Introduction
Although natural aging of the body may be considered
a fundamental biological process (1), it is not clear which molecular and
evolutionary genetic mechanisms trigger the progressive decline in
physiological functions and decrease the rate of reproduction
(2). Aging is most likely not due
to active gene programming (3),
but more likely based on evolved limitations in somatic maintenance
(4). In humans, one of the most
striking features of aging is a gradual reduction in skeletal
muscle mass and a concomitant decline in contractile strength
(5). This age-related progressive
loss of muscle mass and function has been termed sarcopenia and is
believed to be due to a multifactorial etiology (6–8).
As reviewed by Berger and Doherty (9), epidemiological studies of
age-related skeletal muscle wasting indicate that nearly half the
population over 75 years of age is suffering from some form of
sarcopenia leading in severe cases to loss of independence.
Although the findings from a large number of longitudinal and
cross-sectional studies of skeletal muscle aging (10–19) do not concur on the exact rate and
extent of contractile tissue loss (20), these investigations agree that
human aging is associated with a general impairment of structural
and functional elements of the musculoskeletal system (21). Since individual skeletal muscles
are differentially affected during the natural aging process
(22), one should be careful
about extrapolating age-related changes in a particular muscle
group to the aging process of the entire neuromuscular system.
Muscle wasting occurs in all aged individuals to a
varying degree (23), but this
degenerative process may be accelerated by a variety of
exacerbating factors, such as lack of physical activity, improper
nutritional intake, extensive pharmacotherapy and/or chronic
illness (24). For example,
extended periods of bed rest in elderly patients are clearly
related to inactivity-induced insulin resistance and further
complicate the sarcopenic syndrome (25). Advancing age is associated with
the loss of spinal motor neurons due to apoptosis (26), an impaired capacity for axonal
re-innervation of deinnervated muscle fibers (27) and age-induced low-grade
inflammation (28). Histological
hallmarks of sarcopenia are a reduction in fibre numbers, a decline
in fibre size and potential loss of entire motor units, whereby
preferential atrophy of type II fibres has been established as a
consequence of aging (21,27,29).
The diagnosis of sarcopenia is based on the combined presence of a
low muscle mass and a low gait speed (30–32). The clinical cut-off point for
sarcopenia is considered to be a percentage of muscle mass two
standard deviations below the mean measured in young adults of the
same gender and ethnic background, as well as a walking speed below
0.8 m/sec in the 4 m walking test (31).
Since a reduction in skeletal muscle mass is
pronounced in aged lower limb muscle groups, the human vastus
lateralis muscle has been the center of attention in numerous
studies on old age (33). Between
the ages of 20 and 80 years, the cross-sectional area of the
vastus lateralis muscle may be reduced by up to 40%
(34,35). Epidemiological studies have
indicated accelerated muscle wasting after the fifth decade with an
approximately 2% reduction in muscle mass per year (9). We carried out a comparative survey
of total extracts from middle-aged vs. aged vastus lateralis
muscle tissue. Fluorescence difference in-gel electrophoresis
(DIGE) in combination with mass spectrometry (MS) was employed to
study human muscle aging, since this advanced method of modern
protein biochemistry is capable of swiftly evaluating potential
changes in large numbers of protein species (36). Over the last decade, MS-based
proteomics has identified numerous novel protein factors involved
in myogenesis, muscle differentiation, fibre transitions and
various neuromuscular pathologies, as summarised in recent reviews
(37–39). With respect to the natural aging
process, proteomics technology has been applied to study altered
protein expression levels associated with cellular changes in
skeletal muscle tissues (40),
whereby most studies have focused on animal models of aging, such
as the senescent Wistar rat (41–51).
A previous proteomic survey of human muscle aging
has investigated differences in protein expression between young
adults (20–25 years) and aged (70–76 years) individuals (52). In order to build on these findings
and to determine potential changes after the fifth decade of life,
we used fluorescent tagging of the vastus lateralis muscle
proteome from middle-aged (47–62 years) vs. aged (76–82 years)
individuals and conducted a comprehensive gel electrophoresis-based
survey of human skeletal muscle aging. Densitometric scanning
revealed a differential expression pattern for 19 2-dimensional
(2D) protein spots and the subsequent mass spectrometric analysis
identified these muscle-associated proteins as being involved in
contraction, relaxation, ion homeostasis, cellular stress response
and fibre metabolism. Alterations in key muscle proteins from
vastus lateralis muscle support the idea of generally
perturbed protein expression patterns during human aging and agree
with the notion that sarcopenia is associated with the fast-to-slow
and glycolytic-to-oxidative transition processes (52–54). Of note, the muscle-specific
carbonic anhydrase (CA)3 isoform (EC 4.2.1.1) (55), which has not been previously
identified by proteomic surveys of human muscle aging (38,52), showed an increase in senescent
skeletal muscle by both MS-based proteomics and independent
immunoblot analysis. Elevated levels of CA3 may be indicative of an
increased demand for efficient CO2-removal during fibre
aging and/or age-related fibre type shifting to slower-contracting
muscle populations (54). The
establishment of a novel biomarker of muscle aging after the fifth
decade of life may be exploitable for the future development of a
more reliable assay to diagnose sarcopenia in old age.
Materials and methods
Materials
For the fluorescence 2D-DIGE analysis of aged human
skeletal muscles, immobilised pH gradient (IPG) drystrips pH 3–11,
CyDye DIGE Fluor minimal dyes Cy3 and Cy5, Coomassie Brilliant
Blue, ampholytes, acetonitrile, cover fluid, and the 2D-Clean-Up
kit were purchased from Amersham Biosciences/GE Healthcare (Little
Chalfont, UK). Ultrapure protogel acrylamide stock solutions were
from National Diagnostics (Atlanta, GA, USA). Sequencing
grade-modified trypsin for peptide generation was obtained from
Promega (Madison, WI, USA). LC-MS Formic acid and Chromasolv water
were purchased from Fluka Chemical Corp. (Milwaukee, WI, USA) and
spin filters were from Fisher Scientific (Loughborough, UK).
Chemiluminescence substrate, protease inhibitors and nitrocellulose
sheets were from Pierce and Warriner (Chester, UK), Roche
Diagnostics (Manheim, Germany) and Millipore (Bedford, MA, USA),
respectively. For immunoblotting, primary antibodies to the CA3
isoform were purchased from Abcam (Cambridge, UK) and secondary
peroxidase-conjugated antibodies were from Chemicon International,
Inc. (Temecula, CA, USA). Ultrapure lysine for quenching the DIGE
labelling reaction, DNase I and all general reagents were otained
from Sigma Chemical Co. (Dorset, UK).
Human skeletal muscle biopsy samples
Since the neuromuscular system of the vastus
lateralis muscle is intensively studied in human physiology and
biochemistry, and frequently targeted during biopsy procedures for
diagnostic purposes or routine biomedical investigations,
age-related changes in this muscle were studied by MS-based
proteomics. Two groupings of biopsy material representing healthy
middle-aged vs. healthy aged vastus lateralis muscle were
provided by the associated hospitals of the University of Bonn
according to German ethics regulations (56). Middle-aged specimens derived from
47-, 55-, 59- and 62-year-old muscle tissue are marked in this
study as samples MA1 to MA4. Aged specimens derived from 76-, 77-,
81- and 82-year-old muscle tissue are marked as samples OA1 to OA4.
Following biopsy, fresh tissue specimens were immediately frozen in
liquid nitrogen, transported on dry ice and then stored at −80°C
prior to usage.
Preparation of urea-soluble fraction of
skeletal muscle proteome
Freshly thawed skeletal muscle biopsy material (100
mg) was ground into a fine powder in the presence of liquid
nitrogen using a pestle and mortar. Pulverised human muscle
specimens of different ages were resuspended in 1 ml of ice-cold
buffer containing 7 M urea, 2 M thiourea, 65 mM CHAPS, 10 mM Trizma
base, 1% ampholytes pH 3–11, and 100 mM dithiothreitol. In order to
eliminate excessive viscosity of the protein extract due to large
amounts of muscle-derived DNA and to minimise protein degradation
due to the presence of muscle-associated proteases, the solution
was supplemented with 2 μl of DNase I (200 units; Sigma
Chemical Co.) per 100 μl buffer and a protease inhibitor
cocktail (Roche Diagnostics) (57). The muscle tissue homogenate was
gently mixed by vortexing and then placed on a bench top shaker for
2.5 h at room temperature to precipitate the total urea-soluble
protein complement. Following centrifugation of the suspension at
20,000 x g in an Eppendorf 5417R centrifuge (Eppendorf, Hamburg,
Germany) for 20 min, the protein-containing pellet was washed in 5
ml of ice-cold 100% acetone and thoroughly broken up by vortexing
and sonication. Using 80% acetone, washing and centrifugation was
repeated twice and the final protein precipitate was collected by
centrifugation and resuspended in 1 ml of the above described
buffer. Individual protein samples were incubated for 1 h at room
temperature with careful vortexing every 10 min for 5 sec.
Determination of protein concentration and removal of potentially
interfering contaminants was performed using the Bradford assay
system (58) and 2D-Clean-Up kit
from Amersham Biosciences/GE Healthcare (59), respectively. Finally, protein
pellets representing middle-aged and aged skeletal muscle protein
fractions were resuspended in DIGE lysis buffer (7 M urea, 2 M
thiourea, 30 mM Tris, 4% CHAPS, pH 8.5) and adjusted to a protein
concentration of 1 mg/ml.
Fluorescence DIGE analysis
The gel electrophoretic separation of fluorescently
tagged muscle proteins was performed with a total amount of 100
μg protein per 2D-DIGE gel. To keep potential impurities to
a minimum, electrophoretic separation steps were performed under
designated fume hoods. For each protein preparation derived from
differently aged vastus lateralis muscle, a 50 μg
protein aliquote was fluorescently-labelled with 200 pmol DIGE
fluor dyes for 30 min on ice and in the dark. Tissue extracts from
eight different muscle samples representing 47–82 years of age were
each labelled with Cy3 dye. In addition, pooled internal standards
were prepared with Cy5 dye, as previously described in detail
(60). Quenching of the
fluorescent labelling reaction was carried out with 10 mM lysine
for 10 min on ice. For separation in the first dimension,
isoelectric focusing (IEF) was carried out with 24 cm strips in an
Amersham IPGphor system using the following running conditions: 30
V for 2 h, 500 V for 2.5 h, 1,000 V for 1 h, 2,000 V for 1 h, 4000
V for 1 h, 6,000 V for 1 h, 800 V for 3 h, 500 V for 1.5 h and
finally 8,000 V for 2.5 h. Gel strips were then equilibrated for 20
min in reducing buffer containing 100 mM dithiothreitol, followed
by 10 min of alkylation in buffer containing 250 mM iodoacetamide.
For separation in the second dimension, slab gel electrophoresis
was carried out with an Amersham Ettan Dalt-twelve system, using
12.5% gels. Eight slab gels were run in parallel at 0.5 W/gel for
60 min and then 15 W/gel until the blue dye front had disappeared
from the bottom of the gel. Cy3- and Cy5-labelled muscle-associated
proteins were visualised with the assistance of an Amersham Typhoon
Trio variable mode imager. Individual 2D-DIGE gels were warped to a
single master gel prior to analysis. Potential changes in the
protein expression profile of middle-aged vs. aged skeletal muscle
samples were analysed using Progenesis SameSpots analysis software
from Nonlinear Dynamics (Newcastle upon Tyne, UK), using the
following parameters: n=4; t-test P<0.05; and a power value of
>0.8. Muscle proteins with a significantly changed density were
selected for tryptic digestion from Coomassie Brilliant
Blue-stained preparative gels (61).
Mass spectrometric identification of
skeletal muscle proteins
The mass spectrometric identification of proteins of
particular interest was carried out in a dedicated and
air-conditioned proteomics suite with semi-clean analytical status
to avoid potential contamination of samples. Protein spots were
digested by standardised in-gel trypsination to generate distinct
peptide populations (62). MS
analysis was performed on a Model 6340 Ion Trap LC/MS apparatus
from Agilent Technologies (Santa Clara, CA, USA). Spot excision,
washing, destaining and treatment with protease were performed by
previously optimised methods (57,60,61). Trypsin-generated peptide mixtures
were dried through vacuum centrifugation and then resuspended in
MS-grade distilled water and 0.1% (v/v) formic acid, spun down
through spin filter and added to LC-MS viles for identification by
ion trap LC-MS analysis. A nanoflow Aligent 1200 series system was
employed for peptide separation. Analytical samples were loaded
into the enrichment at a capillary flow rate set to 2 μl/min
with a mix of 0.1% (v/v) formic acid and 50% (v/v) acetonitrile and
formic acid at a ratio of 19:1. The voltage was set to 1,700 V.
Database searches were carried out with Mascot MS/MS Ion search
(Matrix Science, London, UK). All searches used ‘Homo
sapiens’ as a taxonomic category and the following parameters:
i) two missed cleavages by trypsin, ii) mass tolerance of precursor
ions ±2.5 Da and product ions ±0.7 Da, iii) carboxymethylated
cysteins fixed modification, and iv) oxidation of methionine as
variable modification. In addition, only hits with at least two
matched distinct peptides and a Mascot score of at least 44 were
considered significant proteomic findings.
Verification of age-related changes in
the expression of carbonic anhydrase by immunoblotting
In order to verify age-related changes in the
abundance of CA, immunoblot analysis was carried out to determine
the expression levels of the CA3 isoform in middle-aged vs. aged
vastus lateralis muscle preparations. The electrophoretic
transfer of muscle proteins to Immobilon NC-pure nitrocellulose
membranes was carried out for 1 h at 100 V and 4°C with a
Mini-Protean transfer system from Bio-Rad Laboratories
(Hemel-Hempstead, UK). Transfer efficiency was routinely evaluated
by reversible Ponceau S-Red staining of membrane sheets.
Nitrocellulose membranes were blocked for 1 h in 5% (w/v) fat-free
milk powder dissolved in phosphate-buffered saline [PBS; 50 mM
sodium phosphate, 0.9% (w/v) NaCl, pH 7.4]. Blocked and washed
membranes were incubated for 3 h at room temperature with
appropriately diluted primary antibody to the CA3 isoform, followed
by a gentle washing in blocking buffer and then incubated with an
appropriate dilution of peroxidase-conjugated secondary antibody
for 1 h at room temperature (57). Immuno-decorated membranes were
washed again in blocking solution and a final rinse in PBS.
Antibody-labelled protein bands were visualised with the
SuperSignal-type enhanced chemiluminescence kit from Pierce and
Warriner. Densitometric scanning of immunoblots was performed on a
300 S computing densitometer (Molecular Dynamics, Sunnyvale, CA,
USA) with ImageJ (NIH, USA) and GraphPad Prism (GraphPad Software,
Inc., San Diego, CA, USA) software.
Results
In order to evaluate age-dependent alterations in
the skeletal muscle proteome after the fifth decade of life, we
applied the fluorescence 2D-DIGE technique and MS for studying
vastus lateralis specimens from middle-aged vs. aged
individuals. In our current study, the comparative gel
electrophoresis-based proteomic study of crude muscle extracts
focused on urea-soluble proteins, which is clearly reflected by the
identification of mostly soluble and relatively abundant protein
species. Shown is an overview of the analytical DIGE gels used in
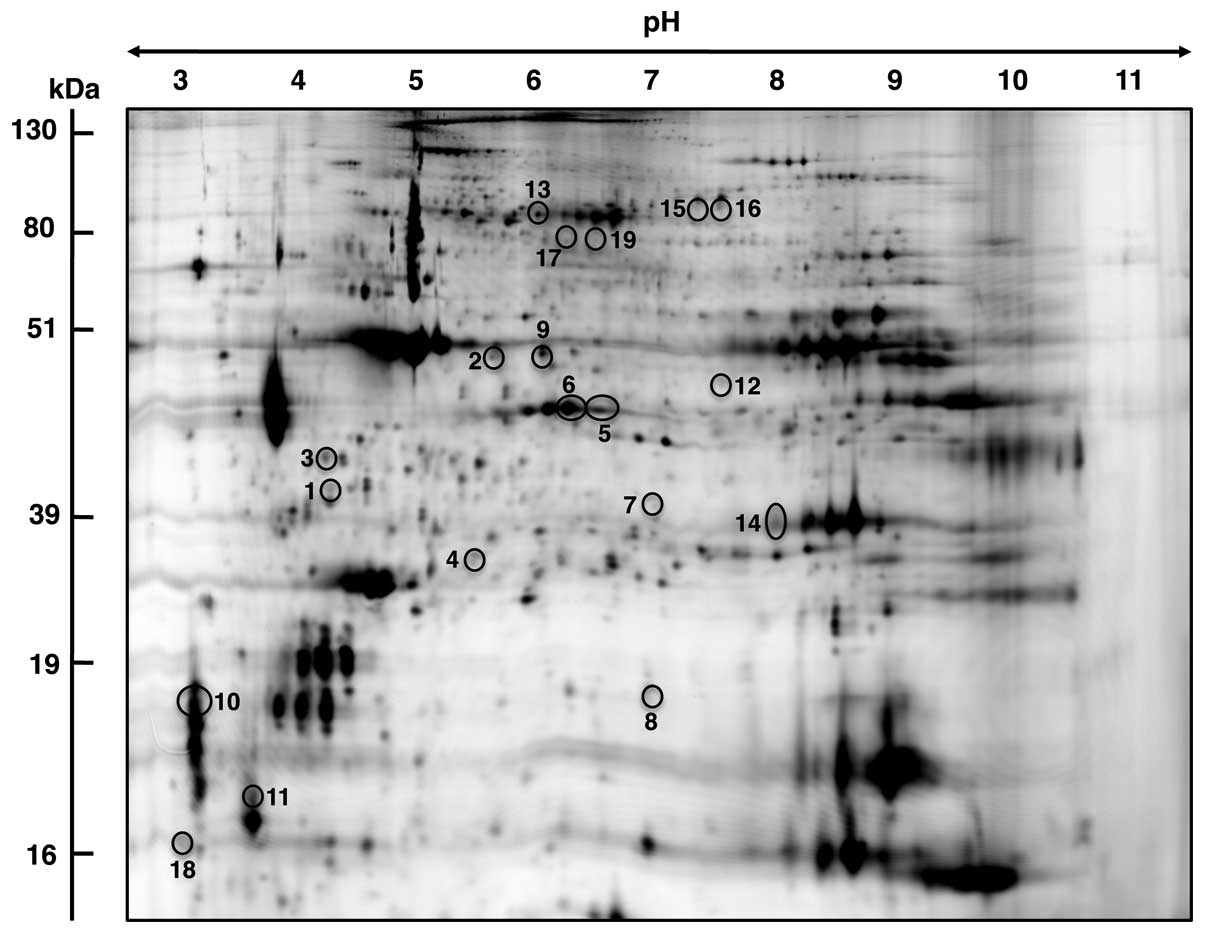
this proteomic study (Fig. 1), a
DIGE master gel outlining protein spots with a significant
age-related change in abundance (Fig.
2), a listing of changed proteins in senescent vastus
lateralis muscle as determined by MS analysis (Table I) and immunoblotting of changed
expression levels in a muscle-specific protein, the CA3 isoform
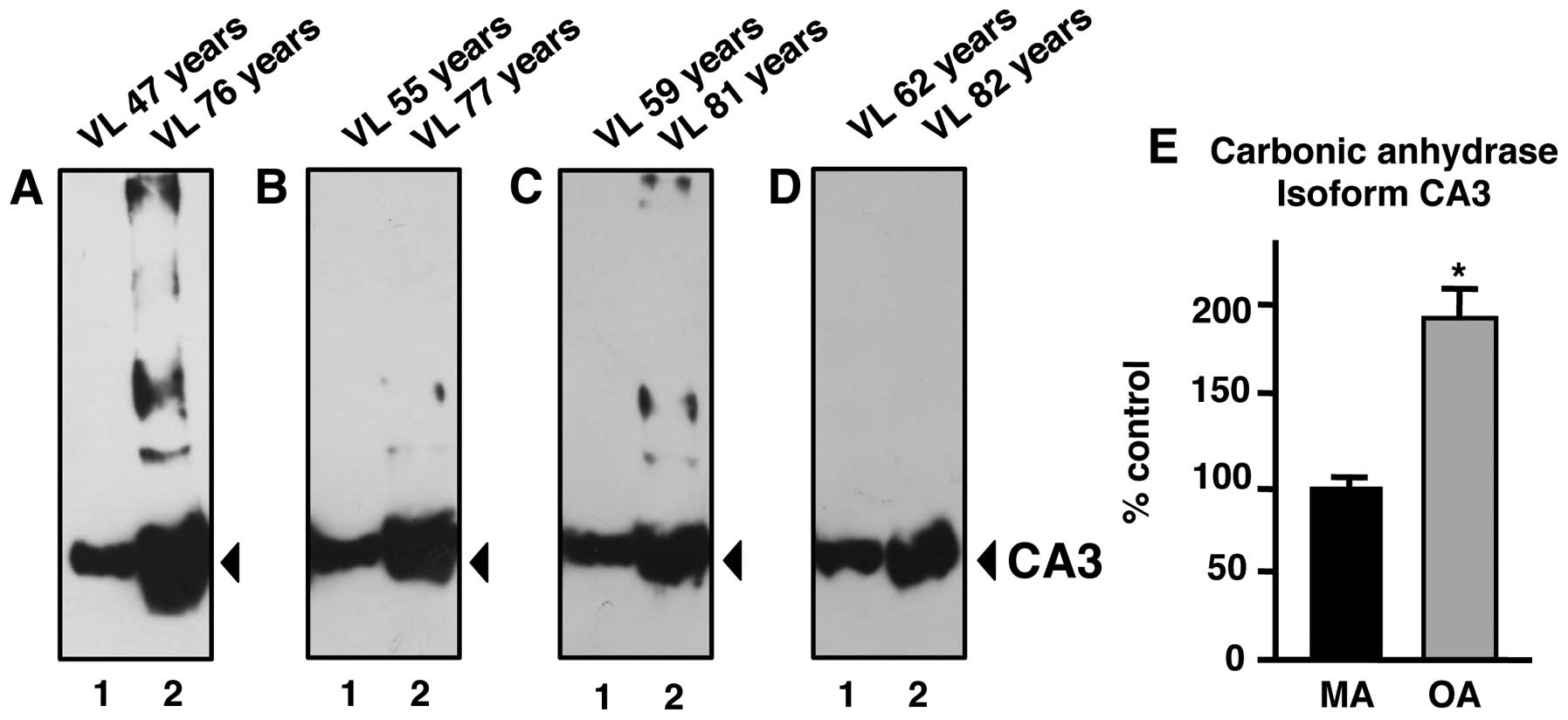
(Fig. 3). In order to correlate
MS-identified protein species listed in Table I with distinct protein spots of
altered concentration in the DIGE master gel shown in Fig. 2, the numbering system of proteins
in the table and corresponding gel is identical.
 | Table I.List of DIGE-identified proteins with
a changed abundance in middle aged vs. aged human vastus lateralis
muscle. |
Table I.
List of DIGE-identified proteins with
a changed abundance in middle aged vs. aged human vastus lateralis
muscle.
| Spot no. | Name of identified
protein | Accession no. | Molecular mass
(kDa) | Iso-electric point
pI | Mascot score | Coverage (%) | Peptides
matched | Anova | Fold change | Sequences |
|---|
| 1 | Actin, α skeletal
muscle | gi|33563240 | 42372 | 5.23 | 135 | 28 | 10 | 2.32E-04 | −3.8 | AGFAGDDAPR,
AVFPSIVGRPR, HQGVMVGMGQK, DSYVGDEAQSK, DSYVGDEAQSKR, LDLAGR,
DLTDYLMK, GYSFVTTAER, LCYVALDFENEMATAASSSSLEK,
SYELPDGQVITIGNER |
| 2 | Actin, α skeletal
muscle | gi|33563240 | 42372 | 5.23 | 435 | 36 | 11 | 3.05E-04 | −2.6 | AGFAGDDAPR,
DSYVGDEAQSK, DSYVGDEAQSKR, VAPEEHPTLLTEAPLNPK, DLTDYLMK,
GYSFVTTAER, LCYVALDFENEMATAASSSSLEK, SYELPDGQVITIGNER,
DLYANNVMSGGTTMYPGIADR, EITALAPSTMK, IIAPPER |
| 3 | Annexin A5 | gi|6753060 | 35788 | 4.83 | 46 | 5 | 2 | 0.002 | −2.5 | VLTEIIASR,
MLVVLLQANR |
| 4 | DJ-1 protein | gi|55741460 | 20240 | 6.32 | 47 | 36 | 6 | 7.00E-03 | −2.1 | ALVILAK,
GAEEMETVIPVDVMR, VTVAGLAGK, GLIAAICAGPTALLAHEVGFGCK, DGLILTSR,
APLVLKD |
| 5 | Troponin T, slow
skeletal muscle | gi|3449358 | 29992 | 6.34 | 198 | 19 | 4 | 1.00E-03 | −2 | VDFDDIHR,
DLLELQTLIDVHFEQR, VLSNMGAHFGGYLVK, YEINVLYNR |
| 6 | Troponin T, slow
skeletal muscle | gi|3449358 | 29992 | 6.34 | 223 | 22 | 6 | 4.00E-03 | −2 | IPEGERVDFDDIHR,
VDFDDIHR, DLLELQTLIDVHFEQR, KVLSNMGAHFGGYLVK, VLSNMGAHFGGYLVK,
YEINVLYNR |
| 7 | Glycogen
phosphorylase | gi|6755256 | 97689 | 6.65 | 60 | 8 | 8 | 6.00E-03 | −1.9 | VIFLENYR,
VIPAADLSEQISTAGTEASGTGNMK, GYNAQEYYDR, GYNAQEYYDRIPELR, IPELR,
DIVNMLMHHDR, TIAQYAR, EIWGVEPSR |
| 8 | Heat shock protein
β-7 | gi|31542970 | 18660 | 5.95 | 44 | 19 | 3 | 1.20E-02 | −1.8 | ALPAQDPPMEK,
LAADGTVMNTFAHK, EDGSLTIR |
| 9 | Actin, β | gi|49868 | 39451 | 5.78 | 101 | 23 | 10 | 2.00E-03 | −1.7 | AVFPSIVGR,
HQGVMVGMGQK, DSYVGDEAQSK, DSYVGDEAQSKR, LDLAGR, DLTDYLMK,
SYELPDGQVITIGNER, EITALAPSTMK, IIAPPER, IIAPPERK |
| 10 | Troponin C,
slow | gi|6678369 | 18525 | 4.04 | 121 | 24 | 3 | 5.00E-03 | −1.7 | AAVEQLTEEQKNEFK,
GKSEEELSDLFR, NADGYIDLDELK |
| 11 | Myosin light chain
MLC2 | gi|199985 | 18870 | 4.71 | 100 | 40 | 6 | 6.00E-03 | −1.5 | DTFAALGR,
EAPGPINFTVFLTMFGEK, GADPEETILNAFK, VFDPEGK, EMLTTQAER,
NLVHIITHGEEKD |
| 12 | Acyl-CoA
dehydrogenase short chain | gi|192659 | 45208 | 8.96 | 153 | 12 | 4 | 1.30E-02 | 1.3 | GISAFLVPMPTPGLTLGK,
IAMQTLDMGR, LADMALALESAR, ITEIYEGTSEIQR |
| 13 | Heat shock-protein
Hsp70 | gi|31560686 | 69889 | 5.51 | 175 | 12 | 7 | 6.00E-03 | 1.4 | VEIIANDQGNR,
TTPSYVAFTDTER, EIAEAYLGGK, DAGTITGLNVLR, IINEPTAAAIAYGLDK,
FEELNADLFR, VCNPIISK |
| 14 | Carbonic anhydrase
3 | gi|31982861 | 29638 | 6.89 | 47 | 8 | 2 | 1.20E-02 | 1.5 | VVFDDTYDR,
YAAELHLVHWNPK |
| 15 | Creatine kinase
M-type | gi|6671762 | 43250 | 6.58 | 86 | 12 | 4 | 1.00E-02 | 1.6 | GGDDLDPNYVLSSR,
LSVEALNSLTGEFK, IEEIFK, GQSIDDMIPAQK |
| 16 | Succinate
dehydrogenase complex subunit A | gi|3851614 | 59266 | 6.16 | 77 | 4 | 2 | 1.30E-02 | 1.6 | LGANSLLDLVVFGR,
SMQNHAAVFR |
| 17 | Actin,
α-cardiac | gi|387090 | 42048 | 5.23 | 358 | 21 | 9 | 3.00E-03 | 1.7 | AGFAGDDAPR,
HQGVMVGMGQK, DSYVGDEAQSK, DSYVGDEAQSKR, DLTDYLMK, GYSFVTTAER,
SYELPDGQVITIGNER, CDIDIRK, IIAPPER |
| 18 | Calmodulin | gi|99032084 | 16696 | 4.09 | 59 | 47 | 6 | 5.00E-03 | 1.7 | EAFSLFDKDGDGTITTK,
MKDTDSEEEIR, DTDSEEEIREAFR, VFDKDGNGYISAAELR,DGNGYISAAELR,
EADIDGDGQVNYEEFVQMMTAK |
| 19 |
Phosphoglucomutase | gi|33416468 | 63705 | 6.02 | 69 | 9 | 5 | 4.35E-05 | 2 | VDLGVLGK,
SMPTSGALDR, FFGNLMDASK, YDYEEVEAEGANK, LSGTGSAGATIR |
Proteomic profiling of aging human vastus
lateralis muscle
In this study, high-resolution fluorescence 2D gel
electrophoresis, in combination with densitometric analysis using a
Typhoon Trio variable imager and image analysis with progenesis 2D
analysis software, resulted in the identification of 19 protein
spots with a significant change in concentration levels in
middle-aged vs. aged individuals. Shown are analytical DIGE gels of
protein extracts from 47-, 55-, 59- and 62-year-old muscle tissue
(Fig. 1A–D) and from 76-, 77-,
81-and 82-year-old muscle tissue (Fig. 1I–L), labelled with Cy3 dye. Pooled
standards, labelled with Cy5 dye, are depicted in Fig. 1E–H and M–P.
MS identification of skeletal muscle
proteins with an age-related change in concentration
A DIGE master gel with electrophoretically separated
human vastus lateralis muscle proteins is shown for a
molecular mass range of approximately 10–150 kDa and a pI
range of pH 3 to pH 11 (Fig. 2).
Muscle-associated proteins with a potential age-dependent
alteration in density ranged in molecular mass from apparent 16.7
kDa (calmodulin) to 97.7 kDa (glycogen phosphorylase) and covered a
pI range from approximately 4.09 (calmodulin) to 8.96
(Acyl-CoA dehydrogenase). MS analysis identified 19 proteins
species, which are listed in Table
I outlining protein name, protein accession number, molecular
mass, pI-value, Mascot score, percentage sequence coverage,
number of matched peptide sequences, Anova score, fold change of
individual proteins affected by muscle aging, and peptide sequences
used to identify proteins. The majority of muscle-associated
proteins identified by DIGE screening in combination with MS
analysis were shown to be associated with the actomyosin apparatus,
the cytoskeleton, metabolism, signalling and the cellular stress
response. A reduced density was determined for 11 proteins and 8
proteins showed an increase in their abundance.
The vastus lateralis muscle protein with the
highest fold decrease was shown in the muscle isoform of α-actin
(spots 1, 2). Phosphoglucomutase (spot 19) was identified as the
significantly increased enzyme in aged muscle tissue. In addition
to muscle α-actin, muscle β-actin (spot 9), Annexin (spot 3), DJ-1
protein (spot 4), troponin subunits (spots 5, 6, 10), glycogen
phosphorylase (spot 7), heat shock protein (Hsp) β-7 (spot 8), and
myosin light chain (spot 11) were decreased in senescent human
muscle. Besides phosphoglucomutase, increased muscle proteins were
identified as Acyl-CoA dehydrogenase (spot 12), Hsp70 (spot 13),
CA3 isoform (spot 14), muscle creatine kinase (spot 15), succinate
dehydrogenase (spot 16), cardiac α-actin (spot 17) and calmodulin
(spot 18).
In addition to the proteins listed in Table I, the DIGE method in combination
with MS analysis identified two other proteins with a changed
abundance in aged muscle; slow myosin light chain MLC3
(gi|33563264|; 22523 kDa, pI 5.03) and the voltage-dependent
anion-selective channel VDAC2 (gi|6755965|; 32351 kDa, pI
7.44). The contractile protein, MLC3, was identified by six
peptides (ALGQNPTQAEVLR, MMDFETFLPMLQHISK, NKDTGTYEDFVEGLR,
EGNGTVMGAELR, HVLATLGER and LTEDEVEK) with a 35% sequence coverage
and a Mascot score of 86, and the mitochondrial ion channel, VDAC2,
was identified by three peptides (LTFDTTFSPNTGK,
VNNSSLIGVGYTQTLRPGVK and LTLSALVDGKSFNAGGHK) with a 17% sequence
coverage and a Mascot score of 64. Although their identification
was based on a sufficient number of peptide sequences, the
DIGE-based determination of their respective 2D spots displayed
only minor fragments. Therefore, these proteins were not included
in the listing of the main findings of this proteomic survey of
human skeletal muscle aging.
Immunoblot analysis of CA3 in aging human
skeletal muscle
Although the DIGE technique represents one of the
most powerful comparative methods in modern biochemistry and MS
analysis is highly accurate in identifying distinct protein
species, we were interested in independently verifying the
differential expression of a muscle-specific protein identified in
this proteomic study on old age. Since the CA3 isoform is a
relatively muscle-specific and fibre type-specific enzyme,
comparative immunoblotting was carried out to confirm a key finding
of the proteomic data presented in this study. The increased
immuno-decoration of CA in aged vs. middle-aged specimens from
vastus lateralis muscle (Fig.
3A–D) is in agreement with the proteomic establishment of
higher levels of this muscle enzyme in senescent contractile tissue
(Fig. 2 and Table I). A comparison of 47- vs.
76-year-old, 55- vs. 77-year-old, 59- vs. 81-year-old and 62- vs.
82-year-old skeletal muscle specimens revealed a statistically
significant increase in CA3 isoform expression in senescent
skeletal muscle (Fig. 3E).
Discussion
The progressive decline in skeletal muscle mass and
the weakening of contractile strength is a major pathophysiological
feature of the aged neuromuscular system. Often the frailty
syndrome and muscular dysfunction present personal care problems
for elderly individuals, limiting their independence and requiring
them to seek outside help, despite other medical ailments (63). This warrants large-scale genomic,
proteomic and metabolomic surveys of aged skeletal muscle in order
to establish the underlying mechanisms of sarcopenia in old age.
Over the last few years, a considerable number of molecular
investigations involved in the genetic basis of sarcopenia have
identified a large collection of differentially expressed genes in
aged muscle tissue (64). Based
on these genetic findings, it is crucial to verify which of the
identified age-dependent alterations in gene expression patterns
translate into changed concentration levels and/or
post-translational modifications in distinct protein products that
markedly modify skeletal muscle functions. The MS-based proteomic
survey of human aging presented in this study focused on the
elucidation of potential alterations in the muscle protein
complement after the fifth decade of life. Our fluorescent DIGE
study of the vastus lateralis muscle compared middle-aged
(47–62 years) vs. aged (76–82 years) individuals in order to
supplement a set of previously determined proteomic data on the
same skeletal muscle from young adults (20–25 years) and aged
(70–76 years) individuals (52).
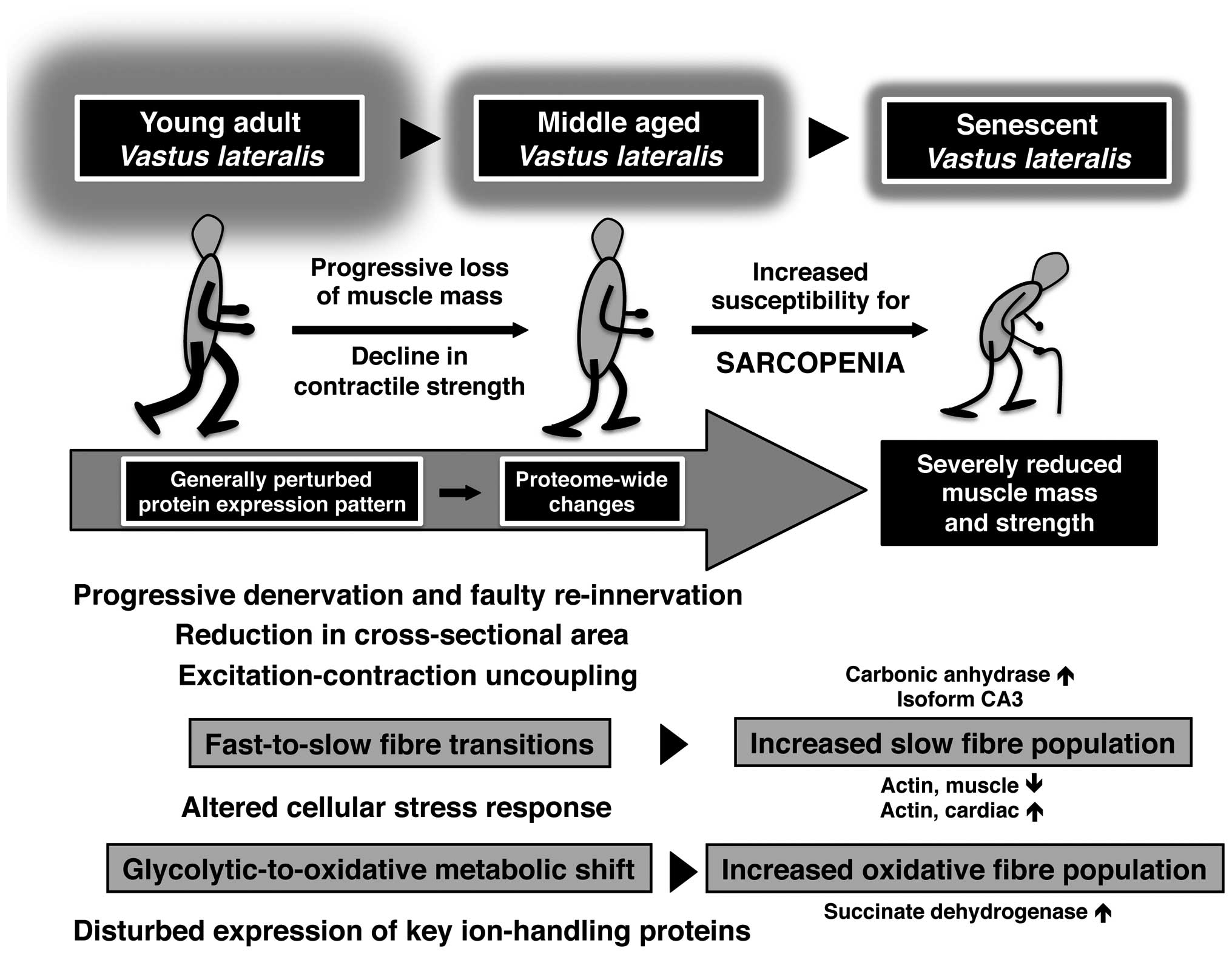
Based on the proteome-wide changes revealed by these extensive gel
electrophoresis-based surveys, the illustrative scheme in Fig. 4 summarises our knowledge of major
molecular and cellular events during human skeletal muscle aging.
Aged human vastus lateralis muscle tissue, which exhibits
progressive denervation, faulty re-innervation and reduction in
cross-sectional area (21,34),
is characterised by excitation-contraction uncoupling, an altered
cellular stress response, impaired ion homeostasis, fast-to-slow
fibre transitions and glycolytic-to-oxidative metabolic shifts
(52–54).
The comparative proteomic study of middle-aged vs.
aged human muscle identified age-related changes in the expression
of key proteins involved in the excitation-contraction-relaxation
cycle, ion handling, the cellular stress response and muscle fibre
bioenergetics. The apparent switch between the muscle isoform of
actin with its cardiac counter-part (65) in aged vastus lateralis
muscle supports the idea of a fast-to-slow transformation process
in the actomyosin apparatus during skeletal muscle aging (54). However, the decrease in slow
subunits of the regulatory element, troponin, and myosin light
chain 2 (MLC2) does not follow this general trend of isoform
switching in a slower-twitching aged muscle population. On the
other hand, increased levels of the mitochondrial enzyme, succinate
dehydrogenase, clearly support the metabolic concept of
glycolytic-to-oxidative transitions during fibre aging (53). This proteomic finding agrees with
the findings from numerous published studies on skeletal muscle
aging in both rodent models of sarcopenia and senescent human
muscle (46,50,52). Most likely the observed
contractile and metabolic transitions are not primary triggering
events that render an aged skeletal muscle more susceptible to the
loss of tissue mass and contractile strength, but a
pathophysiological consequence of age-related abnormalities
(54). It is important to stress
that histological studies have established that certain aged human
skeletal muscles exhibit a considerable reduction in fibre numbers,
a drastic decline in fibre size and high levels of atrophying
fast-twitching type II contractile fibres (21,27,29). Thus, the overall fast-to-slow
transformation and glycolytic-to-oxidative metabolic shift during
human skeletal muscle aging, as revealed by proteomics, possibly
reflects the preferential loss of fast vs. slower contracting
fibres in the senescent organism, and not an age-dependent
adaptation due to enhanced muscle plasticity.
In agreement with the concept of age-related fibre
type shifting, as exemplified by increased levels of slow
contractile proteins and mitochondrial marker enzymes, is the
observation of a higher concentration of the muscle-specific CA3
isoform (55) in aged human
vastus lateralis muscle. This relatively abundant enzyme has
not been previously identified by proteomic surveys of human
skeletal muscle aging (38,52). CAs are widely distributed
throughout the body and catalyse the reversible hydration of
CO2 (66). The various
isoforms play a crucial role in the acid-base balance,
CO2-removal and provision of CO2 for
metabolic processes, such as gluconeogenesis and the urea cycle, as
well as the regulatory processes of ion homeostasis (67–69). Mammalian skeletal muscles express
several CA isoforms in a fiber type-specific manner, whereby the
predominant CA3 isoform is mostly present in the cytosolic fraction
of type I and IIa fibers (55).
Changes in neuromuscular activity patterns, metabolic alterations,
stretch-induced hypertrophy and disuse atrophy have profound
effects on the expression of muscle CAs (70–72). Of note, soluble isoforms of muscle
CA are routinely used in clinical applications for assessing fibre
damage (73). It has been
suggested that the muscle-specific CA3 isoform presents a more
sensitive biomarker of muscle damage compared to creatine kinase in
neuromuscular disorders (74).
Thus, CA is a well-established biomarker routinely used in muscle
pathology (75), making it also
an ideal candidate for evaluating potential fibre type shifting
during muscle aging. The CA3 isoform levels were shown to be
increased in senescent human skeletal muscle by both proteomics and
independent immunoblot analysis. Although a higher concentration of
CA3 may be due to an increased demand for efficient
CO2-removal during fibre aging, the altered density of
this abundant fibre type marker is more likely indicative of
age-related fibre type shifting to slower-contracting muscle
populations. In contrast to elevated levels of CA3 in aging human
skeletal muscle, proteomic studies have recently established that
the same muscle-specific CA isoform is decreased in dystrophic
skeletal muscle (76) and
non-obese, diabetic skeletal muscle (77). This suggests that the
age-dependent increase in CA3 levels may be relatively specific
with respect to sarcopenia in old age.
Other altered muscle-associated proteins of interest
were Annexin A5, DJ-1 protein, glycogen phosphorylase, Hsp β-7 and
Hsp70, creatine kinase, calmodulin and phosphoglucomutase.
Increased levels of creatine kinase indicate a compensatory effect
on the creatine phosphate shuttle system in aged muscle (78). An altered concentration of Annexin
V and calmodulin may be due to altered ion handling during aging
(79). Since calmodulin is a
critical factor for Ca2+-sequestration and
Ca2+-cycling during excitation-contraction coupling
(80) and also plays a critical
role in skeletal muscle plasticity (81), a changed density of this abundant
Ca2+-binding protein presents an excellent new marker of
age-related alterations in ion homeostasis. Since many metabolic
enzymes involved in glycolysis, gluconeogenesis and glycogen
metabolism are multi-functional (82), the alterations in
phosphoglucomutase and glycogen phosphorylase during muscle aging
are difficult to interpret. Both enzymes are of crucial importance
for the bioenergetic utilization of muscle glucose.
Phosphoglucomutase catalyses the inter-conversion of
glucose-1-phosphate and glucose 6-phosphate, and glycogen
phosphorylase facilitates the phosphorolytic cleavage of a
glucosyl-residue from the glycogen polymer (83). The decreased levels of glycogen
phosphorylase and increased density of phosphoglucomutase indicate
altered flux rates of glucose and glycogen metabolism in senescent
fibres. It has previously been shown that proteins involved in the
cellular stress response are changed in various neuromuscular
disorders (84), including
sarcopenia in old age (40). The
expression levels of various molecular chaperones have been shown
to be altered after the fifth decade of life. However, distinct
differences appear to exist between rodent models of aging and
senescent human muscle (43). Of
note, the mostly uncharacterised DJ-1 protein, which has been
identified in this study by MS analysis, has previously been shown
to play a role in neuro-degeneration (85). This makes this muscle-associated
protein a potential new marker of sarcopenia.
In conclusion, aging of the human vastus
lateralis muscle is associated with a plethora of proteome-wide
changes, especially affecting fibre contraction, ion homeostasis,
muscle metabolism and the cellular stress response. These proteomic
findings are in agreement with findings from previous histological,
physiological and biochemical investigations that have established
cycles of denervation and impaired re-innervation causing the loss
of entire motor units and muscular atrophy, excitation-contraction
uncoupling at the triad junction, altered functioning of the
actomyosin apparatus resulting in weaker contractility, impaired
regulation of bioenergetic processes, disturbed ion handling and an
altered cellular stress response by molecular chaperones.
Importantly, the proteomic data presented in this study, are in
agreement with altered levels of key metabolic enzymes and
contractile elements that suggest a fast-to-slow transition of the
contractile apparatus and a glycolytic-to-oxidative shift in
metabolism during human muscle aging (54). The establishment of a novel
biomarker signature of muscle aging after the fifth decade of life
might be exploitable for the future design of a more reliable assay
to diagnose sarcopenia of old age.
Acknowledgements
The present study was supported by
grants from the Science Foundation Ireland, the Health Research
Board and the Higher Education Authority. We thank Helen Kennelly
for providing excellent technical support.
Abbreviations:
|
2D
|
2-dimensional;
|
|
CA
|
carbonic anhydrase;
|
|
DIGE
|
difference in-gel electrophoresis;
|
|
Hsp
|
heat shock protein;
|
|
IEF
|
isoelectric focusing;
|
|
MS
|
mass spectrometry;
|
|
SDS
|
sodium dodecyl sulfate;
|
|
PAGE
|
polyacrylamide gel
electrophoresis;
|
|
PBS
|
phosphate-buffered saline
|
References
|
1.
|
J VijgJY WeiUnderstanding the biology of
aging: the key to prevention and therapyJ Am Geriatr
Soc4342643419957706635
|
|
2.
|
T FlattPS SchmidtIntegrating evolutionary
and molecular genetics of agingBiochim Biophys
Acta1790951962200910.1016/j.bbagen.2009.07.01019619612
|
|
3.
|
TB KirkwoodSN AustadWhy do we
age?Nature408233238200010.1038/3504168211089980
|
|
4.
|
TB KirkwoodS MelovOn the
programmed/non-programmed nature of ageing within the life
historyCurr Biol21R701R707201110.1016/j.cub.2011.07.02021959160
|
|
5.
|
GS LynchSarcopenia - Age-Related Muscle
Wasting and Weakness: Mechanisms and TreatmentsSpringerNew
York480201110.1007/978-90-481-9713-2
|
|
6.
|
LV ThompsonAge-related muscle
dysfunctionExp
Gerontol44106111200910.1016/j.exger.2008.05.00318657920
|
|
7.
|
WJ EvansSkeletal muscle loss: cachexia,
sarcopenia, and inactivityAm J Clin
Nutr911123S1127S201010.3945/ajcn.2010.28608A20164314
|
|
8.
|
DR ThomasSarcopeniaClin Geriatr
Med26331346201010.1016/j.cger.2010.02.012
|
|
9.
|
MJ BergerTJ DohertySarcopenia: prevalence,
mechanisms, and functional consequencesInterdiscip Top
Gerontol3794114201010.1159/00031999720703058
|
|
10.
|
GB ForbesJC ReinaAdult lean body mass
declines with age: some longitudinal
observationsMetabolism19653663197010.1016/0026-0495(70)90062-45459997
|
|
11.
|
RN BaumgartnerPM StauberD McHughKM
KoehlerPJ GarryCross-sectional age differences in body composition
in persons 60+ years of ageJ Gerontol A Biol Sci Med
Sci50M307M31619957583802
|
|
12.
|
RS LindleEJ MetterNA LynchJL FlegJL
FozardJ TobinTA RoyBF HurleyAge and gender comparisons of muscle
strength in 654 women and men aged 20–93 yrJ Appl
Physiol831581158719979375323
|
|
13.
|
RN BaumgartnerKM KoehlerD GallagherL
RomeroSB HeymsfieldRR RossPJ GarryRD LindemanEpidemiology of
sarcopenia among the elderly in New MexicoAm J
Epidemiol147755763199810.1093/oxfordjournals.aje.a0095209554417
|
|
14.
|
LJ Melton IIIS KhoslaCS CrowsonMK
O’ConnorWM O’FallonBL RiggsEpidemiology of sarcopeniaJ Am Geriatr
Soc486256302000
|
|
15.
|
I JanssenSB HeymsfieldR RossLow relative
skeletal muscle mass (sarcopenia) in older persons is associated
with functional impairment and physical disabilityJ Am Geriatr
Soc50889896200210.1046/j.1532-5415.2002.50216.x12028177
|
|
16.
|
BH GoodpasterSW ParkTB HarrisSB
KritchevskyM NevittAV SchwartzEM SimonsickFA TylavskyM VisserAB
NewmanThe loss of skeletal muscle strength, mass, and quality in
older adults: the health, aging and body composition studyJ
Gerontol A Biol Sci Med
Sci6110591064200610.1093/gerona/61.10.105917077199
|
|
17.
|
MJ DelmonicoTB HarrisM VisserSW ParkMB
ConroyP Velasquez-MieyerR BoudreauTM ManiniM NevittAB NewmanBH
GoodpasterHealth, aging, and body. Longitudinal study of muscle
strength, quality, and adipose tissue infiltrationAm J Clin
Nutr9015791585200910.3945/ajcn.2009.2804719864405
|
|
18.
|
KK HedayatiM DittmarPrevalence of
sarcopenia among older community-dwelling people with normal health
and nutritional stateEcol Food
Nutr49110128201010.1080/0367024090354115421883084
|
|
19.
|
HP PatelHE SyddallHJ MartinCE StewartC
CooperAA SayerHertfordshire sarcopenia study: design and methodsBMC
Geriatr1043201010.1186/1471-2318-10-4320587018
|
|
20.
|
DN ProctorPC O’BrienEJ AtkinsonKS
NairComparison of techniques to estimate total body skeletal muscle
mass in people of different age groupsAm J
Physiol277E489E495199910484361
|
|
21.
|
AA VandervoortAging of the human
neuromuscular systemMuscle
Nerve251725200210.1002/mus.121511754180
|
|
22.
|
WR FronteraKF ReidEM PhillipsLS
KrivickasVA HughesR RoubenoffFA FieldingMuscle fiber size and
function in elderly humans: a longitudinal studyJ Appl
Physiol105637642200810.1152/japplphysiol.90332.200818556434
|
|
23.
|
JA FaulknerLM LarkinDR ClaflinSV
BrooksAge-related changes in the structure and function of skeletal
musclesClin Exp Pharmacol
Physiol3410911096200710.1111/j.1440-1681.2007.04752.x17880359
|
|
24.
|
D ScottL BlizzardJ FellG JonesThe
epidemiology of sarcopenia in community living older adults: what
role does lifestyle play?J Cachexia Sarcopenia
Muscle2125134201110.1007/s13539-011-0036-421966639
|
|
25.
|
RH CokerRR WolfeBedrest and sarcopeniaCurr
Opin Clin Nutr Metab Care15711201210.1097/MCO.0b013e32834da629
|
|
26.
|
P AagaardC SuettaP CaserottiSP MagnussonM
KjaerRole of the nervous system in sarcopenia and muscle atrophy
with aging: strength training as a countermeasureScand J Med Sci
Sports204964201010.1111/j.1600-0838.2009.01084.x20487503
|
|
27.
|
E EdstromM AltunE BergmanH JohnsonS
KullbergV Ramirez-LeonB UlfhakeFactors contributing to
neuromuscular impairment and sarcopenia during agingPhysiol
Behav92129135200710.1016/j.physbeh.2007.05.04017585972
|
|
28.
|
I BeyerT MetsI BautmansChronic low-grade
inflammation and age-related sarcopeniaCurr Opin Clin Nutr Metab
Care151222201210.1097/MCO.0b013e32834dd29722108098
|
|
29.
|
L LarssonB SjodinJ KarlssonHistochemical
and biochemical changes in human skeletal muscle with age in
sedentary males, age 22–65 yearsActa Physiol
Scand10331391978208350
|
|
30.
|
M MuscaritoliSD AnkerJ ArgilésZ AversaJM
BauerG BioloY BoirieI BosaeusT CederholmP CostelliConsensus
definition of sarcopenia, cachexia and pre-cachexia: joint document
elaborated by Special Interest Groups (SIG) ‘cachexia-anorexia in
chronic wasting diseases’ and ‘nutrition in geriatrics’Clin
Nutr29154159201020060626
|
|
31.
|
AJ Cruz-JentoftJP BaeyensJM BauerY BoirieT
CederholmF LandiFC MartinJP MichelY RollandSM SchneiderSarcopenia:
European consensus on definition and diagnosis: Report of the
European Working Group on Sarcopenia in Older PeopleAge
Ageing39412423201010.1093/ageing/afq03420392703
|
|
32.
|
AJ Cruz-JentoftF LandiE TopinkovaJP
MichelUnderstanding sarcopenia as a geriatric syndromeCurr Opin
Clin Nutr Metab
Care1317201010.1097/MCO.0b013e328333c1c119915458
|
|
33.
|
H PatelHE SyddallHJ MartinC CooperC
StewartAA SayerThe feasibility and acceptability of muscle biopsy
in epidemiological studies: findings from the Hertfordshire
Sarcopenia Study (HSS)J Nutr Health
Aging151015201110.1007/s12603-011-0006-821267515
|
|
34.
|
J LexellCC TaylorM SjostromWhat is the
cause of the ageing atrophy? Total number, size and proportion of
different fiber types studied in whole vastus lateralis
muscle from 15- to 83-year-old menJ Neurol
Sci8427529419883379447
|
|
35.
|
J LexellHuman aging, muscle mass, and
fiber type compositionJ Gerontol A Biol Sci Med Sci5011161995
|
|
36.
|
C LewisP DoranK OhlendieckProteomic
analysis of dystrophic muscleMethods Mol
Biol798357369201210.1007/978-1-61779-343-1_2022130847
|
|
37.
|
K OhlendieckProteomics of skeletal muscle
differentiation, neuromuscular disorders and fiber agingExpert Rev
Proteomics7283296201010.1586/epr.10.220377394
|
|
38.
|
C GelfiM VassoP CerretelliDiversity of
human skeletal muscle in health and disease: contribution of
proteomicsJ
Proteomics74774795201110.1016/j.jprot.2011.02.02821414428
|
|
39.
|
K OhlendieckSkeletal muscle proteomics:
current approaches, technical challenges and emerging
techniquesSkelet Muscle16201110.1186/2044-5040-1-621798084
|
|
40.
|
P DoranP DonoghueK O’ConnellJ GannonK
OhlendieckProteomics of skeletal muscle
agingProteomics99891003200910.1002/pmic.20080036519180535
|
|
41.
|
I PiecA ListratJ AlliotC ChambonRG TaylorD
BechetDifferential proteome analysis of aging in rat skeletal
muscleFASEB J1911431145200515831715
|
|
42.
|
K O’ConnellJ GannonP DoranK
OhlendieckProteomic profiling reveals a severely perturbed protein
expression pattern in aged skeletal muscleInt J Mol
Med20145153200717611631
|
|
43.
|
D DoranJ GannonK O’ConnellK
OhlendieckAging skeletal muscle shows a drastic increase in the
small heat shock proteins αB-crystallin/HspB5 and cvHsp/HspB7Eur J
Cell Biol86629640200717761354
|
|
44.
|
J GannonL StauntonK O’ConnellP DoranK
OhlendieckPhosphoproteomic analysis of aged skeletal muscleInt J
Mol Med2233422008
|
|
45.
|
K O’ConnellP DoranJ GannonK
OhlendieckLectin-based proteomic profiling of aged skeletal muscle:
decreased pyruvate kinase isozyme M1 exhibits drastically increased
levels of N-glycosylationEur J Cell Biol877938052008
|
|
46.
|
P DoranK O’ConnellJ GannonM KavanaghK
OhlendieckOpposite pathobiochemical fate of pyruvate kinase and
adenylate kinase in aged rat skeletal muscle as revealed by
proteomic DIGE
analysisProteomics8364377200810.1002/pmic.20070047518050275
|
|
47.
|
D CapitanioM VassoC FaniaM MoriggiA
ViganoP ProcacciV MagnaghiC GelfiComparative proteomic profile of
rat sciatic nerve and gastrocnemius muscle tissues in ageing by 2-D
DIGEProteomics920042020200910.1002/pmic.20070116219333999
|
|
48.
|
J GannonP DoranA KirwanK OhlendieckDrastic
increase of myosin light chain MLC-2 in senescent skeletal muscle
indicates fast-to-slow fibre transition in sarcopenia of old ageEur
J Cell Biol88685700200910.1016/j.ejcb.2009.06.00419616867
|
|
49.
|
A LombardiE SilvestriF CioffiR SeneseA
LanniF GogliaP de LangeM MorenoDefining the transcriptomic and
proteomic profiles of rat ageing skeletal muscle by the use of a
cDNA array, 2D- and Blue native-PAGE approachJ
Proteomics72708721200910.1016/j.jprot.2009.02.00719268720
|
|
50.
|
K O’ConnellK OhlendieckProteomic DIGE
analysis of the mitochondria-enriched fraction from aged rat
skeletal muscleProteomics955095524200919834913
|
|
51.
|
P DonoghueL StauntonE MullenG ManningK
OhlendieckDIGE analysis of rat skeletal muscle proteins using
nonionic detergent phase extraction of young adult vs. aged
gastrocnemius tissueJ
Proteomics714411453201010.1016/j.jprot.2010.01.01420153846
|
|
52.
|
C GelfiA ViganoM RipamontiA PontoglioS
BegumMA PellegrinoB GrassiR BottinelliR WaitP CerretelliThe human
muscle proteome in agingJ Proteome
Res513441353200610.1021/pr050414x16739986
|
|
53.
|
L StauntonK O’ConnellK OhlendieckProteomic
profiling of mitochondrial enzymes during skeletal muscle agingJ
Aging Res2011908035201110.4061/2011/90803521437005
|
|
54.
|
K OhlendieckProteomic profiling of
fast-to-slow muscle transitions during agingFront
Physiol2105201122207852
|
|
55.
|
P FremontPM CharestC CotePA RogersCarbonic
anhydrase III in skeletal muscle fibers: an immunocytochemical and
biochemical studyJ Histochem
Cytochem36775782198810.1177/36.7.31334073133407
|
|
56.
|
R SchäferU KnaufM ZweyerO HögemeierF de
GuarriniX LiuHJ EichhornFW KochRR MundegarI ErzenA WernigAge
dependence of the human skeletal muscle stem cell in forming muscle
tissueArtif Organs30130140200616480387
|
|
57.
|
L StauntonH JockuschC WiegandT AlbrechtK
OhlendieckIdentification of secondary effects of hyperexcitability
by proteomic profiling of myotonic mouse muscleMol
Biosyst724802489201110.1039/c1mb05043e21629954
|
|
58.
|
MM BradfordA rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye bindingAnal
Biochem72248254197610.1016/0003-2697(76)90527-3942051
|
|
59.
|
T RabilloudM ChevalletS LucheC
LelongTwo-dimensional gel electrophoresis in proteomics: Past,
present and futureJ
Proteomics7320642077201010.1016/j.jprot.2010.05.01620685252
|
|
60.
|
L StauntonH JockuschK OhlendieckProteomic
analysis of muscle affected by motor neuron degeneration: the
wobbler mouse model of amyotrophic lateral sclerosisBiochem Biophys
Res Commun406595600201110.1016/j.bbrc.2011.02.09921354103
|
|
61.
|
C LewisK OhlendieckMass spectrometric
identification of dystrophin isoform Dp427 by on-membrane digestion
of sarcolemma from skeletal muscleAnal
Biochem404197203201010.1016/j.ab.2010.05.01720507823
|
|
62.
|
A ShevchenkoH TomasJ HavlisJV OlsenM
MannIn-gel digestion for mass spectrometric characterization of
proteins and proteomesNat
Protoc128562860200610.1038/nprot.2006.46817406544
|
|
63.
|
T LangT StreeperP CawthonK BaldwinDR
TaaffeTB HarrisSarcopenia: etiology, clinical consequences,
intervention, and assessmentOsteoporos
Int21543559201010.1007/s00198-009-1059-y19779761
|
|
64.
|
LJ TanSL LiuSF LeiCJ PapasianHW
DengMolecular genetic studies of gene identification for
sarcopeniaHum Genet131131201210.1007/s00439-011-1040-721706341
|
|
65.
|
SY KhaitlinaFunctional specificity of
actin isoformsInt Rev
Cytol2023598200110.1016/S0074-7696(01)02003-411061563
|
|
66.
|
C GeersG GrosCarbon dioxide transport and
carbonic anhydrase in blood and musclePhysiol
Rev80681715200010747205
|
|
67.
|
P WetzelG GrosInhibition and kinetic
properties of membrane-bound carbonic anhydrases in rabbit skeletal
musclesArch Biochem
Biophys356151158199810.1006/abbi.1998.07629705205
|
|
68.
|
P WetzelT KleinkeS PapadopoulosG
GrosInhibition of muscle carbonic anhydrase slows the
Ca2+ transient in rat skeletal muscle fibersAm J
Physiol283C1242C1253200210.1152/ajpcell.00106.200212225987
|
|
69.
|
P FremontH RiverinJ FrenettePA RogersC
CoteFatigue and recovery of rat soleus muscle are influenced by
inhibition of an intracellular carbonic anhydrase isoformAm J
Physiol260R615R62119912001010
|
|
70.
|
C BrownsonH IsenbergW BrownS SalmonsY
EdwardsChanges in skeletal muscle gene transcription induced by
chronic stimulationMuscle
Nerve1111831189198810.1002/mus.8801111133147374
|
|
71.
|
C BrownsonP LoughnaAlterations in the mRNA
levels of two metabolic enzymes in rat skeletal muscle during
stretch-induced hypertrophy and disuse atrophyPfluegers
Arch431990992199610.1007/s0042400500978927521
|
|
72.
|
CH CoteF AmbrosioG PerreaultMetabolic and
contractile influence of carbonic anhydrase III in skeletal muscle
is age dependentAm J Physiol276R559R56519999950937
|
|
73.
|
AH WuMB PerrymanClinical applications of
muscle enzymes and proteinsCurr Opin
Rheumatol481582019921457275
|
|
74.
|
HK VäänänenTE TakalaU TolonenJ VuoriVV
MyllyläMuscle-specific carbonic anhydrase III is a more sensitive
marker of muscle damage than creatine kinase in neuromuscular
disordersArch Neurol451254125619883142447
|
|
75.
|
P BrancaccioG LippiN MaffulliBiochemical
markers of muscular damageClin Chem Lab
Med48757767201010.1515/CCLM.2010.179
|
|
76.
|
P DoranG MartinP DowlingH JockuschK
OhlendieckProteome analysis of the dystrophin-deficient MDX
diaphragm reveals a drastic increase in the heat shock protein
cvHSPProteomics646104621200610.1002/pmic.20060008216835851
|
|
77.
|
E MullenK OhlendieckProteomic profiling of
non-obese type 2 diabetic skeletal muscleInt J Mol
Med25445458201020127051
|
|
78.
|
T WallimannM Tokarska-SchlattnerU
SchlattnerThe creatine kinase system and pleiotropic effects of
creatineAmino
Acids4012711296201110.1007/s00726-011-0877-321448658
|
|
79.
|
M YanezJ Gil-LongoM Campos-ToimilCalcium
binding proteinsAdv Exp Med
Biol740461482201210.1007/978-94-007-2888-2_19
|
|
80.
|
W TangS SencerSL HamiltonCalmodulin
modulation of proteins involved in excitation-contraction
couplingFront Biosci7d1583d1589200210.2741/tang12045019
|
|
81.
|
P TaviH WesterbladThe role of in vivo
Ca²+ signals acting on
Ca²+-calmodulin-dependent proteins for skeletal muscle
plasticityJ Physiol589502150312011
|
|
82.
|
K OhlendieckProteomics of skeletal muscle
glycolysisBiochim Biophys
Acta180420892101201010.1016/j.bbapap.2010.08.00120709194
|
|
83.
|
TE JensenEA RichterRegulation of glucose
and glycogen metabolism during and after exerciseJ
Physiol59010691076201210.1113/jphysiol.2011.22497222199166
|
|
84.
|
RN NishimuraFR SharpHeat shock proteins
and neuromuscular diseaseMuscle
Nerve32693709200510.1002/mus.2037315962334
|
|
85.
|
V BonifatiBA OostraP HeutinkLinking DJ-1
to neuro-degeneration offers novel insights for understanding the
pathogenesis of Parkinson’s diseaseJ Mol
Med82163174200414712351
|