Introduction
Cervical cancer is the third most common cancer
among women worldwide, and nearly half a million cases are
diagnosed annually. In developing countries, it is the leading
cause of cancer mortality in women (1,2).
Even with optimal treatment (primarily surgery, followed by
chemotherapy and radiotherapy), 40% of cervical cancer patients
succumb to this disease annually (3). Human papillomavirus type 16 (HPV16)
is one of the principal pathogens in cervical cancer and is
responsible for >50% of cervical cancer cases (3,4).
The US Food and Drug Administration has approved 2 prophylactic HPV
vaccines, Gardasil (Merck) and Cervarix (GlaxoSmithKline) to
prevent cervical cancer and the prophylactic vaccines can
significantly reduce the incidence of HPV-related cancers (1,5).
However, since the vaccine is based on the virion capsid proteins
(L1, L2), which are absent in cervical cancer, they are unlikely to
be effective in controlling pre-existing HPV infections or
HPV-associated lesions (6).
Therefore, it is necessary to develop both prophylactic and
therapeutic vaccines against cervical cancer. The 2 major HPV
oncoproteins, E6 and E7, which are consistently expressed in
cervical cancer cells, are essential for the malignant
transformation and maintenance of tumor cells (7). Cellular immunity to E7/E6 is
associated with the clearance of premalignant HPV16 lesions,
therefore, E6 and E7 are ideal targets for cervical cancer
immunotherapy (8).
Listeria monocytogenes (LM) as a vaccine
vector has been widely used to deliver tumor-associated antigens
(TAAs) for cancer immunotherapy (9,10).
L. monocytogenes is a facultative intracellular bacterium
that has the unusual ability to escape from the phagosome and
multiply in the cytoplasm of cells (11). L. monocytogenes directly
infects antigen-presenting cells (APCs), such as dendritic cells
and macrophages, thereby delivering TAAs into their cytoplasm,
resulting in processing and presentation of TAAs to the immune
system, which induces antigen-specific CD4+ and
CD8+ T cell responses (12,13). Hence, recombinant L.
monocytogenes-based vaccines expressing TAAs, including
endoglin (CD105) (14), human
prostate-specific antigen (PSA) (15), HER-2/neu (16) and HPV16 E7 (17,18) have been developed for cancer
immunotherapy in preclinical and clinical trials. Previously, Gunn
et al (19) developed an
LM-LLO-E7 vaccine, which carried a plasmid that contained an
hly promoter to drive the expression of the LLO-E7 fusion
protein in the presence of chloramphenicol. Verch et al
reported that Listeria-based antibiotic resistance gene-free
vaccine LMdd (pTV3) was constructed, but only attenuates the vector
by 0.5–1 log (20,21). However, for clinical application,
the safety and ability of a live recombinant vaccine to stably
express foreign antigens need to be assessed.
In the current study, an attenuated L.
monocytogenes with deletions of actA and plcB
(LM▵actA/plcB, in brief: LM1-2) was employed as a
vaccine vector to deliver HPV16 E7. ActA is a major virulence
protein that induces the rapid polymerization of filamentous actin
and propels L. monocytogenes through the cytoplasm and into
neighboring cells (22). Pamer
(23) reported that
ActA-deficient mutants are highly attenuated. Additionally, PlcB
has been demonstrated to be important in second vacuolar escape
(24). Previous studies analyzed
a double mutant with actA and plcB in adult
volunteers and indicated that the mutant was a safe vector for
clinical application (25,26).
Also, the E7 antigen was integrated into the chromosome of L.
monocytogenes using the integration vector pIMK2-SPhly, which
provides an effective tool to secretly express the heterologous
proteins (27).
Results of the present study showed that the highly
attenuated LM1-2-E7 strain induced E7-specific cell-mediated
immunity and exhibited significant prophylactic and therapeutic
efficacy against cervical cancer in a murine model. In addition,
the antitumor efficacy was associated with intratu-moral
CD8+ T cell infiltration and preferential accumulation
of LM1-2-E7 within tumor xenografts.
Materials and methods
Peptide, bacteria and plasmid
HPV16 E749-57 (RAHYNIVTF) peptide for the
H-2Db restricted epitope was synthesized by Beijing
Scilight Biotechnology, LLC. (Beijing, China). The plasmid
pIMK2-SPhly was generously provided by Professor Chakraborty
(Justus Liebig University, Giessen, Germany). Strain yzuLM4
(serotype 1/2a) was isolated and preserved in our laboratory. The
LM1-2 strain was actA and plcB double mutant, which
was previously constructed by homologous recombination (28).
Mice, cell lines and media
C57BL/6 mice (6–8 weeks old) were purchased from the
Comparative Medical Center of Yangzhou University. Animals were
housed and used in accordance with the protocols approved by the
institutional animal experimental committee. The TC-1 cell line was
purchased from Beijing Hualisentai Bio-Scientific, Co., Ltd.
(Beijing, China). TC-1 cells are C57BL/6 lung tumor epithelial
cells immortalized with HPV16 E6/E7 and transformed with the
c-Ha-ras oncogene. Cells were cultured in RPMI-1640, supplemented
with 10% fetal calf serum (FCS), 1 mmol/l sodium pyruvate, 100 U/ml
penicillin and 100 μg/ml streptomycin in a 37°C incubator
with 5% CO2.
Construction of the recombinant LM1-2-E7
strain
The E7 fragment was amplified and cloned into vector
pIMK2-SPhly using primers E7 forward, (GACGGATCCCATGGAGATAC ACCTAC) and
E7 reverse (CCGCTCGAGTTATGGTTTCTG AGAACA) with
restriction enzyme sites BamHI and XhoI. The
resulting plasmid pIMK2-SPhly-E7 and control plasmid pIMK2-SPhly
were electroporated into competent LM1-2, which were designed as
the recombinant L. monocytogenes strain LM1-2-E7 and
LM1-2-control, respectively.
Western blotting
LM1-2-E7 and LM1-2-control were cultured overnight
in brain heart infusion (BHI) broth at 37°C and their supernatants
were collected by centrifugation at 8,000 rpm for 10 min. Secreted
proteins in the culture supernatants were precipitated with
trichloroacetic acid (TCA; Sigma-Aldrich, St. Louis, MO, USA) and
resuspended in 1.5 mM Tris-HCl buffer. Then, the precipitates were
lysed by ultrasonic waves. The secreted proteins and lysates were
separated by 4–12% SDS-PAGE, transferred to a nitrocellulose
membrane and detected using an anti-E7 monoclonal antibody (Clone
8C9; Invitrogen Life Technologies, Carlsbad, CA, USA). The signals
were measured using ECL detection reagents (Thermo Scientific,
Rockford, IL, USA) and exposure to Hyperfilm.
In vitro stability assay
The stability of LM1-2-E7 was determined by serially
passaging for 40 times without antibiotics and subsequently
analyzing the expression of E7 in the 20th, 30th and 40th
passages.
Immunization of mice with LM1-2-E7
C57BL/6 mice (6–8 weeks old) were intraperitoneally
immunized with LM1-2-E7 (0.1 LD50, 5×107 CFU)
or LM1-2-control (0.1 LD50, 5×107 CFU) or
phosphate-buffered saline (PBS) buffer on days 0 and 7. Seven days
after the booster immunization, mice were used for ELISPOT assay,
cytotoxicity assay in vivo and histopathological study. The
groups treated with PBS buffer or LM1-2-control were used as
negative controls.
Enzyme-linked immunosorbent spot
(ELISPOT) assay
The ELISPOT assay was performed as previously
described (29). The spleens were
harvested and processed into single cell suspensions. The cell
suspensions were treated with ammonium chloride (ACK) buffer to
lyse the erythrocytes and washed twice with complete RPMI-1640
medium. A total of 5×105 cells/well was incubated in anti-murine
interferon (IFN)-γ/interleukin (IL)-4-coated ELISPOT plates. The
cells were stimulated in triplicate in C-RPMI medium as a negative
control and pulsed with 2 μM E749-57 (RAHYNIVTF)
peptide or with concanavalin A (ConA, 5 μg/ml; Sigma, St.
Louis, MO, USA) as a positive control. After 48 h of incubation at
37°C, the plates were developed according to the manufacturer’s
protocol (BD Pharmingen, San Diego, CA, USA). The monoclonal
antibodies that were used for the ELISPOT assay were R4-6A2 for
IFN-γ, XMG1.2 for biotinylated IFN-γ, BVD4-1D11 for IL-4 and
BVD6-24G2 for biotinylated IL-4 (BD Pharmingen). The spots were
counted using an automated ELISPOT Bioreader 5000 (ImmunoBioSystem,
The Colony, TX, USA).
Cytotoxicity assay
To determine specific cytotoxicity in vivo,
the splenocytes were pooled from naive C57BL/6 mice and divided
into 2 groups: the cell suspension for 1 group was incubated with
the E7 peptide at 37°C for 45 min and subsequently labeled with 2.5
μM Carboxyfluorescein succinimidyl ester
(CFSEhigh; Molecular Probes, Invitrogen Life
Technologies) buffer at 37°C for 10 min, whereas another group was
incubated without the peptide and labeled with CFSElow
(0.25 μM) buffer. CFSEhigh and CFSElow
cells were mixed in a 1:1 ratio and 107 cells were
intravenously injected into immunized mice. Twenty-four hours
later, the spleens of the immunized mice were processed into single
cell suspensions and analyzed by flow cytometry (FACScan; Becton
Dickinson) to determine the ratio of CFSEhigh to
CFSElow cells. The percentage of specific lysis was
calculated using the following formula: percent specific
lysis=100−[100× (% CFSEhigh immunized/%
CFSElow immunized)/(% CFSEhigh control/%
CFSElow control)] (30).
Histopathological study
The sections of the spleens and livers were fixed in
13% neutral buffered formalin. Paraffin-embedded sections were cut
at 5 μm, stained with hematoxylin and eosin (H&E), and
examined for histological lesions under a microscope (Leica
Microsystems, Wetzlar, Germany).
Prophylactic and therapeutic tumor load
experiments
In a prophylactic experiment, C57BL/6 mice (6–8
weeks old, 8 per group) were intraperitoneally immunized with
LM1-2-E7 or LM1-2-control or PBS buffer on days 0 and 7 and
subcutaneously challenged on day 11 with 2×105 TC-1
cells. The mice were monitored for tumor formation.
In a therapeutic setting, C57BL/6 mice (8 per group)
were subcutaneously injected with 2×105 TC-1 cells on
the left flank. When the tumor size reached an average diameter of
5 mm on day 7 after tumor cell inoculation, the mice received
LM1-2-E7 or LM1-2-control or PBS buffer intraperitoneally on days 7
and 14. The tumors were monitored every 3 days with calipers and
the longest and shortest surface diameters were recorded for each
individual tumor. Tumor volume was calculated as the following:
length × (width)2/2 (31).
Analysis of CD8+ T cells in
the tumor
The tumors in the immunized groups were excised on
day 7 after the second immunization, minced using a sterile razor
blade and digested with a buffer containing 2 mg/ml collagenase
type I and 12 U/ml DNase in PBS buffer. After a 2 h incubation at
37°C with agitation, single cell suspensions were harvested after
filtration through a nylon mesh, stained with anti-CD3-FITC
(clone145-2C11; BD Pharmingen) and anti-CD8-APC monoclonal
antibodies (clone53-6.7; BD Pharmingen) and analyzed by FACS.
For immunohistochemistry analysis, tumor specimens
in the immunized groups were fixed in 13% neutral buffered
formalin. Paraffin-embedded sections were cut at 5 μm and
incubated with anti-mCD8 (clone53-6.7; R&D Systems,
Minneapolis, MN, USA) overnight at 4°C. The anti-mCD8 antibody was
visualized through HRP-DAB Cell and Tissue Staining kit (R&D
Systems) and counterstained with H&E.
Bacterial translocation studies
Tumor-bearing mice (7 days after the tumor cell
inoculation) received a single intraperitoneal immunization (0.1
LD50 LM1-2-E7). The spleens, livers and tumors of 3 mice
were homogenized on days 1, 2, 3 and 5 post-immunization. The
bacterial numbers were determined by plating the cell suspensions
on BHI agar. The tumor tissues at 1 day post-immunization were cut
into smaller sections, fixed in a solution of 2.5% (w/v)
glutaraldehyde in 0.1 M cacodylate buffer, dehydrated and embedded
in Epon. Ultrathin sections were cut and stained with uranyl
acetate and Reynold’s lead citrate. The sections were examined
using a Transmission Electron Microscope (TEM; JEOL Ltd., Tokyo,
Japan).
Statistical analysis
Statistical analyses for in vitro and in
vivo experiments were carried out using the GraphPad Software
package (GraphPad Software, La Jolla, CA, USA). Student’s t-test
and one-way ANOVA were used for analysis of the comparisons between
the groups. Statistical significance was set at
*P<0.05, **P<0.01 and
***P<0.001.
Results
Construction of LM1-2-E7 to express and
secrete the HPV16 E7 protein
The recombinant attenuated LM1-2-E7 strain, which
integrated the encoding gene of the HPV16 E7 into the chromosome of
the LM1-2 strain using pIMK2-SPhly, was constructed. Western
blotting revealed that the recombinant strain LM1-2-E7 expressed
and secreted the HPV16 E7 protein (Fig. 1A). Moreover, E7 was stably
expressed for at least 40 passages indicated by in vitro
stability assay (Fig. 1B).
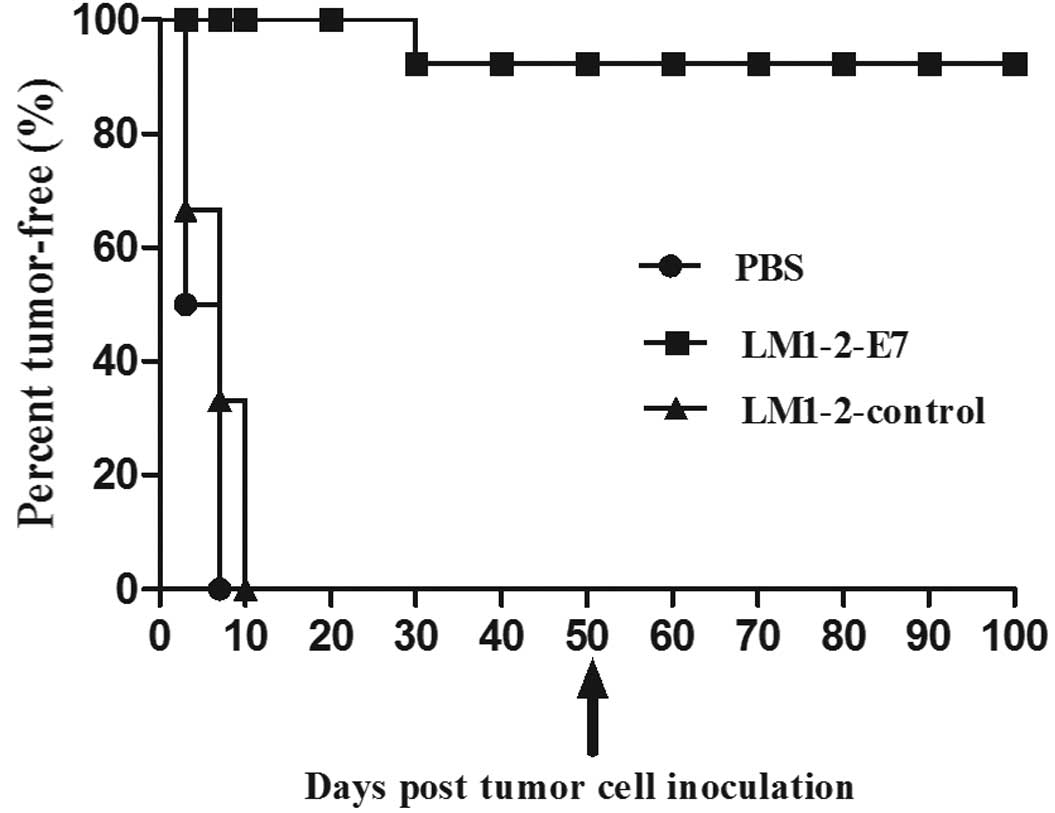
Vaccination with LM1-2-E7 protects mice
against tumor cell challenge
The preventive effect of the LM1-2-E7 vaccine was
assessed in the TC-1 tumor model. Mice were challenged with TC-1
cells on day 4 after the second immunization and were observed for
tumor development. In the LM1-2-E7-vaccinated group, 7 mice were
tumor-free until day 50, excluding 1 mouse (87.5%) that indicated
tumor growth on day 32 after the tumor cell challenge. By contrast,
tumors appeared in all of the mice on day 7 in the PBS group and on
day 10 in the LM1-2-control group (Fig. 2). The results demonstrate that the
LM1-2-E7 strain confers significant preventive efficacy.
Furthermore, 7 tumor-free mice were rechallenged with TC-1 cells on
day 50 and all of the mice rejected the tumor challenge 50 days
after the second tumor cell injection. The tumor-free mice appeared
to be healthy and presented no weight loss by the end of the study.
This finding indicates that the mice which are vaccinated with
LM1-2-E7 have long-lasting protection and elicit a memory response
against HPV16.
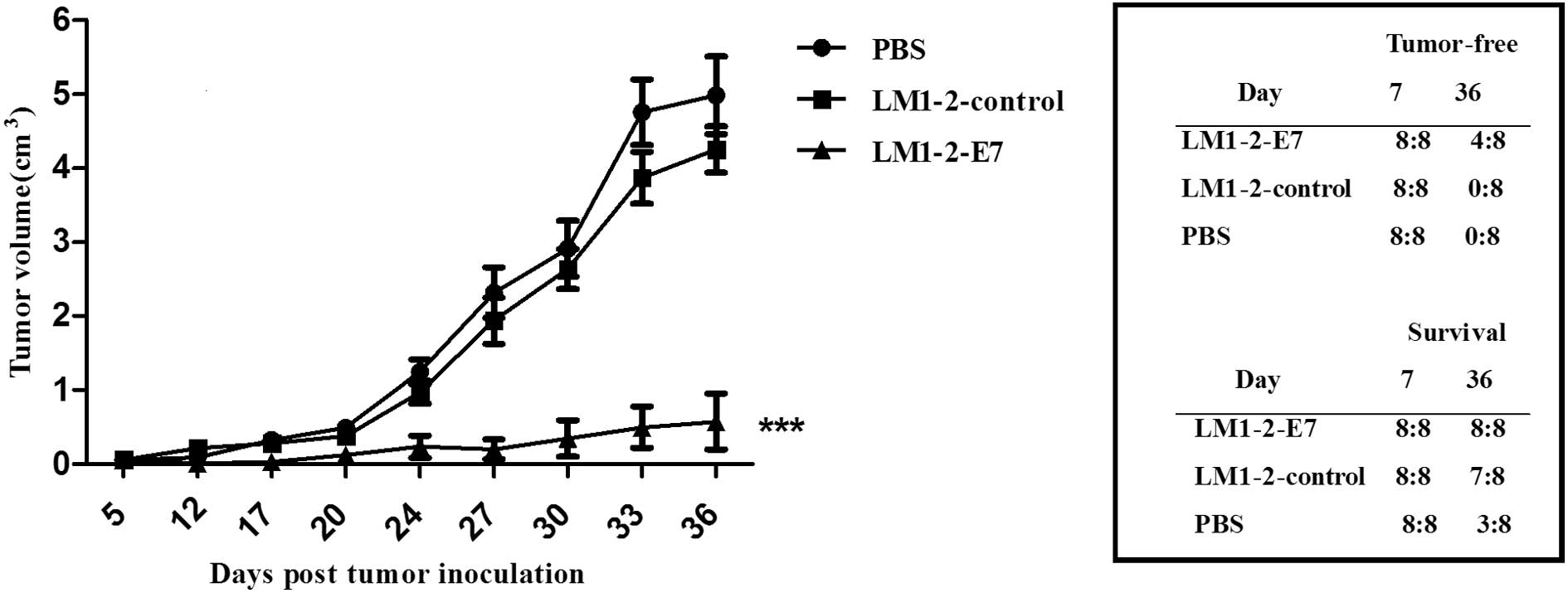
LM1-2-E7 strain causes regression of
established tumors in mice
To examine the therapeutic efficacy of the LM1-2-E7
vaccine candidate, C57BL/6 mice were subcutaneously injected TC-1
cells and intraperitoneally immunized with LM1-2-E7, LM1-2-control,
or PBS buffer on days 7 and 14 and tumor growth were measured. As
shown in Fig. 3, the tumor
volumes in the mice that were immunized with LM1-2-E7 were
significantly reduced compared to those in the LM1-2-control or the
PBS group (P<0.001). Moreover, 4 of the 8 mice in the
LM1-2-E7-immunized group remained tumor-free. Additionally, the
mice immunized with LM1-2-E7 survived tumor invasion on day 36. By
contrast, 7 mice survived tumor invasion in the LM1-2-control group
and only 3 mice in the PBS group. These results indicate that the
LM1-2-E7 strain exhibits therapeutic activity in the TC-1 mouse
tumor model.
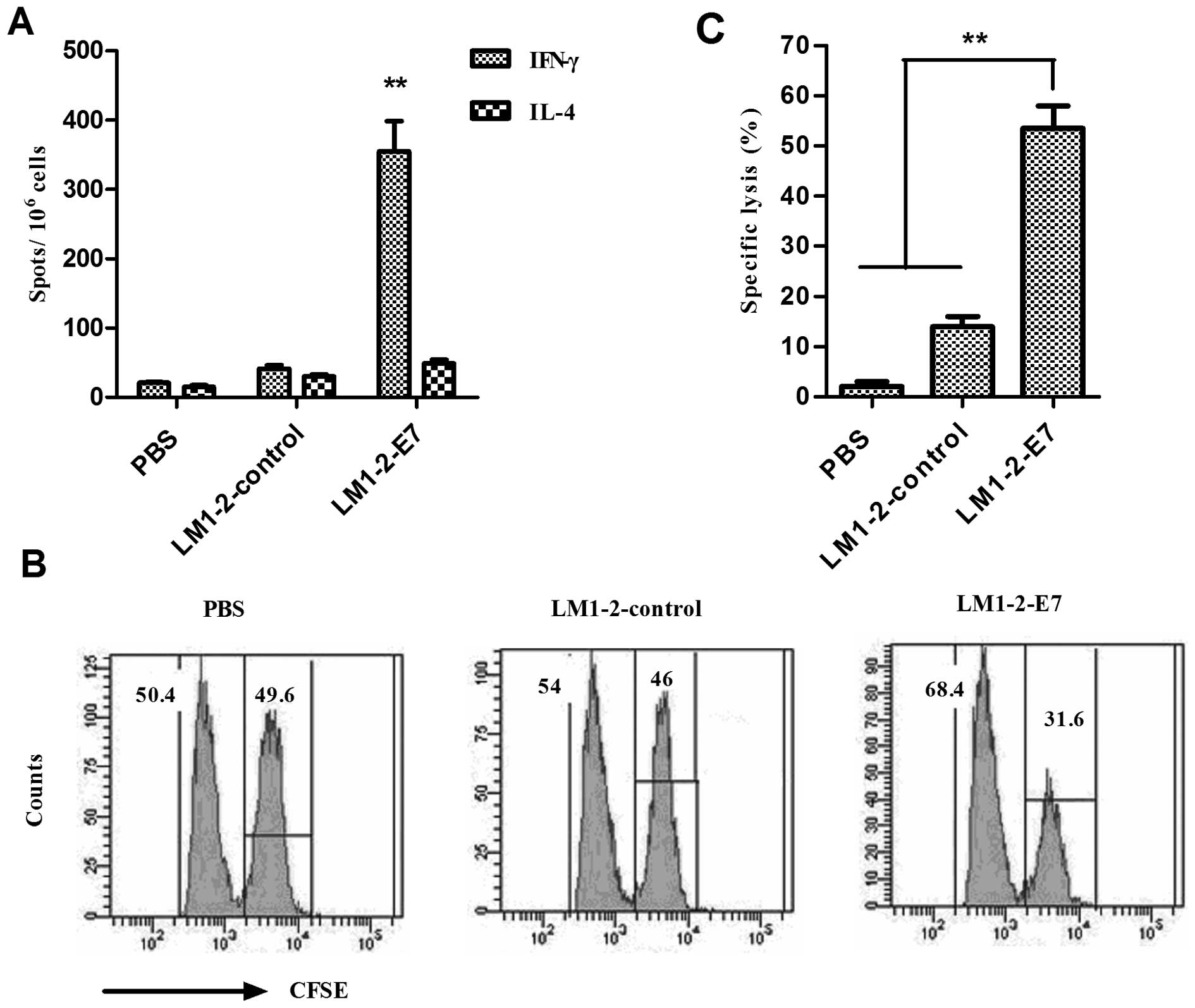
LM1-2-E7 strain elicits E7-specific
cellular immune responses in mice
In order to measure the cellular immune responses
elicited by LM1-2-E7, the splenocytes were isolated on Day 7 from
the second immunization, and the number of IFN-γ/IL-4-secreting
cells after stimulation with E749-57 peptide was
analyzed using an ELISPOT assay. The mean number of IFN-γ-secreting
cells in the LM1-2-E7-immunized group was significantly higher than
the number of IL-4-secreting cells (P<0.01) and significantly
increased compared to those of the control groups (P<0.01).
These results indicate that mice immunized with LM1-2-E7 developed
cellular immune responses against the E7 peptide (Fig. 4).
Cytotoxic T-lymphocytes (CTLs) are a critical
component of the immune response to tumors. Thus, the E7-specific
CTL responses in vivo were determined. The results indicate
that LM1-2-E7-vaccinated mice exhibited the highest cytolytic
activity (53.49±3.63%) against E749-57-loaded target
cells in comparison with the cytolytic activity (13.96±2.05%) found
in the immunized with LM1-2-control and (2.0±0.76%) in the PBS
group.
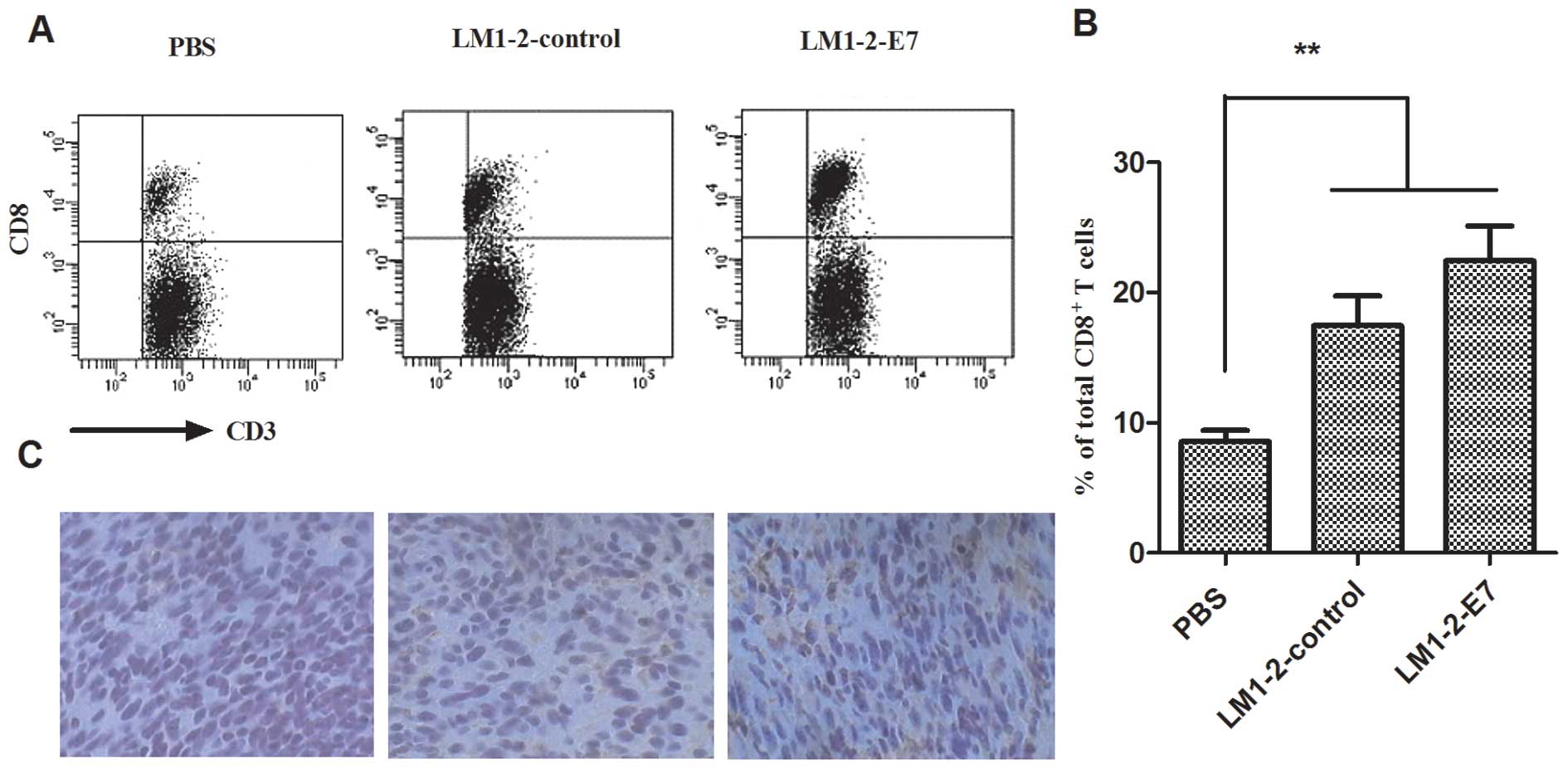
Increased frequency of tumor-infiltrating
CD8+ T cells in tumors
Tumor-infiltrating lymphocytes (TILs) are a part of
the tumor surveillance system. The CD8+ T cells were
analyzed in the tumors from all the groups. A higher number of
tumor-infiltrating CD8+ T cells (22.45±2.66%) was
detected in the LM1-2-E7 group compared to the controls
(17.45±2.1%, LM1-2-control; 8.55±0.96%, PBS group) (Fig. 5A and B). A similar result was also
found in the immunohistochemistry of the excised tumor tissues
(Fig. 5C). These results indicate
that the LM1-2-E7 vaccine could enhance antitumor immunity by
increasing the infiltration of CD8+ T cells into the
tumor.
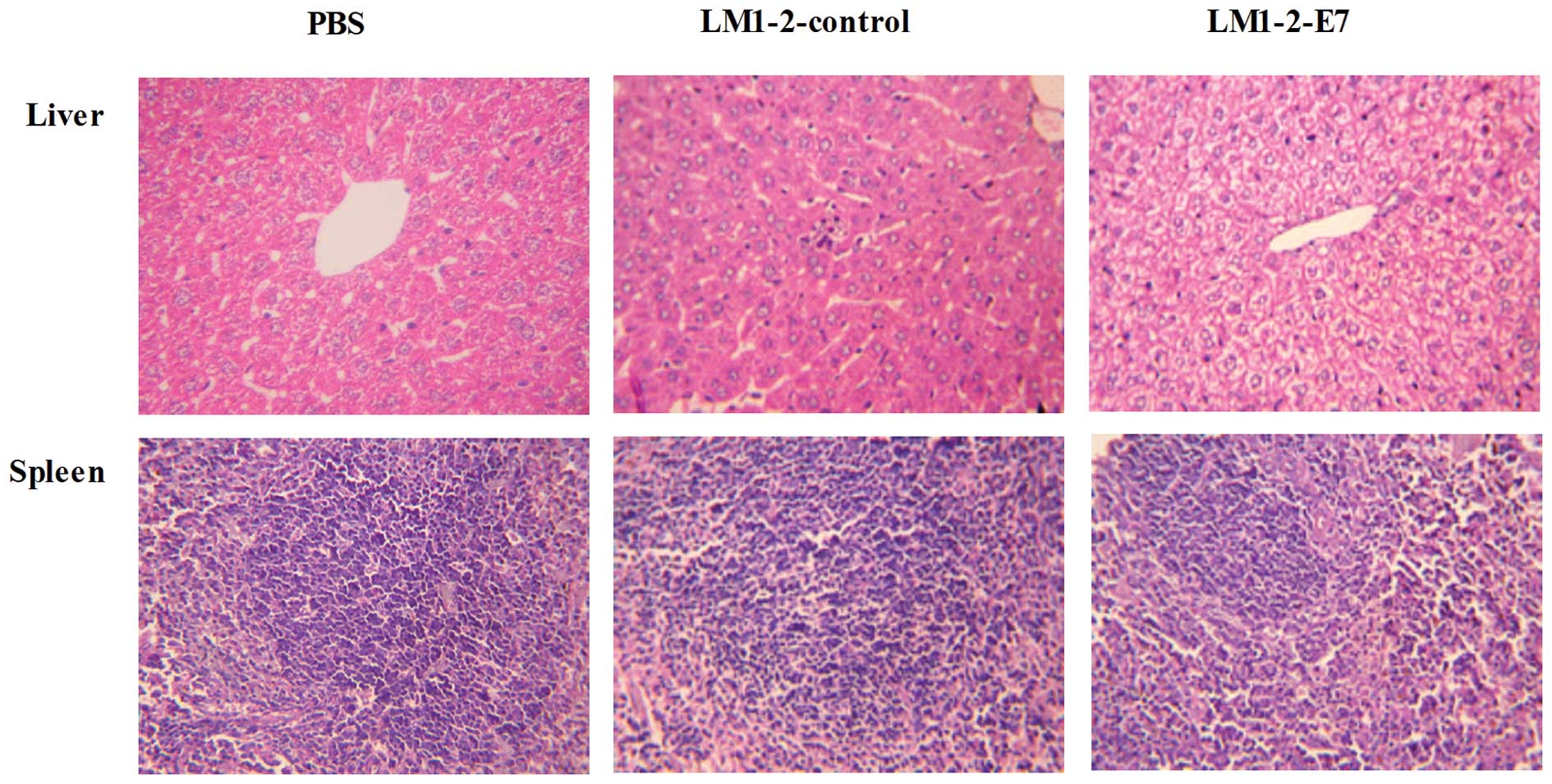
Histopathological study of the organs of
the immunized mice
The spleens from the LM1-2-E7-immunized group and
LM1-2-control group did not reveal any significant pathological
lesions and the liver sections revealed inflammatory cell
infiltration in the hepatic lobules. The results demonstrate that
LM1-2-E7 and LM1-2-control induce a mild inflammatory response with
no necrosis or structure damage (Fig.
6).
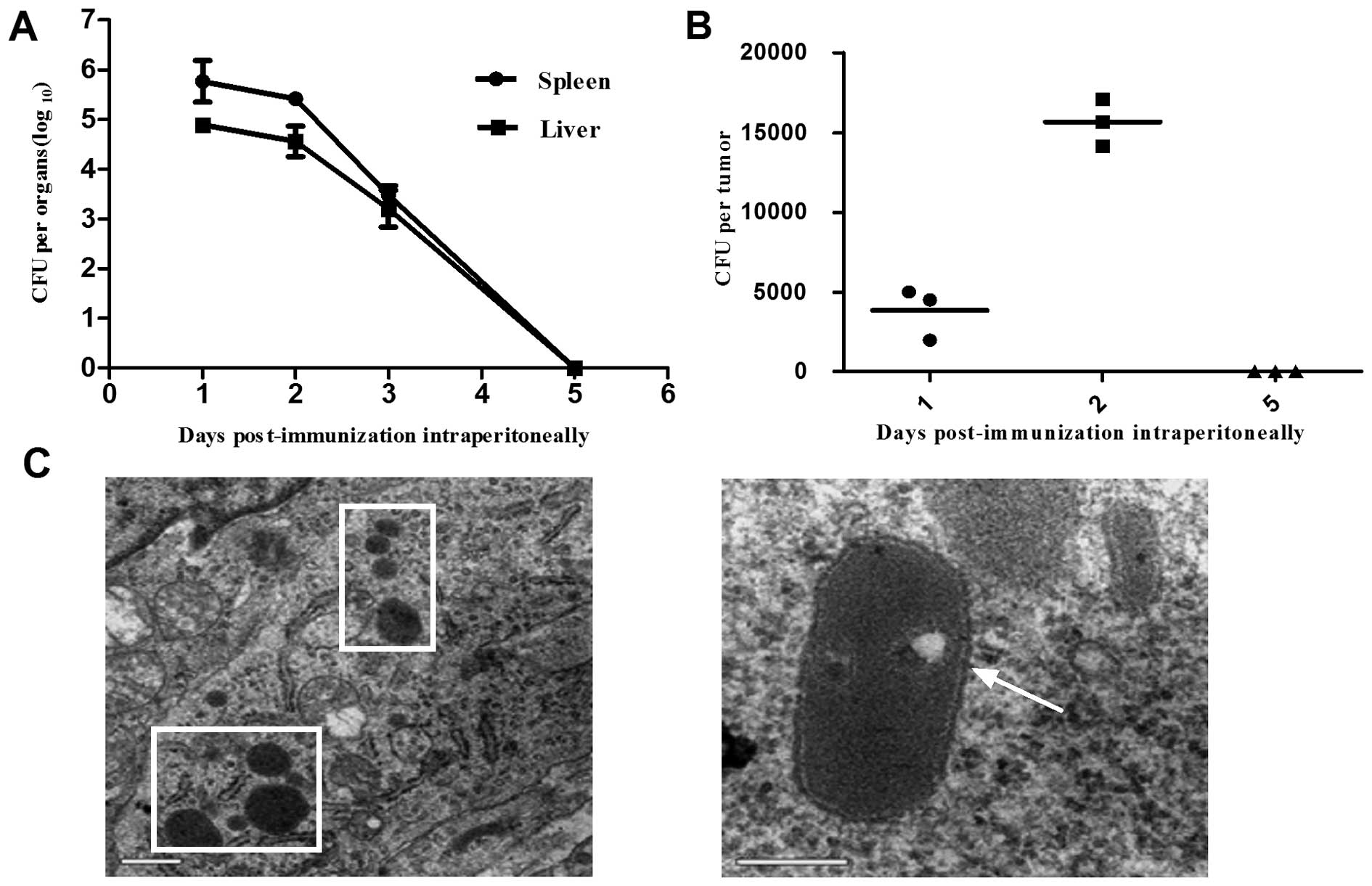
Distribution of the LM1-2-E7 strain after
a single intraperitoneal immunization
Tumor-bearing mice were intraperitoneally immunized
with a single dose of LM1-2-E7 and the number of viable bacteria
was determined in the spleen, liver and tumor homogenates. As shown
in Fig. 7A, the LM1-2-E7 bacteria
were rapidly cleared from the spleen and liver without damaging
their structures. On day 1 post-immunization, LM1-2-E7 bacteria
were detected in tumor tissue and the number of bacteria peaked on
day 2. TEM demonstrated that the LM1-2-E7 strain infected the
tumors in vivo with a high efficacy (Fig. 7C). On day 5 post-immunization,
LM1-2-E7 disappeared from the spleens, livers and tumors of the
mice (Fig. 7A and B).
Discussion
There are currently prophylactic vaccines against
HPV for clinical use, however, the cost of these vaccines is
prohibitive in developing countries (1). In addition, these prophylactic
vaccines do not generate therapeutic effects (32). The prophylactic and therapeutic
L. monocytogenes-based vaccine (LM1-2-E7) was developed in
the present study. LM1-2-E7 was able to protect 87.5% of the mice
even 50 days after the second tumor cell challenge. Also,
immunization with the LM1-2-E7 strain was able to induce tumor
regression in 50% of the mice in the therapeutic experiment.
To avoid serious toxicity of L.
monocytogenes, a highly attenuated strain
LM▵actA/plcB was selected as a vaccine vector.
Due to deletion of the virulence factors actA and
plcB, the pathogenicity of LM▵actA/plcB is
significantly decreased, the LD50 is 3 logs higher than
that of the parent strain yzuLM4 (data not shown). As indicated in
tissue sections, only lymphocyte infiltration was observed in the
livers from LM1-2-E7-immunized mice. Also, the results demonstrated
that the attenuated LM1-2-E7 strain was cleared by the immune
system within 5 days post-immunization, which is in line with the
LM▵actA/plcB strain (33). Although virulence of the LM1-2-E7
strain is reduced, LM1-2-E7 retains the ability to induce strong
E7-specific immune responses.
In addition to choosing the attenuated strain as
backbone, an integration vector pIMK2-SPhly was used for the
construction of recombinant strain. The E7 antigen was stably
integrated into the Listeria chromosome via pIMK2-SPhly and
secretly expressed by recombinant strain, which is the basis of the
E7 antigen entered into the MHC class I pathway (27). Moreover, stability assay in
vitro suggests that the E7 antigen was stably expressed for at
least 40 passages, which is appropriate for clinical
immunization.
Antigen-specific T cell responses are most critical
for the regression of established tumors and protection against
tumor challenge. IFN-γ and IL-4 secreted by Th1 cells and Th2 cells
in LM1-2-E7 immunized mice were determined by ELISPOT assay. The
increased secretion of the Th1 cytokine IFN-γ indicated that the
immune response elicited by LM1-2-E7 was biased toward the Th1 type
against the E749-57 peptide. These Th1 T cells have a
pivotal role in antitumor immunity and contribute to APC maturation
and the release of cytokines during CD8+ T cell
proliferation and differentiation (34). Of note, our cytotoxicity activity
in vivo revealed that the stimulated T cells in LM1-2-E7
immunized mice could destroy the target cells pulsed with the E7
peptide, which indicated that LM1-2-E7 could induce strong
E7-specific CTL response in vivo.
The generation of CD8+ tumor-infiltrating
lymphocytes (TILs) can be used as a surrogate marker for cancer
vaccine efficacy and activated CD8+ T cells release
cytolytic agents that attack and kill tumor cells (35). The number of tumor-infiltrating
CD8+ T cells increased in the LM1-2-E7 group compared to
the controls through the FACS and immunohistochemistry analyses,
which was strongly associated with the better therapeutic efficacy
in the LM1-2-E7-immunized mice. The higher number of
CD8+ T cells may be depended on the cytosolic location
of L. monocytogenes.
Our results show that the recombinant
Listeria strain could infect and reside in the tumors, so E7
antigen delivered by the recombinant Listeria strain is
targeted to the tumor tissue. Yu et al (36) suggested that the survival and
replication of L. monocytogenes in tumors is dependent on
the tumor immune microenvironment and vascularization, which
provides a sanctuary for bacteria to escape clearance by the immune
system. L. monocytogenes in tumors may directly lyse tumor
cells or stimulate homing effector cells by releasing cytokines and
chemokines to destroy tumor cells. Gravekamp and Paterson also
reported that L. monocytogenes can efficiently kill tumor
cells through a dual mode of action, which involves both direct
kill and CTL responses to the Listeria antigen (37).
In the present study, L. monocytogenes exerts
a strong adjuvant effect, which is associated with a strong innate
immunity induced by L. monocytogenes (12). One manifestation is that 2-fold
increase of tumor-infiltrating CD8+ T cells caused in
the LM1-2-control group compared to the PBS group. In vivo
tumor regression assay showed that LM1-2-control can prolong the
survival time of tumor-bearing mice, which mainly produced an
effect by L. monocytogenes.
The TC-1 cell line which expresses HPV E6 and E7
oncoproteins was applied to establish an HPV16-positive
cancer-associated tumor model. The TC-1 tumor model in C57BL/6 mice
is a widely-used model for use in vaccine research against cervical
cancer (38,39). Thus, the antitumor efficacy of
LM1-2-E7 was initially evaluated in the TC-1 tumor model. However,
this transplantable tumor model has several limitations, such as
the inability to accurately mimic the immunoinhibitory effects of
the tumor microenvironment (40).
Hence, other cervical cancer models (a transgenic mouse model) will
be selected to evaluate the potency of this vaccine in future
studies.
In conclusion, our results demonstrate that LM1-2 is
a possible vaccine vector, which is capable of delivering TAAs for
cancer immunotherapy. LM1-2-E7 exerts a prophylactic effect on
tumor growth and leads to the regression of established tumors
expressing E7 antigen. The antitumor efficacy was associated with
E7-specific CTL response and robust cellular immune responses
elicited by recombinant strain. The results may be of importance in
further investigations of this vaccine to combat cervical cancer
and other HPV-associated cancers.
Acknowledgements
The authors thank Professor T.
Chakraborty for providing plasmid pIMK2-SPhly. This study was
supported by grants from the National Basic Research Program of
China (2012CB518805), the Science and Technology Support Program of
Jiangsu Province (BE2012367), the Priority Academic Program
Development of Jiangsu Higher Education Institutions (PAPD), the
National Natural Science Foundation of China (no. 31101841) and the
Provincial Government of Jiangsu, China (nos. BK2011446 and
2009KJA230001).
References
|
1.
|
J CohenPublic health. High hopes and
dilemmas for a cervical cancer
vaccineScience308618621200510.1126/science.308.5722.61815860602
|
|
2.
|
A JemalF BrayMM CenterJ FerlayE WardD
FormanGlobal cancer statisticsCA Cancer J
Clin616990201110.3322/caac.20107
|
|
3.
|
G GattaMB LasotaA VerdecchiaSurvival of
European women with gynaecological tumours, during the period
1978–1989. EUROCARE Working GroupEur J
Cancer3422182225199810070290
|
|
4.
|
A SchäferW FriedmannM MielkeB
SchwartländerMA KochThe increased frequency of cervical
dysplasia-neoplasia in women infected with the human
immunodeficiency virus is related to the degree of
immunosuppressionAm J Obstet Gynecol16459359919911992708
|
|
5.
|
RM HauptC SattlerHPV vaccine continues to
be safe and effective, and its benefits continue to outweigh its
risksExpert Rev Vaccines9697701201010.1586/erv.10.5620624041
|
|
6.
|
TC WuTherapeutic human papillomavirus DNA
vaccination strategies to control cervical cancerEur J
Immunol37310314200710.1002/eji.20063697817273998
|
|
7.
|
H zur HausenPapillomaviruses and cancer:
from basic studies to clinical applicationNat Rev
Cancer2342350200212044010
|
|
8.
|
CF HungTC WuA MonieR RodenAntigen-specific
immunotherapy of cervical and ovarian cancerImmunol
Rev2224369200810.1111/j.1600-065X.2008.00622.x18363994
|
|
9.
|
V ShahabiMM SeaveyPC MaciagS RiveraA
WallechaDevelopment of a live and highly attenuated Listeria
monocytogenes-based vaccine for the treatment of
Her2/neu-overexpressing cancers in humanCancer Gene
Ther185362201120725099
|
|
10.
|
M TangneyCG GahanListeria
monocytogenes as a vector for anti-cancer therapiesCurr Gene
Ther104655201010.2174/156652310790945539
|
|
11.
|
KW BruhnN CraftJF MillerListeria as
a vaccine vectorMicrobes
Infect912261235200710.1016/j.micinf.2007.05.010
|
|
12.
|
LA ZenewiczH ShenInnate and adaptive
immune responses to Listeria monocytogenes: a short
overviewMicrobes
Infect912081215200710.1016/j.micinf.2007.05.00817719259
|
|
13.
|
A DarjiW MohamedE DomannT
ChakrabortyInduction of immune responses by attenuated isogenic
mutant strains of Listeria monocytogenesVaccine21Suppl
2102109200310.1016/S0264-410X(03)00208-112763691
|
|
14.
|
LM WoodZK PanP GuirnaldaP TsaiM SeaveyY
PatersonTargeting tumor vasculature with novel
Listeria-based vaccines directed against CD105Cancer Immunol
Immunother60931942201110.1007/s00262-011-1002-x21431419
|
|
15.
|
A WallechaPC MaciagS RiveraY PatersonV
ShahabiConstruction and characterization of an attenuated
Listeria monocytogenes strain for clinical use in cancer
immunotherapyClin Vaccine Immunol16961032009
|
|
16.
|
MM SeaveyZK PanPC MaciagA novel human
Her-2/neu chimeric molecule expressed by Listeria
monocytogenes can elicit potent HLA-A2 restricted CD8-positive
T cell responses and impact the growth and spread of
Her-2/neu-positive breast tumorsClin Cancer
Res15924932200919188163
|
|
17.
|
PC MaciagS RadulovicJ RothmanThe first
clinical use of a live-attenuated Listeria monocytogenes
vaccine: A Phase I safety study of Lm-LLO-E7 in patients with
advanced carcinoma of the cervixVaccine2739753983200919389451
|
|
18.
|
DA SewellZK PanY
PatersonListeria-based HPV-16 E7 vaccines limit
autochthonous tumor growth in a transgenic mouse model for HPV-16
transformed
tumorsVaccine2653155320200810.1016/j.vaccine.2008.07.036
|
|
19.
|
GR GunnA ZubairC PetersZK PanTC WuY
PatersonTwo Listeria monocytogenes vaccine vectors that
express different molecular forms of human papilloma virus-16
(HPV-16) E7 induce qualitatively different T cell immunity that
correlates with their ability to induce regression of established
tumors immortalized by HPV-16J Immunol16764716479200111714814
|
|
20.
|
A WallechaC FrenchR PetitR SinghA AminJ
RothmanLm-LLO-based immunotherapies and HPV-associated diseaseJ
Oncol2012542851201210.1155/2012/54285122481930
|
|
21.
|
T VerchZK PanY PatersonListeria
monocytogenes-based antibiotic resistance gene-free antigen
delivery system applicable to other bacterial vectors and DNA
vaccinesInfect
Immun7264186425200410.1128/IAI.72.11.6418-6425.2004
|
|
22.
|
P CossartA Toledo-AranaListeria
monocytogenes, a unique model in infection biology: an
overviewMicrobes
Infect1010411050200810.1016/j.micinf.2008.07.043
|
|
23.
|
EG PamerImmune responses to Listeria
monocytogenesNat Rev Immunol4812823200410.1038/nri1461
|
|
24.
|
D SchlüterE DomannC
BuckPhosphatidylcholine-specific phospholipase C from Listeria
monocytogenes is an important virulence factor in murine
cerebral listeriosisInfect Immun66593059381998
|
|
25.
|
H AngelakopoulosK LoockDM SisulER JensenJF
MillerEL HohmannSafety and shedding of an attenuated strain of
Listeria monocytogenes with a deletion of actA/plcB in adult
volunteers: a dose escalation study of oral inoculationInfect
Immun7035923601200212065500
|
|
26.
|
DG BrockstedtTW DubenskyPromises and
challenges for the development of Listeria
monocytogenes-based immunotherapiesExpert Rev
Vaccines710691084200810.1586/14760584.7.7.106918767955
|
|
27.
|
IR MonkCG GahanC HillTools for functional
post-genomic analysis of Listeria monocytogenesAppl Environ
Microbiol7439213934200810.1128/AEM.00314-0818441118
|
|
28.
|
Y YinD TianH JiaoPathogenicity and
immunogenicity of a mutant strain of Listeria monocytogenes
in the chicken infection modelClin Vaccine
Immunol18500505201110.1128/CVI.00445-1021228136
|
|
29.
|
X JiaoR Lo-ManN WinterE DériaudB GicquelC
LeclercThe shift of Th1 to Th2 immunodominance associated with the
chronicity of Mycobacterium bovis bacille Calmette- Guérin
infection does not affect the memory responseJ
Immunol17013921398200310.4049/jimmunol.170.3.139212538699
|
|
30.
|
P BerraondoC NouzéX PrévilleD LadantC
LeclercEradication of large tumors in mice by a tritherapy
targeting the innate, adaptive, and regulatory components of the
immune systemCancer
Res6788478855200710.1158/0008-5472.CAN-07-032117875726
|
|
31.
|
S NaitoAC von EschenbachR GiavazziIJ
FidlerGrowth and metastasis of tumor cells isolated from a human
renal cell carcinoma implanted into different organs of nude
miceCancer Res464109411519863731078
|
|
32.
|
K LinK DoolanCF HungTC WuPerspectives for
preventive and therapeutic HPV vaccinesJ Formos Med
Assoc109424201010.1016/S0929-6646(10)60017-420123582
|
|
33.
|
YL YinCJ ZhangDB TianZM PanXA JiaoKinetics
of mutant Listeria monocytogenes strain yzuLM1-2 in murine
infection modelVet Sci in China378508532007
|
|
34.
|
A de JongJM van der HulstGG KenterRapid
enrichment of human papillomavirus (HPV)-specific polyclonal T cell
populations for adoptive immunotherapy of cervical cancerInt J
Cancer114274282200515540211
|
|
35.
|
MD VeselyMH KershawRD SchreiberMJ
SmythNatural innate and adaptive immunity to cancerAnnu Rev
Immunol29235271201110.1146/annurev-immunol-031210-10132421219185
|
|
36.
|
YA YuS ShabahangTM
TimiryasovaVisualization of tumors and metastases in live animals
with bacteria and vaccinia virus encoding light-emitting
proteinsNat Biotechnol22313320200410.1038/nbt93714990953
|
|
37.
|
C GravekampY PatersonHarnessing
Listeria monocytogenes to target tumorsCancer Biol
Ther92572652010
|
|
38.
|
Q ZengS PengA MonieControl of
cervicovaginal HPV-16 E7-expressing tumors by the combination of
therapeutic HPV vaccination and vascular disrupting agentsHum Gene
Ther22809819201010.1089/hum.2010.07121128743
|
|
39.
|
RK SharmaRH SchabowskyAK Srivastava4-1BB
ligand as an effective multifunctional immunomodulator and antigen
delivery vehicle for the development of therapeutic cancer
vaccinesCancer Res7039453954201010.1158/0008-5472.CAN-09-4480
|
|
40.
|
S Ostrand-RosenbergAnimal models of tumor
immunity, immunotherapy and cancer vaccinesCurr Opin
Immunol16143150200410.1016/j.coi.2004.01.00315023405
|