Introduction
Liriope platyphylla (LP) is a perennial
seed-reproducing plant that is distributed widely throughout the
temperate climate region of the northern hemisphere. In Korea,
these plants grow principally in low mountain areas at altitudes
<500 m above sea level, and their leaves remain green throughout
the year (1). LP is a herb that
has been used for a long time in oriental medicine to treat asthma
as well as bronchial and lung inflammation (2). The effects of LP root extracts on
the prevention of obesity, diabetes, inflammation and
neurodegenerative disease have previously been demonstrated
(3–7). Among these therapeutic effects, LP
exhibited therapeutic potential in human subjects suffering from
obesity and diabetes. In particular, Gyeongshingangjeehwan (GGEx;
mainly containing LP) has been reported to prevent obesity and
hypertriglyceridemia by inhibiting feeding and activating hepatic
peroxisome proliferator-activated receptor-α in OLETF male rats
(6). In addition, the
homoisoflavone-enriched fraction of LP methanol extracts increased
insulin-stimulated glucose uptake in 3T3-L1 adipocytes through
increased glucose transporter (Glut)-4 contents in the plasma
membrane (3). Some novel
compounds have been isolated from LP using methanol extraction. Of
these compounds, LP9M80-H induces insulin secretion and
differentially regulates the expression of Glut-1 and Glut-3
expression through the mitogen-activated protein kinase (MAPK) and
phosphoinisitide-3-kinase (PI3-K) signaling pathways (8). Furthermore, an aqueous extract of LP
induces the upregulation of insulin secretion and the
downregulation of glucose concentration through an increase in the
expression of several Glut proteins regulated by the insulin
signaling pathway (9).
Steaming is often applied to medicinal plants to
increase the levels or efficacy of their functional components as
well as to induce chemical transformations of the specific
components (10). This process
has been applied most successfully to ginseng, derivations of which
are taken orally as adaptogens, aphrodisiacs, nourishing
stimulants, as well as in the treatment of type II diabetes and
sexual dysfunction in men (11–13). Korean ginseng is found in two
forms; Korean white ginseng (KWG) (Panax ginseng C.A.
Meyer), which is air-dried ginseng, and Korean red ginseng (KRG)
(Ginseng Radix Rubra), which is repeatedly steamed (10). During the steaming process,
several important components in ginseng, such as ginsenosides,
acidic polysaccharides and phenolics, are transformed into
different components and several new compounds, such as
non-saponinpolyaceylene, maltol and amino acid (14,15). Recently, RLP was produced by a
steaming process and its effects on the insulin secretion ability
and insulin receptor signaling pathway were examined. The maximum
insulin secretion was observed in INS cells treated with 3-SLP and
9-SALP (16). Despite these
initial results, there are no reports on how RLP affects the
insulin secretion ability and adipocyte hypertrophy to improve
diabetes and obesity.
Therefore, in this study, we investigated the
effects of RLP on insulin secretion and adipocyte hypertrophy using
a type II diabetes model. Significant changes on the insulin
release, lipid concentration, fatty liver formation and fatty acid
oxidation-related genes were observed in the pancreas, abdominal
fat and livers of OLETF rats in response to RLP treatment.
Materials and methods
Preparation of RLP
LP roots were collected from plantations in the
Miryang area of Korea (Fig. 1)
and dried with a hot-air drier (JSR Instruments, Uttaranchal,
India) at 60°C. The voucher specimens (WPC-11-010) were deposited
at the Functional Materials Bank of PNU-Wellbeing RIS Center at
Pusan National University. Six hundred grams of dry roots were
reduced to powder using an electric blender. The water extract was
purified at 100°C for 2 h using circulated extraction equipment
(IKA Labortechnik, Staufen, Germany) after adding 2 liters of
distilled water. The composition of RLP was measured as previously
described (17,18). Extract solutions were concentrated
into dry pellets using a rotary evaporator (Eyela, Tokyo, Japan)
and stored at −80°C until use.
HPLC analysis of RLP
The AEtLP was analyzed by ILC 3000 HPLC system
(Interface Engineering, Co., Ltd., Seoul, Korea) equipped with
Corona® CAD® Detector (ESA Biosciences, Inc.,
Chelmsford, MA, USA). The chromatographic separation was performed
on a CapCell PAK MG C18 (4.6×250 mm, particle size 5 μm;
Shiseido Co., Ltd., Tokyo, Japan). The mobile phase consisted of
solvent A (deionized water) and solvent B (acetonitrile) using the
gradient elution program; 0–25 min, 30–90% of solvent B and 25–40
min, 90% of solvent B. A flow rate of 1.0 ml/min was used for the
sample analysis. The nebulizer gas was compressed nitrogen. The gas
flow rate and gas pressure were maintained at 1.53 l/min and 35±2
psi, respectively. The output signal of the detector was recorded
using Clarity™ Chromatography Software (DataApex, Prague, Czech
Republic).
Care and use of OLETF rats
All animal experimental procedures in this study
were approved by the Institutional Animal Care and Use Committee
(IACUC) at Pusan National University (PNU-2011-000220). The animals
were handled in a Pusan National University-Laboratory Animal
Resources Center accredited by the Korea FDA in accordance with the
USA NIH guidelines (accredited unit no. 000996). All rats were
housed in specified pathogen-free (SPF) conditions under a strict
light cycle (light on at 06:00 h and off at 18:00 h), and given a
standard irradiated chow diet (Purina Mills, Inc.) ad
libitum. The 20-week-old LETO and OLETF rats used in this study
were supplied by Central Laboratory Animals, Inc. (Seoul,
Korea).
Treatment of RLP and detection of glucose
levels
RLP was dissolved in distilled water to give a final
concentration of 9.1 μg/ml. OLETF rats were divided randomly
into 2 groups. The first group received a comparable volume of
daily water via oral administration (vehicle-treated group),
whereas the second group received 50 mg/kg body weight/day of RLP
via oral administration for 3 weeks (RLP-treated group). After 3
weeks, the glucose concentration was detected following 24 h of
fasting using the sensitive strip of the Blood Glucose Monitoring
System (i-Sens, Inc., Seoul, Korea).
Intraperitoneal glucose tolerance
test
The glucose tolerance was determined from 5
rats/group using an intraperitoneal glucose tolerance test (IPGTT;
1.5 mg glucose/g after a 24 h fast with blood glucose measurements
being made at 0, 15, 30, 60 and 120 min). The serum glucose level
from rats was detected using the sensitive strip of the Blood
Glucose Monitoring System (i-Sens, Inc.,).
Serum biochemical analysis
After the final administration of RLP, the rats were
fasted for 24 h and blood was collected from the abdominal vein.
Serum was obtained by the centrifugation of blood incubated for 30
min at room temperature. The biochemical components in the serum
were assayed using an Automated Serum Analyzer (Hitachi 747,
Japan). All assays were measured with fresh serum and carried out
in duplicate.
Quantification of insulin and adiponectin
by ELISA
The concentration of insulin and adiponectin in
serum from the LETO and OLETF rats was detected using an
ultra-sensitive assay procedure and reagents in the Mercodia Rat
Insulin ELISA kit (Cat. 10-1250-01; Mercodia, Sweden) and
Adiponectin ELISA kit (Cat. AG-45A-0005TP-KI01; AdipoGen, Inc.,
Korea). The sera and standards were incubated in a plate shaker at
100–150 rpm at room temperature for 2 h on antibody-coated plates.
The wells were then washed 6 times using an automatic plate washer
(PV100; Hoefer, Inc., USA). HRP conjugate was added to all the
plates, which were then incubated in a shaker for 15 min at room
temperature. The reaction was quenched by adding 50 μl of a
stop solution (0.5 M H2SO4). The plates were
read at 450 nm using a Molecular Devices Vmax plate reader
(Sunnyvale, CA, USA).
Histological analysis and
immunostaining
The abdominal fat and liver collected from the rats
were fixed with 10% formalin for 48 h, embedded in paraffin wax and
sectioned into 3 μm slices. The fat and liver sections were
then stained with hematoxylin & eosin (H&E; Sigma-Aldrich,
St. Louis, MO, USA), and observed by optical microscopy. The size,
number and morphology of the adipocytes were measured using Leica
Application Suite (Leica Microsystems, Switzerland).
Immunohistochemical analysis was performed as
previously described (8).
Briefly, the distribution of insulin protein was observed using
optical microscopy after fixing the tissue samples in 5% formalin
for 48 h, embedding the tissues in paraffin and acquiring 3
μm sections. Each section was de-paraffinized with xylene,
rehydrated and pretreated for 30 min at room temperature with a
phosphate-buffered saline (PBS)-based blocking buffer containing
10% goat serum. The samples were then incubated with mouse
anti-insulin antibody diluted 1:1,000 in PBS-blocking buffer.
Antigen-antibody complexes were visualized with goat anti-rabbit
HRP-conjugated streptavidin secondary antibody (Histostain-Plus
kit; Zymed Laboratories) diluted 1:1,000 in PBS-blocking buffer.
3,3′-Diaminobenzidine (DAB) substrate (Invitrogen Life
Technologies, Carlsbad, CA, USA) and a model GS-690 imaging
densitometer (Bio-Rad Laboratories, Hercules, CA, USA) were used to
detect insulin proteins.
RT-PCR
The frozen fat and liver tissues were chopped with
scissors and homogenized in an RNAzol B solution (CS104; Tet-Test,
Inc.). The isolated RNA was then determined by UV-spectroscopy.
Gene expression was examined by RT-PCR using 5 μg of the
total RNA from each tissue, and 500 ng of the oligo(dT) primer
[(18418-012); Gibco-BRL] was annealed at 70°C for 10 min.
Complementary DNA, which served as the template for further
amplification, was synthesized by adding dATP, dCTP, dGTP and dTTP
with 200 units of reverse transcriptase. Ten pmole of the sense and
antisense primers were added, and the reaction mixture was then
subjected to 25 cycles of amplification. Amplification was carried
out in a Perkin-Elmer Thermal Cycler using the following cycles; 30
sec at 94°C, 30 sec at 62°C and 45 sec at 72°C. In each case, the
minus-RT controls were included to differentiate the DNA and RNA
products. RT-PCR was also performed using the primers specific to
β-actin to ensure the RNA integrity. The primer sequences for
VLCAD, MCAD, PPAR-γ, aP2 and leptin were based on a previous study
(19). The sequences of the
β-actin sense and antisense primers were, 5′-TGGAA TCCTG TGGCA
TCCAT GAAAC-3′ and 5′-TAAAA CGCAG CTCAG TAACA GTCCG-3′,
respectively. The levels of the PCR products were quantified using
a Kodak Electrophoresis Documentation and Analysis System 120 on 1%
agarose gels. The experiment was repeated 3 times.
Statistical analysis
The tests for significance between the vehicle- and
RLP-treated groups in the OLETF rats were performed using a one-way
ANOVA test of variance. In addition, the tests for significance
between the LETO and OLETF groups were performed using a post hoc
test (all were from SPSS for Windows, Release 10.10, Standard
Version; Chicago, IL, USA) of the variance and the significance
levels are given in the text. All values are reported as the mean
standard deviation (SD). P<0.05 was considered to indicate
statistically significant difference.
Results
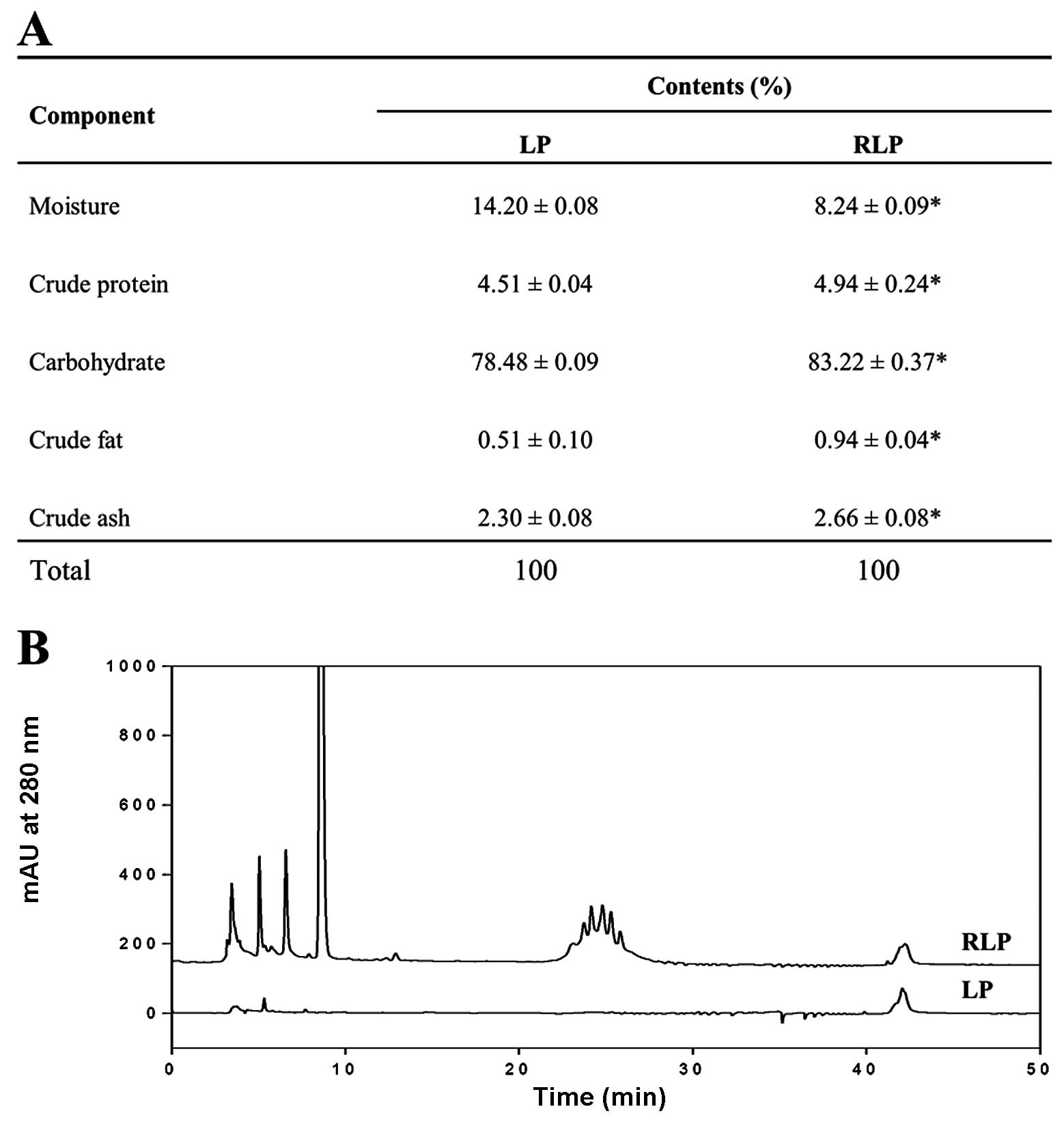
Composition of RLP
RLP consisted mainly of carbohydrates (83.22%) and
moisture (8.24%) and, to a lesser extent, proteins, ash and fat
(Fig. 1A). However, following the
steaming process, their level was significantly changed into a
different concentration. Four components including carbohydrate,
protein, ash and fat were slightly increased in RLP, while the
moisture was greatly decreased. In particular, the HPLC
chromatogram showed a large amount of polyphenolic compounds which
have antioxidant activity contained in RLP when compared with the
LP (Fig. 1B).
RLP effects on the weight of body and
glucose regulation of OLETF rats
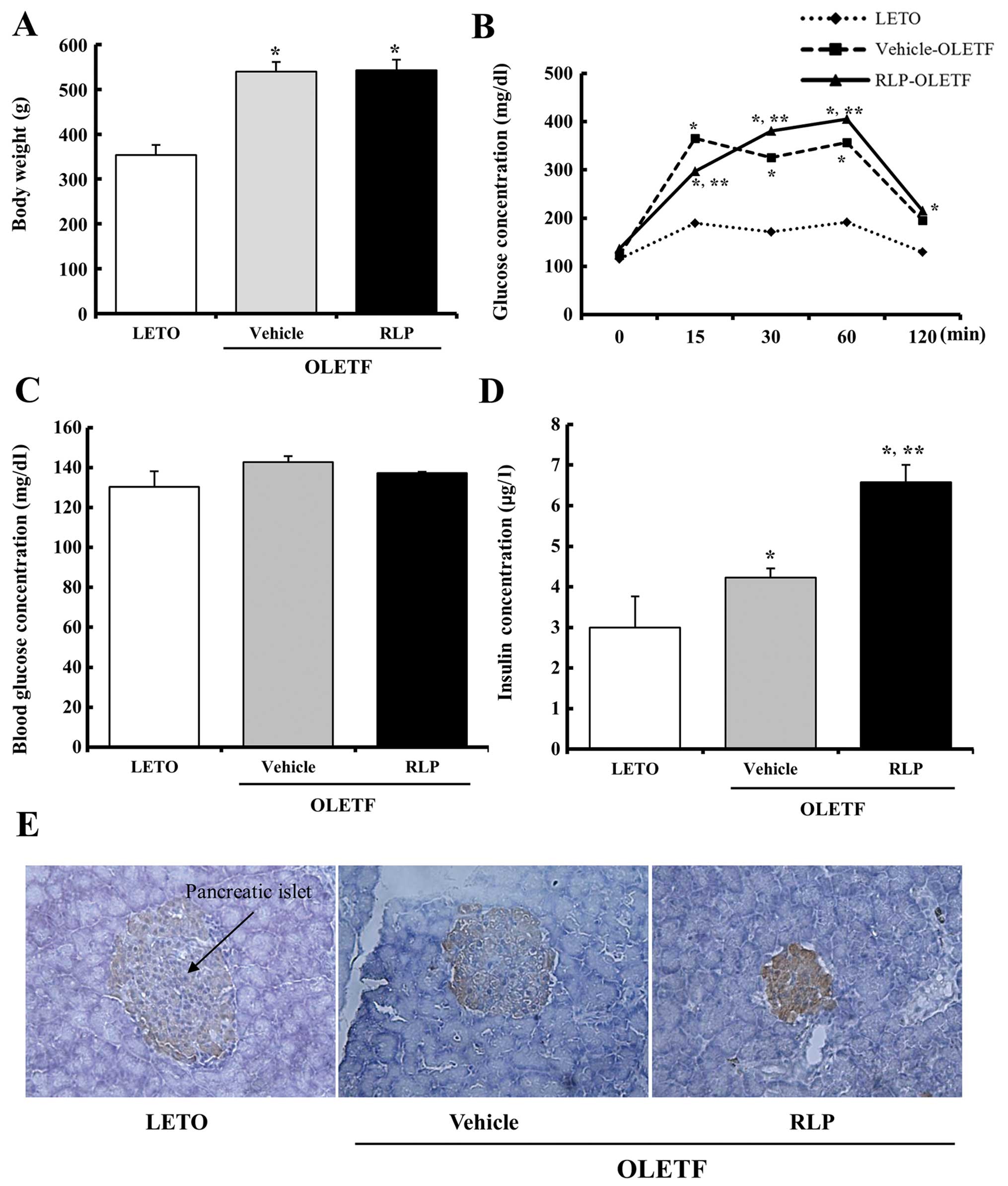
The body weights of the LETO and OLETF rats
following RLP treatment for 3 weeks were firstly measured to
determine if the RLP treatment affects the progress of obesity. The
vehicle-treated OLETF rats had a higher body weight than the LETO
rats. On the other hand, the body weight was not altered in the
RLP-treated group compared to the vehicle-treated group (Fig. 2A). Therefore, the 50 mg/kg RLP
treatment did not work efficiently on the decrease of the body
weight to affect the obesity status of OLETF rats.
In addition, the effect of the RLP treatment on the
glucose tolerance response was determined by measuring the serum
glucose level during the time course of an IPGTT in the 20-week-old
OLETF rats using a post-prandial IPGTT. The OLETF rats showed
significant impairment in glucose tolerance compared to the LETO
rats during the course of an IPGTT. However, no significant
difference was detected between the RLP- and the vehicle-treated
group. Only the glucose concentration was lower in the RLP-treated
group compared to the vehicle-treated group at the early stage
(Fig. 2B). Nevertheless, the
blood concentration on the final day of the experiment did not
change in the RLP-treated OLETF rats compared to the
vehicle-treated rats (Fig. 2C).
Regarding insulin, its concentration was significantly higher in
the OLETF rats compared to the LETO rats (∼1.43-fold). In the
RLP-treated group, the insulin concentration was increased
significantly to 6.4 μg/l (Fig. 2D). Furthermore, a similar pattern
was detected in the pancreatic islets using immunostaining analysis
(Fig. 2E). These results suggest
that RLP treatment can effectively induce an enhancement of insulin
secretion ability in the serum of OLETF rats.
RLP effects on the adiponectin and lipid
concentration in the serum of OLETF rats
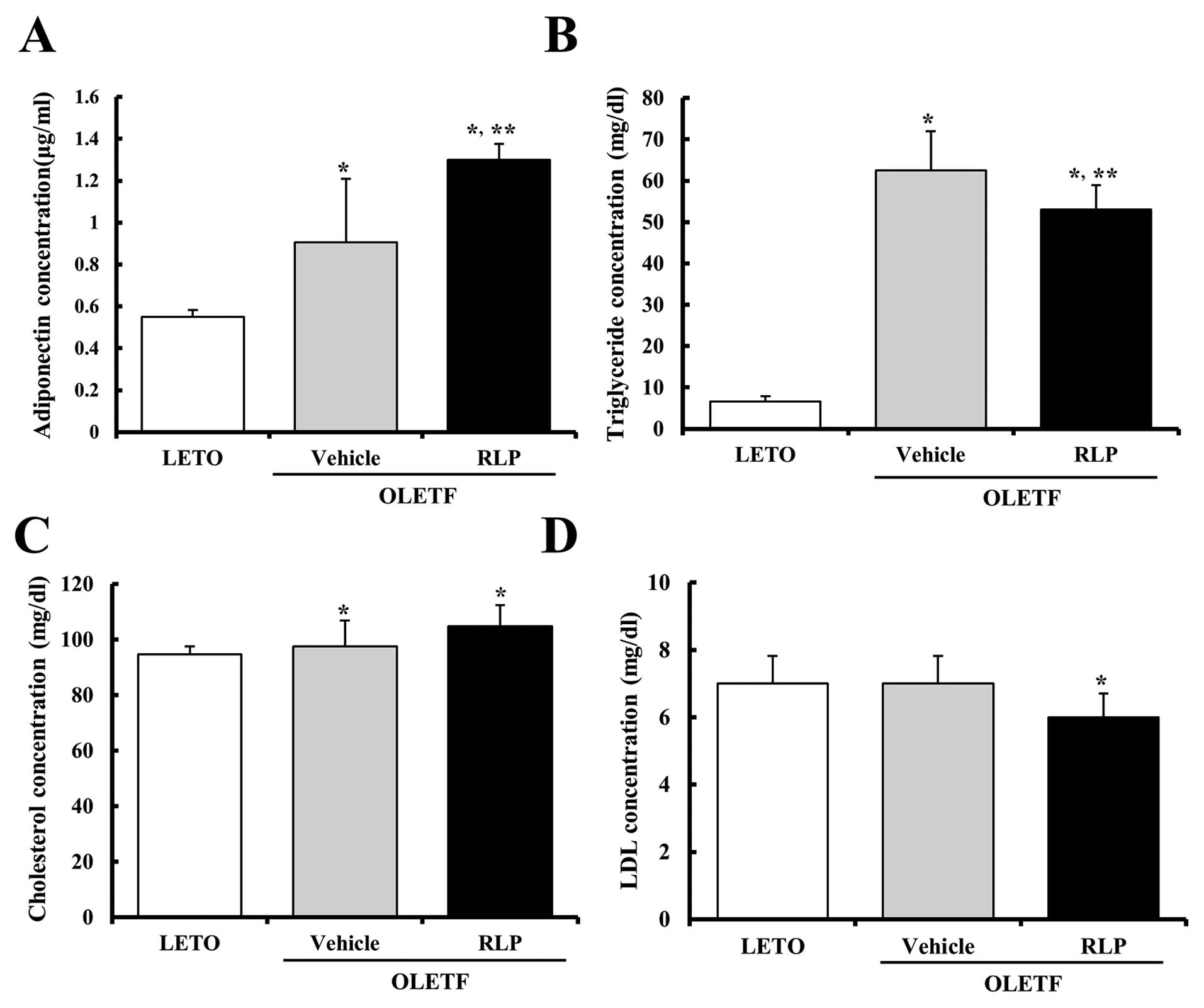
In most animals, adiponectin plays a critical role
in controlling the systemic fat catabolism and glucose metabolism
(20–22). The adiponectin concentration in
the serum of OLETF rats was measured to determine the effect of RLP
treatment on the regulation of adiponectin concentration. As shown
Fig. 3A, the adiponectin
concentration was increased significantly in the vehicle-treated
OLETF rats compared to the LETO rats. In addition, these levels
were significantly higher in the RLP-treated rats than in the
vehicle-treated OLETF rats. Therefore, these results show that the
RLP treatment could induce a strong increase in adiponectin
concentration in OLETF rats.
An increase in lipid concentration including
triglyceride, cholesterol and LDL is one of the important features
observed in obesity (23). The
levels of triglyceride, cholesterol and LDL in the serum of OLETF
rats were measured to determine if the RLP treatment could induce a
change in lipid concentration. Briefly, the triglyceride
concentration was higher in the vehicle-treated OLETF rats than in
the LETO rats. Following the RLP treatment, the triglyceride
concentration was slightly lower in the OLETF rats. However, their
level failed to reach that of the LETO rats (Fig. 3B). The LDL concentrations tended
to decrease in RLP-treated OLETF rats when compared with the
vehicle-treated OLETF rats (Fig.
3D), whereas the cholesterol concentration was maintained at a
constant level in the RLP-treated group (Fig. 3C). Therefore, RLP treatment may
contribute to the reduction of the concentration of some lipids in
the sera of OLETF rats.
RLP effects on abdominal fat and
adipocyte size of OLETF rats
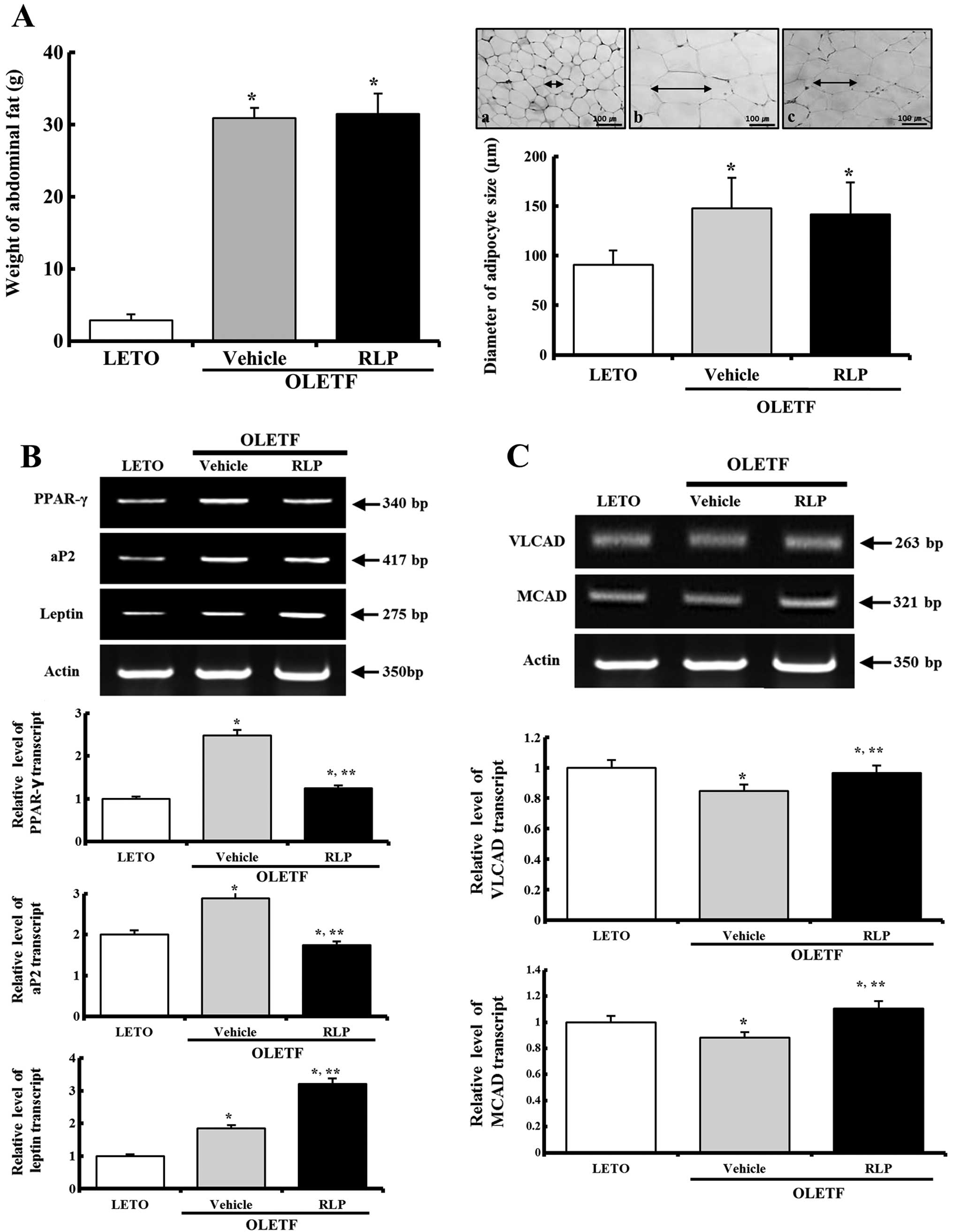
The fat mass and size of adipocytes were measured
under an optical microscope to determine if RLP treatment affects
the abdominal fat. Prior to RLP treatment, vehicle-OLETF rats
showed a higher fat mass than the LETO rats. In particular, the
weight of abdominal fat in the vehicle-treated OLETF rats was
increased by 675% compared to the LETO rats. However, there were no
significant differences between the vehicle-treated and RLP-treated
OLETF rats (Fig. 4A). In
addition, the mean size of the adipocytes was 161% larger in the
vehicle-treated OLETF rats compared to the LETO rats. Nevertheless,
no changes were detected in the RLP-treated group compared with the
vehicle-treated group (Fig. 4A).
These results suggest that RLP treatment has little effect on the
fat mass and adipocyte size in OLETF rats.
RLP effects on the expression of
adipocyte marker genes in abdominal fat
Hypertrophic adipocytes in obese rodents
over-express several genes related to the obese metabolism.
Therefore, the level of PPAR-γ, aP2 and leptin expression was
measured in the abdominal fat of OLETF rats to determine the effect
of the RLP treatment on the expression of adipocyte marker genes.
Prior to RLP treatment, the level of PPAR-γ, aP2 and leptin
expression was slightly higher in the OLETF rats than in the LETO
rats. On the other hand, in the RLP-treated OLETF rats, the levels
of PPAR-γ and aP2 expression were decreased by 49 and 39% compared
to the vehicle-treated OLETF rats (Fig. 4B). Leptin expression was even
higher in the RLP-treated group. These results suggest that RLP can
induce the downregulation of leptin and PPAR-γ to regulate the
obese metabolism.
RLP effects on the expression of the gene
responsible for fatty acid β-oxidation in abdominal fat
VLCAD and MCAD are important enzymes for fatty acid
oxidation (24). The levels of
these enzymes were detected in the abdominal fat by RT-PCR to
determine the effect of RLP treatment on the expression of VLCAD
and MCAD. In the obese condition, the expression of these enzymes
was slightly lower compared to the normal condition. On the other
hand, the RLP-treated OLETF rats showed an increase in VLCAD and
MCAD expression compared to the vehicle-treated OLETF rats
(Fig. 4C). Therefore, these
results show that the expression of VLCAD and MCAD was
significantly increased by the RLP treatment for 3 weeks.
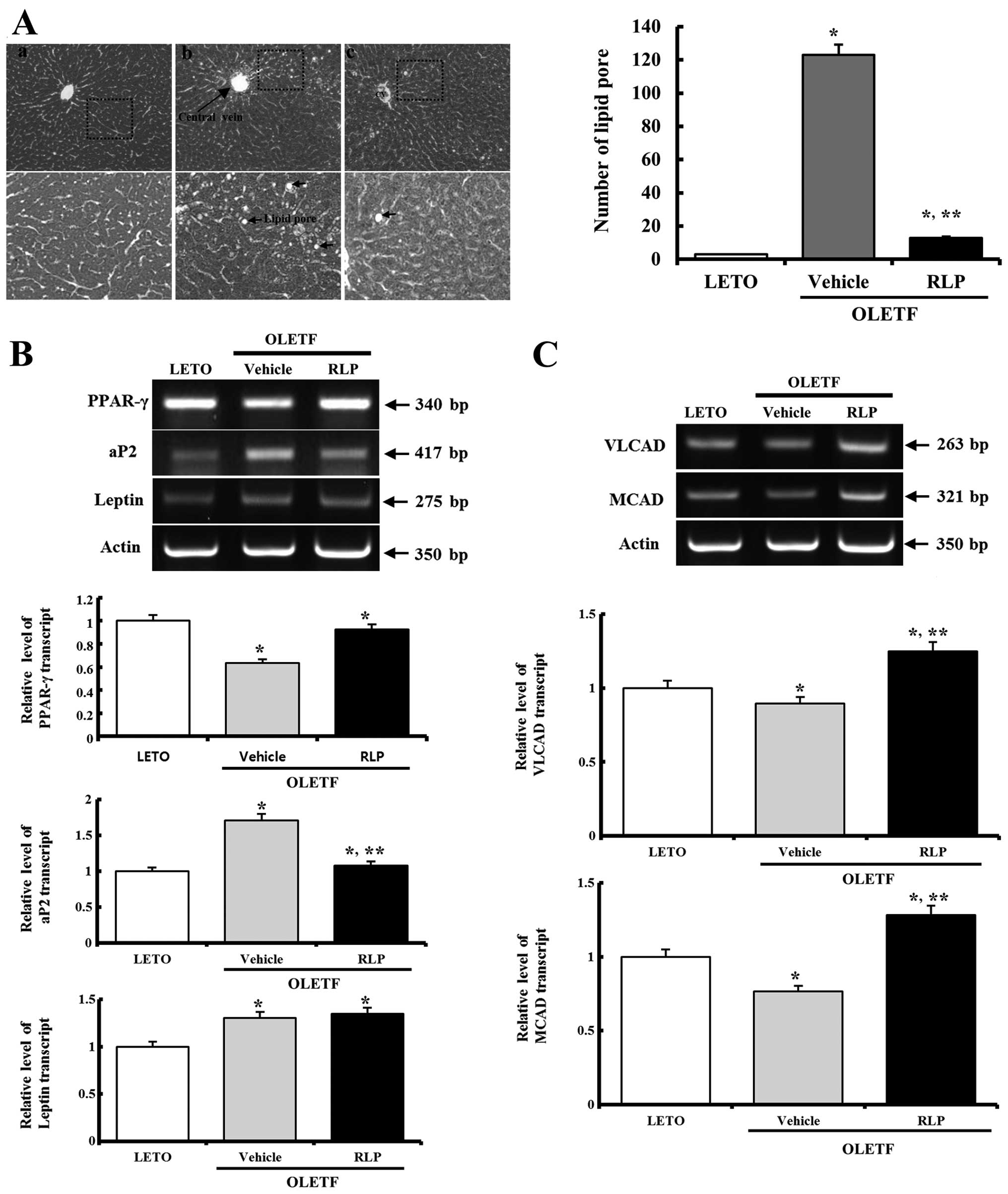
RLP effects on the fatty liver formation
and expression of the gene responsible for fatty acid β-oxidation
in the liver
To determine if the RLP treatment affects the lipid
metabolism of the liver, the alteration of fat accumulation and
gene expression for fatty acid β-oxidation was observed in the
liver tissue. Fatty liver formation was higher in the OLETF rats
compared to the LETO rats. However, following RLP treatment, these
levels in the OLETF rats decreased significantly to near the level
of the LETO rats (Fig. 5A). In
addition, of the 3 adipocyte marker genes, aP2 and leptin
expression was increased significantly in the liver of the OLETF
rats, even though PPAR-γ was decreased in the OLETF rats. However,
these alteration patterns were transformed into different ones. The
expression levels of the PPAR-γ and aP2 genes in RLP-treated OLETF
rats were recovered to those measured in the LETO rats, but leptin
expression was increased to more than that observed in the LETO
rats (Fig. 5B). Therefore, RLP
can induce the regulation of PPAR-γ, aP2 and leptin expressions to
control the obese metabolism.
In the case of gene expression analysis for fatty
acid β-oxidation, the expression level of the VLCAD and MCAD genes
were significantly lower in the OLETF rats than in the LETO rats.
However, these levels were higher in the RLP-treated OLETF rats
than in the vehicle-treated OLETF rats (Fig. 5C). Therefore, RLP treatment can
induce gene expression for fatty acid β-oxidation in the liver of
OLETF rats.
Discussion
LP and RLP have attracted considerable attention as
novel therapeutic drugs (3,8,16).
As part of the research into the development of drugs for obesity
and diabetes, this study examined the effects of RLP on the glucose
and fat metabolism using a type II diabetes model with obesity. The
results suggest that RLP treatment can induce a decrease in the
symptoms developed in diabetes and obesity.
The OLETF rats used in this study were first
developed by the selective breeding of rats showing spontaneous
obesity in an outbred colony of Long-Evans rats at Otsuka
Pharmaceuticals (25). The OLETF
rats consumed 30% more food than the LETO rats due to the fact that
these rats have a food intake disorder induced by a cholecystokinin
(CCK) 1 receptor defect. Therefore, they become 35% more obese than
LETO rats (26). On the other
hand, the OLETF rats showed different properties to the other
obesity rat model on fat deposition. Most of the fat in the OLETF
rats was deposited in the intra-abdominal or visceral areas,
whereas the fat in Zucker rats was deposited in subcutaneous areas
(27). In this study, the body
weight of the OLETF rats was 25% higher than in the LETO rats. In
addition, the abdominal fat was increased significantly in the
OLETF rats. Therefore, the OLETF rats used in this experiment
showed spontaneous obesity.
There are few reports on the relationship between LP
and abdominal fat. In particular, Gyeongshingangjeehwan (GGEx),
which is comprised of Liriope platyphylla F.T. Wang and T.
Tang (Liliaceae), Platycodongrandiflorum A. DC.
(Campanulaceae), Schisandrachinensis K. Koch (Magnoliaceae)
and Ephedra sinica Stapf (Ephedraceae), exhibited
anti-obesity abilities. Compared to the obese OLETF control rats,
administration of GGEx for 8 weeks significantly decreased the food
intake and plasma leptin levels as well as the body weight gain and
abdominal fat in OLETF rats (6).
In the present study, a significant change in the body weight and
abdominal fat was not detected in the vehicle- and RLP-treated rats
for 3 weeks (Figs. 2 and 4). The main reasons for these
differences were the administration period and the composition of
the drug; however, further research is required.
Insulin produced by pancreatic β-cells plays a key
role in regulating the carbohydrate and fat metabolism in an animal
body. Insulin induces glucose uptake from the blood to the cells in
the liver, muscle and fat tissue, and stores it as glycogen in the
liver and muscle (28,29). In vitro analysis showed a
dramatic increase in insulin secretion in RLP-treated INS cells
compared to the vehicle-treated group (16). Therefore, this study aimed to
confirm these results in an animal model for type II diabetes with
obesity. As shown in Fig. 2D, the
insulin concentration was significantly higher in the RLP-treated
OLETF rats than in the vehicle-treated OLETF rats. Therefore, these
results provide strong evidence that RLP treatment can induce an
increase in insulin concentration in OLETF rats.
Adiponectin is considered an important hormone that
is involved in the process of regulating the fatty acid catabolism
and glucose regulation (22).
This hormone also contributed to suppress the metabolic disorders
including diabetes, obesity, atherosclerosis and non-alcoholic
fatty liver disease (22,30). Adiponectin is maintained at
approximately 5–10 μg/ml in the bloodstream of humans and
the level is lower in diabetics than non-diabetics. In healthy
human adults, the concentration of this hormone was inversely
related to the body fat mass. The decrease in body weight induced a
significant increase in adiponectin concentration (31). In the present study, the
concentration of adiponectin was significantly higher in the OLETF
rats. This concentration was also increased following RLP
treatment. This differs from previous findings in that the
adiponectin level was inversely related to the body fat mass in
healthy humans. This difference may be caused by a metabolic
disorder including a CCK1 receptor defect in OLETF rats.
Under diabetic conditions induced by the
administration of streptozotocin, the total cholesterol, HDL,
triglycerides, creatinine and urea concentrations were
significantly higher than under non-diabetic conditions (23). Of these lipids, the circulating
triglycerides were decreased by GGEx treatment for 8 weeks in the
OLETF rats (6). Our findings from
RLP treatment for 3 weeks are in agreement with previous studies,
even though the rate of decrease was different. These results
suggest that LP can effectively induce a decrease of triglycerides
in serum, regardless of the administration period and concentration
of LP used.
Acyl-CoA dehydrogenase is a catalytic enzyme that
participates in the initial step in each cycle of fatty acid
oxidation in the mitochondria of cells. This enzyme can be
classified into 3 groups based on their specificity for long-,
medium- and short-chain fatty acid substrates (24). Of the 3 enzymes, the levels of
VLCAD and MCAD were significantly lower in the visceral white
adipose tissue of the OLETF rats compared to the LETO rats and some
polyherbal drugs and GGEx could recover the levels (19). VLCAD and MCAD were also expressed
in the liver of rats and play a key role in fatty acid oxidation.
Furthermore, their expression was regulated by several diet types.
Following a high fat diet, the mRNA level of the VLCAD gene was
increased considerably in the liver of rats compared to that in the
liver of rats fed low fat chow, whereas there was no change in the
mRNA level of the VLCAD gene in the muscle of the same rats
(32). In addition, 10% TG + 4%
α-linolenic acid-rich diacylglycerol diets for a period of 1 month
induced an upregulation of β-oxidation activity, as well as the
acyl-CoA oxidase (ACO) and MCAD mRNA levels (33). In this study, the levels of VLCAD
and MCAD expression in the abdominal fat and liver of OLETF rats
were also increased by the RLP treatment for 3 weeks, even though
the increase rate was different in both. This study suggests that
RLP treatment can induce VLCAD and MCAD expression in the abdominal
fat and liver tissue of OLETF rats.
Our results suggest that RLP induces an increase in
the insulin and adiponectin concentration, as well as a significant
change in the lipid concentration and expression of adipocyte
marker genes in the diabetic and obese OLETF rats. Therefore, RLP
may be considered a new potential treatment for obesity through its
ability to reduce the lipid metabolism.
Acknowledgements
The authors thank Jin Hyang Hwang, the
animal technicians, for directing the Animal Facility and Care at
the Laboratory Animal Resources Center. This study was supported by
grants to Dr Dae Youn Hwang from the Korea Institute of Planning
Evaluation for Technology of Food, Agriculture, Forestry and
Fisheries (110119-3).
References
|
1.
|
MK HuhHW HuhJS ChoiBK LeeGenetic diversity
and population structure of Liriope platyphylla (Liliaceae)
in KoreaJ Life Sci17328333200710.5352/JLS.2007.17.3.328
|
|
2.
|
YC LeeJC LeeYB SeoYB KookLiriopis
tuber inhibits OVA-induced airway inflammation and bronchial
hyperresponsiveness in murine model of asthmaJ
Ethnopharmacol101144152200510.1016/j.jep.2005.04.030
|
|
3.
|
SB ChoiJD WhaS ParkThe insulin sensitizing
effect of homoisoflavone-enriched fraction in Liriope
platyphylla Wang et Tang via PI3-kinase pathwayLife
Sci7526532664200410.1016/j.lfs.2004.04.03915369701
|
|
4.
|
SW KimIM ChangKB OhInhibition of the
bacterial surface protein anchoring transpeptidase sortase by
medicinal plantsBiosci Biotechnol
Biochem6627512754200210.1271/bbb.66.275112596883
|
|
5.
|
J HurP LeeJ KimAJ KimH KimSY KimInduction
of nerve growth factor by a butanol fraction of Liriope
platyphylla in C6 and primary astrocyte cellsBiol Pharm
Bull2712571260200410.1248/bpb.27.1257
|
|
6.
|
S JeongK ChaeYS JungYH RhoJ LeeJ HaKH
YoonGC KimKS OhSS ShinM YoonThe Korean traditional medicine
Gyeongshingangjeehwan inhibits obesity through the regulation of
leptin and PPARalpha action in OLETF ratsJ
Ethnopharmacol119245251200810.1016/j.jep.2008.06.037
|
|
7.
|
J HurP LeeE MoonI KangSH KimMS OhSY
KimNeurite outgrowth induced by spicatoside A, a steroidal saponin,
via the tyrosine kinase A receptor pathwayEur J
Pharmacol620915200910.1016/j.ejphar.2009.08.01619695245
|
|
8.
|
YK LeeJE KimSH NamJS GooSI ChoiYH ChoiCJ
BaeJM WooJS ChoDY HwangDifferential regulation of the biosynthesis
of glucose transporters by the PI3-K and MAPK pathways of insulin
signaling by treatment with novel compounds from Liriope
platyphyllaInt J Mol Med27319327201121165549
|
|
9.
|
JE KimSH NamSI ChoiIS HwangHR LeeMJ JangCY
LeeHJ SoonHS LeeHS KimAqueous extracts of Liriope
platyphylla are tightly-regulated by insulin secretion from
pancreatic islets and by increased glucose uptake through glucose
transporters expressed in liver hepatocytesBiomol
Ther193483562011
|
|
10.
|
K KimHY KimKorean red ginseng stimulates
insulin release from isolated rat pancreatic isletsJ
Ethnopharmacol120190195200810.1016/j.jep.2008.08.00618773949
|
|
11.
|
JM LuQ YaoC ChenGinseng compounds: an
update on their molecular mechanisms and medical applicationsCurr
Vasc Pharmacol7293302200910.2174/15701610978834076719601854
|
|
12.
|
TB NgPharmacological activity of sanchi
ginseng (Panax notoginseng)J Pharm
Pharmacol5810071019200610.1211/jpp.58.8.000116872547
|
|
13.
|
D KieferT PantusoPanax ginsengAm Fam
Physician6815391542200314596440
|
|
14.
|
NI BaekDS KimYH LeeJD ParkCB LeeSI
KimGinsenoside Rh4, a genuine dammarane glycoside from Korean red
ginsengPlanta Med628687199610.1055/s-2006-9578168720394
|
|
15.
|
TK YunYS LeeKH KwonKJ ChoiSaponin contents
and anticarcinogenic effects of ginseng depending on types and ages
in miceActa Pharmacol Sin1729329819969812705
|
|
16.
|
SI ChoiHR LeeJS GooJE KimSH NamIS HwangYJ
LeeSH PrakHS LeeJS LeeEffects of steaming time and frequency for
manufactured red Liriope platyphylla on the insulin
secretion ability and insulin receptor signaling pathwayLab Anim
Res27117126201110.5625/lar.2011.27.2.11721826171
|
|
17.
|
SD KimYS KuIZ LeeID KimKS YounGerneral
components and sensory evaluation of hot water extract from
Liriopis TuberJ Korean Soc Food Sci Nutr3020242001
|
|
18.
|
JE KimYK LeeSH NamSI ChoiJS GooMJ JangHS
LeeHJ SonCY LeeDY HwangThe symptoms of atopic dermatitis in NC/Nga
mice were significantly relieved by the water extract of Liriope
platyphyllaLab Anim
Res26377384201010.5625/lar.2010.26.4.377
|
|
19.
|
SS ShinYS JungKH YoonS ChoiY HongD ParkH
LeeBI SeoHY LeeM YoonThe Korean traditional medicine
gyeongshingangjeehwan inhibits adipocyte hypertrophy and visceral
adipose tissue accumulation by activating PPARalpha actions in rat
white adipose tissuesJ
Ethnopharmacol1274754201010.1016/j.jep.2009.09.052
|
|
20.
|
Y AritaS KiharaN OuchiM TakahashiK MaedaJ
MiyagawaK HottaI ShimomuraT NakamuraK MiyaokaParadoxical decrease
of an adipose-specific protein, adiponectin, in obesityBiochem
Biophys Res Commun2577983199910.1006/bbrc.1999.025510092513
|
|
21.
|
WS YangWJ LeeT FunahashiS TanakaY
MatsuzawaCL ChaoCL ChenTY TaiLM ChuangWeight reduction increases
plasma levels of an adipose-derived anti-inflammatory protein,
adiponectinJ Clin Endocrinol
Metab8638153819200110.1210/jcem.86.8.774111502817
|
|
22.
|
JJ DiezP IglesiasThe role of the novel
adipocyte-derived hormone adiponectin in human diseaseEur J
Endocrinol148293300200310.1530/eje.0.148029312611609
|
|
23.
|
S RoySK DontamallaAK MondruS SannigrahiPR
VeerareddyDownregulation of apoptosis and modulation of TGF-β1 by
sodium selenate prevents streptozotocin-induced diabetic rat renal
impairmentBiol Trace Elem Res13955712011
|
|
24.
|
JJ KimM WangR PaschkeCrystal structures of
medium-chain acyl-CoA dehydrogenase from pig liver mitochondria
with and without substrateProc Natl Acad Sci
USA9075237527199310.1073/pnas.90.16.75238356049
|
|
25.
|
K KawanoT HirashimaS MoriT NatoriOLETF
(Otsuka Long-Evans Tokushima Fatty) rat: a new NIDDM rat
strainDiabetes Res Clin
Pract24S317S320199410.1016/0168-8227(94)90269-07859627
|
|
26.
|
TH MoranUnraveling the obesity of OLETF
ratsPhysiol
Behav947178200810.1016/j.physbeh.2007.11.03518190934
|
|
27.
|
Y MoriY IkedaSimilarity and dissimilarity
between the OLETF rats and obese subjects with NIDDMObesity and
NIDMM. Lessons from the OLETF RatK ShimaElsevier Science
BVAmsterdam2372441999
|
|
28.
|
D Le RoithY ZickRecent advances in our
understanding of insulin action and insulin resistanceDiabetes
Care24588597200111289486
|
|
29.
|
C TahaA KlipThe insulin signaling pathwayJ
Membr Biol169112199910.1007/PL00005896
|
|
30.
|
O UkkolaM SantaniemiAdiponectin: a link
between excess adiposity and associated comorbidities?J Mol
Med80696702200210.1007/s00109-002-0378-712436346
|
|
31.
|
A CoppolaR MarfellaL CoppolaE TagliamonteD
FontanaE LiguoriT CirilloM CafieroS NataleC AstaritaEffect of
weight loss on coronary circulation and adiponectin levels in obese
womenInt J
Cardiol134414416200910.1016/j.ijcard.2007.12.08718378021
|
|
32.
|
H JiMI FriedmanReduced capacity for fatty
acid oxidation in rats with inherited susceptibility to
diet-induced
obesityMetabolism5611241130200710.1016/j.metabol.2007.04.00617618960
|
|
33.
|
T MuraseM AokiI TokimitsuSupplementation
with alpha-linolenic acid-rich diacylglycerol suppresses fatty
liver formation accompanied by an up-regulation of beta-oxidation
in Zucker fatty ratsBiochim Biophys
Acta1733224231200510.1016/j.bbalip.2004.12.01515863369
|