Introduction
Pancreatic cancer, a highly lethal disease, is
always diagnosed at an advanced stage for which there is little
effective treatment. It remains the fourth most common cause of
cancer-related death in the Western world (1). Due to the aggressive nature of this
disease, most patients with pancreatic cancer present with local
invasion or distant metastasis at the time of diagnosis, and less
than 20% of patients are candidates for surgery with curative
intent (2). Metastatic pancreatic
cancer is relatively incurable. For pancreatic cancer, the overall
5 year survival rates are reported to be below 5% (3). Thus, understanding the molecular
mechanisms of pancreatic cancer progression should be helpful to
develop efficient treatments for the disease. Among new approaches,
gene therapy is definitely required to improve treatment results
(4–7).
B7-H3, a member of the B7 immunoregulatory family,
was identified in 2001 by database searches of a human dendritic
cell derived cDNA library (8).
Previous studies showed that B7-H3 protein can be expressed in
dendritic cells, and in the liver, lung, prostate as well as in
similar tumor cell lines (9–12).
However, the physiological and pathological role of B7-H3 is
largely unknown. In an early study, human B7-H3 was reported to be
a co-stimulator of T cells, promoting T cell proliferation and
cytokine production (8).
Subsequently, it was reported that in several mouse cancer models,
B7-H3 ectopic expression enhanced the induction of tumor-specific
CD8 cytotoxic T cells, which may slow tumor growth or even
completely eradicate tumors (13,14). More recently, B7-H3 was repeatedly
implicated as a potent inhibitor of T cell activity (15). Previous studies found that B7-H3
deficient mice show airway inflammation (16), experimental autoimmune
encephalitis (17) and allergic
conjunctivitis in an accelerated pattern (18). In contrast to these studies,
Steinberger et al(10)
suggested that B7-H3 has no characteristics of a co-signaling
molecule and it does not act as a regulator of immune responses.
Therefore, the biological functions of B7-H3 are still unclear.
Metastasis, the spread of cancer cells from the
primary tumor sites to distant organs, is a complex process that
involves induction of cell motility, activation of extracellular
matrix proteases, intravasation to vessels, travel via the
circulatory system, and survival and establishment of secondary
tumors in a new microenvironment (19,20). The same process occurs in
metastatic pancreatic cancer.
It has been suggested that B7-H3 is a
tumor-associated antigen that regulates important cellular
responses, such as proliferation, adhesion and metastasis,
indicating its novel role in tumor progression (21,22). In this study, we focused on B7-H3
in pancreatic cancer tissue as well as in the human pancreatic
cancer cell line Patu8988.
Materials and methods
Reagents
The anti-human B7-H3 antibody was purchased from
R&D Systems, Inc. The horseradish peroxidase-conjugated
secondary anti-mouse antibody was from Bio-Rad Laboratories, Inc.
TRIzol reagent and MMLV were purchased from Gibco-BRL. TaqDNA
polymerase, dNTPs and DNA marker were purchased from Takara.
Patients
This study was approved by the Ethics Committee of
The First Affiliated Hospital of Soochow University for Clinical
Investigation. Included in the study were 26 patients with
pancreatic cancer who underwent surgery for radical resection.
Patients were excluded from analysis if they received chemotherapy
or radiation therapy prior to the surgical operation or underwent
previous pancreatic surgery. Specimens of pancreatic cancer were
obtained from the patients during surgical operation, following
written consent. At the same time, specimens of normal pancreatic
tissues distant to the tumor were obtained as controls. The
diagnosis of each tissue was confirmed by a frozen section stained
with hematoxylin and eosin. After dissection under sterile
conditions, each tissue sample was collected, separated and divided
into 2 groups during preparation and analysis. One group was fixed
in 10% buffered methanol for immunohistochemical estimation of
B7-H3 expression, and another group was used for B7-H3
enzyme-linked immunosorbent assay (ELISA).
Tissue extracts and B7-H3 enzyme-linked
immunosorbent assay (ELISA)
Each tissue sample was collected and weighed in the
same manner during preparation. Extract preparation was performed
as recommended by the manufacturer (Cell Signaling Technology).
Briefly, each tissue sample was prepared with PBS and homogenized
in cell lysis buffer containing Protease Inhibitor Cocktail Set I
(Calbiochem). After incubation on ice for 30 min, the homogenate
was centrifuged at 14,000 x g for 10 min at 4°C, and the
supernatant was collected for ELISA assay. ELISA kits produced at
our laboratory were used to measure B7-H3, as described previously
(19). B7-H3 concentrations were
determined using a standard curve with an 8-parameter curve fit
analysis program. We calculated the B7-H3 level/gram (g) of each
tissue sample.
Cells and cell culture
Pancreatic cancer cell line Patu8988 was kindly
provided by Professor Chang-Geng Ruan from the Jiangsu Provincial
Institute of Hematology, China. Patu8988 cells were cultured in
RPMI-1640 (Gibco Inc.), and the medium was supplemented with 10%
fetal bovine serum (Atlanta Biologicals, Inc.) and 1%
penicillin-streptomycin (Gibco, Inc.) at 37°C in an atmosphere of
5% CO2. After cells attained 80 to 90% confluence, they
were harvested with 0.25% trypsin and split at a 1:3 ratio.
Generation of stable cell lines
Small hairpin RNA (shRNA) of the human B7-H3
(NM_001024736; GenBank) lentiviral gene transfer vector encoding
the green fluorescent protein (GFP) sequence was constructed by
Shanghai GeneChem Co. (Shanghai, China). The targeting sequence of
B7-H3 was 5′-GAGCAGGGCTTGTTTGATGTG-3′, and it was confirmed by
sequencing. The recombinant lentivirus of small hairpin
interference RNA targeting B7-H3 (LV-B7-H3 virus) and the
nontargeted control mock lentivirus (LV-NC virus) were prepared and
titered to 5×109 Tu/ml (transfection unit). Cells were
subcultured at 5×104 cells/well into 6-well tissue
culture plates overnight. The viral supernatant was then added into
cells at a multiplicity of infection (MOI) of 10 with ENi.S and 5
μg/ml Polybrene. GFP was evaluated by fluorescence microscopy to
estimate the infection efficiency. The infected Patu8988 cells were
termed the LV-B7-H3 group and LV-NC group, respectively, and the
Patu8988 cells without infection were the control group. The 3
groups mentioned above were used in subsequent experiments.
Real-time reverse transcriptase-polymerase chain reaction (RT-PCR)
was carried out to confirm the knockdown of B7-H3 mRNA, and B7-H3
protein expression was analyzed by FCM using a Cytomics™ FC 500
device.
Real-time reverse
transcriptase-polymerase chain reaction (RT-PCR)
RT-PCR was performed to confirm the knockdown of
B7-H3 mRNA in the transfectants. Total RNA was collected using
TRIzol reagent following the manufacturer’s instructions. The
concentration and purity of the total RNA were detected with an
ultraviolet spectrophotometer and then reversely transcribed into
cDNA with MMLV. Quantitative real-time PCR assays were carried out
using SYBR-Green Real-time PCR Master Mix and real-time PCR
amplification equipment. GAPDH was used as an internal control. The
PCR conditions consisted of 1 cycle at 95°C for 15 sec followed by
45 cycles at 95°C for 5 sec and at 60°C for 30 sec. The primer
sequences were as follows: 5′-CTCTGCCTTCTCACCTCTTTG-3′ (sense) and
5′-CCTTGAGGGAGGAACTTTATC-3′ (antisense) for B7-H3 (134 bp);
5′-TGACTTCAACAGCGACACCCA-3′ (sense) and 5′-CACCCTGTTGCTGTAGCCAAA-3′
(antisense) for GAPDH (121 bp).
In another experiment of RT-PCR for B7-H3, products
were electrophoresed on 1.8% agarose gel containing 0.1% ethidium
bromide. Images of the fluorescent bands were captured by use of
the Bio-Rad gel documentation system.
Cell proliferation by MTT assay
The MTT assay was used to study the effect of B7-H3
RNA interference on Patu8988 cell proliferation. Cells of each
group were plated at 10,000 cells/ well in a 96-well plate for 24,
48 or 72 h. At each time point after discarding the medium, 100 μl
RPMI-1640 containing 20 μl MTT (Sigma) (5 mg/ml) was added to each
well. After incubation at 37°C for 4 h the MTT solution was
removed. Dimethyl sulfoxide (100 μl) was added to each well and
mixed to dissolve the dark blue formazan crystals that formed. The
proportion of viable cells was determined by reading the optical
density using test wave length (570 nm) and reference wave length
(630 nm) with a Multiskan™ MK3 ELISA reader. The assay was carried
out in quintuplicate for each group and repeated in triplicate.
In vitro wound scrape assay
Cells of each group were incubated in 6-well plates.
A small wound area was made in the confluent monolayer with a
200-μl pipette tip in a lengthwise stripe. Cells were then washed
twice with PBS and incubated in serum-free RPMI-1640 medium at 37°C
in a 5% CO2 incubator for 24 h (23,24). Images were captured at different
times from 0 to 48 h. Wound width was measured at a ×100
magnification using a BX50 microscope (Olympus) with a calibrated
eyepiece grid (1 mm/100 μm graduation). Ten measurements were
determined at random intervals along the wound length. This
experiment was carried out in triplicate.
In vitro invasion assay
A co-culture system was used as an alternative
method to evaluate cancer cell invasiveness (25). Briefly, the upper portion of
Transwell inserts with an 8-μm pore size and a 6.5-mm diameter was
coated with 20 μl Matrigel diluted 1:3 in serum-free RPMI-1640 and
incubated at 37°C for 4 h. The coated inserts were placed in the
well of a 24-well plate with 600 μl RPMI-1640 containing 10% FBS in
the bottom chamber. After 12 h of serum starvation, the trypsinized
cells were harvested and diluted to a 5×106/ml cell
suspension with serum-free RPMI-1640. Each cell suspension (100 μl)
was added to the upper chambers. After incubation at 37°C for 48 h
in a 5% CO2 atmosphere, the non-invading cells and gel
were removed from the upper chamber with cotton tipped swabs. The
cells were rinsed with PBS, and cells on the filters were fixed
with methanol for 30 min and stained with crystal violet solution
(Sigma). The number of invading cells on the filters was counted in
5 random fields/filter at ×100 magnification in triplicate wells of
each group.
Subcutaneous transplantation model
study
Three groups of 6 male Balb/c nude mice (5- to
6-weeks old and 20–24 g in weight) were bred in an
aseptic-specified pathogen-free (SPF) condition and kept at a
constant humidity and temperature (25–28°C). Animal experiments
were carried out according to protocols approved by the Animal Care
and Use Committee and were in compliance with the Guidelines on
Animal Welfare of the China National Committee for Animal
Experiments. Cells (2×107) (LV-B7-H3, LV-NC or control
cells) in 0.2 ml normal sodium were injected subcutaneously in the
right inguinal region of nude mice, respectively. The size of
tumors was measured twice a week with calipers, and the volume was
determined using the simplified formula of a rotational ellipsoid
(L x W2 × 0.5). Growth curves were constructed, and the
data are presented as means ± SD. Tumors were harvested from mice 6
weeks after tumor cell injection. B7-H3 expession was detected by
immunohistochemistry of the tumor xenografts.
Immunohistochemistry
Clinical specimens and the tumor xenografts were
used for immunohistochemical studies. Specimens were fixed in
formalin overnight and embedded in paraffin. Serial sections (4 μm)
were prepared for immunohistological staining. Tissue sections were
quenched for endogenous peroxidase with freshly prepared 3%
H2O2 with 0.1% sodium azide and then placed
in an antigen retrieval solution for 15 min. After incubation in a
casein block, primary antibodies such as anti-B7-H3 (1:50 dilution)
were applied to the sections for 1 h at room temperature, followed
by incubation with the secondary antibody and Extravidinconjugated
horseradish peroxidase. The immune reaction was counterstained with
hematoxylin, dehydrated, and mounted. Sections were then evaluated
for the presence of brown diaminobenzidine precipitates indicative
of positive reactivity by microscopy. The brown staining in the
cytoplasm was read as positive reactivity for B7-H3.
Orthotopic transplantation pancreatic
cancer model study
The establishment of an inguinal region subcutaneous
transplantation tumor model of 3 groups (LV-B7-H3, LV-NC and
control cells) was carried out respectively, as described above.
After growing to a specific certain volume, the tumors were
resected under aseptic environment and washed twice in
antibiotic-containing RPMI-1640 to prevent possible infection.
Necrotic tissues were removed, and the remaining viable tumor
tissues were cut into small pieces of 1 mm3.
Five-week-old BALB/c-nu mice, weighing 20–24 g, were anesthetized
with urethane (4 ml/kg) by intramuscular injection. After the
abdominal skin was sterilized, an incision was made in the upper
left abdomen, and the pancreas was exposed. Tumor pieces were
attached to the pancreas using absorbable sutures. The pancreas was
then returned to the peritoneum, and the abdominal wall and the
skin were closed with silk sutures. The animals were allowed to
recover for 24 h. Three groups of 6 surviving mice were bred in an
aseptic-specified pathogens-free (SPF) condition and kept at a
constant humidity and temperature (25–28°C). All of the mice were
sacrificed 7 weeks after the orthotopic transplantation operation.
Metastatic visceral tumors out of the pancreas, such as metastatic
tumors in liver, on the small intestine serous membrane surface or
on the peritoneum, were excised carefully and weighed as described
previously (26).
Statistical analysis
B7-H3 expression in pancreatic cancer and normal
pancreas tissues as determined by immunohistochemical staining was
compared and assessed using the Chi-square test. Other data are
shown as means ± SD. Statistical comparisons were performed using
the Student’s t-test. All P-values were determined by 2-sided tests
with significance considered at <0.05. These analyses were
performed using SPSS 13.0 software.
Results
Tissue samples and immunohistochemical
staining
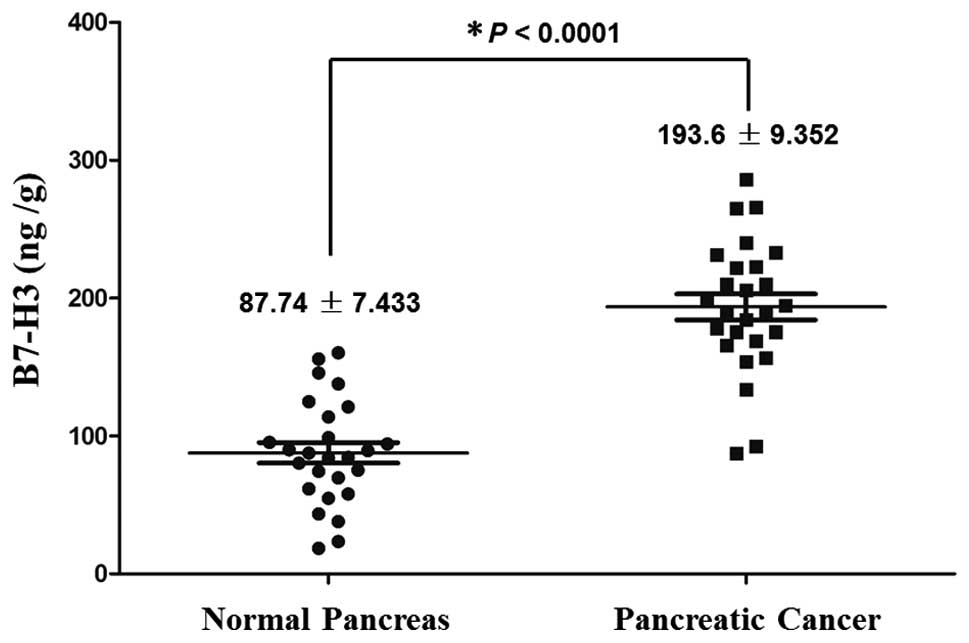
B7-H3 levels in the pancreatic cancer group were
significantly higher than that in the normal pancreas group (mean
193.6±9.352 vs. 87.74±7.433 ng/g, P<0.0001) (Fig. 1). Immunohistochemical staining
revealed significantly overexpressed B7-H3 in tumor tissue
(Chi-square test 15.341; P<0.001). Positive staining for B7-H3
expression was detected in more than 50% of cells in 17 of the 26
pancreatic cancer specimens while no positive cells were detected
in normal pancreas specimens (Fig.
2).
B7-H3 downregulation by RNA interference
in Patu8988 cells
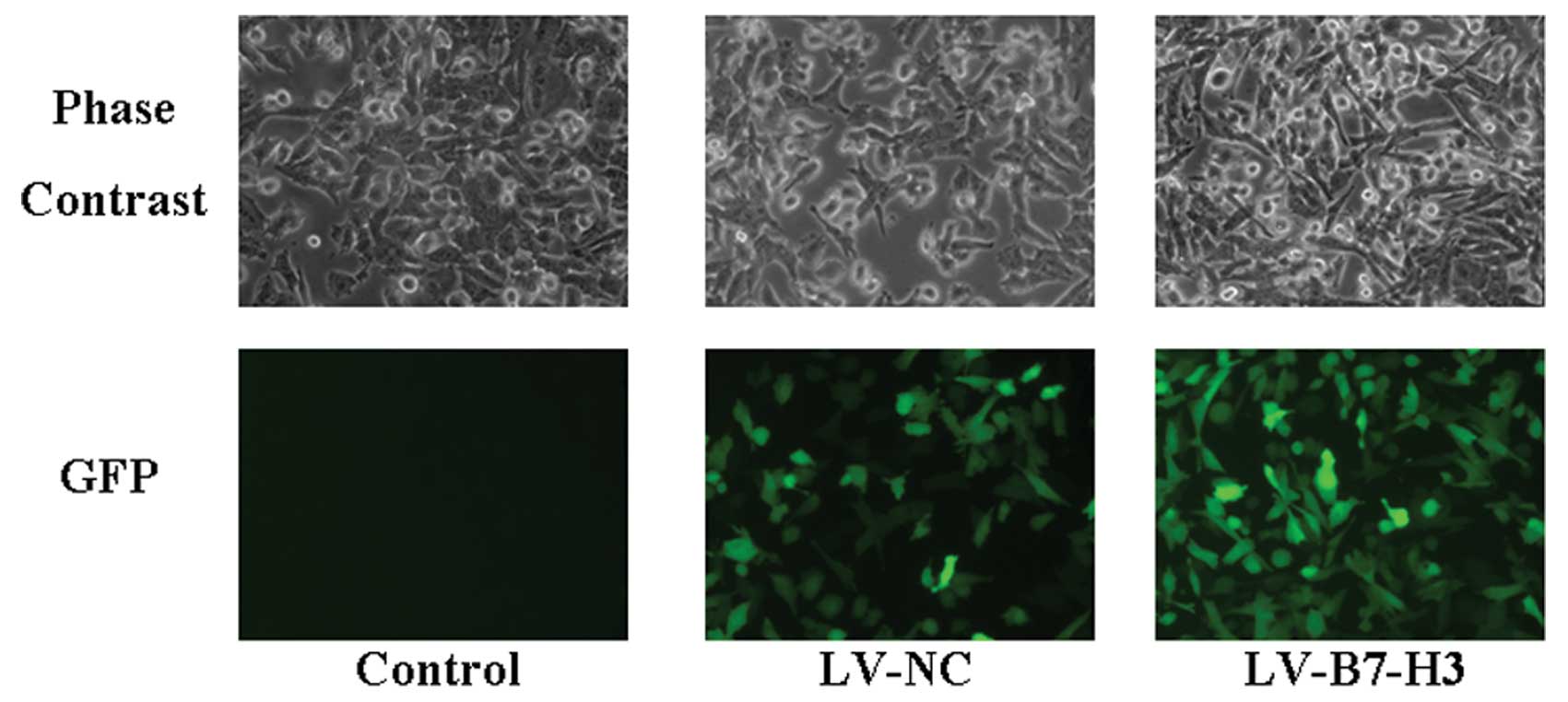
After infection with the lentiviral vector, Patu8988
cells were examined by fluorescence microscopy (Fig. 3). The result showed high
efficiency of the lentiviral infection. To determine the efficiency
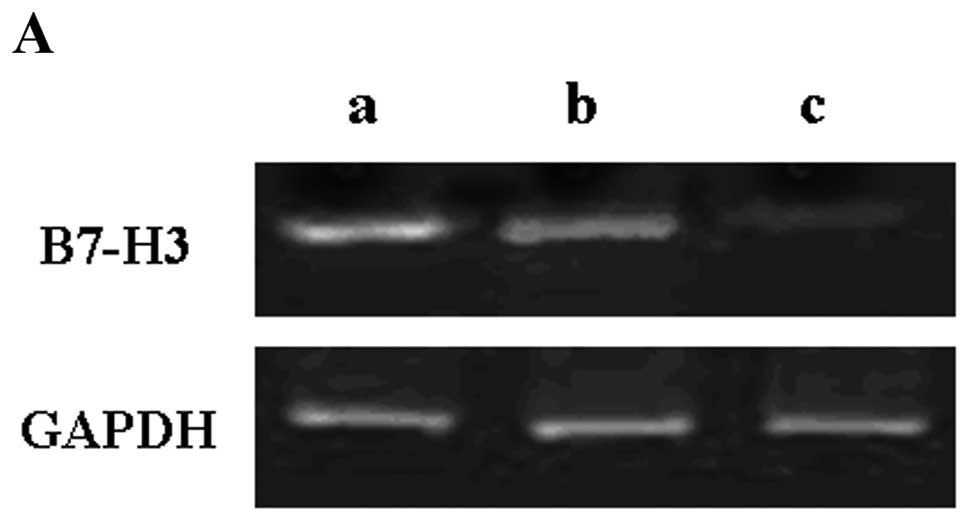
of RNA interference, we analyzed the levels of B7-H3 mRNA and
protein expression in the 3 groups. Fig. 4A and B shows B7-H3 mRNA expression
in the 3 groups. B7-H3 mRNA expression was obviously decreased in
the LV-B7-H3 group compared with the LV-NC or the control groups
(mean 8.4±2.15%, P<0.01) (Fig.
4B). The inhibition rate was 91.6%. However, there was no
significant difference between the LV-NC group and the control
group (P>0.05). A similar decrease was found in protein
synthesis by FCM assay (mean 20.6±5.9%, P<0.01) (Fig. 4C). The mean inhibition rate was
79.4% vs. the control group. These findings indicate that the
downregulation of the B7-H3 gene, by RNA interference was specific
and efficient.
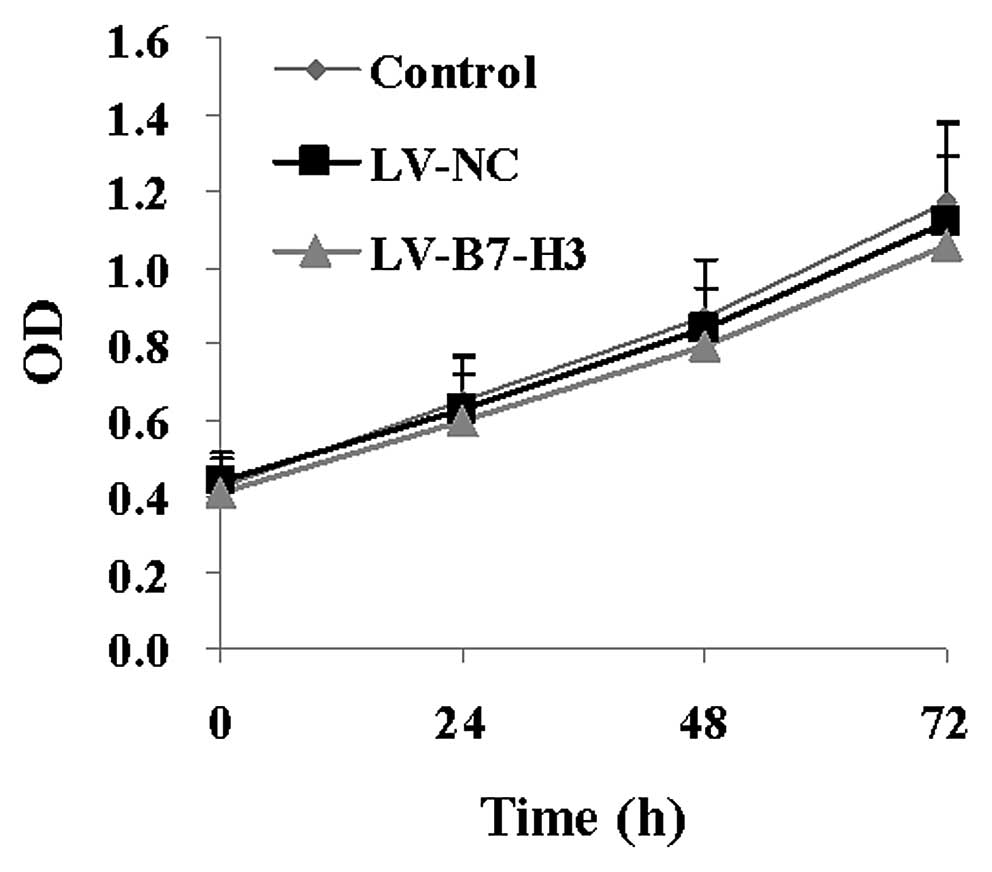
Proliferation in vitro by MTT assay
To characterize the role of B7-H3 in Patu8988 cell
growth, we measured the cell proliferation rate in vitro by
MTT assay. There was no statistical significance in cellular
proliferation between the control and the experimental groups
(P>0.05) (Fig. 5).
Migration on wound scrape assay in
vitro
To determine whether B7-H3 acts as a cell migration
regulator, we used the wound scrape assay to evaluate cell
motility. RNA interference resulting in inhibition of B7-H3
significantly decreased Patu8988 cell migration in the wound scrape
model (Fig. 6). Time course
analysis of the wound closure showed that a monolayer was
re-established within a significantly shorter period in the LV-NC
and control groups than that in the LV-B7-H3 group.
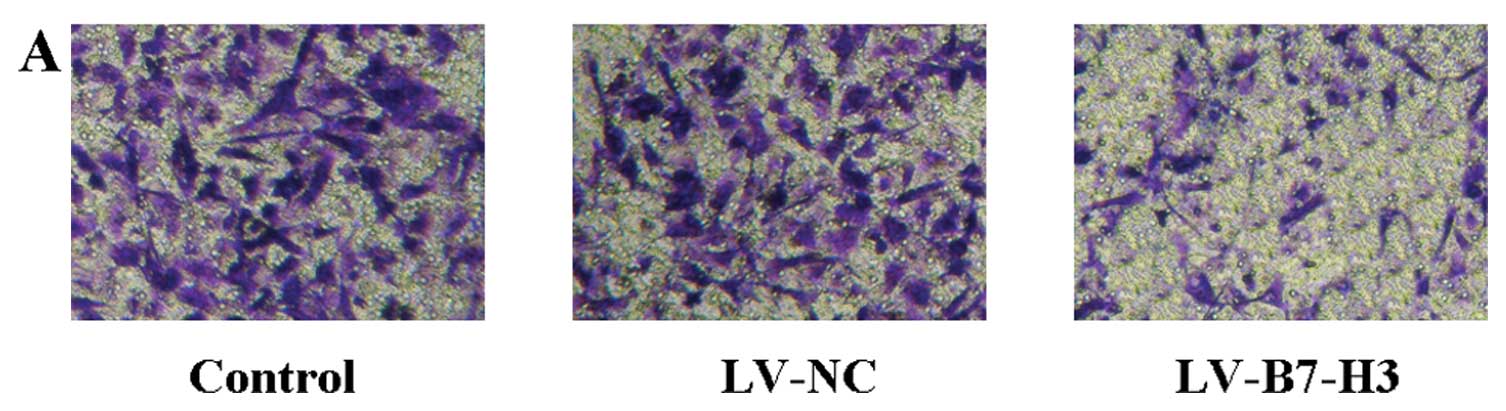
Invasive ability in the Transwell assay
in vitro
After down-regulation of the expression of B7-H3 by
RNA interference, an in vitro assay on Matrigel filters
revealing that the number of invading Patu8988 cells was decreased
up to 50% (P<0.05, LV-B7-H3 group vs. the control group)
(Fig. 7). There was no
statistical significance in the number of invading cells between
the LV-NC and the control group (P>0.05).
Tumor growth in the subcutaneous
transplantation mouse model
The in vitro experiments with the Patu8988
cells showed the effects of B7-H3 on tumor progression. Hence, we
examined whether this could be observed in vivo. LV-B7-H3,
LV-NC and control cells were injected subcutaneously into nude
mice. All of the 18 mice developed detectable tumors at the
beginning of this experiment. The growth rate was reduced by the
knockdown of B7-H3 (Fig. 8).
Inhibition of tumor growth was observed in the LV-B7-H3 group at 6
weeks, when compared to the LV-NC group (211±47 mm3) or
the control group (235±57 mm3). The average tumor volume
(22±5 mm3) in the LV-B7-H3 group was significantly lower
than that in the LV-NC and control groups (P<0.01). There was no
statistical significance in tumor volume between the LV-NC and the
control group (P>0.05).
The knockdown of B7-H3 in the xenografts was
confirmed by immunohistochemical staining. While the level of B7-H3
expression retaimed low in the LV-B7-H3 group tumors, the LV-NC and
control tumors showed strong staining (Fig. 9).
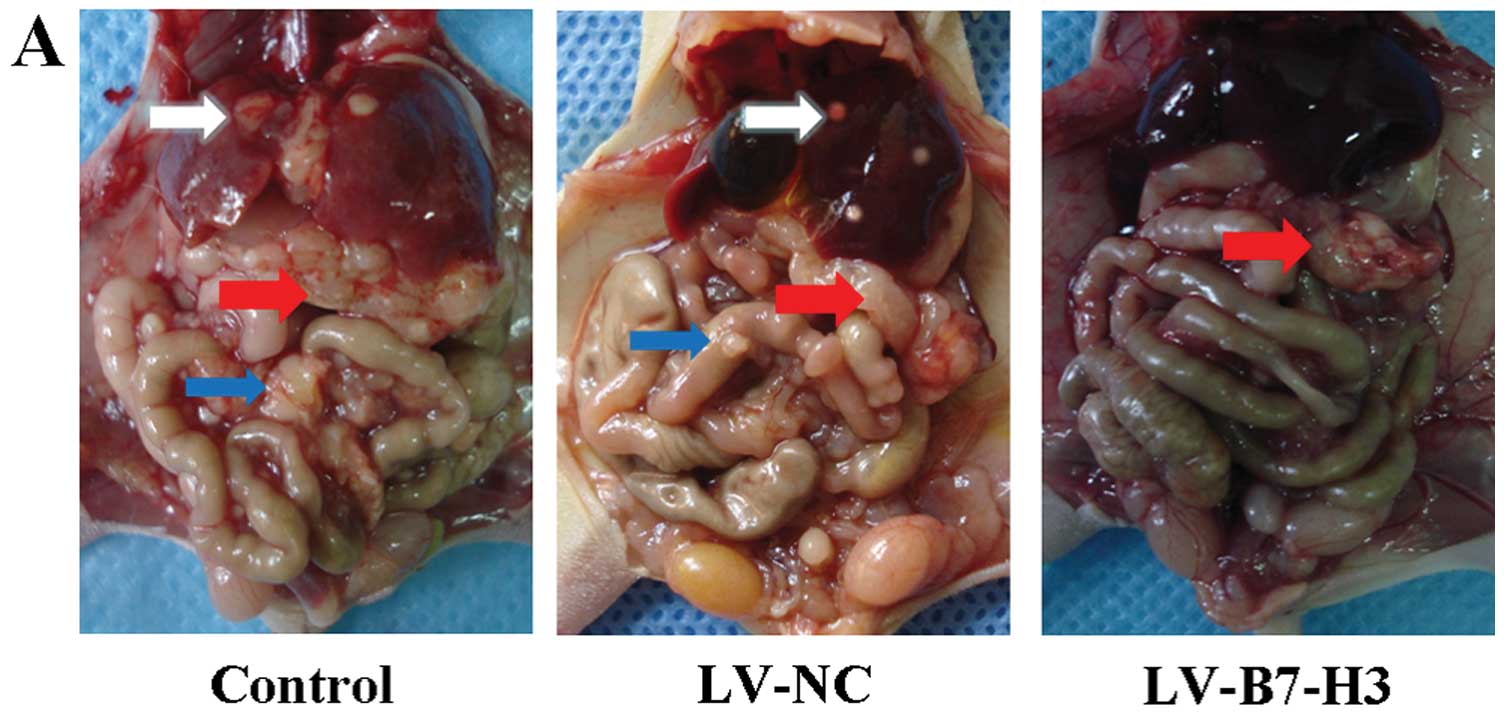
Metastatic tumors in the orthotopic
transplantation pancreatic cancer mouse model
All of the 18 mice were sacrificed 7 weeks after the
transplantation operation. All of the mice developed orthotopic
transplantation pancreatic cancer tumors in this experiment.
Abdominal visceral metastatic tumors were detected, excised and
weighed (Fig. 10). The number of
cases of liver metastasis in the LV-B7-H3 group (1/6, 16.67%) was
less than the number of cases in the LV-NC group (4/6, 66.67%) or
the control group (5/6, 83.33%). The effect of inhibiting
metastasis by knockdown of B7-H3 was assessed in terms of the
average postmortem abdominal visceral metastatic tumor weight.
Inhibition of metastasis was observed in the LV-B7-H3 group, when
compared to the LV-NC group (1.28±0.41 g) or the control group
(1.33±0.38 g). The average weight of the abdominal visceral
metastatic tumors (0.26±0.13 g) in the LV-B7-H3 group was
significantly lower than that in LV-NC and control groups (Fig. 10B) (P<0.01). There was no
statistical significance in metastatic visceral tumor weight
between the LV-NC and the control group (P>0.05). These
indicated that inhibition of B7-H3 expression reduced pancreatic
cancer metastasis in vivo. It strongly supports the effects
observed in vitro indicating that B7-H3 plays a vital role
in invasion and migration of pancreatic cancer cells.
Discussion
In recent years increasing evidence indicates that
B7-H3 plays an important role in tumor progression and metastasis.
Wu et al(27) reported
that B7-H3 expression is related to survival time and tumor
infiltration depth in gastric cancer cases. Zhang et
al(28) found that
circulating B7-H3 in serum is a highly sensitive biomarker for
non-small cell lung cancer (NSCLC) and increased circulating B7-H3
suggests a poor clinical prognosis for NSCLC patients. Sun et
al(29) reported that higher
B7-H3 expression in colorectal cancer was positively correlated
with a more advanced tumor grade, and the level of soluble B7-H3 in
serum from colorectal cancer patients was higher than healthy
donors. This suggests that both soluble and membranous B7-H3
proteins are involved in colorectal cancer progression. Yamato
et al(12) found that
B7-H3 expression was significantly more intense in cases with lymph
node metastasis and advanced pathological stage in pancreatic
cancer. B7-H3 blockade induced a substantial antitumor effect on
murine pancreatic cancer. B7-H3 overexpression was also reported to
correlate with tumor aggressiveness and poor clinical outcome,
suggesting that B7-H3 has a critical role in tumor progression.
Since we realized the limitation of using
immunohistochemical methods for semi-quantitative analysis
(30), we also used ELISA. Our
results showed aberrant B7-H3 expression in pancreatic cancer, in
accord with the findings of Yamato et al(12). However, to ascertain why B7-H3
overexpression correlates with pathological indicators of
aggressive cancer and clinical outcome and its role in tumor
progression, we further investigated the effects of low B7-H3
expression on the biological features of human pancreatic cancer
Patu8988 cells.
Carcinogenesis is a multiple step process in which
cancer cells lose proliferation control, disseminate from a
localized primary tumor mass to invading adnexa and metastasize to
distant organs. Limitless cell growth is an important alteration in
cancer cell phenotype (31). We
first determined the effects of B7-H3 depletion on Patu8988 cell
growth. The proliferation of B7-H3-knockdown cells was the same as
that of controls and there was no obvious difference. To further
investigate whether B7-H3 contributes to tumor metastasis, we
performed a wound scrape assay to evaluate cell motility and a
Transwell invasion assay to assess cell invasiveness in
vitro to determine the mechanisms of cell metastasis toward
distant tissue. The results indicated that B7-H3 has a putatively
important role in tumor migration and invasiveness, indicating
higher aggressiveness and poor clinical outcome.
The results in vitro were confirmed in our
studies in vivo. In the subcutaneous transplantation model,
the growth rate of established B7-H3-knockdown xenografts was
slower than that of the LV-NC and the control groups. Although the
MTT assay in vitro showed that the proliferation rate was
not decreased by B7-H3 knockdown, the growth rate of
B7-H3-knockdown tumors in the subcutaneous transplantation model
showed a significant decrease. Nevertheless, the tumor cell growth
microenvironment in vivo is quite more complicated than that
in vitro. The underlying molecular mechanisms of this
phenomenon still require further research. Immunohistochemical
analysis of the xenograft tissue confirmed that the tumors
originating from LV-B7-H3 cells retained low expression levels of
the B7-H3 protein, whereas the LV-NC group and the control group
tumors showed strong B7-H3 staining. Furthermore, in the orthotopic
transplantation pancreatic cancer model, we found that decreased
B7-H3 expression reduced tumor metastasis. Compared to the control
group, there was a dramatic reduction in the weight of the
abdominal visceral metastatic tumors in the LV-B7-H3 group. The
underlying reasons may at least partly be that B7-H3 knockdown
inhibits pancreatic cancer migratory and invasive ability.
In summary, our study investigating the role of
B7-H3 in pancreatic cancer progression shows that this protein
promotes cancer cell migration and invasiveness in vitro and
in vivo. Furthermore, in contrast to previous reports
focusing on the immunoregulatory effects of B7-H3, which are
involved in evasion of cancer immune surveillance, our data show
that it plays a critical role in pancreatic cancer progression
through cell migration and invasiveness via non-immunomechanisms.
These findings provide new insight into the role of B7-H3 in
pancreatic cancer and may have important implications in the
development of targeted therapeutics for this disease. However,
whether B7-H3 regulates cancer progression directly or through
various important intracellular pathways, still requires
investigation, and we will engage in this field further.
In conclusion, B7-H3 is aberrantly expressed in
pancreatic cancer. Our study indicates that B7-H3 may regulate
tumor progression by promoting cell migration and invasiveness, in
addition to acting as an immunoregulatory protein. B7-H3 may serve
as a potential molecular target for pancreatic cancer therapy.
Although preliminary data are significant, a precise mechanism of
B7-H3 expression regulation in the tumor environment, overall
knowledge of its clinical implications and targeted therapeutic
interventions in pancreatic cancer require further
investigation.
Acknowledgements
This study was supported by grants
from the National Natural Science Foundation of China (no.
30901789), and the Post-graduate Scientific Research Innovation
Project of the Education Department of Jiangsu Province (no.
XZZ11_0125), and the Science and Technology Research Project of
Science and Technology Bureau of Suzhou City (no. YS201120),
China.
References
|
1
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics, 2007. CA Cancer J Clin. 57:43–66. 2007. View Article : Google Scholar
|
|
2
|
Neoptolemos JP, Cunningham D, Friess H, et
al: Adjuvant therapy in pancreatic cancer: historical and current
perspectives. Ann Oncol. 14:675–692. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar
|
|
4
|
Ghaneh P, Costello E, Neoptolemos JP, et
al: Biology and management of pancreatic cancer. Postgrad Med J.
84:478–497. 2008. View Article : Google Scholar
|
|
5
|
Pan X, Sheng W, Zhu Q, et al: Inhibition
of pancreatic carcinoma growth by adenovirus-mediated human
interleukin-24 expression in animal model. Cancer Biother
Radiopharm. 23:425–434. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou J, Zhang ZX, Zhao H, et al:
Anti-angiogenesis by lentivirus-mediated small interfering RNA
silencing of angiopoietin-2 gene in pancreatic carcinoma. Technol
Cancer Res Treat. 10:361–369. 2011.PubMed/NCBI
|
|
7
|
Zhao X, Li DC, Zhao H, et al: A study of
the suppressive effect on human pancreatic adenocarcinoma cell
proliferation and angiogenesis by stable plasmid-based siRNA
silencing of c-Src gene expression. Oncol Rep. 27:628–636.
2012.PubMed/NCBI
|
|
8
|
Chapoval AI, Ni J, Lau JS, et al: B7-H3: a
costimulatory molecule for T cell activation and IFN-gamma
production. Nat Immunol. 2:269–274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Suh WK, Wang SX, Jheon AH, et al: The
immune regulatory protein B7-H3 promotes osteoblast differentiation
and bone mineralization. Proc Natl Acad Sci USA. 101:12969–12973.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Steinberger P, Majdic O, Derdak SV, et al:
Molecular characterization of human 4Ig-B7-H3, a member of the B7
family with four Ig-like domains. J Immunol. 172:2352–2359. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang GB, Zhou H, Chen YJ, et al:
Characterization and application of two novel monoclonal antibodies
against 2IgB7-H3: expression analysis of 2IgB7-H3 on dendritic
cells and tumor cells. Tissue Antigens. 66:83–92. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamato I, Sho M, Nomi T, et al: Clinical
importance of B7-H3 expression in human pancreatic cancer. Br J
Cancer. 101:1709–1716. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Luo L, Chapoval AI, Flies DB, et al: B7-H3
enhances tumor immunity in vivo by costimulating rapid clonal
expansion of antigen-specific CD8+ cytolytic T cells. J
Immunol. 173:5445–5450. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lupu CM, Eisenbach C, Kuefner MA, et al:
An orthotopic colon cancer model for studying the B7-H3 antitumor
effect in vivo. J Gastrointest Surg. 10:635–645. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Castriconi R, Dondero A, Augugliaro R, et
al: Identification of 4Ig-B7-H3 as a neuroblastoma associated
molecule that exerts a protective role from an NK cell-mediated
lysis. Proc Natl Acad Sci USA. 101:12640–12645. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suh WK, Gajewska BU, Okada H, et al: The
B7 family member B7-H3 preferentially down-regulates T helper type
1-mediated immune responses. Nat Immunol. 4:899–906. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Prasad DV, Nguyen T, Li Z, et al: Murine
B7-H3 is a negative regulator of T cells. J Immunol. 173:2500–2506.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fukushima A, Sumi T, Fukuda K, et al:
B7-H3 regulates the development of experimental allergic
conjunctivitis in mice. Immunol Lett. 113:52–57. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ghadially R: The role of stem and
circulating cells in cancer metastasis. J Surg Oncol. 103:555–557.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen YW, Tekle C and Fodstad O: The
immunoregulatory protein human B7-H3 is a tumor-associated antigen
that regulates tumor cell migration and invasion. Curr Cancer Drug
Targets. 8:404–413. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang G, Hou J, Shi J, et al: Soluble
CD276 (B7-H3) is released from monocytes, dendritic cells and
activated T cells and is detectable in normal human serum.
Immunology. 123:538–546. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsai CY, Lee TS, Kou YR, et al:
Glucosamine inhibits IL-1 beta-mediated IL-8 production in prostate
cancer cells by MAPK attenuation. J Cell Biochem. 108:489–498.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fernandez-Martinez AB, Bajo AM,
Sanchez-Chapado M, et al: Vasoactive intestinal peptide behaves as
a pro-metastatic factor in human prostate cancer cells. Prostate.
69:774–786. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yaqinuddin A, Qureshi SA, Qazi R, et al:
DNMT1 silencing affects locus specific DNA methylation and
increases prostate cancer derived PC3 cell invasiveness. J Urol.
182:756–761. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
ElBayoumi TA and Torchilin VP:
Tumor-targeted nano-medicines: enhanced antitumor efficacy in vivo
of doxorubicin-loaded, long-circulating liposomes modified with
cancer-specific monoclonal antibody. Clin Cancer Res. 15:1973–1980.
2009. View Article : Google Scholar
|
|
27
|
Wu CP, Jiang JT, Tan M, et al:
Relationship between co-stimulatory molecule B7-H3 expression and
gastric carcinoma histology and prognosis. World J Gastroenterol.
12:457–459. 2006.PubMed/NCBI
|
|
28
|
Zhang G, Xu Y, Lu X, et al: Diagnosis
value of serum B7-H3 expression in non-small cell lung cancer. Lung
Cancer. 66:245–249. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun J, Chen LJ, Zhang GB, et al: Clinical
significance and regulation of the costimulatory molecule B7-H3 in
human colorectal carcinoma. Cancer Immunol Immunother.
59:1163–1171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou GX, Ireland J, Rayman P, et al:
Quantification of carbonic anhydrase IX expression in serum and
tissue of renal cell carcinoma patients using enzyme-linked
immunosorbent assay: prognostic and diagnostic potentials. Urology.
75:257–261. 2010. View Article : Google Scholar
|
|
31
|
Yang QS, Gu JL, Du LQ, et al:
ShRNA-mediated Ku80 gene silencing inhibits cell proliferation and
sensitizes to gamma-radiation and mitomycin C-induced apoptosis in
esophageal squamous cell carcinoma lines. J Radiat Res. 49:399–407.
2008. View Article : Google Scholar : PubMed/NCBI
|