Introduction
Almost simultaneously with the first space flight,
the potential hazard of the space environment became of particular
interest. Especially in the context of long-term space flight and
extra-terrestrial habitation, it is imperative to characterize the
potential impact of space conditions, most prominently ionizing
radiation and microgravity, on development. Performing animal
experiments in space is non-trivial and developmental biology
studies in space are even more complicated as mating and sampling
at set time points are difficult to achieve. On the other hand,
space conditions are very difficult to reproduce on Earth: space
radiation is too complex and variable in terms of composition and
energy (1), and there is no
present way to cancel gravity on Earth.
Animal studies have shown that ionizing radiation
has a detrimental impact on embryonic development (2). While lethality is the main effect of
irradiation during the pre-implantation period, embryos may show a
variety of congenital anomalies after irradiation during the
organogenesis period which follows. Those could be caused by the
arrest of the development of a structure at an early stage of a
specific organ development (2).
Albeit being less radiosensitive than in younger embryos, the fetal
period is affected by radiation, especially at the level of the
central nervous system, the hematopoietic system and tissue
formation (2). The origin of
these developmental defects at the molecular level is assumed to
lie in the generation of DNA double strand breaks (DSBs) by
ionizing radiation (3–7). If not repaired properly, these
lesions can lead to cell death, proliferation arrest or mutations
which increase the risk of genetic diseases or cancer.
In-flight experiments have shown an impact of
microgravity during critical periods of mammalian development,
especially on bones, muscles and brain (8). However, most of the observed effects
were recovered upon return to Earth. Despite extensive research,
risk assessment of development in space will depend on the
understanding of how space conditions affect organs and tissues at
the molecular and cellular levels. To this end, robust models are
required that allow for simulating space conditions on Earth. For
microgravity simulation, various devices are currently used such as
rotating wall vessel (RWV) or the random positioning machine (RPM)
(9,10). The latter randomizes in three
dimensions the direction component of the gravity vector in order
to obtain, on average, a net force close to zero as the sample is
taken as a reference. In this way, the RPM has effects comparable
to real microgravity if the changes in direction are faster than
the response time of the sample and it is therefore considered as a
valuable microgravity simulation model (9–13).
We investigated the impact of simulated microgravity
on the cellular response to ionizing radiation by incubating cells
for 24 h in a RPM after exposure to increasing doses of X-rays.
Fetal skin fibroblasts were used as an in vitro model. While
simulated microgravity had no impact on radiation-induced
morphological or cell cycle alterations, it induced a significant
decrease in apoptosis.
Materials and methods
Cell culture, X-irradiation and simulated
microgravity
The STO [Sandos inbred mice (SIM)] thioguanine- and
ouabain-resistant) cell line is derived from a continuous line of
SIM skin fetal fibroblasts (14,15). Two milliliters of cells at a
density of 2.5×105 cells/ml were seeded in
12.5-cm2 flasks and cultured at 37°C and 5%
CO2 in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal bovine serum (FBS) and 1%
penicillin/streptomycin (all were from Invitrogen, Merelbeke,
Belgium).
Twenty-four hours after plating, cells were
X-irradiated with the following doses: 0, 0.5, 1 or 4 Gy (250 keV,
15 mA, 1-mm Cu). The dose rate was 0.273±0.006 Gy/min and the
accuracy of the given dose was higher than 99.99%.
Immediately after irradiation, the flasks were
filled up with culture medium and half of them were submitted to
simulated microgravity for 24 h while the others were placed in the
same incubator and served as gravity-controls. The medium was
placed in the same incubation conditions as during the experiment
(37°C, 5% CO2) for 24 h prior to the experiment in order
to be equilibrated in terms of temperature and gas. The desktop
random positioning machine (RPM) (Dutch Space, Leiden, The
Netherlands) was used for microgravity simulation at a rotational
velocity between 55°/sec and 65°/sec. Direction, speed and interval
were set as random. Directly after RPM treatment and for all
measurements, all cells (adherent and floating) were washed with
phosphate-buffered saline (PBS) (Invitrogen) and collected after
exposure to 0.05% trypsin-EDTA (Invitrogen).
Cellular morphology and cell area
measurement
After trypsinization, cells were washed twice with
PBS supplemented with 10% FBS and resuspended in 1 ml of PBS
supplemented with 1% FBS. Next, cells were centrifuged onto a slide
by cytospinning at 500 rpm for 5 min. The slides were stained with
May-Grünwald Giemsa and were analyzed by eye with light microscopy.
Cells (n=100) from the 3 biological replicates prepared for each of
the doses were scored for normality, vacuoles and morphological
signs of apoptosis (cell blebs).
Cell area was measured on the slides prepared for
cellular morphology assessment, with ImageJ freeware, 1.45h [W.S.
Rasband, ImageJ; US National Institutes of Health, Bethesda,
Maryland, USA; http://rsb.info.nih.gov/ij/ (1997–2010)]. In brief,
color images were converted to 8-bit grayscale and binarized using
a fixed threshold. After watershed-based segmentation, the
projected area was derived for the cellular regions of interest.
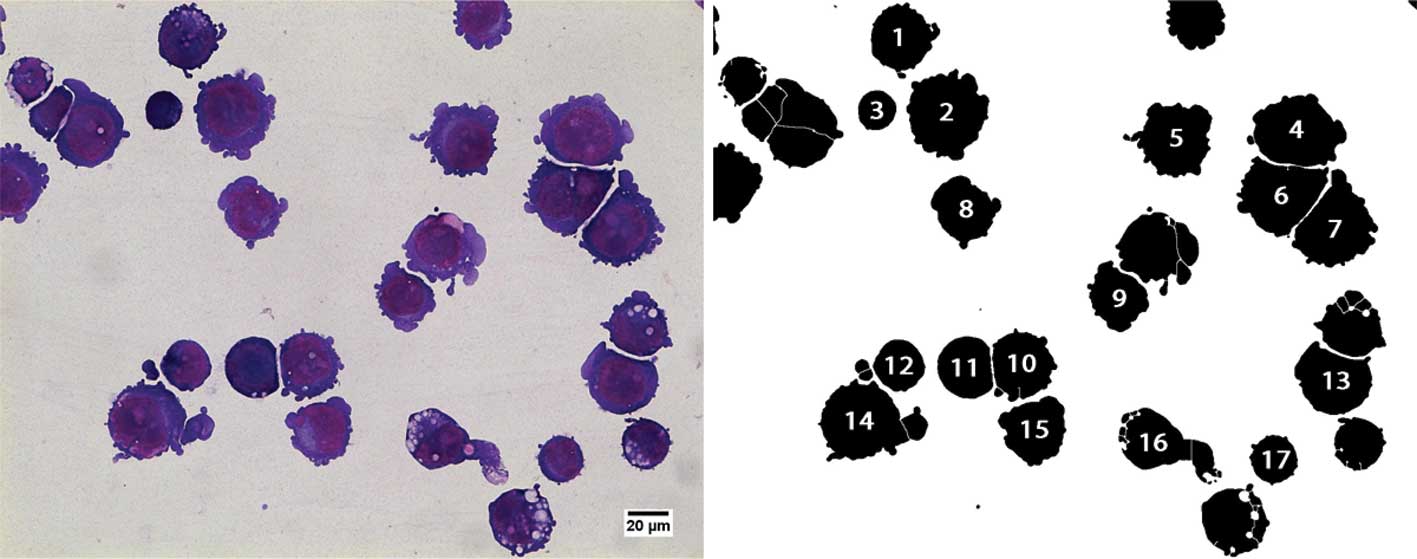
Incorrectly segmented cells were manually removed by visual
inspection. Fig. 1 shows an
example of a 4 Gy irradiated sample before and after
segmentation.
Flow cytometry
Cell cycle analysis and caspase-3 activity were
measured using a Beckman-Coulter EPICS XL flow cytometer. For all
assays and each condition, 20,000 cells were analyzed in
triplicates.
Cell cycle analysis
After trypsinization, cells were fixed in cold
ethanol (80%) for 1 h, after which they were washed in PBS and
suspended in a solution of propidium iodide (PI) red
(Sigma-Aldrich, Belgium) containing RNase (10 mM Tris, 5 mM
MgCl2, 10 μg/ml RNase, 40 μg/ml PI) before
being incubated at 37°C for 1 h. Estimation of G1, S and G2
fractions was performed by flow cytometry after discrimination of
doublets and aggregates using red fluorescence pulse width
analysis. In addition to gating away doublets and aggregates,
signals from cells with less than half the DNA content of the G1
peak were excluded to avoid micronuclei and apoptotic bodies. The
mitotic index (MI) was calculated as follows: MI =
[(1xG1)+(1.5xS)+(2xG2)]/100.
Apoptosis (caspase-3 activity and sub-G1
peak)
Caspase-3 activity assessment was performed
following the manufacturer’s instructions using the CaspGlow™ kit
(Medical and Biological Laboratories Co., Ltd., Woburn, MA,
USA).
While performing cell cycle analysis, cells that
showed a red fluorescence corresponding to a DNA content situated
between half the mean channel of G1 phase and the G1 phase mean
were referred to as sub-G1 cells.
Statistics
Graphics were performed using GraphPad Prism version
5.00 (GraphPad Software Inc., USA), while statistics were performed
with SPSS version 17.0 (IBM Corporation, USA). When data were
parametric (Kolmogorov-Smirnov test) one-way or two-way ANOVA was
performed with Sidak post hoc test to analyze differences
between treatments. All data were tested for homogeneity of
variance before ANOVA analysis. If not parametric, 2 sample tests
were performed by Kolmogorov-Smirnov and k sample tests were
performed by Kruskal-Wallis. Post hoc test for
Kruskal-Wallis tests were performed according to Chan and Walmsley
(16). Differences between means
were considered as significant when P-values <0.05.
Results
Cell morphology
In order to assess the effect of simulated
microgravity in combination to irradiation, STO cells were
irradiated and subsequently exposed to RPM for 24 h. After RPM
treatment, a portion of the cells appeared to have detached from
the flasks and formed globular cell aggregates (data not shown).
The occurrence of these aggregates was radiation-independent and
found at all doses, non-irradiated cells included. For
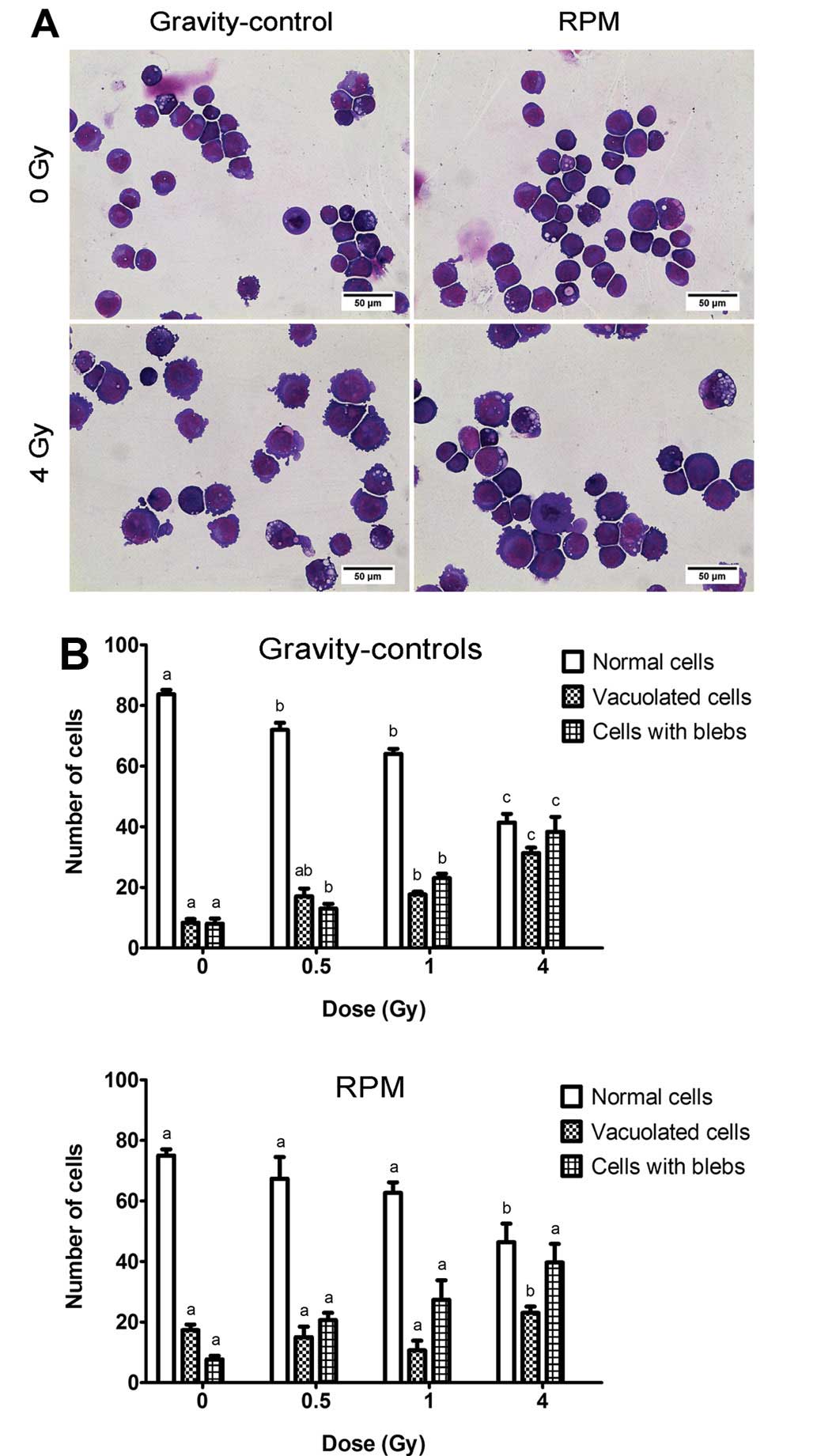
morphological assessment, the cells were mounted by cytospinning
and May-Grünwald-Giemsa stained (Fig.
2A). Subsequent visual inspection of stained slides showed an
increase in the number of vacuolated and blebbing cells with
radiation dose but no significant differences in the number of
normal and apoptotic cells (blebs) between RPM-treated cells and
gravity-controls, irrespective of the dose (Fig. 2B).
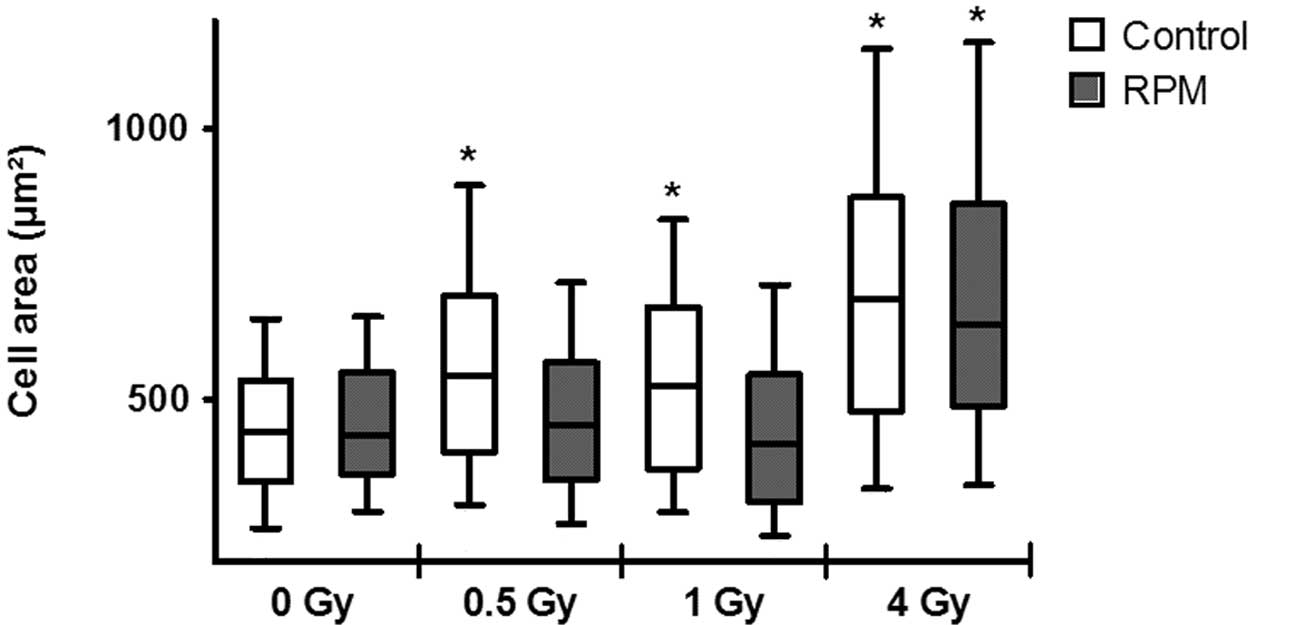
Using automated cell size analysis, a significant
increase was detected in gravity-controls for all doses (Fig. 3). However, this increase was not
observed for RPM treatment exposed to 0.5 Gy and 1 Gy of X-rays,
but only at 4 Gy, where it was comparable to the gravity-controls
irradiated with the same dose.
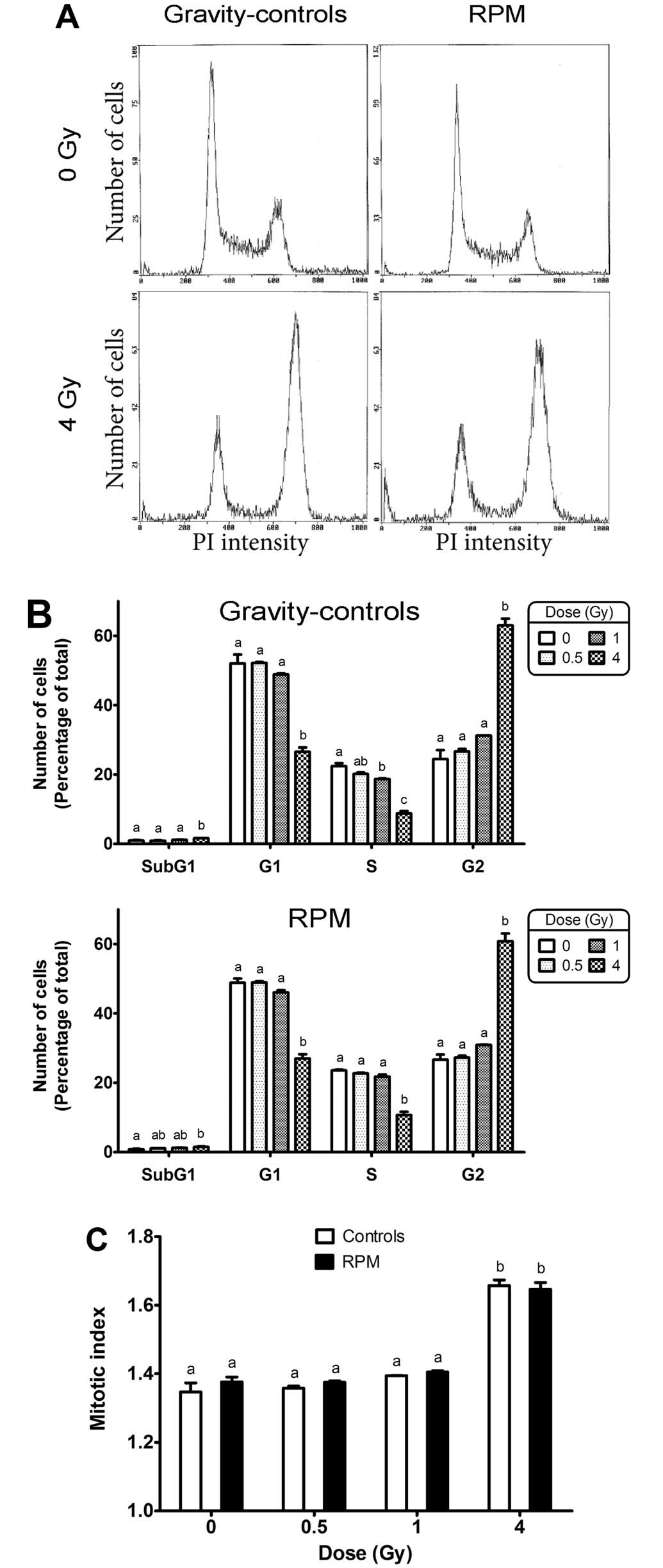
Cell cycle
The observations of increased cell area after
exposure to X-rays indicated a possible shift in cell cycle
distribution towards the G2 phase. To verify this, a cell cycle
analysis was performed by flow cytometry (see Fig. 4A for representative examples of
cell cycle distributions). Irradiation of gravity-control samples
with 4 Gy significantly increased the number of cells in G2 phase
and decreased the number of cells in G1 and S phases (Fig. 4B), which induced an increase of
the mitotic index (Fig. 4C). RPM
treatment had no significant effect on the number of cells in the
G2 phase, irrespective of the X-ray dose (Fig. 4B). Likewise, no difference was
observed in the mitotic index between RPM-treated and
gravity-control cells.
Apoptosis
The sub-G1 peak measured by cell cycle analysis
increased significantly after exposure of gravity-control cells to
4 Gy, but no significant effect of the RPM was detected on the
sub-G1 peak, irrespective of the dose (Fig. 4B). However, the accuracy of this
assay is limited by its sensitivity and the underestimation of
cells encountering apoptosis while undergoing the S and G2 phases
of the cell cycle. We therefore measured a second marker of
apoptosis based on caspase-3 activity that we had previously
described as a more suitable assay to measure radiation-induced
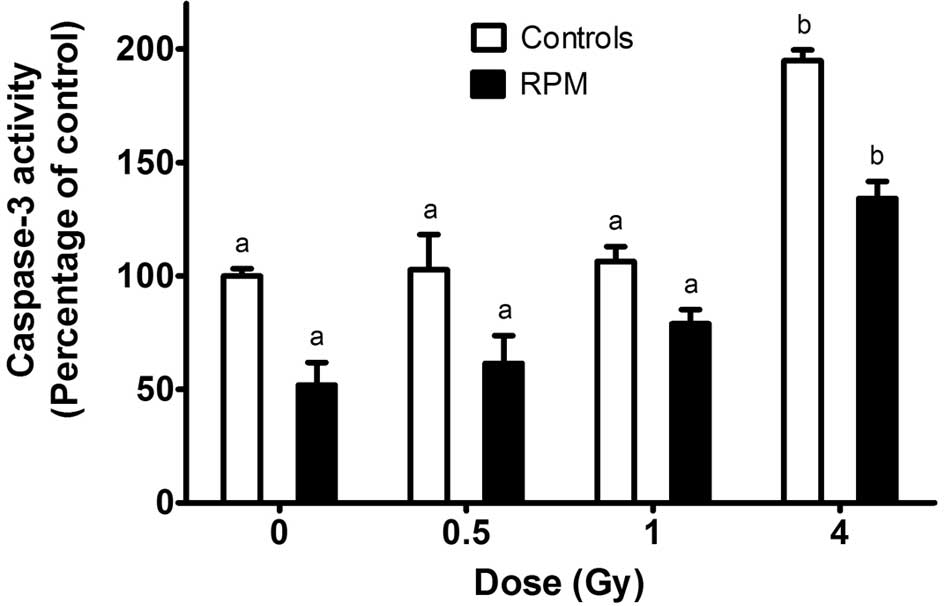
apoptosis in STO cells (17). In
the gravity-control cells, no increase in the caspase-3 activity
was observed following X-ray doses of 0.5 or 1 Gy while a 2-fold
increase in caspase-3 activity was observed after exposure of the
cells to 4 Gy (Fig. 5). Cells
from the samples submitted to simulated microgravity for 24 h
exhibited a significantly lower caspase-3 activity at all doses
compared to the gravity-controls. On the other hand, 4 Gy caused a
relative increase in caspase-3 activity which was similar to the
one observed for the gravity-control cells exposed to the same
dose.
Discussion
Microgravity is believed to induce various effects
on many cellular processes. Among others, alterations in the
cytoskeletal organization have been reported in many cell types,
including lymphocytes (18),
endothelial cells (19) and
osteoblasts (20). It has been
hypothesized that the cytoskeleton could act as a mechanosensitive
element that would trigger cell signaling changes leading to
differences in cell proliferation and cell death (18,20). A possible interaction between
irradiation and simulated microgravity has already been
investigated (21). Indeed,
microgravity-induced alterations in DNA damage repair kinetics
following irradiation could have dramatic consequences for cell
survival, which is of particular interest for space mission risk
assessment. Although Mognato et al (22) observed a slower re-joining of
radiation-induced DSBs and a significant increase in apoptosis in
human lymphocytes irradiated with an acute high dose of gamma-rays
(5 Gy) and subsequently submitted to the rotating wall vessel (RWV)
for 24 h (22), most studies
showed no difference in repair kinetics or cell death upon
simulated microgravity as reviewed by Manti (21). Our data showed that irradiated STO
cells which had been exposed to simulated microgravity immediately
after irradiation exhibited the same relative radiation-induced
increase of apoptosis as those which had not been exposed to
microgravity. On the other hand, we observed an overall decrease in
the level of apoptosis, evidenced by a lower level of caspase-3
activity, in RPM-treated cells with respect to gravity-controls,
irrespective of the irradiation dose. This indicates a lower
background level of apoptosis, but no change in radiation-induced
apoptosis. Apparently conflicting with this observation, no
difference was detected in the percentage of sub-G1 cells between
RPM and gravity-control conditions. However, sub-G1 peak assessment
is known to have a low specificity to detect apoptotic cells
(23).
In contrast to Canova et al (24), we observed no change in
radiation-induced G2 phase arrest between RPM-treated cells and
gravity-controls. Although a significant increase in cell area was
measured at all doses in gravity-controls, the radiation-induced G2
phase arrest was significant only at 4 Gy. On the other hand, no
increase in cell area was observed up to 1 Gy in RPM-treated cells.
Based on these observations, we hypothesize that RPM treatment
could protect STO cells from undergoing apoptosis after exposure to
moderate doses of radiation, possibly by decreasing the basal
levels of caspase-3 activity. Such decrease in apoptosis could have
dramatic consequences at the organismal level, by allowing
accumulation of cells with persistent DNA damage and/or mutations
(25). Indeed, an increase in
mutation frequency was observed in human lymphocytes
gamma-irradiated (1–3 Gy) and subsequently submitted to the RWV for
24 h (26).
In conclusion, we showed that simulated microgravity
(RPM) decreased the level of apoptosis in fetal skin fibroblasts.
This could have important consequences for mutation frequency in
cells subjected to the combination of microgravity and
irradiation.
Acknowledgements
This study was performed in the
context of the ESA Topical Team on ‘Developmental Biology in
Vertebrates’ and was financially supported by 2 PRODEX/ESA
contracts (C90-303 and C90-391). The financial support of the
Hercules Foundation to E.J.M. Van Damme (project AUGE/013) is also
gratefully acknowledged. The STO cell line was generously donated
by Professor Luc Leyns, Lab Cell Genetics, VUB, Brussels,
Belgium.
References
|
1.
|
O GoossensF VanhavereN LeysRadiation
dosimetry for microbial experiments in the International Space
Station using different etched track and luminescent
detectorsRadiat Prot
Dosimetry120433437200610.1093/rpd/nci65216644947
|
|
2.
|
P JacquetSensitivity of germ cells and
embryos to ionizing radiationJ Biol Regul Homeost
Agents18106114200415471212
|
|
3.
|
SV CostesA BoissiereS RavaniR RomanoB
ParvinMH Barcellos-HoffImaging features that discriminate between
foci induced by high- and low-LET radiation in human
fibroblastsRadiat Res165505515200610.1667/RR3538.116669704
|
|
4.
|
M BelliO SaporaMA TabocchiniMolecular
targets in cellular response to ionizing radiation and implications
in space radiation protectionJ Radiat
Res43SupplS13S19200210.1269/jrr.43.S1312793724
|
|
5.
|
B StenerlöwL EkerljungJ CarlssonJ
LennartssonRadiation induced DNA-damage/repair and associated
signaling pathwaysTargeted Radionuclide Tumor TherapyT StigbrandJ
CarlssonGP AdamsSpringer Science2492662008
|
|
6.
|
PL OliveJP BanathPhosphorylation of
histone H2AX as a measure of radiosensitivityInt J Radiat Oncol
Biol Phys58331335200410.1016/j.ijrobp.2003.09.02814751500
|
|
7.
|
IR RadfordThe level of induced DNA
double-strand breakage correlates with cell killing after
X-irradiationInt J Radiat Biol Relat Stud Phys Chem
Med484554198510.1080/095530085145510513874180
|
|
8.
|
AE RoncaMammalian development in spaceAdv
Space Biol Med9217251200310.1016/S1569-2574(03)09009-914631635
|
|
9.
|
RH HuijserDesktop RPM: New small size
microgravity simulator for the bioscience laboratoryFokker Space:
FS-MGR00-017, 2000.
|
|
10.
|
JJWA van LoonSome history and use of the
random positioning machine, RPM, in gravity related researchAdv
Space Res39116111652007
|
|
11.
|
A BorstJ van LoonTechnology and
developments for the Random positioning machine, RPMMicrogravity
Sci Technol21287292200910.1007/s12217-008-9043-2
|
|
12.
|
TF KraftJJ van LoonJZ KissPlastid position
in Arabidopsis columella cells is similar in microgravity
and on a random-positioning machinePlanta211415422200010987561
|
|
13.
|
A VillaS VersariJA MaierS BradamanteCell
behavior in simulated microgravity: a comparison of results
obtained with RWV and RPMGravit Space Biol
Bull188990200516038099
|
|
14.
|
LM WareAA AxelradInherited resistance to
N- and B-tropic murine leukemia viruses in vitro: evidence that
congenic mouse strains SIM and SIM. R differ at the Fv-1
locusVirology50339348197210.1016/0042-6822(72)90385-64344190
|
|
15.
|
KS KochKH SonR MaehrImmune-privileged
embryonic Swiss mouse STO and STO cell-derived progenitor cells:
major histocompatibility complex and cell differentiation antigen
expression patterns resemble those of human embryonic stem cell
linesImmunology11998115200610.1111/j.1365-2567.2006.02412.x
|
|
16.
|
Y ChanRP WalmsleyLearning and
understanding the Kruskal-Wallis one-way
analysis-of-variance-by-ranks test for differences among three or
more independent groupsPhys Ther771755176219979413454
|
|
17.
|
M BeckM MoreelsP JacquetP van OostveldtWH
De VosS BaatoutX-irradiation induces cell death in fetal
fibroblastsInt J Mol Med30114118201222505139
|
|
18.
|
A CogoliSignal transduction in T
lymphocytes in microgravityGravit Space Biol
Bull10516199711540120
|
|
19.
|
M InfangerP KossmehlM ShakibaeiInduction
of three-dimensional assembly and increase in apoptosis of human
endothelial cells by simulated microgravity: impact of vascular
endothelial growth
factorApoptosis11749764200610.1007/s10495-006-5697-7
|
|
20.
|
A GuignandonO AkhouayriY UssonFocal
contact clustering in osteoblastic cells under mechanical stresses:
microgravity and cyclic deformationCell Commun
Adhes106983200310.1080/cac.10.2.69.8314681058
|
|
21.
|
L MantiDoes reduced gravity alter cellular
response to ionizing radiation?Radiat Environ
Biophys4518200610.1007/s00411-006-0037-416523345
|
|
22.
|
M MognatoC GirardiS FabrisL CelottiDNA
repair in modeled microgravity: double strand break rejoining
activity in human lymphocytes irradiated with gamma-raysMutat
Res6633239200910.1016/j.mrfmmm.2009.01.00219428367
|
|
23.
|
GD WilsonB MarplesFlow cytometry in
radiation research: past, present and futureRadiat
Res168391403200710.1667/RR1042.117903043
|
|
24.
|
S CanovaF FiorasiM Mognato‘Modeled
microgravity’ affects cell response to ionizing radiation and
increases genomic damageRadiat Res1631911992005
|
|
25.
|
D RisinNR PellisModeled microgravity
inhibits apoptosis in peripheral blood lymphocytesIn Vitro Cell Dev
Biol
Anim376672200110.1290/1071-2690(2001)037%3C0066:MMIAIP%3E2.0.CO;211332739
|
|
26.
|
M MognatoL Celotti‘Modeled microgravity’
affects cell survival and HPRT mutant frequency, but not the
expression of DNA repair genes in human lymphocytes irradiated with
ionising radiationMutat Res5784174292005
|