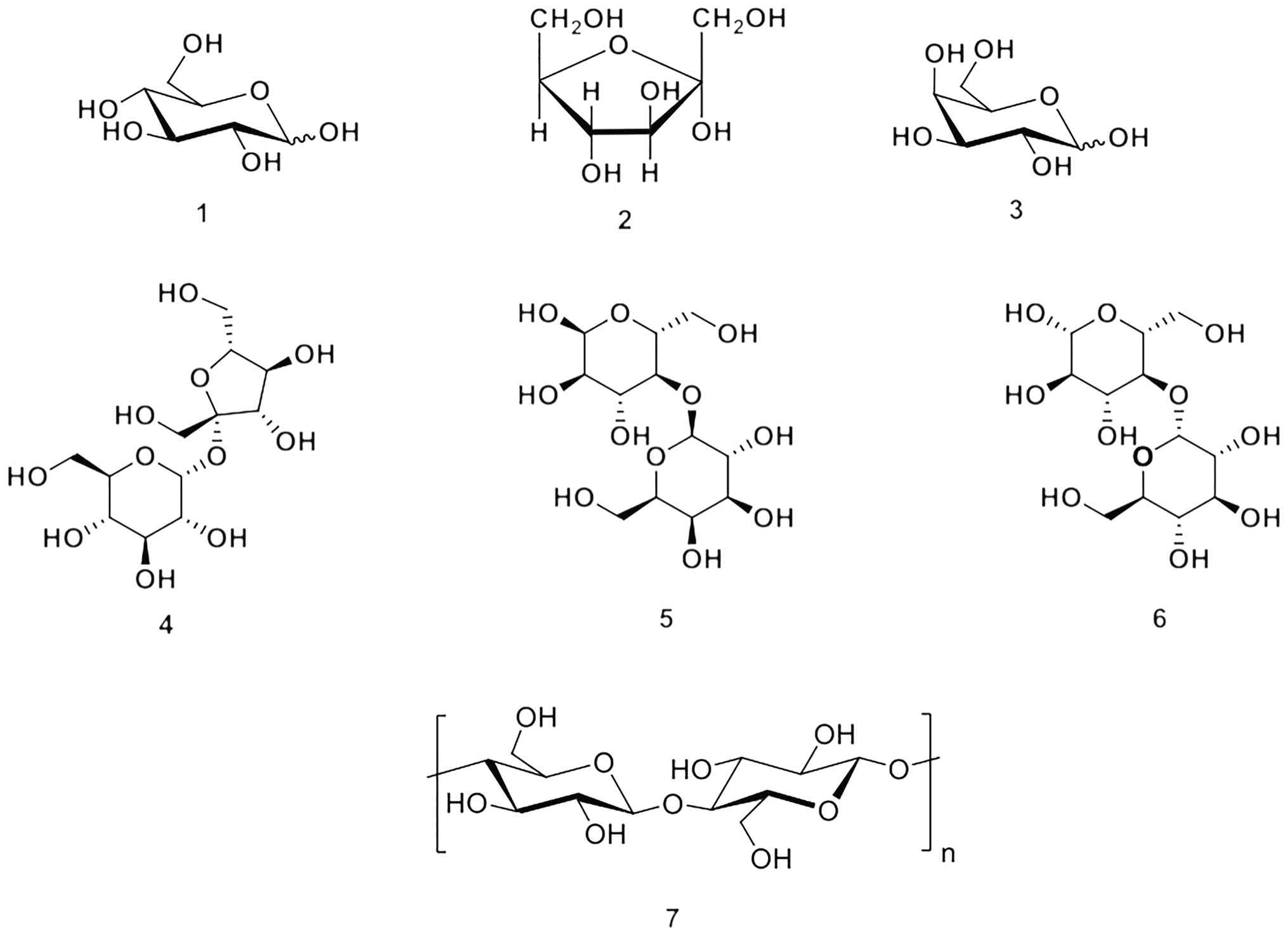
The structural hierarchy and complex structural
features of carbohydrates influence their nutritional values to a
great extent. Monosaccharides, such as glucose and fructose, are
the simplest monomeric forms of sugar. Through the formation of
glycosidic bonds, these units assemble into disaccharides (for
example, sucrose) and eventually into polysaccharides with vast,
complex chains that define the texture and energy profile of the
human diet. A few common examples of carbohydrates are illustrated
in Fig. 1 (18,19).
Furthermore, polysaccharides are utilized in various functions,
including energy storage in plants (primarily starch) and as
structural support for plant cells (primarily cellulose). They
function as a gelling component of the intercellular matrix to help
hold ions, such as sodium, calcium and magnesium. Polysaccharides,
such as cellulose, starch, chitin and seaweed polysaccharides have
high commercial value across various industries, from paper
manufacturing to food processing (20).
The physicochemical properties of carbohydrates are
fundamental factors that affect their functions in food matrices
and biological evaluation. These properties, ranging from
solubility and hygroscopicity to gelation capacity and reactivity,
arise directly from molecular structure, including monomer
composition, glycosidic linkage stereochemistry (α vs. β),
branching density and molecular weight (21). Such structural features influence
not only the texture, stability and sensory properties of foods,
but also the rate and extent of their digestion, thus affecting
glycemic response and interactions with the gut microbiota
(22).
Solubility and hygroscopicity are vital properties
influencing food quality and shelf life. Monosaccharides, such as
glucose and fructose exhibit high water solubility due to their
multiple hydroxyl groups, thereby reducing the reactivity of water
with food and inhibiting microbial growth (23), which is utilized in confectionery
and baked goods to maintain softness and moisture. However,
polysaccharides, such as cellulose are insoluble due to β(1→4)
linkages that result in extensive intermolecular hydrogen bonding,
which contributes to dietary fiber content and textural bulk
without significantly affecting water activity (24). The solubility of carbohydrates also
affects phase behavior in complex matrices; for instance, during
the preparation of gummy sweets, incompatibility between gelatin
and long-chain polysaccharides in glucose syrups can lead to phase
separation, which is modulated by moisture content and the presence
of small-molecule additives, such as citric acid or allulose
(25).
Carbohydrate physicochemistry is also controlled by
viscosity and gelation. Starch, composed of amylose [linear glucan
connected through α(1→4)-linkage] and amylopectin [branched with
α(1→6) linkages], undergoes gelation upon heating in aqueous media,
leading to swelling of the system, loss of crystallinity and
formation of a viscoelastic network (26). The ratio of amylose to amylopectin
critically influences the gel strength and retrogradation kinetics;
a higher amylose content improves gelation but also increases the
risk of staling in bakery products (27). Similarly, pectin, a
heteropolysaccharide rich in galacturonic acid, forms gels in
acidic, high-sugar environments through charge neutralization and
water loss, a mechanism essential for jam and jelly preparation.
Recent research has demonstrated that processing technologies,
including ultrasound-assisted heat-moisture treatment, can modify
starch crystallinity and reduce rapidly digestible starch content,
thereby lowering glycemic potential while preserving functional
properties (28).
Carbohydrate reactivity, particularly through the
Maillard reaction, markedly affects both food quality and
nutritional outcomes. This non-enzymatic browning reaction occurs
between reducing sugars (e.g., glucose, fructose) and the amino
groups of proteins at elevated temperatures and in the presence of
moisture, generating flavor compounds, brown pigments and advanced
glycation end-products (29).
While it improves the flavor in roasted coffee or seared meat,
etc., uncontrolled Maillard reactions in processed foods can
produce potentially harmful compounds and decrease protein
bioavailability (30). On the
contrary, controlled Maillard reactions between proteins and
prebiotic fibers can enhance emulsifying properties, solubility and
antioxidant activity, thereby enhancing the functionality of
plant-based products (31).
Non-reducing sugars such as sucrose do not participate in the
Maillard reaction if not hydrolyzed first to glucose and fructose,
emphasizing how glycosidic bond affects reactivity.
The biological significance of these physicochemical
traits becomes evident in their modulation of carbohydrate
bioavailability and gut health. Rapidly digestible starches, often
resulting from extensive gelatinization and low amylose content,
elicit sharp postprandial glucose spikes, which can contribute to
metabolic disorders when consumed chronically (24). By contrast, resistant starches and
non-digestible oligosaccharides, characterized by β-linkages or
complex branching that are inaccessible to human amylases, reach
the colon intact, where they serve as prebiotics, selectively
stimulating beneficial bacteria such as Lactobacilli and
Bifidobacteria (32,33).
The fermentation of these fibers produces short-chain fatty acids
(SCFAs), such as butyrate, which nourish colonocytes, regulate
immune function and improve insulin sensitivity (34). The efficacy of prebiotic action is
highly dependent on physicochemical parameters: Molecular weight,
solubility and glycosidic bond type determine fermentability rates
and microbial selectivity (35).
Moreover, the food matrix itself modulates
carbohydrate bioaccessibility. Encapsulation within cell walls (as
in whole grains) or complexation with proteins and lipids can
attenuate enzymatic hydrolysis, reducing glycemic impact (36). Processing methods, both thermal
(e.g., baking and extrusion) and non-thermal (e.g., high-pressure
processing), can disrupt these matrices, increasing starch
digestibility, or they can promote retrogradation and resistant
starch formation, depending on conditions (37). Thus, the interplay between
intrinsic carbohydrate properties and extrinsic matrix factors
ultimately shapes nutritional outcomes.
The physicochemical properties of carbohydrates are
not only technical aspects for food formulation but are crucial to
their biological outcomes. These properties influence texture and
stability in food products, regulate glycemic response and modulate
the gut microbiota, thereby connecting food science and human
physiology (9,11). The strategic manipulation of
carbohydrate structure through breeding, enzymatic modification, or
customized processing provides promising opportunities to create
foods that meet sensory expectations while enhancing metabolic
health (38).
3. Nutritional aspects of carbohydrates in
the human diet
Dietary sources and
bioavailability
The bioavailability of carbohydrates, the rate and
extent to which they are absorbed and utilized, is intrinsically
linked to their structural hierarchy and the food matrix in which
they reside (9). Dietary
carbohydrates range from simple monosaccharides to complex
polysaccharides. Monosaccharides, such as glucose and fructose are
present in their free form in honey, fruits and corn syrups,
providing immediate bioavailability. Disaccharides, such as the
lactose found in mammalian milk or the sucrose concentrated in
sugar beets and cane, require minimal enzymatic cleavage prior to
absorption (39). While
animal-derived foods are generally carbohydrate-poor, dairy
products are a unique exception, providing the essential
disaccharide lactose, which accounts for almost 40% of the energy
in human breast milk and is vital for neonatal development
(40,41).
Contemporary nutritional science emphasizes that the
health impact of these sugars is heavily dictated by their source.
For instance, while refined sucrose and fructose in high-fructose
corn syrup are associated with rapid glycemic spikes, the same
sugars found within the fibrous matrix of whole fruits such as
grapes or kiwi exhibit different kinetic profiles (42,43).
As the structural complexity of polysaccharides, such as the
starches found in legumes, tubers and cereal grains, increases, the
pace of digestion typically slows. However, modern processing
techniques have challenged the traditional dichotomy between simple
and complex. Current evidence indicates that highly processed
complex carbohydrates, such as white bread or instant rice, can
have a glycemic index (GI) equal to or higher than that of simple
table sugar, leading to significant insulin demand and increasing
the risk of metabolic syndromes, obesity and ovulatory infertility
(44). The role of carbohydrates
in the diet is presented in Table
I (41,44-61).
Table I provides an overview of
dietary carbohydrates, which are vital macronutrients; their types,
quantity and metabolic fate critically influence health
outcomes.
 | Table ISource of various carbohydrates and
diseases related to excess intake. |
Table I
Source of various carbohydrates and
diseases related to excess intake.
| Category of
carbohydrates | Type of
carbohydrate | Sweetness (relative
to sucrose=1) | Food source | Daily amount | Amount of energy
(caloric value kcal/g) | Activity on the
body parts | Carbohydrate
toxicity or diseases | (Refs.) |
|---|
|
Monosaccharides | Glucose | 0.75 | Dates, currants,
figs, grapes, dried apricot, sweet corn, and honey | 25 g | 4 | Blood | Hyperglycemia,
diabetes | (44-48) |
| | fructose | 1.74 | Honey, dates, fruit
jams, grapes, apples, pear, kiwi and banana | ≤50 g | 4 | Liver | non-alcoholic fatty
liver disease (NAFLD) | (45,47,49-51) |
| | Galactose | 0.32 | Flavored yogurts,
lactose free milk, ground black peppers | 5-20 g | 4 | ovaries | Ovarian cancer | (44,47,52) |
| Disaccharides | Sucrose | 1 | Cakes, cookies, and
dark chocolate, beetroot, carrots, sugar cane and sugar beet | 20-32 g | 4 | Heart, breast | heart disease,
breast cancer | (47,53-55) |
| | lactose | 0.2 | Milk, buttermilk,
yogurt, sour cream, condensed milk, Frozen, cottage cheese,
evaporated milk, goats milk and ice creams | 12 g | 4 | intestine | Lactose
intolerance | (47,54,56,41) |
| | maltose | 0.4 | Malted wheat and
barley, Breads, bagels, Malt extract, molasses Beer | 75 mg | 4 | Digestive
system | Chronic
diarrhea | (47,54,55,4) |
|
Oligosaccharide | Fructooligosacch
aride | 0.3-0.6 | Legumes, cabbage,
broccoli, Onion, fennel, peas, wheat, bananas, asparagus,
garlic | 10-2 0 g | 4 | NA | Not specific
toxicity | (47,57,58) |
| | Galactooligosacch
arides | 0.3-0.6 | Cow milk, yogurt,
beans. | 12-15 g | 4 | NA | NA | (47,59) |
|
Polysaccharides | Starch | Not sweet | Oats, rice,
chickpeas, potato, pasta, corn, flours. | NA | 4 | NA | NA | (47) |
| | Fiber | Not sweet | Cereal grains,
legumes, nuts, seeds. | 28-36 g | 2 | NA | NA | (60,61) |
The interaction between dietary carbohydrates, the
gut microbiome, and host health constitutes a dynamic and
reciprocal relationship that is fundamental to modern nutritional
frameworks. Dietary fibers and other indigestible carbohydrates
serve as pivotal environmental factors influencing the composition
and metabolic output of the gut microbial ecosystem (62). Subsequently, the gut microbiota
converts these dietary elements into bioactive metabolites that
affect host metabolism, immune function and even neurological
processes (63). This relationship
is increasingly recognized in the prevention and management of
chronic diseases, including obesity, cardiometabolic disorders and
certain types of cancer (62). The
notion of carbohydrate quality currently includes not only its
glycemic effects, but also its capacity to support a beneficial gut
microbiota, thereby enhancing overall health (9,11).
This integrated viewpoint emphasizes that the simplistic perception
of carbohydrates as mere caloric sources is outdated; they should
be regarded as information-rich molecules whose chemical language
is deciphered by both human enzymes and the trillions of microbial
entities in the gut, ultimately defining their significant
biological roles in food and nutrition sciences (34,64).
Role as a primary energy
substrate
Carbohydrates are the main source of oxidative fuel
for the human body, providing with ~4 kcal/g (17 kJ/g) of energy.
Glucose is particularly critical as it is the only metabolic
substrate for the central nervous system and red blood cells, which
lack the enzymes to efficiently oxidize other fuels under normal
physiological conditions (65,66).
In the fed state, dietary carbohydrates maintain blood glucose
homeostasis and replenish glycogen stores in the liver and skeletal
muscle. Under restricted exogenous intake, the body demonstrates
metabolic flexibility by activating gluconeogenesis and ketogenesis
to maintain brain function. This change indicates that the quality
of carbohydrates is critical for overall health. A balanced intake
(usually 45-65% of daily calories) maintains a stable metabolism;
however, consuming high-GI foods continuously break down these
homeostatic mechanisms (67-70).
Non-digestible polysaccharides, or dietary fiber,
have become an integral part of modern preventive medicine. Unlike
starch, which is broken down by enzymes in the small intestine,
fibers such as cellulose and hemicellulose remain whole until they
reach the colon. There, they add bulk to the stool and serve as
fermentable substrates for the gut microbiota (71). The SCFAs produced by this process
serve as an alternative energy source for colonocytes and exert
systemic anti-inflammatory effects (72). Current clinical guidelines suggest
that individuals should consume ~30 g of fiber per day, which is
linked to lower rates of coronary heart disease, type 2 diabetes
and colorectal cancer (73). This
indicates that carbohydrate chemistry can affect long-term health
outcomes.
Despite the extensive understanding of carbohydrate
metabolism, a significant knowledge gap persists regarding
inter-individual variability in glycemic response. While tools such
as the GI and glycemic load provide a standardized framework for
ranking foods, contemporary big data nutrition studies (74) have revealed that two individuals
consuming the identical carbohydrate source can exhibit vastly
different postprandial glucose excursions. This variability is
deemed to be driven by a complex interplay between host genetics,
sleep patterns, physical activity, and most crucially the unique
composition of the gut microbiome of an individual (74,75).
Furthermore, a vigorous scientific debate remains
unresolved concerning the carbohydrate-insulin model (CIM) of
obesity. Proponents of the CIM argue that high-carbohydrate diets
drive obesity by promoting insulin secretion, which partitions
energy toward fat storage rather than oxidation, whereas critics
maintain that total energy balance (calories in vs. calories out)
remains the primary driver (76,77).
Resolving these questions is essential for moving beyond
one-size-fits-all dietary guidelines and toward personalized
nutritional interventions that can effectively combat the global
rise in metabolic disease.
Authoritative international dietary
guidelines and the disadvantages of the overconsumption of refined
carbohydrates
Authoritative international dietary guidelines
consistently identify refined carbohydrates as a key dietary
component to limit due to their well-established association with
chronic disease risk. The World Health Organization (WHO), in its
global recommendations, explicitly advises that the intake of free
sugars, which encompass added sugars and those naturally present in
honey, syrups and fruit juices, should be limited to <10% of
total energy intake for both adults and children, with a further
conditional recommendation to reduce this to <5% for additional
health benefits (78,79). This guidance is grounded in
extensive epidemiological evidence linking a high sugar consumption
to weight gain, dental caries and metabolic dysfunction. Similarly,
the Dietary Guidelines for Americans (DGA), jointly issued by the
US Department of Health and Human Services and the US Department of
Agriculture, emphasize minimizing the intake of refined grains and
added sugars, recommending that <10% of daily calories should be
derived from added sugars and that at least half of all grain
consumption should be whole grains (80). The European Food Safety Authority
(EFSA) and national bodies such as the UK Scientific Advisory
Committee on Nutrition (SACN) echo these principles, advocating for
a shift from refined to whole-grain carbohydrate sources to improve
long-term health outcomes (81).
The scientific foundation for these guidelines rests
on a robust body of epidemiological research. The landmark
Prospective Urban Rural Epidemiology (PURE) study, which followed
>135,000 individuals across 18 countries, provided critical
population-level evidence that high carbohydrate intake,
particularly from refined sources, is associated with an increased
total mortality (82). This
finding challenged older paradigms that focused primarily on fat
reduction and highlighted the importance of carbohydrate quality
over mere quantity. Further analysis from the PURE cohort
specifically linked the higher consumption of refined grains
(median intake of 350 g/day in the highest quintile) to a
significantly elevated risk of developing major cardiovascular
events and total mortality. By contrast, whole grain intake
exhibited neutral or protective associations (83). These results underscore the
differential health impacts of carbohydrate subtypes, a nuance
increasingly emphasized in modern nutritional science.
Longitudinal cohort studies from the USA have
provided granular insights into specific refined carbohydrate
sources. Data from the Nurses' Health Study and the Health
Professionals Follow-Up Study (84) demonstrate a clear dose-response
association between the consumption of sugar-sweetened beverages
(SSBs) and the incidence of type 2 diabetes and cardiovascular
disease. A meta-analysis of prospective cohort studies confirmed
that each daily serving of SSBs is associated with an 18% higher
risk of coronary heart disease (85). Similarly, a previous systematic
review and meta-analysis found that the higher intake of refined
carbohydrates predicts a significantly increased risk of developing
type 2 diabetes, with the association persisting even after
adjustment for body mass index and other confounders (86). The mechanisms underlying these
associations are multifactorial: Refined carbohydrates drive rapid
postprandial glycemia and hyperinsulinemia, promote hepatic DNL
leading to dyslipidemia (elevated triglycerides and reduced
high-density lipoprotein cholesterol) and induce systemic
inflammation, all key pathways in the development of
cardiometabolic diseases (87).
The concept of ultra-processed foods (UPFs), as
defined by the NOVA classification system, has further refined the
understanding of the risks associated with the consumption of
refined carbohydrates. UPFs are industrial formulations that
typically contain high levels of refined starches, added sugars and
unhealthy fats, while being low in fiber and phytonutrients. A
recent umbrella review of epidemiological meta-analyses concluded
that a higher consumption of UPFs is consistently associated with
increased risks of obesity, type 2 diabetes, cardiovascular disease
and all-cause mortality (88). The
Framingham Offspring Study reported that for every 10% increase in
the proportion of UPFs in the diet, there was a 12% higher risk of
overall cardiovascular disease and a 13% higher risk of
cerebrovascular disease (89).
These findings suggest that the health risks of consuming refined
carbohydrates may be amplified in the context of ultra-processing,
where the food matrix is degraded and hyper-palatable combinations
override natural satiety signals, leading to excess energy intake
and metabolic stress (87).
Despite this strong consensus, some nuances and
ongoing debates exist. For instance, not all refined carbohydrate
sources carry equal risk; the consumption of white rice in Asian
populations exhibits complex associations that may be modified by
the overall dietary pattern and glycemic load (83). Furthermore, the focus is shifting
from isolated nutrients to holistic dietary patterns. The
Mediterranean diet, rich in whole grains, legumes, fruits and
vegetables, has been shown in randomized trials, such as the
PREDIMED study, to significantly reduce cardiovascular events,
demonstrating that the adverse effects of the consumption of
refined carbohydrates can be mitigated by replacing them with
high-quality carbohydrate sources within a balanced dietary pattern
(90). Overall, authoritative
guidelines and contemporary epidemiological evidence converge on a
clear message that reducing the intake of refined carbohydrates,
particularly in the form of added sugars and UPFs, is a critical
public health strategy for preventing chronic diseases
globally.
4. Metabolic processes and bioenergetics
involving carbohydrates
The transition from the structural complexity of
dietary carbohydrates to systemic energy requires a highly
coordinated sequence of enzymatic transformations. For the human
body, the critical connection between food and health is realized
through these metabolic pathways, which determine whether a
carbohydrate is partitioned toward immediate adenosine triphosphate
(ATP) production, biosynthetic precursors, or long-term energy
storage. Cells use different enzymatic activities in different
phases to convert ingested glucose into carbon dioxide and water.
Two metabolic pathways, glycolysis and the tricarboxylic acid (TCA)
cycle, also known as the Krebs or citric acid cycle, are involved
in the breakdown of glucose (91).
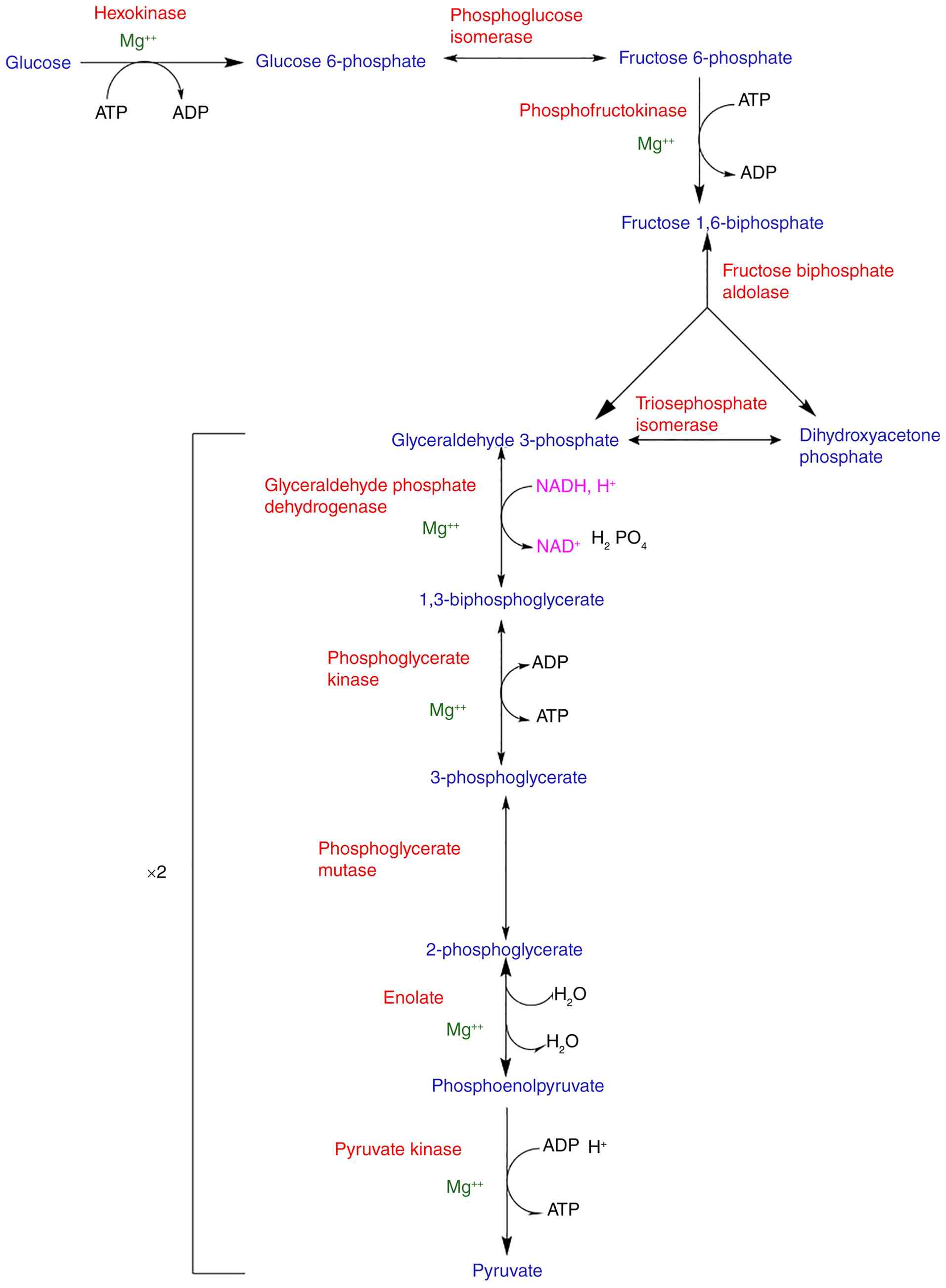
Glycolysis
Glycolysis is a catalytic process that converts
glucose into pyruvate via 10 enzyme steps. These highly regulated
processes of glycolysis have two stages: The energy input for Step
1 requires two ATPs per glucose molecule, followed by the energy
release as four ATPs, two for each glyceraldehyde molecule, which
is described as Step 2(90).
Glycolysis, a process in living cells, can also be used in the
citric acid cycle or fermentation, and is the primary source of
acetyl coenzyme A (acetyl-CoA), the primary energy source. The
structures of glycolysis intermediates are depicted in Fig. 2 (92).
The initial stage of glycolysis requires energy in
the form of ATP. ATP phosphorylates D-glucose at the six carbons
via the enzyme hexokinase to produce D-glucose-6-phosphate (G-6-P).
This regulatory process is inhibited by the presence of
glucose-6-phosphate. Phosphoglucoisomerase (class: Isomerase) then
converts it to D-fructose-6-phosphate (F-6-P), which is
phosphorylated again by ATP, this time at the one-carbon position,
by the enzyme phosphofructokinase (class: Transferase) to produce
D-fructose-1,6-bisphosphate. Owing to its high G value, this is the
committed stage of glycolysis. The enzyme, fructose bisphosphate
aldolase, then cleaves D-fructose-1,6-bisphosphate into two
three-carbon molecules, dihydroxyacetone phosphate (DHAP) and
D-glyceraldehyde-3-phosphate (G-3-P). DHAP is transformed into
G-3-P as the following step in glycolysis requires the molecule
G-3-P by the enzyme triose phosphate isomerase (93).
The second stage involves producing four ATP
molecules for each glucose molecule. The glyceraldehyde-3-phosphate
dehydrogenase enzyme phosphorylates G-3-P at the one carbon to
produce the high-energy molecule, 1,3-bisphosphoglycerate (BPG).
Phosphoglycerate kinase then phosphorylates adenosine diphosphate
(ADP) at the expense of BPG to produce ATP and 3-phosphoglycerate.
Phosphoglycerate mutase then converts 3-phosphoglycerate to
2-phosphoglycerate to produce another high-energy molecule. Enolase
then converts 2-phosphoglycerate to phosphoenolpyruvate, releasing
water, potassium and manganese ions. Pyruvate kinase phosphorylates
ADP to produce ATP and pyruvate, and cellular energy controls this
reaction. High energy levels indicate inhibited pyruvate kinase.
Stage 2 occurs twice due to glucose division (93).
In parallel, G-6-P can be diverted into the hexose
monophosphate shunt (pentose phosphate pathway). Unlike glycolysis,
this pathway is not primarily concerned with ATP production.
Instead, it serves two vital biosynthetic roles: Generating
nicotinamide adenine dinucleotide phosphate, which provides
reducing power for fatty acid synthesis and antioxidant defense
(reducing glutathione), and producing ribose-5-phosphate, a
precursor for nucleotide synthesis (94). In the context of nutrition, the
balance between these two pathways is essential; for instance, a
diet chronically high in refined sugars can over-activate these
shunt pathways, potentially fueling DNL and contributing to
metabolic dysfunction (95).
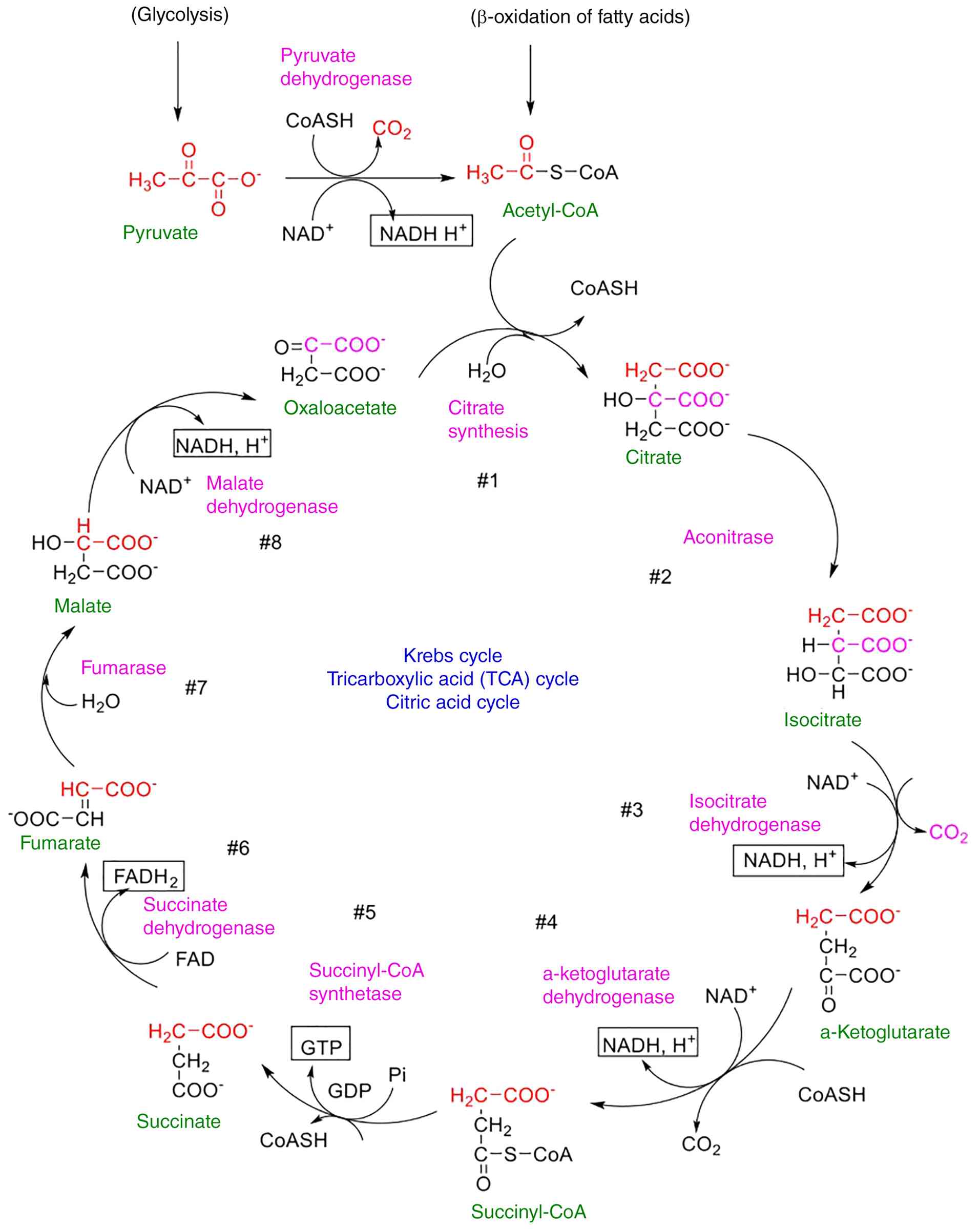
Krebs cycle
The citric acid cycle is the ultimate oxidative
process in which carbs, lipids and amino acids interact. It is the
most significant metabolic pathway for the energy source of the
body. TCA is the most essential core metabolic pathway, connecting
nearly all metabolic processes. The Krebs cycle (Fig. 3) is an aerobic biodegradation
process that begins with acetyl-CoA and ends with CO2.
Patients with diabetes have lower oxaloacetic acid and Krebs cycle
activity due to reduced glucose and pyruvate levels (96).
At higher energy levels, the citrate synthesis rate
increases, while the flux through the Krebs cycle decreases. This
excess citrate is subsequently transported to the cytosol and
degraded to acetyl-CoA. Acetyl-CoA can then be used to produce
fatty acids and cholesterol. Citrate has a positive regulatory
effect on lipogenic enzymes (e.g., acetyl-CoA carboxylase) and a
negative regulatory effect on phosphofructokinase-1 (a glycolytic
enzyme). Citrate is also necessary for ketogenesis in non-hepatic
tissues (96).
The α-ketoglutarate dehydrogenase complex converts
α-KG to succinyl coenzyme A (succinyl-CoA) through multiple steps
(Fig. 3). Succinyl-CoA is a highly
energetic molecule that generates the only guanosine triphosphate
molecule in this pathway when succinate thiokinase is present
(96). Nicotinamide adenine
dinucleotide (NADH2) and flavin adenine dinucleotide
(FADH2) are the energy molecules created during this
oxidative decarboxylation cycle. These molecules release energy
during the oxidative phosphorylation reaction; thus, their power is
retained in ATP as high-energy phosphate (97). Contemporary biochemical evidence
suggests that the total aerobic yield from a single glucose
molecule is ~30-32 ATP. This efficiency highlights why
carbohydrates remain the preferred primary energy substrate of the
human body. However, this high-efficiency system is also sensitive
to nutrient overload. When carbohydrate intake exceeds the capacity
for oxidation and glycogen storage, the excess acetyl-CoA is
redirected toward triglyceride synthesis, reinforcing the link
between high-glycemic diets and adipose tissue expansion (98).
Energy balance and glycemic
response
One glucose molecule is transformed into 38 ATP
molecules during cellular respiration. One glucose molecule is
broken up to produce two molecules of pyruvate, producing eight ATP
directly from anaerobic glycolysis. Each three-carbon pyruvate
yields one NADH2 (equivalent to three ATP) during the
pyruvate decarboxylation reaction. As a result, each six-carbon
glucose molecule in this process generates six ATP. The total
energy produced by each three-carbon pyruvate in the Krebs cycle is
12 ATP (91).
This quantity is generated by isocitrate
dehydrogenase (+NADH2 3ATP), alpha-ketoglutarate
dehydrogenase (+NADH2=3ATP), malate dehydrogenase
(+NADH2=3 ATP), succinate dehydrogenase
(+FADH2=2 ATP) and succinate thiokinase (+GTP). As each
six-carbon glucose molecule generates two three-carbon pyruvate
molecules, the total quantity produced by one glucose molecule is
24 ATP. The glycolysis 8 ATP, the pyruvate dehydrogenase complex 6
ATP, and the Krebs cycle 24 ATP combine to yield 38 ATP (91).
The systemic health impact of carbohydrate
metabolism is best observed through the glycemic response, the
postprandial rise and fall of blood glucose. This response is
governed by a delicate hormonal ‘tug-of-war’ between insulin and
glucagon (99).
High-bioavailability carbohydrates (simple sugars and refined
starches) elicit a rapid insulin spike, which promotes glucose
uptake via GLUT4 transporters in muscle and adipose tissue, while
simultaneously suppressing lipolysis (99,100).
The connection here involves metabolic flexibility.
In a healthy individual, the body can switch seamlessly between
carbohydrate oxidation in the fed state and fat oxidation in the
fasted state. However, a diet dominated by high-GI foods can lead
to chronic hyperinsulinemia and eventual insulin resistance. This
state represents a breakdown in systemic energy balance: Cells
become starved of glucose despite elevated blood glucose levels,
leading to increased hunger and a cycle of metabolic stress
(44). Understanding these
pathways emphasizes that the nutritional value of a carbohydrate is
defined not only by its caloric content, but by its kinetic
interaction with the body's metabolic machinery.
Divergent hepatic kinetics: Fructose
vs. glucose metabolism
While glucose and fructose are structural isomers
sharing the same caloric density, their metabolic destinies in the
human body are fundamentally different due to divergent hepatic
processing (101). Glucose
metabolism is systemic; although the liver is a primary site for
its regulation, glucose is utilized by nearly every cell in the
body, and its entry into glycolysis is strictly governed by the
energy status of the cell. By contrast, fructose is almost
exclusively metabolized by the liver, and its entry into central
metabolic pathways bypasses the most critical regulatory
checkpoints (101,102).
The primary kinetic distinction lies in the first
steps of phosphorylation. Glucose is converted to G-6-P by
hexokinase or glucokinase. Fructose, however, is phosphorylated by
fructokinase (ketohexokinase or KHK) to form fructose-1-phosphate
(F-1-P). This enzyme is not inhibited by ATP or citrate.
Furthermore, F-1-P is cleaved by aldolase B into DHAP and
glyceraldehyde. These triose phosphates enter the glycolytic
sequence downstream of the phosphofructokinase-1 (PFK-1) regulatory
step (103,104). Essentially, the excess of
fructose in the metabolic machinery provides a continuous supply of
carbon precursors, such as acetyl-CoA, regardless of the actual
energy requirements of the cell (105). Additionally, the subsequent
conversion of fructose-6-phosphate to fructose-1,6-bisphosphate by
PFK-1 is the rate-limiting step of glycolysis. When cellular ATP
levels are high, PFK-1 is inhibited, decelerating glucose oxidation
to prevent an unnecessary surplus of metabolic intermediates
(103).
This unregulated flux has profound implications for
metabolic health. As the liver is suddenly presented with an
abundance of acetyl-CoA that it cannot oxidize in the TCA cycle, it
must divert these carbons toward DNL. This leads to the synthesis
of palmitate and other fatty acids, contributing to the
accumulation of intrahepatic fat (non-alcoholic fatty liver
disease, NAFLD) and the secretion of very-low-density lipoprotein,
which raises blood triglyceride levels (106).
Furthermore, the rapid, uncontrolled phosphorylation
of fructose by KHK leads to transient intracellular ATP depletion.
As ATP is rapidly consumed to phosphorylate fructose without the
brake of feedback inhibition, the resulting rise in AMP triggers
the purine degradation pathway, leading to the production of uric
acid. Elevated uric acid is not only a risk factor for gout, but
also functions as a pro-oxidant within the hepatocyte, potentially
inducing mitochondrial oxidative stress and further impairing
insulin sensitivity (107,108).
This kinetic reality explains why high-fructose diets, particularly
those rich in refined syrups, are uniquely linked to metabolic
syndrome, independent of total caloric intake.
Role of resistant starches and
oligosaccharides in shaping the gut microbiota and the risk of
metabolic disease
Resistant starches and oligosaccharides represent
two critical classes of non-digestible carbohydrates that exert
profound influence on gut microbial ecology and, consequently, on
the risk of developing metabolic diseases, such as obesity, type 2
diabetes and cardiovascular disorders. Their biological
significance stems not from caloric contribution, since they
largely escape digestion in the small intestine, but from their
role as selective substrates for beneficial gut commensals, thereby
modulating microbial composition, metabolic output and host
physiology (15).
Resistant starch is a form of starch that resists
enzymatic hydrolysis by human amylases and reaches the colon
intact, where it serves as a fermentable substrate for specific
members of the gut microbiota. Primary degraders of resistant
starch include Ruminococcus bromii and Bifidobacterium
adolescentis, with emerging evidence identifying additional
specialists such as Ruminococcoides bili (109). These bacteria initiate the
breakdown of complex resistant starch granules through specialized
carbohydrate-active enzymes (CAZymes), enabling secondary
fermenters to access the resulting oligosaccharides and
monosaccharides. This hierarchical degradation process fosters
cross-feeding networks, thereby enhancing microbial diversity and
stability. The fermentation of resistant starch predominantly
yields SCFAs, notably butyrate, acetate and propionate, which serve
as key mediators of metabolic health. Butyrate, in particular, is
the preferred energy source for colonocytes, strengthens the gut
barrier by upregulating tight junction proteins, and exerts
anti-inflammatory effects by inhibiting histone deacetylases and
activating G-protein-coupled receptors (e.g., GPR109A). Clinical
and preclinical studies consistently link higher a intake of
resistant starch with improved insulin sensitivity, reduced
adiposity and lower systemic inflammation, all of which are
protective against cardiometabolic diseases (24,62).
Oligosaccharides, such as fructooligosaccharides
(FOS), galactooligosaccharides and human milk oligosaccharides,
function similarly as prebiotics by selectively stimulating the
growth and activity of beneficial bacteria, especially
Bifidobacteria and certain Lactobacilli. Their
structural complexity, defined by glycosidic linkage types [e.g.,
β(2→1) in inulin-type FOS] and the degree of polymerization,
determines their resistance to host enzymes and specificity for
microbial utilization. Research profiling the metabolic capacities
of 17 gut commensal strains has revealed marked strain-specific
differences in oligosaccharide fermentation, underscoring that
prebiotic efficacy is contingent on both substrate structure and
the recipient's microbial repertoire (110). This individualized response
explains the variable outcomes observed in dietary interventions
and highlights the need for personalized nutrition approaches. The
SCFAs produced from oligosaccharide fermentation regulate host
metabolism through multiple pathways: propionate reduces hepatic
lipogenesis and gluconeogenesis, while acetate influences appetite
regulation via central nervous system signaling (62). Moreover, oligosaccharides can
directly inhibit pathogen adhesion by acting as decoy receptors,
thereby enhancing gut defense mechanisms.
The interplay between these carbohydrates and the
gut microbiome has significant implications for the prevention of
chronic disease. Dietary fibers, including RS and oligosaccharides,
are associated with a reduced risk of obesity, type 2 diabetes and
types of certain cancer, largely through microbiota-dependent
mechanisms (62,111). For instance, low-carbohydrate
diets that inadvertently reduce fiber intake may diminish SCFA
production, potentially compromising gut barrier integrity and
promoting endotoxemia, a known driver of insulin resistance
(111). Conversely, the strategic
inclusion of high-quality, fermentable carbohydrates supports a
resilient microbial ecosystem that buffers against dysbiosis
induced by Western diets high in refined sugars and saturated fats
(112). Notably, the concept of
‘carbohydrate quality’ now supersedes mere quantity; slowly
digestible or non-digestible forms such as resistant starch and
oligosaccharides are favored for their ability to modulate
postprandial glycemia and sustain microbial health without
contributing to hyperinsulinemia or DNL (24,112). Hence, resistant starches and
oligosaccharides function as keystone dietary components that shape
a health-promoting gut microbiota, drive beneficial metabolic
signaling via SCFAs, and mitigate risk factors for major chronic
diseases, positioning them at the nexus of food science, microbiome
research and precision nutrition.
Despite these advances however, challenges remain in
translating findings into universal dietary recommendations due to
inter-individual variability in gut microbiota composition, host
genetics and baseline metabolic status. Furthermore, the structural
heterogeneity of dietary carbohydrates complicates analytical
characterization and standardization. Emerging glycomic platforms
combining chromatography and mass spectrometry are beginning to
address these gaps by enabling precise mapping of carbohydrate
structures to microbial responses (34).
5. The global intake trends of dietary
fiber
In the 1970s, Burkitt's research (113,114) led to a resurgence of scientific
interest in dietary fiber (97).
Subsequent research has demonstrated a connection between the
consumption of dietary fiber, body weight, metabolic processes and
overall health. The consumption of dietary fiber is associated with
a favorable body weight, a healthy gut microbiota and a lower risk
of cardiovascular disease and mortality. It also enhances
intestinal health and lowers the riss of developing cancer. The
primary health benefits of dietary fiber and the risks of
deficiency are discussed in this synopsis, with particular
attention to the effects of abdominal obesity and overall metabolic
health (61).
Global dietary fiber intake trends reveal a
persistent and concerning gap between recommended levels and actual
consumption across most populations worldwide. Current evidence
indicates that median intake values typically range from 15 to 20 g
per day, substantially below the widely accepted recommendations of
25-38 g per day for adults (8,115).
This deficit is particularly pronounced in high- and middle-income
countries where dietary patterns have markedly shifted toward
energy-dense, nutrient-poor options. Recent global macrotrend
analyses demonstrated that while some middle-income countries have
experienced modest increases in fiber consumption over the past
three decades, these improvements are often offset by concurrent
rises in sodium intake and the consumption of UPFs (116). The Swiss National Nutrition
Survey provides a compelling case study, demonstrating that a
higher consumption of UPFs is significantly associated with a lower
dietary fiber intake, highlighting the inverse association between
industrial food processing and fiber adequacy (117). Similarly, data from the USA
spanning 1999=2017 reveal only minimal increases in fiber
consumption, with persistent racial disparities; African Americans
consistently exhibit a lower fiber intake compared to other
demographic groups, potentially contributing to elevated colorectal
cancer rates in this population (118). In Chile, national surveys
indicate that fiber intake remains suboptimal despite public health
initiatives, with primary sources being traditional staples rather
than diverse plant-based foods (119,120).
For example, according to a previous analysis,
adults in the USA and Europe ingest 18-24 g of dietary fiber daily
for men and 16-20 g for women, with grain products as the main
source. This is approximately one-third of what is advised; the
majority of Western populations wish to increase their intake by
~50%. According to the NHANES (National Health and Nutrition
Examination Survey), dietary fiber intake was higher in European
nations than in North American nations (97).
An emerging scientific debate centers on the
differential physiological effects of various fiber types and
sources, challenging the traditional view of fiber as a homogeneous
nutrient category. Contemporary research increasingly distinguishes
between isolated and intrinsic fiber, fermentable and
non-fermentable varieties, and the impact of fiber across different
dietary contexts, including low-carbohydrate and ultra-processed
food frameworks. The controversy extends to regulatory definitions,
particularly regarding low-molecular-weight carbohydrates, such as
oligosaccharides and inulin, which are accepted as dietary fiber in
many jurisdictions, but remain optional in the Codex Alimentarius
definition, creating inconsistencies in nutritional labeling and
research interpretation (121).
This debate is further complicated by the recognition that the
health benefits of fiber are not merely mechanical, but are
profoundly mediated through interactions with the gut microbiome.
Different fiber types serve as selective substrates for specific
microbial taxa, leading to varied production profiles of SCFAs,
such as acetate, propionate and butyrate, which function as
critical signaling molecules influencing immune regulation,
metabolic homeostasis and even neurocognitive function (62,122,123).
Contemporary evidence from large-scale prospective
cohort studies and comprehensive meta-analyses robustly supports
the protective role of higher total fiber intake against major
chronic diseases. The landmark Lancet systematic review established
that every 8-g increase in daily fiber consumption is associated
with a 5-27% reduction in mortality and the incidence of coronary
heart disease, stroke, type 2 diabetes and colorectal cancer
(8). This finding has been
reinforced by more recent umbrella reviews encompassing millions of
individuals, which confirm strong, credible associations between
adequate fiber intake and reduced risk across diverse health
outcomes (115). The mechanisms
underlying these benefits extend beyond the traditional
understanding of the role of fiber in gastrointestinal health to
include modulation of the gut-brain axis, the enhancement of gut
barrier integrity and the epigenetic regulation of immune function
through SCFA-mediated histone deacetylase inhibition (123-125).
Furthermore, fiber consumption has demonstrated specific protective
effects in vulnerable populations, including the reduced risk of
surgery for inflammatory bowel disease (126), improved metabolic parameters in
obesity (127,128) and improved glycemic control in
diabetes (129,130).
Despite this substantial body of evidence, several
critical scientific questions remain unresolved. First, the causal
vs. the associative nature of fiber-microbiome-host interactions
requires further elucidation through well-designed intervention
trials that account for interindividual variability in baseline
microbiota composition and host genetics, such as the FUT2 secretor
status, which influences microbial niche availability (62,122). Second, optimal dose-response
associations for specific fiber types and distinct health endpoints
have not been definitively established, rendering precise dietary
recommendations challenging (115). Third, the relative efficacy of
whole food vs. supplements or fortified foods in promoting
long-term adherence and sustained health benefits remains unclear,
particularly given the complex food matrix effects that may enhance
fiber bioactivity (131,132). Fourth, the impact of modern food
processing methods on fiber structure and functionality requires
systematic investigation, as industrial processing may alter the
physicochemical properties of fiber and, consequently, its
biological effects (133,134). Finally, the development of
personalized nutrition approaches necessitates a better
understanding of how individual factors, including age, sex,
metabolic status and existing disease conditions, influence
responses to different fiber interventions, which could
revolutionize dietary guidance from population-based to
precision-based recommendations (135).
6. Medicinal aspects of carbohydrates
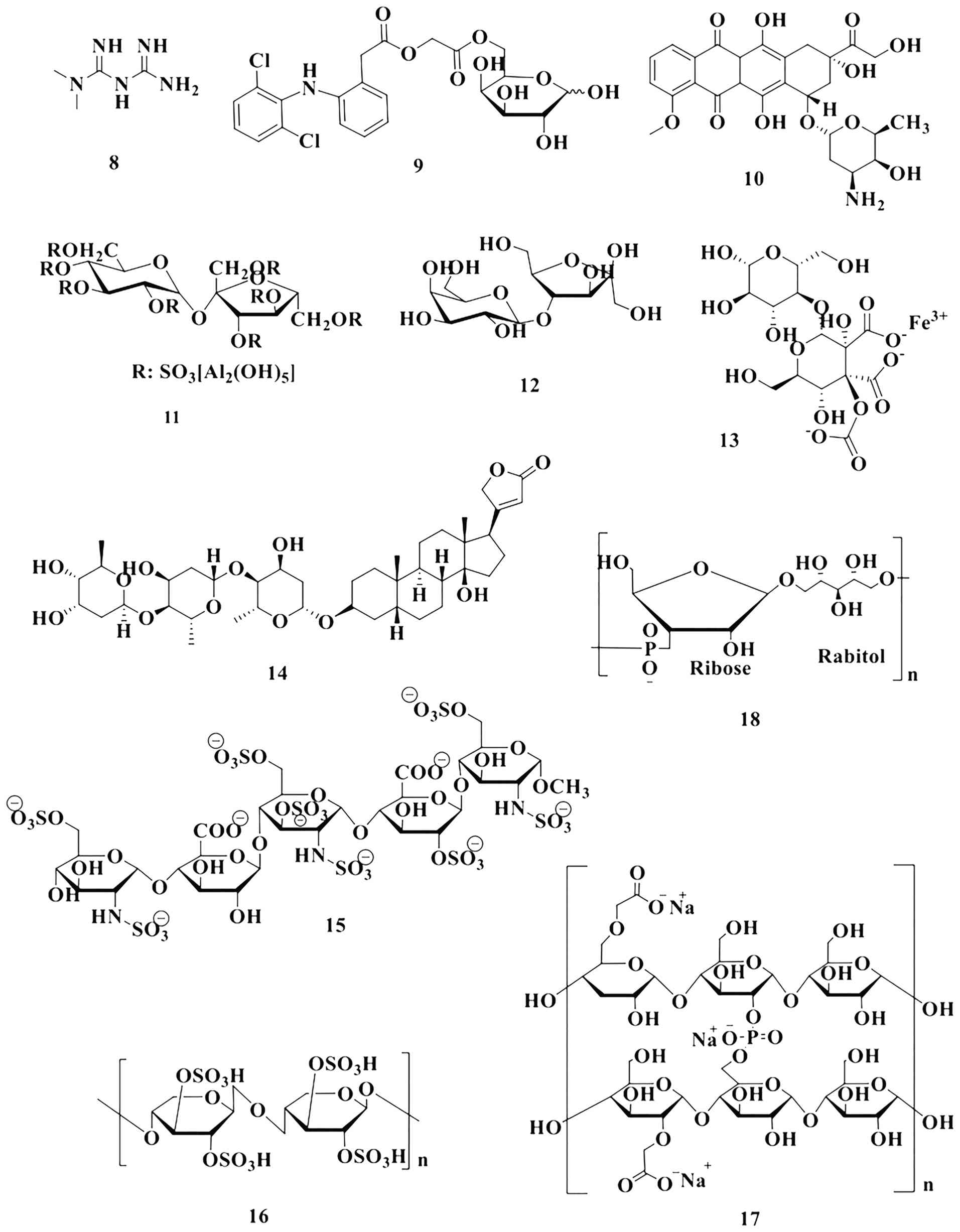
Among the bioactive glycosides that have been
naturally extracted or derived, O- and N-glycosides
are abundant, while C-glycosides (both naturally occurring
and laboratory synthesized) make more stable bio-mimics than native
O-glycosides as they are not prone to catalytic or enzymatic
cleavage (136). These widespread
classes of biomolecules play vital roles in immune system function
and the stimulation of the immune response, which have yet to be
properly explored by researchers. Several such carbohydrate-based
drugs developed with potential biological activities are
illustrated in Fig. 4 (137,138). Additionally, they have inherent
chiral complexity, high bioavailability and excellent
biocompatibility, rendering this class of compounds an excellent
option for the development of prodrugs (139). Depending on the evidence, some
carbohydrate-based medicines are given in Table II (140-157).
It underscores the versatility of carbohydrates in drug design,
delivery and therapeutic function, while emphasizing the importance
of dosage control to minimize adverse effects.
| Table IIMedicines derived from carbohydrate
sources and their functions. |
Table II
Medicines derived from carbohydrate
sources and their functions.
| Category of
carbohydrates | Types of
carbohydrate | Medicine derived
from the carbohydrate (Fig.
4)a | Activity on the
body parts | Function of the
medicine | Diseases caused by
overdosage of the medicine (side-effects) | (Refs.) |
|---|
|
Monosaccharides | Glucose | Metformin (no.
8) | Liver | To balance the
production of glucose in the liver. | Induces
gastrointestinal disturbance, lactic acidosis, hepatotoxicity,
vitamin B12 deficiency, acute pancreatitis, hypoglycemia | (140,141) |
| | Galactose | ACEgal
anti-inflammatory drug (aceclofenac with D-galactose) (no. 9) | Inflammatory
tissues | Aceclofenac works
by blocking the enzyme called cyclooxygenase (COX) which generates
prostaglandins that causes pain, fever and inflammation | NA | (142) |
| |
3-amino-2,3,6-trideoxy-L-lyxo-hexopyranose | Doxorubicin (no.
10) | Cancer cells | It regulates the
growth of cancer cells by intercalating into DNA and inhibits the
topoisomerase II and further it blocks DNA and RNA synthesis. | Cardiotoxicity,
neuropathy, hepatotoxicity, nephrotoxicity, alopecia, typhlitis,
myelosuppression, neutropenia, anemia, thrombocytopenia, nausea,
and diarrhea | (143,144) |
| Disaccharides | Sucrose | Sucralfate (no.
11) | gastrointestina l
(GI) tract | It stimulates the
healing of duodenal ulcers by reducing pepsin activity in gastric
juice in the stomach. | Constipation,
aluminium toxicity, dry mouth & skin, headaches, nausea,
vomiting, flatulence, low phosphate levels (hypophosphatemia) | (143,145) |
| | lactose | Lactulose (no.
12) | colon | It enhances the
growth of beneficial bacteria such as Lactobacilli. It draws
the water into colon and reduces constipation. It promotes gut
health. | Lactulose affects
the intestine, plasma metabolomes, and nitrogen metabolism. | (146,147) |
| | maltose | Ferric carboxymalt
ose (FCM)- Medication (no. 13) | Bone marrow | It promotes ferric
iron into the bloodstream then utilized for the production of red
blood cells in the bone marrow and it will be stored in the liver
and spleen. | Acute renal
failure, headache and aseptic meningitis, blood clots | (148,149) |
| Trisaccharides | A trisaccharide
moiety attached to a steroidal aglycone | Digoxin (no.
14) | Heart | It inhibits
sodium-potassium ATPase which leads to the increase of
intracellular concentration of sodium and calcium. This leads to a
chain of biochemical reactions that show multiple effects on
cardiac muscle and the cardiovascular system. | Irregular heart
rhythms, visual disturbances, confusion, dizziness, nausea and
vomiting | (143,150) |
|
Oligosaccharides | Heparin-derived
pentasaccharide | Fondaparinu
x-Injection (prefilled syringe) (no. 15) | Blood | It stimulates the
action of antithrombin III against Factor Xa and it prevents blood
clotting. | Headache and
gastrointestinal disorders, including diarrhea, nausea and
vomiting. | (151,152) |
| | Multiple
β-D-xylopyranose units are chemically sulfated and linked
together | Pentosan
polysulfate | Blood (no. 16) | It works on the
blood where it functions as a heparin-like anticoagulant &
reduces the risk of blood clotting and intensifies the process of
fibrinolysis. | Headache,
Nausea, | (143,153) |
|
Polysaccharides | Starch | Sodium starch
glycolate (SSG)- pharmaceutic al excipient (no. 17) | Stomach and small
intestine. | It induces the
effectiveness of medicines and by speeding up the process of tablet
disintegration and drug release. | NA | (154) |
| | Ribosylribitol
phosphate-carbohydrate-containing medication | Haemophilus
influenzae type b (Hib) Vaccine (no. 18) | Immune cells in the
bloodstream and lymphoid tissues. | It is a preventive
vaccine protect the infections of the blood. | Redness, swelling
or pain at the injection site, Low-grade fever, Fatigue. | (155-157) |
Natural sources
Carbohydrates have not only been a major source of
food and nutrition but also have excellent pharmaceutical
applications. Carbohydrates derived from natural sources have been
studied as therapeutic agents for a number of years. Among all
carbohydrate-based drugs developed between 2000 and 2021, the
majority have been derived from naturally occurring carbohydrate
moieties, of which 37% are nucleosides. Furthermore, 60% of all
carbohydrate-based drugs have broad applications in the preparation
of antiviral, antibacterial/antiparasitic, anticancer and
antidiabetic drugs (156). For
instance, saponins, a diverse family of glycosides having brilliant
adjuvant activity, such as Quillaja brasiliensis saponins
(triterpene glycosides). Its source is the Chilean tree Quillaja
saponaria bark. It has been reported to have >20
water-soluble Quillaja saponaria saponins, from which
several led to the eliciting of humoral and cell-mediated responses
(158). Similarly, in Asian
countries, Coriolus versicolor polysaccharides
(polysaccharides Krestin and Lentinan) have been used for >40
years as antitumor adjuvants (159). Pectin, a family of
polysaccharides, is a fine source of dietary fiber. It has been
reported to exhibit inhibitory activity against various cancer cell
lines (160-164).
Synthetic carbohydrate-based
materials
Structures of naturally occurring carbohydrates are
often very complex due to their complex biosynthetic pathway.
Hence, isolating structurally defined pure compounds from natural
sources becomes cumbersome. To better address demands in
glycobiology, synthetic carbohydrate derivatives or glycoconjugates
comprise a fascinating alternative. Synthetic or non-natural
glycoconjugates can aid in understanding structure-activity
associations in biological responses and mimic the functions of
native saccharide derivatives (165).
Some of the advantages of these synthetic
carbohydrate-based materials stem from their biocompatibility and
intrinsic biological functionality, resulting in generally low
toxicity and excellent biodegradability, which are crucial for
clinical translation (166).
Their most significant asset is their ability to engage in
specific, high-affinity interactions with carbohydrate-binding
proteins (lectins, selectins and siglecs) that are central to
numerous disease processes, including cancer, inflammation and
infection This allows for the rational design of therapeutics that
can modulate immune responses; for instance, synthetic
glycopolymers can be engineered to mimic the multivalent
presentation of glycans on cell surfaces, amplifying binding
avidity through the ‘glycocluster effect’ to either activate or
suppress immune cells for applications in tumor immunotherapy
(167,168). Furthermore, carbohydrate
polymers, such as chitosan, alginate and hyaluronic acid serve as
excellent scaffolds for targeted drug delivery, particularly for
colon-specific release, as they can be degraded by colonic
microflora enzymes, thereby minimizing systemic side-effects
(169,170). The structural diversity of the
sugar scaffold also provides multiple points for chemical
modification, enabling fine-tuning of pharmacokinetic properties
such as solubility and half-life (171).
Automated glycan assembly (AGA) has expedited access
to synthetic glycans up to 50-mers (172), while other automated platforms
based on electrochemical assembly (173), fluorous-assisted solution-phase
(174) and HPLC-assisted
synthesis (175) have been
limited to a few examples not exceeding hexasaccharides (176). From the proof-of-concept using a
modified peptide synthesizer in 2001 to the first commercial
Glyconeer 2.1 synthesizer (177),
AGA has been developed using glycans of mammalian, bacterial and
plant origin as a precursor (178,179).
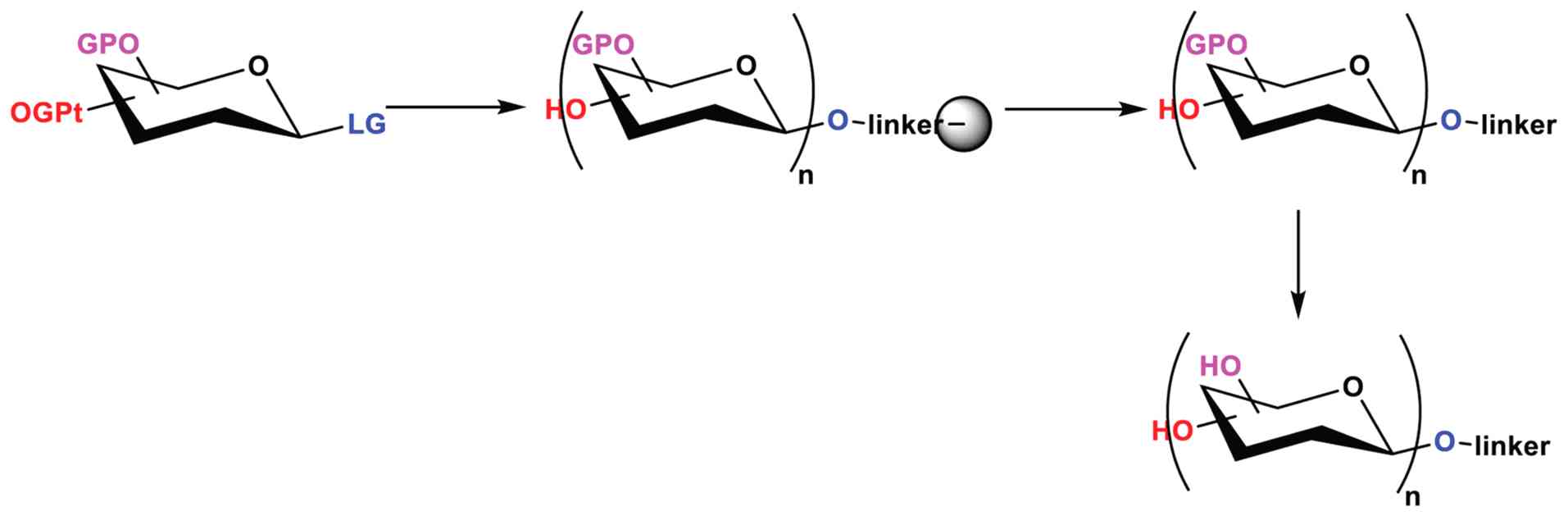
The AGA oligosaccharide synthesis workflow is
designed to minimize the number of purification steps and
manipulations (Fig. 5). Inside the
reaction vessel of the synthesizer, a resin-bound linker serves as
an anchor for sequentially attaching monosaccharide building
blocks. In this manner, excess reagents can be washed away, and
time-consuming intermediate purification steps can be avoided.
Following synthesis, the resin-bound oligosaccharide is removed
from the synthesizer and cleaved from the solid support. Analytical
normal-phase high-performance liquid chromatography (NP-HPLC) and
MALDI analysis of the crude product after cleavage are used to
assess the success of the synthesis qualitatively. The protected
glycan is purified using preparative NP-HPLC. Global deprotection
removes all permanent protecting groups and following reverse-phase
HPLC (RP-HPLC), the unprotected glycan is obtained (177,178,180,181). This enhanced accessibility has
directly fueled progress in glycomimetic design, in which natural
carbohydrate leads are modified to overcome inherent limitations
such as poor metabolic stability and low bioavailability (182). Common strategies include
replacing the endocyclic oxygen with carbon (carbasugars) or sulfur
(thiosugars), or substituting the glycosidic oxygen with a
methylene group to form C-glycosides, all of which confer
resistance to enzymatic hydrolysis by glycosidases (183).
Cyclodextrin (CD)-based materials have garnered
applications in the food and pharmaceutical industries since the
1980s. For example, Sugammadex (Bridion), a modified γ-CD, has been
used in anesthesia since 2008 and also functions as an antidote to
some curare-like muscle relaxants (184). The electrostatic interaction and
complex formation of CD with drugs can significantly alter the
pharmacokinetic profile of a drug, and CD inclusion complex-based
nanofibers have great applications in drug discovery and delivery,
as they tend to increase the bioavailability of the drugs (185-187).
For instance, the oral delivery of insulin is difficult owing to
its instability in the gastric conditions of patients. However,
reports suggest that carboxymethyl-β-CD-grafted chitosan
(CMCD-g-CS) leads to the more effective oral delivery of insulin at
pH 7.4 due to increased insulin bioavailability after pH-triggered
complexation with CMCD-g-CS (188).
Other saccharide derivatives, such as imine sugars,
have been suggested to function as potent inhibitors to several
glycosidases, such as glycogen phosphorylases, glycoside hydrolases
and glycosyltransferases. Their inhibitory potency against these
carbohydrate-processing enzymes is suggested to depend on
structural and electronic similarity to the oxocarbenium transition
state of the natural substrate at a physiological pH (189,190).
Additionally, research on carbohydrate-conjugated
noble metal-based nanoparticles, such as Au, Ag and Pt, has seen
tremendous growth over the past few decades due to their
applications in bioimaging, diagnostic imaging, biosensing,
gene/drug delivery (targeted/triggered/controlled), targeted
therapeutics. Despite their inertness, they can form a stable
covalent linkage via facile synthetic methods with compounds
containing functional groups, such as thiols and disulfides
(16).
7. Carbotoxicity and its relevance to
fructose-induced lipogenesis
Even though lipotoxicity (excess of lipids or a
specific lipid class) is a more familiar term in the biomedical
field, carbotoxicity is gaining increased recognition. The
excessive ingestion of carbohydrates has the potential to degrade
human health and ultimately cause the development of metabolic
syndrome, diabetes, obesity and their multiple co-morbidities.
Hence, the concept of ‘carbotoxicity’ was proposed in 2018, stating
that non-cellulose-digestible carbohydrates can be toxic to a
certain extent (12).
Additionally, the PURE cohort study established a link between
excessive carbohydrate consumption and adverse health effects
worldwide (82). The study
involved human subjects, demonstrating that a high intake of
carbohydrates could increase the risk of mortality with a hazard
ratio (HR) of 1.28 for highest 77.2% vs. lowest 46.4% quintile
carbohydrate energy, which is significantly low with increasing fat
intake with a HR of 0.77 (35.3 vs. 10.6% quintile energy) or excess
protein consumption with a HR 0.88 (19.7 vs. 10.8% quintile energy)
(82,191). Additional research has
demonstrated that the excess intake of free sugars (monosaccharides
and disaccharides) is associated with the risk of DNL in the liver,
leading to the accumulation of a large number of lipids and finally
resulting in NAFLD; an increase in the hepatic DNL following a
high-carbohydrate meal was found in ob/ob mice, Western-diet-fed
mice and healthy individuals, which may increase the risk of
hepatic steatosis (192). Another
study with human trials demonstrated that carbohydrate-restricted
diets, otherwise known as ketogenic diets, can serve as a good
technique for weight loss and the management of type 2 diabetes
(193), along with various
antidiabetic therapies, including antidiabetic drugs that
effectively improve hyperglycemia and hyperinsulinemia (194-196).
The investigation of fructose-induced lipogenesis
through human clinical trials has been pivotal in establishing the
mechanistic link between excessive dietary fructose consumption and
metabolic dysfunction, forming a core component of carbotoxicity.
Carbotoxicity posits that the chronic overconsumption of specific
carbohydrates, particularly fructose, drives metabolic disease
through the substrate overload of hepatic metabolic pathways,
leading to ATP depletion, uric acid generation, mitochondrial
stress and transcriptional upregulation of lipogenic enzymes
(108). Unlike glucose, which is
metabolized in a regulated manner under insulin control, fructose
bypasses the rate-limiting enzyme phosphofructokinase, enabling
unregulated entry into glycolysis and acetyl-CoA production,
thereby fueling DNL, independent of insulin signaling. This
property renders it uniquely hepatotoxic at high doses (108,197).
Several landmarks and recent clinical trials have
provided robust evidence for this pathway. The foundational study
by Schwarz et al (198)
demonstrated that even a moderate increase in fructose intake can
significantly affect liver metabolism. In that trial, 40 children
with obesity were placed on an isocaloric diet, with fructose
restricted to 4% of total calories, for 9 days. This intervention
resulted in a marked 56% decrease in DNL, a 22% reduction in liver
fat and significant improvements in insulin kinetics, all without
any change in body weight. This finding was critical, as it
established that the lipogenic effects of fructose are direct and
rapid, and that its restriction can reverse key features of NAFLD
and insulin resistance, independently of weight loss (198).
Building on this, the 2018 randomized crossover
trial by Beysen et al (199) provided detailed dose-response
data on acute fructose administration in healthy men. The study
revealed that fructose feeding acutely and dose-dependently
stimulates hepatic fractional DNL, with considerable
inter-individual variability in susceptibility. That study
quantified the immediate lipogenic potential of fructose and
highlighted that not all individuals respond equally, suggesting a
role for genetic or metabolic predisposition (199).
More recently, the narrative review by Geidl-Flueck
and Gerber (200) and a
post-publication peer review (letter to the editor) by Prinz
(201) synthesized findings from
multiple controlled feeding trials. Their research directly
compared beverages sweetened with glucose, fructose and sucrose in
healthy men. These studies demonstrated that fructose and sucrose,
but not glucose, significantly increased hepatic DNL. These
findings were instrumental in differentiating the metabolic effects
of monosaccharides, confirming that fructose, as a component of
both sucrose and high-fructose corn syrup, is the primary driver of
sugar-induced lipogenesis.
The relevance of these trials to carbotoxicity is
profound. They provide human experimental evidence validating the
biochemical hypothesis: Excess fructose functions as a chronic
metabolic stressor. The process begins with fructose metabolism in
the liver, which rapidly depletes ATP and generates uric acid, a
known inducer of oxidative stress and inflammation (202). Concurrently, the flood of carbon
from fructose metabolism provides abundant substrate for DNL. This
is further amplified by the activation of key transcription
factors, such as carbohydrate-responsive element-binding protein
and sterol regulatory element-binding protein-1c, which upregulate
the entire lipogenic gene program (203). The resulting increase in the
intrahepatic triglyceride content is a hallmark of NAFLD, which is
now recognized as the hepatic manifestation of metabolic syndrome
(204).
Furthermore, the gut microbiome has emerged as a
crucial intermediary in this process. The pivotal study by Zhao
et al (205) demonstrated
that dietary fructose is not only metabolized by the liver, but
also by the gut microbiota, which convert it to acetate. This
microbiota-derived acetate is then used by the liver as an
additional substrate for DNL, effectively creating a second pathway
through which fructose promotes fat synthesis. This finding,
highlighted in a commentary by Postic (206), underscores the systemic nature of
the metabolic impact of fructose.
The cumulative evidence from these trials paints a
consistent picture: Fructose is a potent, direct stimulator of
hepatic DNL in humans. This mechanism is central to the
pathophysiology of NAFLD, dyslipidemia and insulin resistance, all
key components of metabolic syndrome. The concept of carbotoxicity
integrates these findings, framing high fructose intake not merely
as a source of empty calories, but as a specific metabolic toxin
that disrupts hepatic homeostasis through defined molecular and
physiological pathways. This understanding shifts the focus of
dietary recommendations from simple caloric restriction to the
quality of carbohydrate intake, emphasizing the need to limit added
sugars, particularly those rich in fructose, to mitigate the risk
of chronic metabolic diseases. The slow yet optimal strategy to
address carbotoxicity, a potential cause of obesity, is to
incorporate physical activity, dietary changes and lifestyle
modifications, which can help manage body weight and contribute to
improving metabolic syndrome, cardiovascular disease and premature
mortality.
8. Future perspectives
Carbohydrates and their derivatives play a crucial
role in several biological processes. Researchers have been
involved in the synthesis, characterization and applications of
carbohydrate derivatives for the past two decades. In continuation
of the ongoing projects in carbohydrate chemistry (207-210),
the present review attempted to elucidate the role of carbohydrates
in nutrition, which further broadened our research area. Scientists
are likely to focus on precision nutrition, tailoring carbohydrate
composition in foods to individual needs based on genetic and
metabolic factors. Socially, a growing demand for sustainable,
plant-based options is driving the food industry to innovate with
alternative carbohydrate sources. Health-conscious consumers may
drive the development of functional carbohydrates with specific
health benefits. Advances in biotechnology may lead to genetically
modified crops with improved carbohydrate profiles. Additionally,
smart packaging technologies could help preserve the freshness and
quality of carbohydrate-containing products.
9. Conclusion
Carbohydrates remain one of the most abundant,
versatile and biologically influential macronutrients in the human
diet. Far from functioning solely as sources of metabolic energy,
carbohydrates operate at the intersection of food structure,
digestion kinetics, metabolic regulation, microbial ecology and
therapeutic innovation. Developments in glycomimetics,
carbohydrate-based polymers and automated glycan assembly highlight
their expanding role in therapeutics, immunomodulation and targeted
drug delivery. Looking ahead, emerging paradigms, such as precision
nutrition, microbiome-informed dietary strategies and
structure-guided food processing approaches are poised to redefine
carbohydrate functionality in both health and disease. In
continuation of the ongoing project in carbohydrate chemistry, the
present review attempted to briefly consolidate the roles of
carbohydrates in the food industry, their nutritional value and
their medicinal aspects, emphasizing their function as a critical
macronutrient. Understanding the numerous functions of
carbohydrates will aid in humans make better dietary decisions that
will support their general well-being with healthier diets,
functional foods and innovative therapeutics as aligning
carbohydrate intake with physiological needs, while minimizing
exposure to refined and ultra-processed forms, remains a central,
evidence-based strategy for promoting metabolic health and
preventing chronic disease in the modern era. The present review
emphasizes the connection between carbohydrate-rich foods and human
health, while promoting knowledgeable, sensible eating habits. By
adopting a narrative review and perspective-based approach, the
present review consolidates the current understanding of
carbohydrates across nutrition, metabolism and therapeutics, while
highlighting emerging concepts, such as carbotoxicity and precision
nutrition.
Acknowledgements
SP would like to thank the Central University of
Tamil Nadu (CUTN), Thiruvarur, Tamil Nadu, India for the research
fellowship and infrastructure facilities. All the authors would
like to thank the Central University of Tamil Nadu (CUTN) and
DST-FIST (Department of Science and Technology-Fund for Improvement
of S&T Infrastructure), for the infrastructure facility. TMD
acknowledges Ms. Maruvada Samyuktha and Ms. Banoth Jahnavi,
Department of Chemistry, Venkatraman Ramakrishnan Block, School of
Basic and Applied Sciences, Central University of Tamil Nadu,
Thiruvarur, Tamil Nadu, India for taking part in the preparation of
the manuscript by collecting literature.
Funding
Funding: The present study received financial support from SERC
DST (Science and Engineering Research Council, Department of
Science and Technology), CSIR (Council of Scientific and Industrial
Research) and UGC (University Grants Commission), New Delhi,
India.
Availability of data and materials
Not applicable.
Authors' contributions
SP and TMD conceptualized the review topic and
designed the manuscript framework. VA and JV conducted the
literature search and screening of suitable literature. SP and VA
wrote the initial draft of the manuscript and the figures/tables
were prepared by VA and JV. TMD critically revised the manuscript
for important intellectual content and supervised the project. All
authors have read and approved the final manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Carocho M, Morales P and Ferreira ICFR:
Natural food additives: Quo vadis? Trends Food Sci Technol.
45:284–295. 2015.
|
|
2
|
Chen Y, Michalak M and Agellon LB:
Importance of nutrients and nutrient metabolism on human health.
Yale J Biol Med. 91:95–103. 2018.PubMed/NCBI
|
|
3
|
Moco S, Martin FPJ and Rezzi S:
Metabolomics view on gut microbiome modulation by polyphenol-rich
foods. J Proteome Res. 11:4781–4790. 2012.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Román S, Sánchez-Siles LM and Siegrist M:
The importance of food naturalness for consumers: Results of a
systematic review. Trends Food Sci Technol. 67:44–57. 2017.
|
|
5
|
Saleh ASM, Zhang Q, Chen J and Shen Q:
Millet grains: Nutritional quality, processing, and potential
health benefits. Compr Rev Food Sci Food Saf. 12:281–295. 2013.
|
|
6
|
Lu X, Chen J, Guo Z, Zheng Y, Rea MC, Su
H, Zheng X, Zheng B and Miao S: Using polysaccharides for the
enhancement of functionality of foods: A review. Trends Food Sci
Technol. 86:311–327. 2019.
|
|
7
|
Mozaffarian D: Dietary and policy
priorities for cardiovascular disease, diabetes, and obesity: A
comprehensive review. Circulation. 133:187–225. 2016.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Reynolds A, Mann J, Cummings J, Winter N,
Mete E and Te Morenga L: Carbohydrate quality and human health: A
series of systematic reviews and meta-analyses. Lancet.
393:434–445. 2019.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Razouki AS, Hameed WR, Naeem BA, Falaih
HH, Nassar SA and Mashchal MB: Metabolic effects of dietary fiber,
carbohydrate, glucose, lipid metabolism on human health and
nutritional therapy. Curr Clin Med Educ. 3:11–23. 2025.
|
|
10
|
Svihus B and Hervik AK: Digestion and
metabolic fates of starch, and its relation to major
nutrition-related health problems: A review. Starch - Stärke.
68:302–313. 2016.
|
|
11
|
Gao Q, Li Y and Wang L: Dietary
Carbohydrate modulation in metabolic syndrome: Mechanistic
insights, gut microbiota interactions, and precision nutrition
approaches. Food Rev Int. 1–29. 2026.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Kroemer G, López-Otín C, Madeo F and de
Cabo R: Carbotoxicity-noxious effects of carbohydrates. Cell.
175:605–614. 2018.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Kushwaha R, Vardhan PS and Kushwaha PP:
Chronic kidney disease interplay with comorbidities and
carbohydrate metabolism: A review. Life (Basel).
14(13)2023.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Topping DL and Clifton PM: Short-chain
fatty acids and human colonic function: Roles of resistant starch
and nonstarch polysaccharides. Physiol Rev. 81:1031–1064.
2001.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Liu H, Zhang M, Ma Q, Tian B, Nie C, Chen
Z and Li J: Health beneficial effects of resistant starch on
diabetes and obesity via regulation of gut microbiota: A review.
Food Funct. 11:5749–5767. 2020.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Thodikayil AT, Sharma S and Saha S:
Engineering carbohydrate-based particles for biomedical
applications: Strategies to construct and modify. ACS Appl Bio
Mater. 4:2907–2940. 2021.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Suri J and Gilmour R: Expediting
glycospace exploration: Therapeutic glycans via automated
synthesis. Angew Chem Int Ed Engl. 64(e202422766)2025.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Barrett JS and Gibson PR: Fermentable
oligosaccharides, disaccharides, monosaccharides and polyols
(FODMAPs) and nonallergic food intolerance: FODMAPs or food
chemicals? Ther Adv Gastroenterol. 5:261–268. 2012.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Gilbert RG, Wu AC, Sullivan MA, Sumarriva
GE, Ersch N and Hasjim J: Improving human health through
understanding the complex structure of glucose polymers. Anal
Bioanal Chem. 405:8969–8980. 2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Qi X and Tester RF: Fructose, galactose
and glucose-In health and disease. Clin Nutr ESPEN. 33:18–28.
2019.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Duceac IA, Stanciu MC, Nechifor M, Tanasă
F and Teacă CA: Insights on some polysaccharide gel type materials
and their structural peculiarities. Gels. 8(771)2022.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Tharanathan RN: Food-derived
carbohydrates-structural complexity and functional diversity. Crit
Rev Biotechnol. 22:65–84. 2002.PubMed/NCBI View Article : Google Scholar
|
|
23
|
K R, S VK, Saravanan P, Rajeshkannan R,
Rajasimman M, Kamyab H and Vasseghian Y: Exploring the diverse
applications of Carbohydrate macromolecules in food,
pharmaceutical, and environmental technologies. Environ Res.
240(117521)2024.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Garrido-Romero M, Montilla A and Moreno
FJ: Dietary carbohydrates: A trade-off between appealing
organoleptic and physicochemical properties and ability to control
glucose release and weight management. Curr Opin Food Sci.
49(100976)2023.
|
|
25
|
Wang R, Hartel RW, Zhai X, Fu W, Sun Y and
Wang S: Phase separation and gelation of gelatin-glucose syrup
mixtures and gummy confections: Effects of moisture content,
sugars, citric acid, and citrates. Food Hydrocoll.
153(110006)2024.
|
|
26
|
Wang Q and Wood P: Carbohydrates: Physical
properties. In: Handbook of Food Science, Technology, and
Engineering. Hui YH (Ed). Vol. 1. CRC Press: Taylor & Francis,
2005.
|
|
27
|
Wang Y, Bai Y, Ji H, Dong J, Li X, Liu J
and Jin Z: Insights into rice starch degradation by maltogenic
α-amylase: Effect of starch structure on its rheological
properties. Food Hydrocoll. 124(107289)2021.
|
|
28
|
Han L, Wei Q, Cao S, Yu Y, Cao X and Chen
W: The assisting effects of ultrasound on the multiscale
characteristics of heat-moisture treated starch from Agriophyllum
squarrosum seeds. Int J Biol Macromol. 187:471–480. 2021.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Trần TN: The role of physical chemistry in
food and beverage. Int J Sci Res Arch. 13:2959–2968. 2024.
|
|
30
|
Wang R, Zhai X, Hartel RW, Chang Y, Pang
W, Han W, Lv H and Wang S: Effects of saccharide type and extended
heating on the Maillard reaction and physicochemical properties of
high-solid gelatin gels. Food Chem. 459(140249)2024.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Urango ACM, Meireles MAA and Silva EK:
Maillard conjugates produced from proteins and prebiotic dietary
fibers: Technological properties, health benefits and challenges.
Trends Food Sci Technol. 147(104438)2024.
|
|
32
|
Wang W, Fan Z, Yan Q, Pan T, Luo J, Wei Y,
Li B, Fang Z and Lu W: Gut microbiota determines the fate of
dietary fiber-targeted interventions in host health. Gut Microbes.
16(2416915)2024.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Lu X, Dai X, He W, Ma S and Gong N:
Prebiotic Effects of Polysaccharides and Their Influence on
Lactobacilli. In: Exploring Lactobacilli: Biology, Roles and
Potential Applications in Food Industry and Human Health. Laranjo M
(Ed.), IntechOpen, 2025.
|
|
34
|
Lee H, Song J, Lee B, Cha J and Lee H:
Food carbohydrates in the gut: Structural diversity, microbial
utilization, and analytical strategies. Food Sci Biotechnol.
33:2123–2140. 2024.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Hernandez-Hernandez O, Sabater C,
Calvete-Torre I, Doyagüez EG, Muñoz-Labrador AM, Julio-Gonzalez C,
de las Rivas B, Muñoz R, Ruiz L, Margolles A, et al: Tailoring the
natural rare sugars D-tagatose and L-sorbose to produce novel
functional carbohydrates. NPJ Sci Food. 8(74)2024.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Sęczyk Ł, Sugier D, Świeca M and
Gawlik-Dziki U: The effect of in vitro digestion, food matrix, and
hydrothermal treatment on the potential bioaccessibility of
selected phenolic compounds. Food Chem. 344(128581)2021.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Song P, Huang Y, Li J, Shan S, Zhou Z, Cao
H and Zhao C: The influence of processing technologies on the
biological activity of carbohydrates in food. Food Chem X.
23(101590)2024.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Chen X, Zhang W, Quek SY and Zhao L:
Flavor-food ingredient interactions in fortified or reformulated
novel food: Binding behaviors, manipulation strategies, sensory
impacts, and future trends in delicious and healthy food design.
Compr Rev Food Sci Food Saf. 22:4004–4029. 2023.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Urminská D, Haring N, Fábry V and Urminská
J: Fermentable oligosaccharides, disaccharides, monosaccharides and
polyols and their role in food digestion. J Microbiol Biotech Food
Sci. 11(e5521)2022.
|
|
40
|
Borer KT: Relevance of milk composition to
human longitudinal growth from infancy through puberty: Facts and
controversies. Nutrients. 17(827)2025.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Romero-Velarde E, Delgado-Franco D,
García-Gutiérrez M, Gurrola-Díaz C, Larrosa-Haro A, Montijo-Barrios
E Muskiet FAJ, Vargas-Guerrero B and Geurts J: The importance of
lactose in the human diet: Outcomes of a Mexican consensus meeting.
Nutrients. 11(2737)2019.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Stanhope KL, Griffen SC, Bair BR,
Swarbrick MM, Keim NL and Havel PJ: Twenty-four-hour endocrine and
metabolic profiles following consumption of high-fructose corn
syrup-, sucrose-, fructose-, and glucose-sweetened beverages with
meals. Am J Clin Nutr. 87:1194–1203. 2008.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Khorshidian N, Shadnoush M, Zabihzadeh
Khajavi M, Sohrabvandi S, Yousefi M and Mortazavian AM: Fructose
and high fructose corn syrup: Are they a two-edged sword? Int J
Food Sci Nutr. 72:592–614. 2021.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Aghaei B, Moradi F, Soleimani D,
Moradinazar M, Khosravy T and Samadi M: Glycemic index, glycemic
load, dietary inflammatory index, and risk of infertility in women.
Food Sci Nutr. 11:6413–6424. 2023.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Grembecka M: Natural sweeteners in a human
diet. Rocz Panstw Zakl Hig. 66:195–202. 2015.PubMed/NCBI
|
|
46
|
den Berghe GV: How does blood glucose
control with insulin save lives in intensive care? J Clin Invest.
114:1187–1195. 2004.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wirfält E, McTaggart A, Pala V, Gullberg
B, Frasca G, Panico S, Bueno-de-Mesquita H, Peeters PHM, Engeset D,
Skeie G, et al: Food sources of carbohydrates in a European cohort
of adults. Public Health Nutr. 5:1197–1215. 2002.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Sünram-Lea SI, Foster JK, Durlach P and
Perez C: Glucose facilitation of cognitive performance in healthy
young adults: Examination of the influence of fast-duration, time
of day and pre-consumption plasma glucose levels.
Psychopharmacology (Berl). 157:46–54. 2001.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Alwahsh SM and Gebhardt R: Dietary
fructose as a risk factor for non-alcoholic fatty liver disease
(NAFLD). Arch Toxicol. 91:1545–1563. 2017.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Yu S, Li C, Ji G and Zhang L: The
contribution of dietary fructose to non-alcoholic fatty liver
disease. Front Pharmacol. 12(783393)2021.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Rizkalla SW: Health implications of
fructose consumption: A review of recent data. Nutr Metab (Lond).
7(82)2010.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Liu G, Hale GE and Hughes CL: Galactose
metabolism and ovarian toxicity. Reprod Toxicol. 14:377–384.
2000.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Jiang Y, Pan Y, Rhea PR, Tan L, Gagea M,
Cohen L and Yang P: A sucrose-enriched diet promotes tumorigenesis
in mammary gland in part through the 12-lipoxygenase pathway.
Cancer Res. 76:24–29. 2016.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Qi X and Tester RF: Lactose, maltose, and
sucrose in health and disease. Mol Nutr Food Res.
64(1901082)2020.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Misra V, Shrivastava AK, Shukla SP and
Ansari MI: Effect of sugar intake towards human health. Saudi J
Med. 1:29–36. 2016.
|
|
56
|
Misselwitz B, Butter M, Verbeke K and Fox
MR: Update on lactose malabsorption and intolerance: Pathogenesis,
diagnosis and clinical management. Gut. 68:2080–2091.
2019.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Poothullil JM: Maltose: The primary signal
of hunger and satiation in human beings. Physiol Behav. 52:27–31.
1992.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Costa GT, Guimarães SB and de Carvalho
Sampaio HA: Fructo-oligosaccharide effects on blood glucose: An
overview. Acta Cir Bras. 27:279–282. 2012.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Niittynen L, Kajander K and Korpela R:
Galacto-oligosaccharides and bowel function. Scand J Food Nutr.
51:62–66. 2007.
|
|
60
|
Otles S and Ozgoz S: Health effects of
dietary fiber. Acta Sci Pol Technol Aliment. 13:191–202.
2014.PubMed/NCBI
|
|
61
|
Anderson JW, Baird P, Davis RH JR, Ferreri
S, Knudtson M, Koraym A, Waters V and Williams CL: Health benefits
of dietary fiber. Nutr Rev. 67:188–205. 2009.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Delzenne NM, Bindels LB, Neyrinck AM and
Walter J: The gut microbiome and dietary fibers: implications in
obesity, cardiometabolic diseases and cancer. Nat Rev Microbiol.
23:225–238. 2024.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Tan Z, Yang C, Zhao Y, Yang X and Li T:
Interactional effects of food macronutrients with gut microbiome:
Implications for host health and risk. J Agric Food Chem.
73:19109–19132. 2025.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Mora-Flores LP, Moreno-Terrazas Casildo R,
Fuentes-Cabrera J, Pérez-Vicente HA, de Anda-Jáuregui G and
Neri-Torres EE: The role of carbohydrate intake on the gut
microbiome: A weight of evidence systematic review. Microorganisms.
11(1728)2023.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Frayn KN: Understanding human metabolism.
Cambridge University Press, Cambridge, 2022.
|
|
66
|
Wilkinson JG and Liebman M: Carbohydrate
metabolism in sport and exercise. In: Nutrition in Exercise and
Sport, third edition. CRC Press, Boca Raton, FL, pp63-99, 2022.
|
|
67
|
Garrido G, Guzmán M and Odriozola JM:
Effect of different types of high carbohydrate diets on glycogen
metabolism in liver and skeletal muscle of endurance-trained rats.
Eur J Appl Physiol Occup Physiol. 74:91–99. 1996.PubMed/NCBI View Article : Google Scholar
|
|
68
|
Paoli A, Bosco G, Camporesi EM and Mangar
D: Ketosis, ketogenic diet and food intake control: A complex
relationship. Front Psychol. 6(27)2015.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Dreher ML: Whole fruits and fruit fiber
emerging health effects. Nutrients. 10(1833)2018.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Yu W: Metabolic effects in C57B/6J mice
fed a high-fat diet supplemented with chromium picolinate and
chromium yeast. Honors College Thesis, 2019. https://ir.library.oregonstate.edu/concern/honors_college_theses/s1784s179.
|
|
71
|
Baky MH, Salah M, Ezzelarab N, Shao P,
Elshahed MS and Farag MA: Insoluble dietary fibers: structure,
metabolism, interactions with human microbiome, and role in gut
homeostasis. Crit Rev Food Sci Nutr. 64:1954–1968. 2024.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Yao Y, Cai X, Fei W, Ye Y, Zhao M and
Zheng C: The role of short-chain fatty acids in immunity,
inflammation and metabolism. Crit Rev Food Sci Nutr. 62:1–12.
2022.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Song M, Wu K, Meyerhardt JA, Ogino S, Wang
M, Fuchs CS and Chan AT: Fiber intake and survival after colorectal
cancer diagnosis. JAMA Oncol. 4:71–79. 2018.PubMed/NCBI View Article : Google Scholar
|
|
74
|
Tily H, Patridge E, Cai Y, Gopu V, Gline
S, Genkin M, Lindau H, Sjue A, Slavov I and Perlina A: Gut
microbiome activity contributes to prediction of individual
variation in glycemic response in adults. Diabetes Ther. 13:89–111.
2022.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Carmody RN, Varady K and Turnbaugh PJ:
Digesting the complex metabolic effects of diet on the host and
microbiome. Cell. 187:3857–3876. 2024.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Ludwig DS, Apovian CM, Aronne LJ, Astrup
A, Cantley LC, Ebbeling CB, Heymsfield SB, Johnson JD, King JC,
Krauss RM, et al: Competing paradigms of obesity pathogenesis:
Energy balance versus carbohydrate-insulin models. Eur J Clin Nutr.
76:1209–1221. 2022.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Ludwig DS: Carbohydrate-insulin model:
Does the conventional view of obesity reverse cause and effect?
Philos Trans R Soc Lond B Biol Sci. 378(20220212)2023.PubMed/NCBI View Article : Google Scholar
|
|
78
|
World Health Organization: Sugars intake
for adults and children. Geneva, 2015. https://www.who.int/publications/i/item/9789241549028.
|
|
79
|
Ludwig DS, Hu FB, Tappy L and Brand-Miller
J: Dietary carbohydrates: Role of quality and quantity in chronic
disease. BMJ. 361(k2340)2018.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Williams KA, Fughhi I, Fugar S, Mazur M,
Gates S, Sawyer S, Patel H, Chambers D, McDaniel R, Reiser JR, et
al: Nutrition intervention for reduction of cardiovascular risk in
african americans using the 2019 American college of
cardiology/american heart association primary prevention
guidelines. Nutrients. 13(3422)2021.PubMed/NCBI View Article : Google Scholar
|
|
81
|
Singh M, Hung ES, Cullum A, Allen RE,
Aggett PJ, Dyson P, Forouhi NG, Greenwood DC, Pryke R, Taylor R,
Twenefour D, et al: Lower carbohydrate diets for adults with type 2
diabetes. Br J Nutr. 127:1352–1357. 2021.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Dehghan M, Mente A, Zhang X, Swaminathan
S, Li W, Mohan V, Iqbal R, Kumar R, Wentzel-Viljoen E, Rosengren A,
et al: Associations of fats and carbohydrate intake with
cardiovascular disease and mortality in 18 countries from five
continents (PURE): A prospective cohort study. Lancet.
390:2050–2062. 2017.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Swaminathan S, Dehghan M, Raj JM, Thomas
T, Rangarajan S, Jenkins D, Mony P, Mohan V, Lear SA, Avezum A, et
al: Associations of cereal grains intake with cardiovascular
disease and mortality across 21 countries in prospective urban and
rural epidemiology study: Prospective cohort study. BMJ.
372(m4948)2021.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Malik VS, Li Y, Pan A, De Koning L,
Schernhammer E, Willett WC and Hu FB: Long-term consumption of
sugar-sweetened and artificially sweetened beverages and risk of
mortality in US adults. Circulation. 139:2113–2125. 2019.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Yin J, Zhu Y, Malik V, Li X, Peng X, Zhang
FF, Shan Z and Liu L: Intake of sugar-sweetened and low-calorie
sweetened beverages and risk of cardiovascular disease: A
meta-analysis and systematic review. Adv Nutr. 12:89–101.
2021.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Mahzari M and Mamun A: Does consumption of
refined carbohydrates predict the incidence of type 2 diabetes
mellitus? A systematic review and meta-analysis. Rom J Diabetes
Nutr Metab Dis. 27:168–179. 2020.
|
|
87
|
Maffetone P and Laursen PB: Refined
carbohydrates and the overfat pandemic: Implications for brain
health and public health policy. Front Public Health.
13(1585680)2025.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Lane MM, Gamage E, Du S, Ashtree DN,
McGuinness AJ, Gauci S, Baker P, Lawrence M, Rebholz CM, Srour B,
et al: Ultra-processed food exposure and adverse health outcomes:
Umbrella review of epidemiological meta-analyses. BMJ.
384(e077310)2024.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Juul F, Vaidean G, Lin Y, Deierlein AL and
Parekh N: Ultra-processed foods and incident cardiovascular disease
in the framingham offspring study. J Am Coll Cardiol. 77:1520–1531.
2021.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Lacroix S, Cantin J and Nigam A:
Contemporary issues regarding nutrition in cardiovascular
rehabilitation. Ann Phys Rehabil Med. 60:36–42. 2017.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Maughan R: Carbohydrate metabolism.
Surgery (Oxf). 27:6–10. 2009.
|
|
92
|
Bolaños JP, Almeida A and Moncada S:
Glycolysis: A bioenergetic or a survival pathway? Trends Biochem
Sci. 35:145–149. 2010.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Chandel NS: Glycolysis. Cold Spring Harb
Perspect Biol. 13(a040535)2021.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Akram M, Ali Shah SM, Munir N, Daniyal M,
Tahir IM, Mahmood Z, Irshad M, Akhlaq M, Sultana S and Zainab R:
Hexose monophosphate shunt, the role of its metabolites and
associated disorders: A review. J Cell Physiol. 234:14473–14482.
2019.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Machado MG: The role of acetate in
macrophages response against Streptococcus pneumoniae. Human health
and pathology (unpublished thesis). Université de Lille, 2022.
|
|
96
|
Dashty M: A quick look at biochemistry:
Carbohydrate metabolism. Clin Biochem. 46:1339–1352.
2013.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Stephen AM, Champ MMJ, Cloran SJ, Fleith
M, van Lieshout L, Mejborn H and Burley VJ: Dietary fibre in
Europe: Current state of knowledge on definitions, sources,
recommendations, intakes and relationships to health. Nutr Res Rev.
30:149–190. 2017.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Theodorakis N, Kreouzi M, Pappas A and
Nikolaou M: Beyond calories: Individual metabolic and hormonal
adaptations driving variability in weight management-a
state-of-the-art narrative review. Int J Mol Sci.
25(13438)2024.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Vergari E, Knudsen JG, Ramracheya R,
Salehi A, Zhang Q, Adam J, Asterholm IW, Benrick A, Briant LJ,
Chibalina MV, et al: Insulin inhibits glucagon release by
SGLT2-induced stimulation of somatostatin secretion. Nat Commun.
10(139)2019.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Tiller NB, Roberts JD, Beasley L, Chapman
S, Pinto JM, Smith L, Wiffin M, Russell M, Sparks SA, Duckworth L,
et al: International society of sports nutrition position stand:
Nutritional considerations for single-stage ultra-marathon training
and racing. J Int Soc Sports Nutr. 16(50)2019.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Tappy L: Metabolism of sugars: A window to
the regulation of glucose and lipid homeostasis by splanchnic
organs. Clin Nutr. 40:1691–1698. 2021.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Li Z, Fan X, Gao F, Pan S, Ma X, Cheng H,
Nakatsukasa H, Zhang W and Zhang D: Fructose metabolism and its
roles in metabolic diseases, inflammatory diseases, and cancer. Mol
Biomed. 6(43)2025.PubMed/NCBI View Article : Google Scholar
|
|
103
|
Cirillo P, Gersch MS, Mu W, Scherer PM,
Kim KM, Gesualdo L, Henderson GN, Johnson RJ and Sautin YY:
Ketohexokinase-dependent metabolism of fructose induces
proinflammatory mediators in proximal tubular cells. J Am Soc
Nephrol. 20:545–553. 2009.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Federico A, Rosato V, Masarone M, Torre P,
Dallio M, Romeo M and Persico M: The role of fructose in
non-alcoholic steatohepatitis: Old relationship and new insights.
Nutrients. 13(1314)2021.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Tappy L and Lê KA: Metabolic effects of
fructose and the worldwide increase in obesity. Physiol Rev.
90:23–46. 2010.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Zhu Z, Zhang X, Pan Q, Zhang L and Chai J:
In-depth analysis of de novo lipogenesis in non-alcoholic fatty
liver disease: Mechanism and pharmacological interventions. Liver
Res. 7:285–295. 2023.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Khitan Z and Kim DH: Fructose: A key
factor in the development of metabolic syndrome and hypertension. J
Nutr Metab. 2013(682673)2013.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Takir M, Kostek O, Ozkok A, Elcioglu OC,
Bakan A, Erek A, Mutlu HH, Telci O, Semerci A, Odabas AR, et al:
Lowering uric acid with allopurinol improves insulin resistance and
systemic inflammation in asymptomatic hyperuricemia. J Investig
Med. 63:924–929. 2015.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Kim YJ, Jung DH and Park CS: Important
roles of Ruminococcaceae in the human intestine for resistant
starch utilization. Food Sci Biotechnol. 33:2009–2019.
2024.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Bedu-Ferrari C, Biscarrat P, Pepke F, Vati
S, Chaudemanche C, Castelli F, Chollet C, Rué O, Hennequet-Antier
C, Langella P and Cherbuy C: In-depth characterization of a
selection of gut commensal bacteria reveals their functional
capacities to metabolize dietary carbohydrates with prebiotic
potential. mSystems. 9(e0140123)2024.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Liu L, Sun T, Liu H, Li J and Tian L:
Carbohydrate quality vs quantity on cancer risk: Perspective of
microbiome mechanisms. J Funct Foods. 118(106246)2024.
|
|
112
|
Pavani M, Geethika KV, Ramya D, Patibandla
J and Sunitha A: The role of carbohydrate intake in obesity:
Implications for diet and weight management. J Integral Sci.
8:16–21. 2025.
|
|
113
|
Burkitt DP and Trowell HC: Dietary fibre
and western diseases. Ir Med J. 70:272–277. 1977.PubMed/NCBI
|
|
114
|
Painter NS and Burkitt DP: Diverticular
disease of the colon: A deficiency disease of western civilization.
Br Med J. 2:450–454. 1971.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Veronese N, Gianfredi V, Solmi M,
Barbagallo M, Dominguez LJ, Mandalà C, Di Palermo C, Carruba L,
Solimando L, Stubbs B, et al: The impact of dietary fiber
consumption on human health: An umbrella review of evidence from
17,155,277 individuals. Clin Nutr. 51:325–333. 2025.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Tao J, Quan J, El Helali A, Lam WWT and
Pang H: Global trends indicate increasing consumption of dietary
sodium and fiber in middle-income countries: A study of 30-year
global macrotrends. Nutr Res. 118:63–69. 2023.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Schönenberger KA, Huwiler VV, Reber E,
Mühlebach S, Stanga Z, Pestoni G and Faeh D: Dietary fibre intake
and its association with ultraprocessed food consumption in the
general population of Switzerland: Analysis of a population-based,
cross-sectional national nutrition survey. BMJ Nutr Prev Health.
7:26–37. 2024.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Cifuentes L and O'Keefe S: Analysis of
1999-2017 NHANES data: Minimal increase and racial disparities in
U.S. Fiber consumption over 18 years. Nutr Cancer. 76:345–351.
2024.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Guzmán C, Espinoza J and Fuentealba F:
Pilot study to estimate dietary fiber intake in adults residing in
chile. Nutrients. 15(900)2023.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Guzmán C, Espinoza J, Durán-Agüero S,
Obregón AM and Fuentealba F: Dietary fiber intake in chile: 13
years after the last national report. Nutrients.
15(3671)2023.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Stribling P and Ibrahim F: Dietary fibre
definition revisited-The case of low molecular weight
carbohydrates. Clin Nutr ESPEN. 55:340–356. 2023.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Li L, Yan S, Liu S, Wang P, Li W, Yi Y and
Qin S: In-depth insight into correlations between gut microbiota
and dietary fiber elucidates a dietary causal relationship with
host health. Food Res Int. 172(113133)2023.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Xie L, Alam MJ, Marques FZ and Mackay CR:
A major mechanism for immunomodulation: Dietary fibres and acid
metabolites. Semin Immunol. 66(101737)2023.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Gill SK, Rossi M, Bajka B and Whelan K:
Dietary fiber in gastrointestinal health and disease. Nat Rev
Gastroenterol Hepatol. 18:101–116. 2020.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Schneider E, Balasubramanian R, Ferri A,
Cotter PD, Clarke G and Cryan JF: Fiber & fermented foods:
Differential effects on the microbiota-gut-brain axis. Proc Nutr
Soc. 84:365–380. 2025.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Deng M, Dan L, Ye S, Chen X, Wang X, Tian
L and Chen J: Higher fiber intake is associated with reduced risk
of related surgery among individuals with inflammatory bowel
disease in a prospective cohort study. J Nutr. 153:2274–2282.
2023.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Deehan EC, Mocanu V and Madsen KL: Effects
of dietary fiber on metabolic health and obesity. Nat Rev
Gastroenterol Hepatol. 21:301–318. 2024.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Sharma D, Sharma L, Banerjee D and Dhama
K: Effect of dietary fiber on obese/overweight persons and its
mechanism of action: A systematic and meta-analysis on randomized
controlled trials. Nutr Clin Métabol. 39:164–179. 2025.
|
|
129
|
Liu M: A review of the effects of dietary
fiber on type 2 diabetes. Theor Nat Sci. 124:114–119. 2025.
|
|
130
|
Miketinas DC, Tucker WJ, Douglas CC and
Patterson MA: Usual dietary fibre intake according to diabetes
status in USA adults-NHANES 2013-2018. Br J Nutr. 130:1056–1064.
2023.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Biscarrat P, Bedu-Ferrari C, Langella P
and Cherbuy C: Pulses: A way to encourage sustainable fiber
consumption. Trends Food Sci Technol. 143(104281)2024.
|
|
132
|
Sempio R, Segura Godoy C, Nyhan L, Sahin
AW, Zannini E, Walter J and Arendt EK: Closing the fibre gap-the
impact of combination of soluble and insoluble dietary fibre on
bread quality and health benefits. Foods. 13(1980)2024.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Wang Y, Jian C, Salonen A, Dong M and Yang
Z: Designing healthier bread through the lens of the gut
microbiota. Trends Food Sci Technol. 134:13–28. 2023.
|
|
134
|
Wang J, He S, Tao S, Ma S, Luo Y, Wu M and
Zhou M: Structural, physicochemical, prebiotic properties of guava
pulp insoluble dietary fiber and its quality enhancement ability on
cow/goat yogurt: Impacts of ultrasound-assisted enzyme treatment.
Food Biosci. 58(103797)2024.
|
|
135
|
Aksornkitti V, Visuthranukul C,
Leelahavanichkul A, Joyjinda Y and Sriswasdi S: Gut microbiome
enterotypes and temporal variations during a six-month
inulin-supplemented weight loss randomized controlled trial in Thai
children with obesity. Comput Struct Biotechnol J. 27:5007–5019.
2025.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Wu X, Li S, Chen L, Ma S, Ma B, Song L and
Qian D: Stereoselective construction of multifunctional
C-glycosides enabled by nickel-catalyzed tandem
borylation/glycosylation. J Am Chem Soc. 146:22413–22423.
2024.PubMed/NCBI View Article : Google Scholar
|
|
137
|
van Kooyk Y and Rabinovich GA:
Protein-glycan interactions in the control of innate and adaptive
immune responses. Nat Immunol. 9:593–601. 2008.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Fernández-Tejada A, Cañada FJ and Jiménez
Barbero J: Glycans in medicinal chemistry: An underexploited
resource. ChemMedChem. 10:1291–1295. 2015.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Vasanthan RJ, Pradhan S and Thangamuthu
MD: Emerging aspects of triazole in organic synthesis: Exploring
its potential as a gelator. Curr Org Synth. 21:456–512.
2024.PubMed/NCBI View Article : Google Scholar
|
|
140
|
LaMoia TE and Shulman GI: Cellular and
molecular mechanisms of metformin action. Endocr Rev. 42:77–96.
2021.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Shurrab NT and Arafa ESA: Metformin: A
review of its therapeutic efficacy and adverse effects. Obes Med.
17(100186)2020.
|
|
142
|
Magliocca S, De Caro C, Lazzarato L, Russo
R, Rolando B, Chegaev K, Marini E, Nieddu M, Burrai L, Boatto G, et
al: Aceclofenac-galactose conjugate: design, synthesis,
characterization, and pharmacological and toxicological
evaluations. Mol Pharm. 15:3101–3110. 2018.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Klyosov AA: Carbohydrates and drug design.
ACS Symp Ser. 1021:3–22. 2012.
|
|
144
|
Ajaykumar C: Overview on the side effects
of doxorubicin. IntechOpen, 2021.
|
|
145
|
Putri SA, Purnomo RW and Savitri IJ: The
potential of sucralfate as adjunctive therapy in the treatment of
periodontal disease: A review. World J Adv Res Rev. 25:490–495.
2025.
|
|
146
|
Cao X, Du X, Jiao H, An Q, Chen R, Fang P,
Wang J and Yu B: Carbohydrate-based drugs launched during
2000-2021. Acta Pharm Sin B. 12:3783–3821. 2022.PubMed/NCBI View Article : Google Scholar
|
|
147
|
Panesar PS and Kumari S: Lactulose:
Production, purification and potential applications. Biotechnol
Adv. 29:940–948. 2011.PubMed/NCBI View Article : Google Scholar
|
|
148
|
Ruszkowski J and Witkowski JM: Lactulose:
Patient- and dose-dependent prebiotic properties in humans.
Anaerobe. 59:100–106. 2019.PubMed/NCBI View Article : Google Scholar
|
|
149
|
Bregman DB and Goodnough LT: Experience
with intravenous ferric carboxymaltose in patients with iron
deficiency anemia. Ther Adv Hematol. 5:48–60. 2014.PubMed/NCBI View Article : Google Scholar
|
|
150
|
Boots JMM and Quax RAM: High-dose
intravenous iron with either ferric carboxymaltose or ferric
derisomaltose: A benefit-risk assessment. Drug Saf. 45:1019–1036.
2022.PubMed/NCBI View Article : Google Scholar
|
|
151
|
Patocka J, Nepovimova E, Wu W and Kuca K:
Digoxin: Pharmacology and toxicology-a review. Environ Toxicol
Pharmacol. 79(103400)2020.PubMed/NCBI View Article : Google Scholar
|
|
152
|
Hricovíni M: Structural aspects of
carbohydrates and the relation with their biological properties.
Curr Med Chem. 11:2565–2583. 2004.PubMed/NCBI View Article : Google Scholar
|
|
153
|
Donat F, Duret JP, Santoni A, Cariou R,
Necciari J, Magnani H and de Greef R: The pharmacokinetics of
fondaparinux sodium in healthy volunteers. Clin Pharmacokinet. 41
(Suppl 2):S1–S9. 2002.PubMed/NCBI View Article : Google Scholar
|
|
154
|
Anderson VR and Perry CM: Pentosan
polysulfate: A review of its use in the relief of bladder pain or
discomfort in interstitial cystitis. Drugs. 66:821–835.
2006.PubMed/NCBI View Article : Google Scholar
|
|
155
|
Putra ON, Musfiroh I, Elisa S, Musa M,
Ikram EHK, Chaidir C and Muchtaridi M: Sodium starch glycolate
(SSG) from sago starch (metroxylon sago) as a superdisintegrant:
Synthesis and characterization. Molecules. 29(151)2023.PubMed/NCBI View Article : Google Scholar
|
|
156
|
Stallforth P, Lepenies B, Adibekian A and
Seeberger PH: 2009 Claude S. Hudson award in carbohydrate
chemistry. Carbohydrates: A frontier in medicinal chemistry. J Med
Chem. 52:5561–5577. 2009.PubMed/NCBI View Article : Google Scholar
|
|
157
|
Campbell H and Carter H: Rational use of
haemophilus influenzae type b vaccine. Drugs. 46:378–383.
1993.PubMed/NCBI View Article : Google Scholar
|
|
158
|
Pifferi C, Fuentes R and Fernández-Tejada
A: Natural and synthetic carbohydrate-based vaccine adjuvants and
their mechanisms of action. Nat Rev Chem. 5:197–216.
2021.PubMed/NCBI View Article : Google Scholar
|
|
159
|
Pan L, Cai C, Liu C, Liu D, Li G, Linhardt
RJ and Yu G: Recent progress and advanced technology in
carbohydrate-based drug development. Curr Opin Biotechnol.
69:191–198. 2021.PubMed/NCBI View Article : Google Scholar
|
|
160
|
Maxwell EG, Colquhoun IJ, Chau HK,
Hotchkiss AT, Waldron KW, Morris VJ and Belshaw NJ: Modified sugar
beet pectin induces apoptosis of colon cancer cells via an
interaction with the neutral sugar side-chains. Carbohydr Polym.
136:923–929. 2016.PubMed/NCBI View Article : Google Scholar
|
|
161
|
Ji X, Peng Q, Yuan Y, Shen J, Xie X and
Wang M: Isolation, structures and bioactivities of the
polysaccharides from jujube fruit (Ziziphus jujuba Mill.): A
review. Food Chem. 227:349–357. 2017.PubMed/NCBI View Article : Google Scholar
|
|
162
|
Shao P, Chen X and Sun P: Chemical
characterization, antioxidant and antitumor activity of sulfated
polysaccharide from Sargassum horneri. Carbohydr Polym.
105:260–269. 2014.PubMed/NCBI View Article : Google Scholar
|
|
163
|
Shao P, Pei Y, Fang Z and Sun P: Effects
of partial desulfation on antioxidant and inhibition of DLD cancer
cell of Ulva fasciata polysaccharide. Int J Biol Macromol.
65:307–313. 2014.PubMed/NCBI View Article : Google Scholar
|
|
164
|
Yu Y, Shen M, Song Q and Xie J: Biological
activities and pharmaceutical applications of polysaccharide from
natural resources: A review. Carbohydr Polym. 183:91–101.
2018.PubMed/NCBI View Article : Google Scholar
|
|
165
|
Bhavya PV, Rabecca Jenifer V, Ahmad Y and
Mohan Das T: A simple perspective of glycosciences. J Indian Chem
Soc. 97:157–176. 2020.
|
|
166
|
Lalhmangaihzuala S, Vanlaldinpuia K,
Khiangte V, Laldinpuii Z, Liana T, Lalhriatpuia C and Pachuau Z:
Therapeutic applications of carbohydrate-based compounds: A sweet
solution for medical advancement. Mol Divers. 28:4553–4579.
2024.PubMed/NCBI View Article : Google Scholar
|
|
167
|
Chen Y, Song J, Chen X and Chen G:
Synthetic glycopolymers in tumor immunotherapy. Macromol Rapid
Commun. 46(e2401089)2025.PubMed/NCBI View Article : Google Scholar
|
|
168
|
Hulugalla K, Abodunrin OD, Anderson J,
Smith AE and Werfel T: A ‘sweeter’ approach to cancer
immunotherapy: Glycopolymers as diverse tools for immune
modulation. J Control Release. 388(114372)2025.PubMed/NCBI View Article : Google Scholar
|
|
169
|
Bhirud D, Bhattacharya S and Prajapati BG:
Bioengineered carbohydrate polymers for colon-specific drug
release: Current trends and future prospects. J Biomed Mater Res A.
112:1860–1872. 2024.PubMed/NCBI View Article : Google Scholar
|
|
170
|
Udaipuria N and Bhattacharya S: Novel
carbohydrate polymer-based systems for precise drug delivery in
colon cancer: Improving treatment effectiveness with intelligent
biodegradable materials. Biopolymers. 116(e23632)2025.PubMed/NCBI View Article : Google Scholar
|
|
171
|
Mishra VK, Khanna A, Tiwari G, Tyagi R and
Sagar R: Recent developments on the synthesis of biologically
active glycohybrids. Bioorg Chem. 145(107172)2024.PubMed/NCBI View Article : Google Scholar
|
|
172
|
Plante OJ, Palmacci ER and Seeberger PH:
Automated solid-phase synthesis of oligosaccharides. Science.
291:1523–1527. 2001.PubMed/NCBI View Article : Google Scholar
|
|
173
|
Nokami T, Hayashi R, Saigusa Y, Shimizu A,
Liu CY, Mong KKT and Yoshida JI: Automated solution-phase synthesis
of oligosaccharides via iterative electrochemical assembly of
thioglycosides. Org Lett. 15:4520–4523. 2013.PubMed/NCBI View Article : Google Scholar
|
|
174
|
Tang SL, Linz LB, Bonning BC and Pohl NLB:
Automated solution-phase synthesis of insect glycans to probe the
binding affinity of pea enation mosaic virus. J Org Chem.
80:10482–10489. 2015.PubMed/NCBI View Article : Google Scholar
|
|
175
|
Ganesh NV, Fujikawa K, Tan YH, Stine KJ
and Demchenko AV: HPLC-assisted automated oligosaccharide
synthesis. Org Lett. 14:3036–3039. 2012.PubMed/NCBI View Article : Google Scholar
|
|
176
|
Panza M, Pistorio SG, Stine KJ and
Demchenko AV: Automated chemical oligosaccharide synthesis: Novel
approach to traditional challenges. Chem Rev. 118:8105–8150.
2018.PubMed/NCBI View Article : Google Scholar
|
|
177
|
Hahm HS, Schlegel MK, Hurevich M, Eller S,
Schuhmacher F, Hofmann J, Pagel K and Seeberger PH: Automated
glycan assembly using the glyconeer 2.1 synthesizer. Proc Natl Acad
Sci USA. 114:E3385–E3389. 2017.PubMed/NCBI View Article : Google Scholar
|
|
178
|
Seeberger PH: The logic of automated
glycan assembly. Acc Chem Res. 48:1450–1463. 2015.PubMed/NCBI View Article : Google Scholar
|
|
179
|
Naresh K, Schumacher F, Hahm HS and
Seeberger PH: Pushing the limits of automated glycan assembly:
Synthesis of a 50mer polymannoside. Chem Commun (Camb).
53:9085–9088. 2017.PubMed/NCBI View Article : Google Scholar
|
|
180
|
Guberman M and Seeberger PH: Automated
glycan assembly: A perspective. J Am Chem Soc. 141:5581–5592.
2019.PubMed/NCBI View Article : Google Scholar
|
|
181
|
Niggemeyer GB, Danglad-Flores JA and
Seeberger PH: Automated synthesis of C1-functionalized
oligosaccharides. J Am Chem Soc. 147:1649–1655. 2024.PubMed/NCBI View Article : Google Scholar
|
|
182
|
Yuan L, Hua Y and Wang X: Recent progress
of glycomimetics in drug development. Org Biomol Chem.
23:7671–7680. 2025.PubMed/NCBI View Article : Google Scholar
|
|
183
|
Martínez-Pascual R, Valera-Zaragoza M,
Fernández-Bolaños JG and López Ó: Exploring the chemistry and
applications of Thio-, Seleno-, and tellurosugars. Molecules.
30(2053)2025.PubMed/NCBI View Article : Google Scholar
|
|
184
|
Brandariz I and Iglesias E: Local
anesthetics: Acid base behaviour and inclusion with cyclodextrin.
Curr Org Chem. 17:3050–3063. 2013.
|
|
185
|
Jacob S and Nair AB: Cyclodextrin
complexes: Perspective from drug delivery and formulation. Drug Dev
Res. 79:201–217. 2018.PubMed/NCBI View Article : Google Scholar
|
|
186
|
Jansook P, Ogawa N and Loftsson T:
Cyclodextrins: Structure, physicochemical properties and
pharmaceutical applications. Int J Pharm. 535:272–284.
2018.PubMed/NCBI View Article : Google Scholar
|
|
187
|
Gadade DD and Pekamwar SS: Cyclodextrin
based nanoparticles for drug delivery and theranostics. Adv Pharm
Bull. 10:166–183. 2020.PubMed/NCBI View Article : Google Scholar
|
|
188
|
Mansourpour M, Mahjub R, Amini M, Ostad
SN, Shamsa ES, Rafiee-Tehrani M and Dorkoosh FA: Development of
acid-resistant alginate/trimethyl chitosan nanoparticles containing
cationic β-cyclodextrin polymers for insulin oral delivery. AAPS
PharmSciTech. 16:952–962. 2015.PubMed/NCBI View Article : Google Scholar
|
|
189
|
Lahiri R, Ansari AA and Vankar YD: Recent
developments in design and synthesis of bicyclic azasugars,
carbasugars and related molecules as glycosidase inhibitors. Chem
Soc Rev. 42:5102–5118. 2013.PubMed/NCBI View Article : Google Scholar
|
|
190
|
Tamburrini A, Colombo C and Bernardi A:
Design and synthesis of glycomimetics: Recent advances. Med Res
Rev. 40:495–531. 2020.PubMed/NCBI View Article : Google Scholar
|
|
191
|
Wali JA, Raubenheimer D, Senior AM, Le
Couteur DG and Simpson SJ: Cardio-metabolic consequences of dietary
carbohydrates: Reconciling contradictions using nutritional
geometry. Cardiovasc Dis. 117:386–401. 2021.PubMed/NCBI View Article : Google Scholar
|
|
192
|
Gao Y, Hua R, Hu K and Wang Z:
Carbohydrates deteriorate fatty liver by activating the
inflammatory response. Nutr Res Rev. 35:252–267. 2022.PubMed/NCBI View Article : Google Scholar
|
|
193
|
Webster C: Preliminary investigations for
studying the effects of low carbohydrate high fat diets on
gluconeogenesis in type 2 diabetes patients (unpublished thesis).
University of Cape Town, 2020.
|
|
194
|
Bray GA, Fruhbeck G, Ryan DH and Wilding
JPH: Management of obesity. Lancet. 387:1947–1956. 2016.
|
|
195
|
Patnode CD, Evans CV, Senger CA, Redmond N
and Lin JS: Behavioral counseling to promote a healthful diet and
physical activity for cardiovascular disease prevention in adults
without known cardiovascular disease risk factors: Updated evidence
report and systematic review for the US preventive services task
force. JAMA. 318:175–193. 2017.PubMed/NCBI View Article : Google Scholar
|
|
196
|
Nakamura M and Sadoshima J: Cardiomyopathy
in obesity, insulin resistance and diabetes. J Physiol.
598:2977–2993. 2020.PubMed/NCBI View Article : Google Scholar
|
|
197
|
Liao Y: Computational modelling of
fructose metabolism and lipid deposition in non-alcoholic fatty
liver disease (unpublished thesis). University College London,
2021.
|
|
198
|
Schwarz JM, Noworolski SM, Erkin-Cakmak A,
Korn NJ, Wen MJ, Tai VW, Jones GM, Palii SP, Velasco-Alin M, Pan K,
et al: Effects of dietary fructose restriction on liver fat, de
novo lipogenesis, and insulin kinetics in children with obesity.
Gastroenterology. 153:743–752. 2017.PubMed/NCBI View Article : Google Scholar
|
|
199
|
Beysen C, Ruddy M, Stoch A, Mixson L,
Rosko K, Riiff T, Turner SM, Hellerstein MK and Murphy EJ:
Dose-dependent quantitative effects of acute fructose
administration on hepatic de novo lipogenesis in healthy humans. Am
J Physiol Endocrinol Metab. 315:E126–E132. 2018.PubMed/NCBI View Article : Google Scholar
|
|
200
|
Geidl-Flueck B and Gerber PA: Fructose
drives de novo lipogenesis affecting metabolic health. J
Endocrinol. 257(e220270)2023.PubMed/NCBI View Article : Google Scholar
|
|
201
|
Prinz P: Comments on ‘Fructose- and
sucrose- but not glucose-sweetened beverages promote hepatic de
novo lipogenesis-a randomized controlled trial’. J Hepatol.
75:753–754. 2021.PubMed/NCBI View Article : Google Scholar
|
|
202
|
Brouwers MCGJ: Fructose 1-phosphate, an
evolutionary signaling molecule of abundancy. Trends Endocrinol
Metab. 33:680–689. 2022.PubMed/NCBI View Article : Google Scholar
|
|
203
|
Iizuka K: The roles of carbohydrate
response element binding protein in the relationship between
carbohydrate intake and diseases. Int J Mol Sci.
22(12058)2021.PubMed/NCBI View Article : Google Scholar
|
|
204
|
Coronati M, Baratta F, Pastori D, Ferro D,
Angelico F and Del Ben M: Added fructose in non-alcoholic fatty
liver disease and in metabolic syndrome: A narrative review.
Nutrients. 14(1127)2022.PubMed/NCBI View Article : Google Scholar
|
|
205
|
Zhao S, Jang C, Liu J, Uehara K, Gilbert
M, Izzo L, Zeng X, Trefely S, Fernandez S, Carrer A, et al: Dietary
fructose feeds hepatic lipogenesis via microbiota-derived acetate.
Nature. 579:586–591. 2020.PubMed/NCBI View Article : Google Scholar
|
|
206
|
Postic C: Conversion of a dietary
fructose: New clues from the gut microbiome. Nat Metab. 2:217–218.
2020.PubMed/NCBI View Article : Google Scholar
|
|
207
|
Pradhan S, Małecki JG and Mohan Das T:
Highly selective iodide detection in solution and gel state using
tuneable benzoate N-glucosides. Anal Sens. 6(e202500083)2026.
|
|
208
|
V RJ, Malecki JG and Thangamuthu MD:
Photo-responsive organogelator based on cholesterol incorporated
sugar-azobenzene derivatives. Carbohydr Res.
549(109356)2025.PubMed/NCBI View Article : Google Scholar
|
|
209
|
Bhavya PV, Malecki JG and Mohan Das T:
Isoniazid-sugar-triazole-based fluorescent ‘turn-on’ chemosensor
for Zn2+ and Mn2+ detection with potential
biological applications. ChemistrySelect. 10(e202405443)2025.
|
|
210
|
Pradhan S, Hussain F, Małecki JG and Mohan
Das T: Pyridine hydrazide glycoconjugate-based gelators: Facile
synthesis and exploring their application in iodide sensing. J Mol
Liq. 416(126529)2024.
|