Introduction
Proprotein convertase subtilisin/kexin 9 (PCSK9), a
member of the protein-converting enzyme family, is highly expressed
in adult hepatocytes and small intestinal enterocytes (1). PCSK9 is found to be related to
autosomal dominant hypercholesterolemia (2,3).
Recent studies have revealed that PCSK9 plays a unique role in the
control of plasma low-density lipoprotein cholesterol (LDL-C)
levels by regulating LDL receptor (LDLR) in the hepatocyte cell
membrane (4–6). Thus, PCSK9 has emerged as a new
target for the treatment of dyslipidemia (7,8).
Certain studies have shown that PCSK9 may regulate neuronal
apoptosis and affect the nervous system differentiation. These data
support a novel role for PCSK9 in central nervous system (CNS)
development, different from that in cholesterogenic organs
(9). Therefore, PCSK9 may have
biological activities beyond LDLR homeostasis. A recent in
vivo study revealed that infection and inflammation stimulate
PCSK9 expression (10). Lan et
al (11) used microarray
analysis to identify possible genome-wide gene expression and
pathway alterations in HepG2 cells treated with wild-type or D374Y
gain-of-function PCSK9 proteins. They identified novel pathways
that are presumably regulated by PCSK9 and are independent of its
effects on cholesterol uptake. These pathways are involved in
protein ubiquitination, xenobiotic metabolism, cell cycle and
inflammation and stress responses. Both the upregulation and
downregulation of genes in the inflammation and stress response
pathway were also observed in their study.
Atherosclerosis is a vascular disease, characterized
by chronic inflammation of the arterial wall (12). Previous studies have proposed that
the inflammatory response of macrophages induced by oxidized LDL
(oxLDL) is a key event in the pathogenesis of atherosclerosis
(13,14). Previously, we found that PCSK9 is
involved in the apoptosis of human umbilical vein endothelial cells
(HUVECs) induced by oxLDL (15).
We also showed that PCSK9 is expressed in THP-1-derived macrophages
and in New Zealand rabbit atherosclerotic lesion foam cells
(16).
The effect of PCSK9 on the inflammatory response of
macrophages induced by oxLDL remains unknown. The present study
indicates that PCSK9 affects the biological effect induced by oxLDL
in THP-1-derived macrophages. Therefore, we used THP-1-derived
macrophages to examine the effect of oxLDL on PCSK9 expression. The
role of PCSK9 in the oxLDL-induced inflammatory response of these
macrophages was examined using small interfering RNA (siRNA) to
knockdown PCSK9 expression.
Materials and methods
Cell culture and treatments
THP-1 cells were purchased from the Cell Center of
the Institute of Biochemistry and Cell Biology, Shanghai, China.
Cells grown in a monolayer were maintained in RPMI-1640 medium
containing 10% fetal bovine serum and 10 mM HEPES at 37°C in 5%
CO2. The cells were induced to differentiate into
macrophages by adding 160 nM phorbol 12-myristate 13-acetate and
incubated for 24 h. Healthy human plasma was purchased from the
Hengyang Blood Center. Native LDL was separated via
ultracentrifugation, oxidized using 10 μM CuSO4,
and then stored at 4°C. Agarose (0.5%) gel electrophoresis was used
to verify the oxidation of the native LDL. THP-1-derived
macrophages were incubated with various concentrations of oxLDL
(10–80 μg/ml) under different time courses (6–24 h) to
examine the expression levels of IL-1α, IL-6, TNF-α and PCSK9.
siRNA transfection
PCSK9 siRNA was designed based on the PCSK9 mRNA
sequence (GenBank, no. NM_174936) and synthesized by Guangzhou
Biotechnology Co., Guangzhou, China. Sequences of the PCSK9 siRNA
sense and antisense strands are presented in Table I. The negative control siRNA was
provided by the same company. The PCSK9 siRNA was diluted in
Opti-MEM I and added to the culture medium at a final concentration
of 20, 40 or 80 nM. The transfection of siRNA into THP-1-derived
macrophages in suspension was conducted using Lipofectamine 2000
according to the manufacturer’s instructions (Invitrogen, Carlsbad,
CA, USA). To select the most effective siRNA concentration, reverse
transcription-polymerase chain reaction (RT-PCR) and western blot
analysis were performed 24 h after transfection. The concentrations
of PCSK9 siRNA for maximal effects were used to conduct the
following experiments.
 | Table IsiRNAs and PCR primers used in this
study. |
Table I
siRNAs and PCR primers used in this
study.
| Name | Primer
sequences | Products |
|---|
| PCSK9 siRNA | Sense
5′-GGCAGAGACUGAUCCACUUdTdT-3′ | |
| Antisense
3′-dTdTCCGUCUCUGACUAGGUGAA-5′ | |
| PCSK9 | Forward
5′-ACGATGCCTGCCTCTACTCC-3′ | 205 bp |
| Reverse
5′-GCCTGTGATGTCCCACTCTGT-3′ | |
| IL-1α | Forward
5′-AGTAGCAACCAACGGGAA-3′ | 374 bp |
| Reverse
5′-CATCTTGGGCAGTCACATA-3′ | |
| IL-6 | Forward
5′-ACATTTGCCGAAGAGCC-3′ | 234 bp |
| Reverse
5′-GTAGTGAGGAACAAGCCAGAG-3′ | |
| TNF-α | Forward
5′-CTGCCTGCTGCACTTTGGAG-3′ | 348 bp |
| Reverse
5′-TGGTAGGAGACGGCGATGC-3′ | |
| GAPDH | Forward
5′-CAAGGTCATCCATGACAACTTTG-3′ | 496 bp |
| Reverse
5′-GTCCACCACCCTGTTGCTGTAG-3′ | |
Cytokine enzyme-linked immunosorbent
assay (ELISA)
Cells were plated in 6-well plates and treated as
mentioned above. Culture supernatants were collected and stored at
−20°C until analysis. The concentrations of IL-1α, IL-6 and TNF-α
in the supernatants were measured using ELISA (DuoSet ELISA
Development System; R&D Systems, Abingdon, UK) following the
manufacturer’s instructions. The cytokine standards were used to
generate standard curves. Quantitative determinations in 3
different experiments were performed.
RT-PCR
Total RNA was extracted from the cells using TRIzol
reagent in accordance with the manufacturer’s instructions.
Specific PCR primers for human PCSK9, IL-1α, IL-6, and TNF-α are
presented in Table I. RT-PCR
products were analyzed via 1.2% agarose gel electrophoresis and a
UVP gel image analysis system.
Western blot analysis
Total, cytoplasmic and nuclear protein were
extracted from the cells using the Nuclear and Cytoplasmic Protein
Extraction kit (Auragene Bioscience, Changsha, China) according to
the manufacturer’s instructions. The protein concentration was
determined using the bicinchoninic acid method. The samples were
analyzed by western blot analysis using 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, and then transferred to
a polyvinylidene fluoride membrane. Subsequently, the membrane was
blocked in TBST (20 mM Tris base pH 7.6, 150 mM NaCl, 0.1%
Tween-20) containing 5% non-fat milk for 4 h at room temperature.
Antibodies against PCSK9 (Cayman), IκBα and NF-κB (Bioword) were
added to the blocking solution, and the membrane was incubated at
4°C overnight. After the overnight incubation, the membrane was
washed 3 times with TBST for 30 min, and then incubated with the
horseradish peroxidase-conjugated anti-rabbit immunoglobulin G
antibodies (Zhongshan Golden Bridge Inc., Beijing, China) for an
additional 50 min at room temperature. Finally, the membrane was
washed 3 times with TBST for 30 min. Immunoreactivity was detected
using enhanced chemiluminescence. Signal intensities were
quantified via densitometry using Labwords analysis software.
Statistical analysis
Data are expressed as the means ± SEM. The results
were analyzed using one-way analysis of variance (ANOVA), the
Student’s t-test and SPSS 13.0 software. P<0.05 was considered
to indicate a statistically significant difference.
Results
OxLDL upregulates the mRNA expression and
secretion of inflammatory cytokines in a dose- and time-dependent
manner
To determine whether oxLDL affects the synthesis and
secretion of IL-1α, IL-6 and TNF-α, the 3 inflammatory mediators
involved in atherosclerosis, THP-1-derived macrophages were treated
with various doses of oxLDL (0, 10, 20, 40 and 80 μg/ml) for
24 h. Total-RNAs were extracted and analyzed for the mRNA
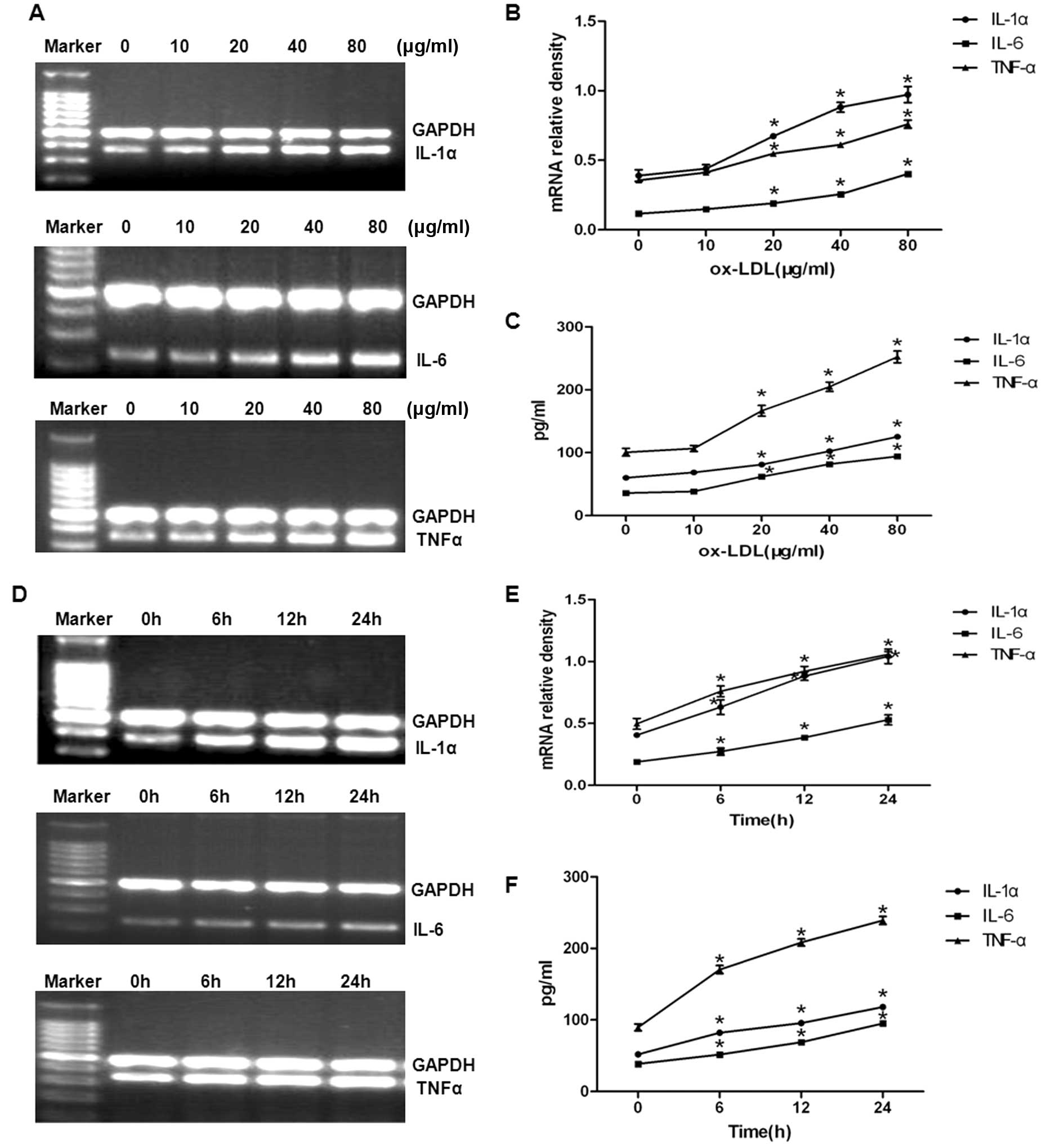
expression levels of these cytokines using RT-PCR. As shown in
Fig. 1A and B, oxLDL increased
the amounts of IL-1α, IL-6 and TNF-α mRNA in a dose-dependent
manner. The amounts of IL-1α, IL-6 and TNF-α secreted from the
cells were determined by ELISA (Fig.
1C). oxLDL increased the amounts of IL-1α, IL-6 and TNF-α in a
dose-dependent manner. These results suggest that oxLDL upregulates
both the mRNA expression and secretion of inflammatory cytokines in
a dose-dependent manner.
Experiments were also performed to determine the
time course of the oxLDL-induced upregulation in IL-1α, IL-6 and
TNF-α mRNA levels and the amounts of IL-1α, IL-6 and TNF-α secreted
from the cells (Fig. 1D and E).
oxLDL increased the levels of IL-1α, IL-6 and TNF-α mRNA in a
time-dependent manner. This upregulation also led to increases in
the amounts of IL-1α, IL-6 and TNF-α secreted into the culture
medium (Fig. 1F). Our results
suggest that OxLDL upregulates the mRNA expression and secretion of
inflammatory cytokines in a dose-and time-dependent manner
(Fig. 1).
OxLDL upregulates PCSK9 expression in
THP-1-derived macrophages in a dose-dependent manner
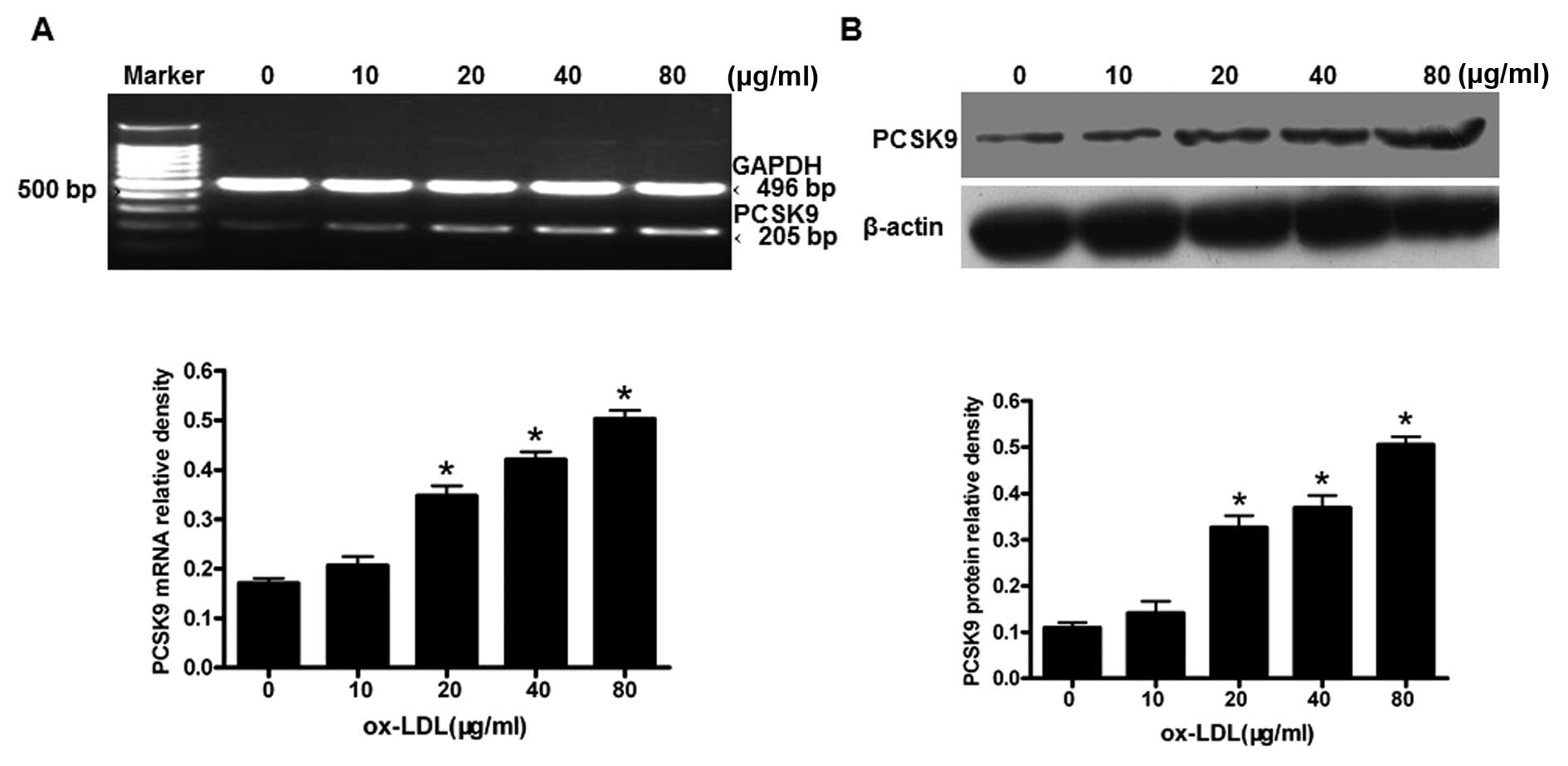
We then investigated whether the expression of PCSK9
in THP-1-derived macrophages is affected by oxLDL stimulation.
THP-1-derived macrophages were treated with oxLDL (0, 10, 20, 40
and 80 μg/ml) for 24 h. The levels of PCSK9 mRNA and protein
were measured using RT-PCR and western blot analysis, respectively.
The expression of PCSK9 mRNA and protein were upregulated in a
concentration-dependent manner (P<0.05) (Fig. 2), with the highest expression of
PCSK9 resulting from treatments of 80 μg/ml oxLDL
(P<0.05). These results suggest that oxLDL upregulates the PCSK9
expression in THP-1-derived macrophages in a dose-dependent
manner.
PCSK9 siRNA reduces the expression of
PCSK9 in THP-1-derived macrophages
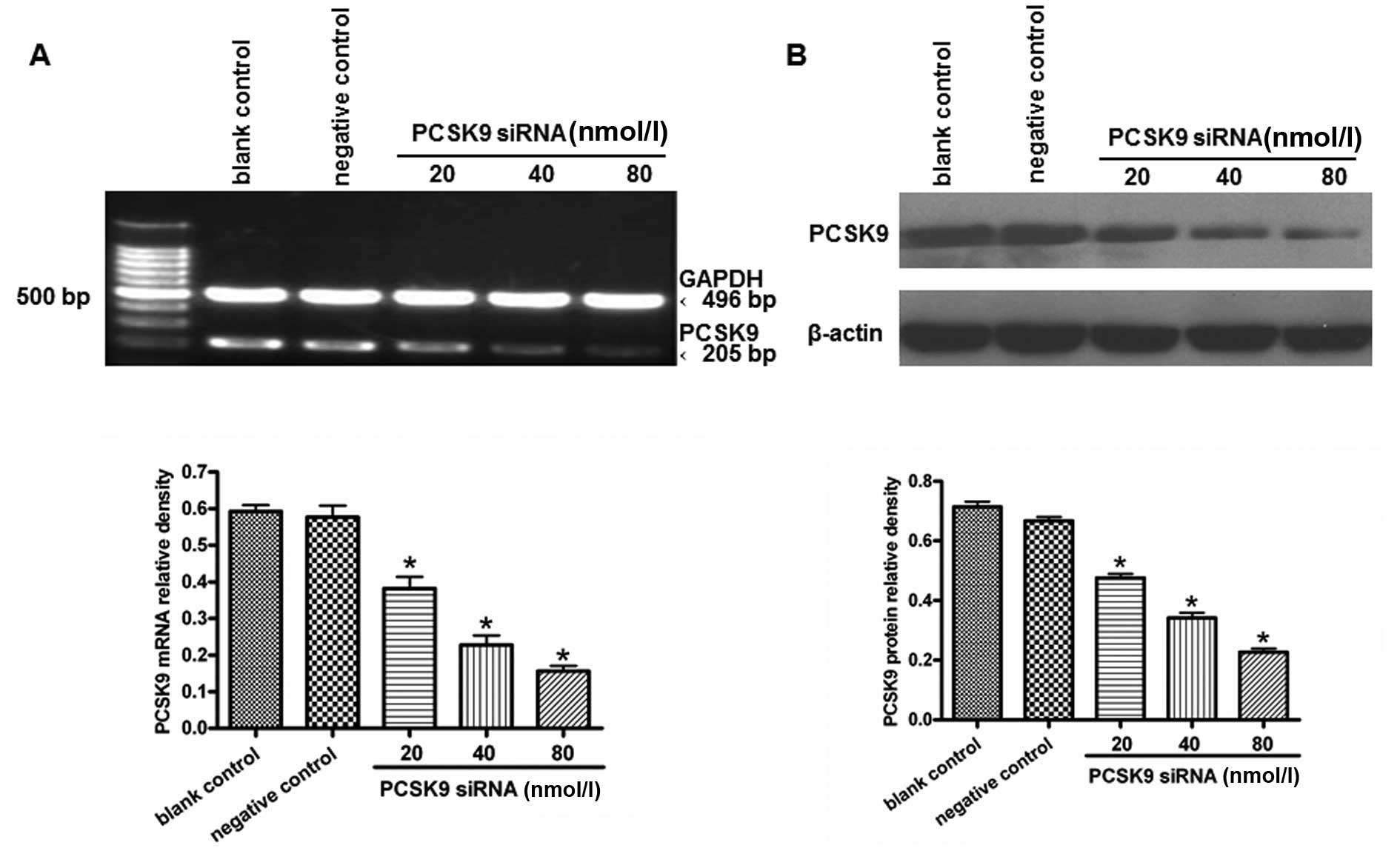
THP-1-derived macrophages were transfected with
various concentrations of siRNA (20, 40 and 80 nM) for 24 h. The
total-RNAs and proteins were extracted. PCSK9 mRNA was detected by
RT-PCR. The treatment with 80 nM PCSK9 siRNA significantly knocked
down the expression of the PCSK9 gene (Fig. 3A). Western blot analysis showed
that PCSK9 protein levels were reduced in PCSK9 siRNA-transfected
cells when compared with cells transfected with the siRNA controls
(P<0.05) (Fig. 3B). Therefore,
80 nM PCSK9 siRNA was used in the subsequent experiments.
PCSK9 siRNA suppresses the oxLDL-induced
upregulation of IL-1α, IL-6 and TNF-α in THP-1-derived
macrophages
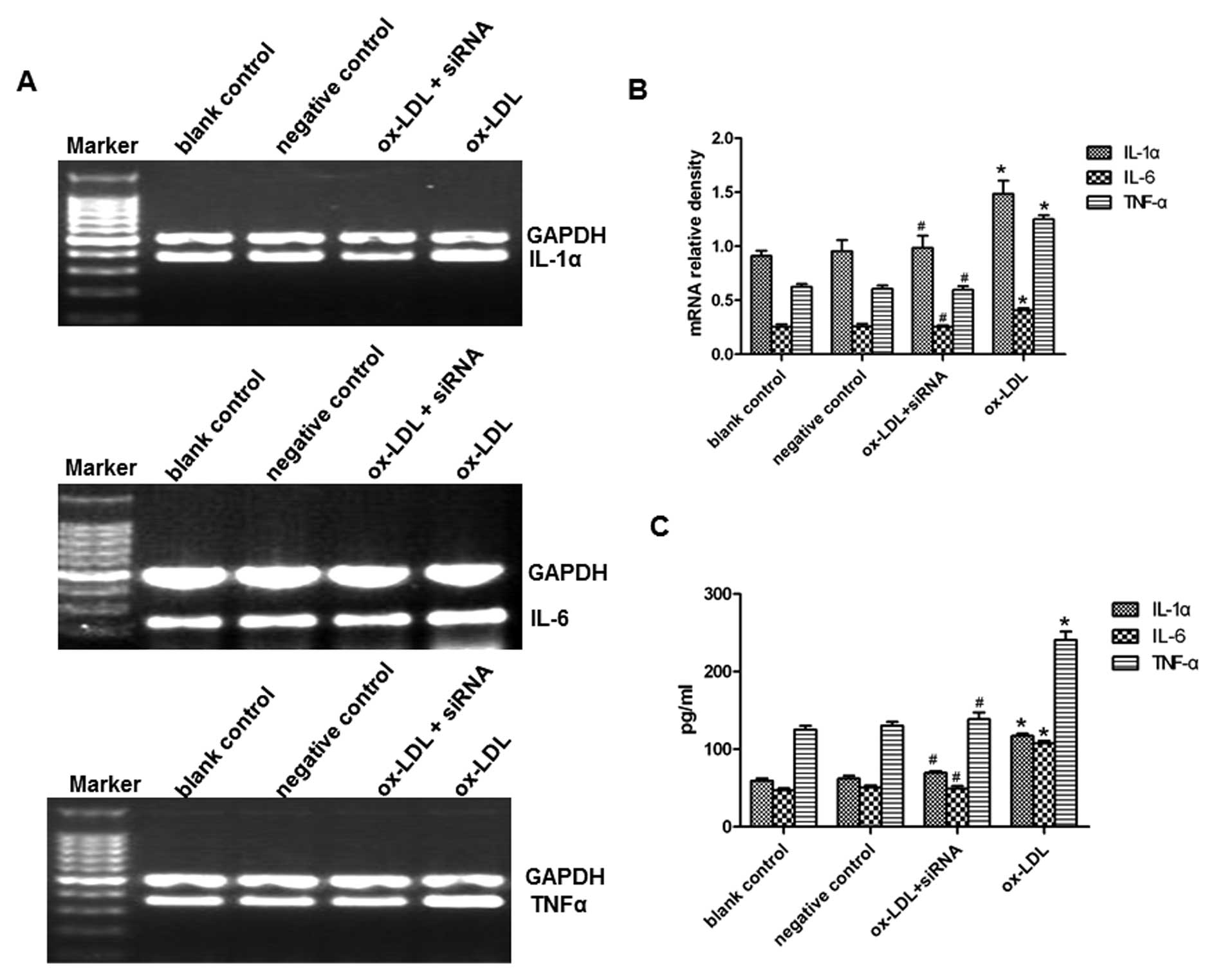
We then investigated whether transfection with PCSK9
siRNA can regulate the inflammatory response in oxLDL-activated
macrophages. THP-1-derived macrophages were transfected with the
control siRNA or PCSK9 siRNA (80 nM), cultured for 24 h in medium,
and then treated with oxLDL (80 μg/ml) for 24 h. First, we
examined the mRNA expression of IL-1α, IL-6 and TNF-α in
oxLDL-induced macrophages. PCSK9 siRNA significantly reduced
oxLDL-induced mRNA expression of IL-1α, IL-6 and TNF-α, as
determined by RT- PCR (Fig. 4A and
B). The concentrations of IL-1α, IL-6 and TNF-α in the medium
were further determined by ELISA. Consistently, the results showed
that PCSK9 siRNA suppressed the oxLDL-induced upregulation of
IL-1α, IL-6 and TNF-α (Fig. 4C).
These results suggest that PCSK9 siRNA suppresses the oxLDL-induced
upregulation of IL-1α, IL-6 and TNF-α in THP-1-derived
macrophages.
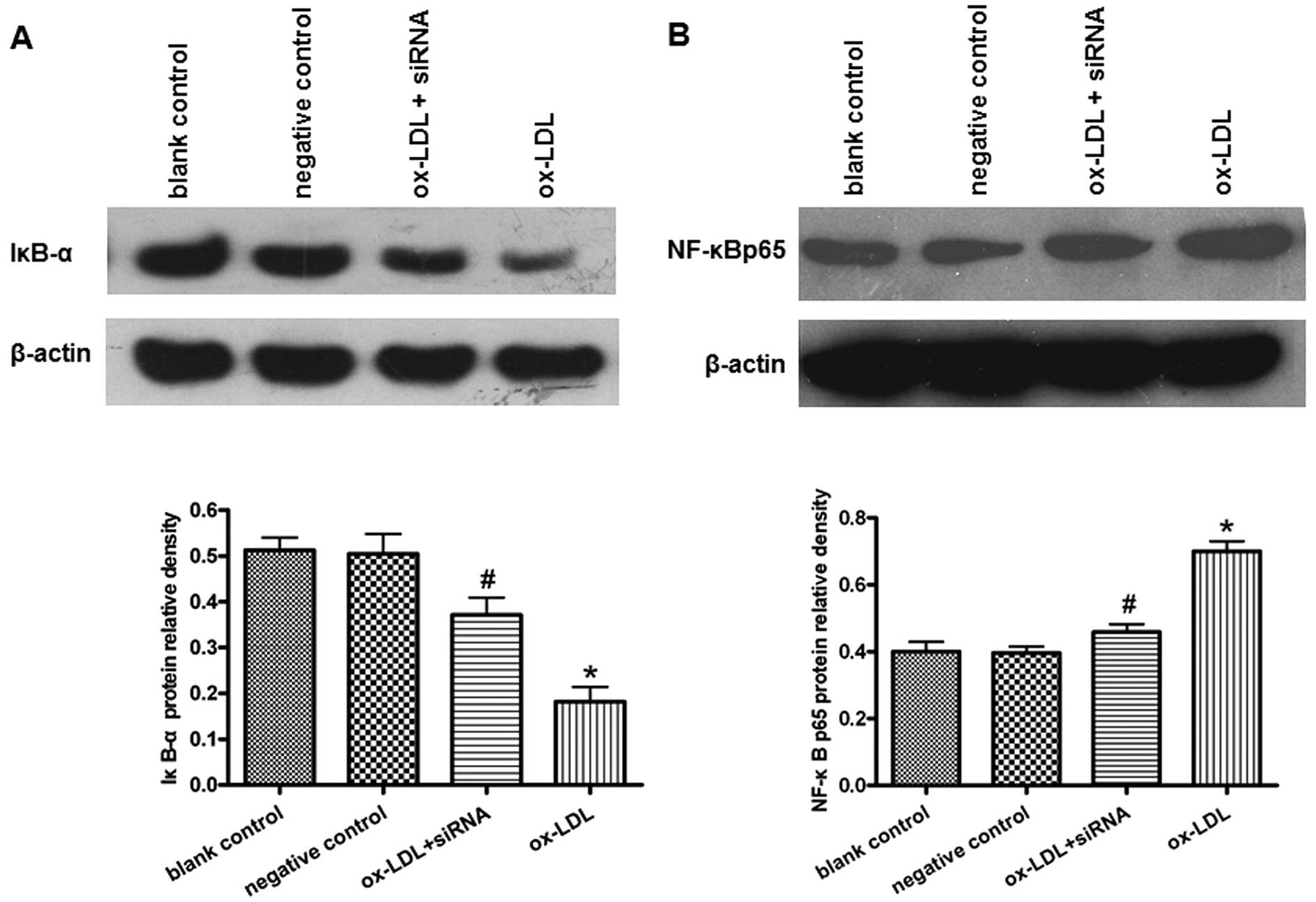
PCSK9 siRNA suppresses oxLDL-induced
IκB-α degradation and NF-κB nuclear translocation in THP-1-derived
macrophages
NF-κB is a major transcription factor regulating the
induction of various inflammatory mediators. To elucidate the
molecular mechanism underlying the anti-inflammatory effects of
PCSK9 siRNA in THP-1-derived macrophages, we further investigated
the effect of PCSK9 siRNA on the NF-κB activity in THP-1-derived
macrophages treated with or without oxLDL. Following cellular
stimulation with oxLDL, IκB-α (a protein inhibiting the
translocation of NF-κB into the nucleus) was degraded significantly
in macrophages (Fig. 5A). The
levels of NF-κB p65 protein in the nucleus were significantly
increased in the cells treated with oxLDL (Fig. 5B). By contrast, PCSK9 siRNA
partially suppressed the degradation of IκB-α and the nuclear
translocation of NF-κB p65 (P<0.05) (Fig. 5).
Discussion
Atherosclerosis is generally considered a
pathological process caused by hyperlipidemia. Recently,
atherosclerosis was found to be a chronic inflammatory response
process beyond lipid metabolism disturbance (17). Many genes related to lipid
metabolism or inflammation are involved in the pathogenesis of
atherosclerosis, such as LDLR, apolipoprotein E (ApoE), C-reactive
protein (CRP) and TNF-α (18). Ma
et al (19) investigated
the effects of inflammatory stress on cholesterol accumulation in
the liver and revealed the correlation between lipid metabolism and
inflammatory response. ATP-binding membrane cassette transporter A1
has been found to be a possible link between inflammation and
reverse cholesterol transport (20,21). Much attention has been focused on
genes that play important roles in both lipid metabolism and
inflammatory response.
PCSK9 is a secreted glycoprotein that regulates the
degradation of LDLR. The function of PCSK9 is currently focused on
lipid metabolism regulation (22–24). Gain-of-function PCSK9 mutations
cause hypercholesterolemia and coronary heart disease (CHD)
(25,26). Loss-of-function PCSK9 mutations
decrease LDL levels and reduce CHD (27,28). As shown in a previous study,
PCSK9-deficient mice exhibited impaired glucose tolerance and
pancreatic islet abnormalities (29). Langhi et al (30) showed that PCSK9 does not alter
insulin secretion, although PCSK9 is expressed in pancreatic δ
cells. Previously, to our knowledge, we reported for the first time
that oxLDL-induced HUVEC THP-1-derived macrophage apoptosis is
effectively inhibited by PCSK9 siRNA via the Bcl/Bax-caspase-9-3
pathway (15). We also found that
PCSK9 is expressed in THP-1-derived macrophages and New Zealand
rabbit atherosclerotic lesion macrophages (16). PCSK9 is markedly induced by
diverse inflammatory stimuli, such as lipopolysaccharide, zymosan
and turpentine (10). PCSK9 may
be a key regulator in lipid metabolism, inflammation, and
atherosclerosis.
Considering that oxLDL is a major pro-inflammatory
factor in the development of atherosclerosis, we selected
oxLDL-stimulated THP-1-derived macrophages as the cell model. In
the present study, IL-1α, IL-6 and TNF-α expression and secretion
increased with oxLDL in a dose- and time-dependent manner.
Importantly, oxLDL also increased the PCSK9 expression in
THP-1-derived macrophages in a dose-dependent manner. The change in
the PCSK9 expression is consistent with the change in inflammatory
cytokine secretion in the oxLDL-stimulated THP-1-derived
macrophages.
Subsequently, we investigated the role of PCSK9 in
oxLDL-induced inflammatory response in THP-1-derived macrophages
using RNA interference. In the present study, we used 3 siRNA
concentrations (20, 40 and 80 nM) and found that the silencing
effect was more potent with increasing concentrations of siRNA, as
detected at both the mRNA and protein levels. The final siRNA
concentration of 80 nM showed the most dramatic silencing effects.
Therefore, we used 80 nM PCSK9 siRNA in all subsequent experiments.
The cells were transfected with PCSK9 siRNA, cultured for 24 h, and
then exposed to oxLDL for 24 h. The expression and secretion of
inflammatory cytokines were determined. Compared to the cells
incubated with oxLDL only, PCSK9 siRNA significantly suppressed the
expression and secretion of IL-1α, IL-6 and TNF-α. These results
further suggest that PCSK9 is involved in the regulation of the
oxLDL-induced inflammatory response in THP-1-derived
macrophages.
Therefore, we attempted to identify the detailed
mechanism of PCSK9 siRNA in regulating the inflammatory response.
NF-κB is a critical transcriptional regulator of a number of genes
involved in immune and inflammatory responses (31). In the resting state, NF-κB is
sequestered in the cytoplasm by binding to an inhibitory factor,
IκBα. With cell stimulation, IκBα is phosphorylated and degraded.
Unbound NF-κB is then released and translocated into the nucleus
where it activates the transcription of target genes, such as
cytokines, chemokines, adhesion molecules and apoptosis inhibitors
(32,33). NF-κB activation also plays a key
role in the oxLDL-induced expression of inflammatory cytokines in
macrophages (34–35). Thus, we speculate that the
suppression of pro-inflammation cytokine secretion by PCSK9 siRNA
is related with NF-κB activation. In the present study, the results
suggest that PCSK9 siRNA reduces the degradation of IκBα and
suppresses the nuclear translocation of NF-κB p65 induced by oxLDL
in macrophages, indicating that the NF-κB activation is inhibited
by PCSK9 siRNA. Collectively, PCSK9 siRNA suppresses not only the
IL-1α, IL-6 and TNF-α expression and secretion but also NF-κB
activation. Further, the activation of NF-κB is responsible for
inflammatory cytokine expression and secretion. Therefore, in
oxLDL-stimulated macrophages, the inhibitory effect of PCSK9 siRNA
on inflammatory cytokines expression and secretion is partly
associated with the down-regulation of the NF-κB activation.
In conclusion, to our knowledge, the results of the
present study strongly indicate for the first time that PCSK9 may
be induced in THP-1-derived macrophages in response to oxLDL. We
also demonstrate that PCSK9 siRNA suppresses the oxLDL-induced
pro-inflammatory cytokine synthesis and secretion by inhibiting
NF-κB translocation into the nucleus, which exhibits an
anti-inflammatory role in macrophages. We believe that further
clarifying the functional role of PCSK9 will hasten its application
as a unique aid for atherosclerosis therapeutic targets.
Acknowledgements
This study was supported by grants
from the Key Project of Educational Department of Hunan Province
(10A105), the Project of Science and Technology Department of Hunan
Province (2009TP4057-2, 2010TP4008-2), and the Project of Health
Department of Hunan Province (B2011-042), and by the Aid Program
for Science and Technology Innovative Research Team in Higher
Educational Institutions of Hunan Province.
References
|
1.
|
B CariouC Le MayP CostetClinical aspects
of
PCSK9Atherosclerosis216258265201110.1016/j.atherosclerosis.2011.04.01821596380
|
|
2.
|
KN MaxwellRE SoccioEM DuncanE SehayekJL
BreslowNovel putative SREBP and LXR target genes identified by
microarray analysis in liver of cholesterol-fed miceJ Lipid
Res4421092119200310.1194/jlr.M300203-JLR20012897189
|
|
3.
|
NG SeidahS BenjannetL WickhamThe secretory
proprotein convertase neural apoptosis-regulated convertase 1
(NARC-1): liver regeneration and neuronal differentiationProc Natl
Acad Sci USA100928933200310.1073/pnas.0335507100
|
|
4.
|
T YamamotoC LuRO RyanA two-step binding
model of PCSK9 interaction with the low density lipoprotein
receptorJ Biol
Chem28654645470201110.1074/jbc.M110.19904221149300
|
|
5.
|
J DavignonG DubucNG SeidahThe influence of
PCSK9 polymorphisms on serum low-density lipoprotein cholesterol
and risk of atherosclerosisCurr Atheroscler
Rep12308315201010.1007/s11883-010-0123-620623344
|
|
6.
|
SA MousaviKE BergeTP LerenThe unique role
of proprotein convertase subtilisin/kexin 9 in cholesterol
homeostasisJ Intern
Med266507519200910.1111/j.1365-2796.2009.02167.x19930098
|
|
7.
|
AJ MarianPCSK9 as a therapeutic target in
atherosclerosisCurr Atheroscler
Rep12151154201010.1007/s11883-010-0099-220425252
|
|
8.
|
CJ DuffNM HooperPCSK9: an emerging target
for treatment of hypercholesterolemiaExpert Opin Ther
Targets15157168201110.1517/14728222.2011.54748021204732
|
|
9.
|
S PoirierA PratE MarcinkiewiczImplication
of the proprotein convertase NARC-1/PCSK9 in the development of the
nervous systemJ
Neurochem98838850200610.1111/j.1471-4159.2006.03928.x16893422
|
|
10.
|
KR FeingoldAH MoserJK ShigenagaSM PatzekC
GrunfeldInflammation stimulates the expression of PCSK9Biochem
Biophys Res
Commun374341344200810.1016/j.bbrc.2008.07.02318638454
|
|
11.
|
H LanL PangMM SmithProprotein convertase
subtilisin/kexin type 9 (PCSK9) affects gene expression pathways
beyond cholesterol metabolism in liver cellsJ Cell
Physiol224273281201020333646
|
|
12.
|
R RossAtherosclerosis is an inflammatory
diseaseAm Heart
J138S419S420199910.1016/S0002-8703(99)70266-810539839
|
|
13.
|
G RodriguezN MagoF RosaRole of
inflammation in atherogenesisInvest Clin501091292009
|
|
14.
|
P SahaB ModaraiJ HumphriesK MattockM
WalthamKG BurnandA SmithThe monocyte/macrophage as a therapeutic
target in atherosclerosisCurr Opin
Pharmacol9109118200910.1016/j.coph.2008.12.01719230773
|
|
15.
|
CY WuZH TangL JiangXF LiZS JiangLS
LiuPCSK9 siRNA inhibits HUVEC apoptosis induced by ox-LDL via
Bcl/Bax-caspase9-caspase3 pathwayMol Cell
Biochem359347358201210.1007/s11010-011-1028-621847580
|
|
16.
|
ZH TangLS LiuJ PengQ YangHT LiuZS
JiangInvestigation of NARC-1 protein expression in atherosclerotic
lesion of New Zealand rabbitChin J Arterioscler1735372009(in
Chinese)
|
|
17.
|
P LibbyY OkamotoVZ RochaE
FolcoInflammation in atherosclerosis: transition from theory to
practiceCirc J74213220201020065609
|
|
18.
|
K YinC TangInflammation, lipid metabolism
dysfunction, and hypertension: Active research fields in
atherosclerosis-related cardiovascular disease in ChinaSci China
Life Sci54976979201110.1007/s11427-011-4225-3
|
|
19.
|
KL MaXZ RuanSH PowisY ChenJF MoorheadZ
VargheseInflammatory stress exacerbates lipid accumulation in
hepatic cells and fatty livers of apolipoprotein E knockout
miceHepatology48770781200810.1002/hep.2242318752326
|
|
20.
|
K YinDF LiaoCK TangATP-binding membrane
cassette transporter A1 (ABCA1): a possible link between
inflammation and reverse cholesterol transportMol
Med16438449201020485864
|
|
21.
|
K YinX DengZC MoTristetraprolin-dependent
post-transcriptional regulation of inflammatory cytokine mRNA
expression by apolipoprotein A–I: role of ATP-binding membrane
cassette transporter A1 and signal transducer and activator of
transcription 3J Biol Chem2861383413845201121339300
|
|
22.
|
A RoubtsovaMN MunkondaZ AwanCirculating
proprotein convertase subtilisin/kexin 9 (PCSK9) regulates VLDLR
protein and triglyceride accumulation in visceral adipose
tissueArterioscler Thromb Vasc
Biol31785791201110.1161/ATVBAHA.110.220988
|
|
23.
|
OL HollaJ CameronK TvetenTB StromKE
BergeJK LaerdahlTP LerenRole of the C-terminal domain of PCSK9 in
degradation of the LDL receptorsJ Lipid
Res5217871794201110.1194/jlr.M01809321771976
|
|
24.
|
AK SoutarUnexpected roles for PCSK9 in
lipid metabolismCurr Opin
Lipidol22192196201110.1097/MOL.0b013e32834622b521494143
|
|
25.
|
TS FisherP Lo SurdoS PanditEffects of pH
and low density lipoprotein (LDL) on PCSK9-dependent LDL receptor
regulationJ Biol
Chem2822050220512200710.1074/jbc.M70163420017493938
|
|
26.
|
D CunninghamDE DanleyKF
GeogheganStructural and biophysical studies of PCSK9 and its
mutants linked to familial hypercholesterolemiaNat Struct Mol
Biol14413419200710.1038/nsmb123517435765
|
|
27.
|
DW ZhangTA LagaceR GarutiZ ZhaoM
McDonaldJD HortonJC CohenHH HobbsBinding of proprotein convertase
subtilisin/kexin type 9 to epidermal growth factor-like repeat A of
low density lipoprotein receptor decreases receptor recycling and
increases degradationJ Biol
Chem2821860218612200710.1074/jbc.M702027200
|
|
28.
|
J CameronOL HollaT RanheimMA KulsethKE
BergeTP LerenEffect of mutations in the PCSK9 gene on the cell
surface LDL receptorsHum Mol
Genet1515511558200610.1093/hmg/ddl07716571601
|
|
29.
|
M MbikayF SiroisJ MaynePCSK9-deficient
mice exhibit impaired glucose tolerance and pancreatic islet
abnormalitiesFEBS
Lett584701706201010.1016/j.febslet.2009.12.01820026049
|
|
30.
|
C LanghiC Le MayV GmyrPCSK9 is expressed
in pancreatic delta-cells and does not alter insulin
secretionBiochem Biophys Res
Commun39012881293200910.1016/j.bbrc.2009.10.13819878649
|
|
31.
|
K De BosscherBW VandenG HaegemanThe
interplay between the glucocorticoid receptor and nuclear
factor-kappaB or activator protein-1: molecular mechanisms for gene
repressionEndocr Rev244885222003
|
|
32.
|
G GloireS Legrand-PoelsJ PietteNF-kappaB
activation by reactive oxygen species: fifteen years laterBiochem
Pharmacol7214931505200616723122
|
|
33.
|
MP de WintherE KantersG KraalMH
HofkerNuclear factor kappaB signaling in atherogenesisArterioscler
Thromb Vasc Biol25904914200515731497
|
|
34.
|
C XieJ KangME FergusonS NagarajanTM
BadgerX WuBlueberries reduce pro-inflammatory cytokine TNF-α and
IL-6 production in mouse macrophages by inhibiting NF-κB activation
and the MAPK pathwayMol Nutr Food Res55158715912011
|
|
35.
|
L LeiH LiF YanY LiY XiaoPorphyromonas
gingivalis lipopolysaccharide alters atherosclerotic-related
gene expression in oxidized low-density-lipoprotein-induced
macrophages and foam cellsJ Periodontal
Res46427437201110.1111/j.1600-0765.2011.01356.x
|