Introduction
Lung cancer has emerged as the leading cause of
cancer-related mortalities, with an increasing incidence worldwide.
The long-term survival of lung cancer patients treated with
conventional therapies, including surgery, radiation therapy and
chemotherapy, remains poor and has changed little in decades.
Therefore, a more rational approach to lung cancer therapy is
required.
Osteopontin (OPN) is an acidic, glycosylated,
phosphorylated and secreted protein that is crucial in determining
the aggressiveness and oncogenic potential of several types of
cancer, including lung cancer (1,2).
It interacts with specific receptors that affect different cascades
of the signaling pathway and modulate the expression of several
downstream molecules associated with cancer development. Thus,
investigators have attempted to inhibit cancer progression by
targeting the OPN function (3,4).
The OPN function mainly depends on post-translational modification
(PTM) and regulation of the processes that involve OPN may be
mediated through glycosylation and phosphorylation (5,6).
O-glycosylation is considered a highly complex process in which
carbohydrate chains are created and extended through the sequential
addition of sugars in a Golgi apparatus wherein transferases are
located (7,8).
Five O-glycosylation sites have been identified in
humans, three of which are also conserved in a mouse OPN (9–11).
However, no reports have indicated the importance of the
O-glycosylation sites in OPN, the function of OPN and in regulating
its downstream signaling pathway.
OPN induces NF-κB-mediated signaling pathways by
binding to its receptor. NF-κB is constitutively active in most
tumor cells and its suppression inhibits the growth and migration
of tumor cells (12–14). Translation plays an important role
in controlling cell growth and it can be regulated via the
mammalian target of rapamycin (mTOR) signaling pathway through
p70S6K, of which OPN-activated Akt is a critical intermediate.
Translation initiation has been shown to be a common downstream
target of signal transduction pathways deregulated in several types
of cancer, including lung cancer (15–17). However, the mechanism by which OPN
regulates mTOR/p70S6K and NF-κB signaling pathways as well as how
inhibition of these pathways may alter cell migration and growth in
lung cancer cells remain to be clarified.
In the present study, it was hypothesized that the
specific inhibition of OPN downstream signaling by introducing
triple mutant (TM) OPN may alter lung cancer cell growth and
motility. Therefore, wild-type (WT) OPN and OPN TM were introduced
to lung cancer cells and their effects on cap-dependent protein
translation, NF-κB activity and glucose uptake were examined. The
effects of OPN WT and TM on lung cancer cell growth and migration
both in vitro and in a xenograft mouse model were also
investigated.
The present study demonstrated that unlike OPN WT
expression, OPN TM expression did not increase cap-dependent
protein translation, NF-κB activity and glucose uptake. The study
results also showed that OPN TM decreased lung cancer cell growth
and migration, suggesting that targeting OPN by introducing OPN TM
is a promising strategy for lung cancer therapy.
Materials and methods
Cell culture and materials
H226 and H322 lung cancer cells were purchased from
the American Type Culture Collection (CCL-185; Rockville, MD, USA).
The cells were grown in RPMI-1640 supplemented with 10% FBS and 1%
penicillin/streptomycin from Gibco BRL (Grand Island, NY, USA). The
mTOR and phospho-mTOR antibodies were obtained from Cell Signaling
Technology, Inc. (Beverly, MA, USA). The other indicated antibodies
were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). LY294002 and monensin were purchased from Calbiochem (San
Diego, CA, USA); PDTC and rapamycin from Sigma- Aldrich (St. Louis,
MO, USA); and NBD-C6-ceramide from Molecular Probes (Eugene, OR,
USA). The bicistronic construct, pcDNA-fLUC-polIRES-rLUC, in which
firefly luciferase represents cap-dependent and the renilla
luciferase activity represents cap-independent protein translation,
was provided by Dr Gram (Novartis Pharma AG, Basel, Switzerland).
The NF-κB luciferase reporter vector was a gift from Dr Nancy
Colburn (National Cancer Institute, Frederick, MD, USA). The
transfection reagent LT1 was obtained from Mirus Biotechnology
(Madison, WI, USA). The Luciferase assay kit and the pRA-SV40
control vector were purchased from Promega (Madison, WI, USA). The
lentivirus that contained shRNA targeting p70S6K (shp70S6K), the
BLOCK-iT™ Lentiviral RNAi Expression System (Invitrogen, Grand
Island, NY, USA), was generated according to the manufacturer’s
instructions, after which the virus titer was determined using an
HIV-1 p24 ELISA kit from PerkinElmer Life Sciences (Boston, MA,
USA).
Plasmid constructs and generation of a
stable cell line
The human OPN was generated via reverse
transcription-PCR (RT-PCR) using the total RNA. Primers used for
RT-PCR were: a forward primer that contained an XbaI
restriction site (5′-GCTCTAGAATGAGAATTGCAGTGATTTG-3′) and a reverse
primer that contained a KpnI restriction site (5′-CGG
GGTACCTTAATTGACCTCAGAAGATGC-3′). The PCR product was inserted into
the corresponding sites of pcDNA3.1(-) to generate pcDNA3.1(-)/OPN
WT. Point mutations on threonine in each glycosylation site (T120A,
T124A, T129A, T133A and T138A) of OPN and three point mutations of
OPN (T124A, T133A and T138A; OPN TM), as well as five point
mutations of OPN in which all five OPN glycosylation sites were
converted to alanine (5 μm), were generated using pcDNA3.1(-)/OPN
WT as a template. The final constructs were confirmed via
restriction enzyme analysis and sequencing. To generate a stable
cell line, pcDNA3.1(-)/OPN WT or TM was transfected into the H226
cells using the TransIT®-LT1 reagent. The transfected
cells were selected with a medium that contained 100 g/ml of G418
disulfate salt from Sigma- Aldrich and the selected cells were
maintained in 50 g/ml of G418 disulfate salt for future
experiments.
RT-PCR
Total RNA was isolated using a TRIzol reagent, after
which RT-PCR was performed with the One-Step RT-PCR kit (Intron,
Seongnam, Korea), according to the manufacturer’s instructions. The
primers used were: OPN forward, 5′-GCAG AATCTCCTACCCCAC-3′ and
reverse, 5′-TCGGAATGC TCATTGCTCTC-3′; GALNac-T1 forward,
5′-CTGCCATGG TAGGTGTCCTG-3′ and reverse, 5′-TGAGGCTTGGAG
CACACTTC-3′; and GAPDH forward, 5′-GAAGGACTCAGA CCACAG-3′ and
reverse, 5′-CTTCACCACCTTCTTGATG-3′. GAPDH was amplified and used as
an internal control. The products were analyzed via electrophoresis
on 1% agarose gels.
Western blot analysis
The total protein concentration in the cell lysates,
media and homogenized tumor samples was determined using the
Bio-Rad Protein Assay reagent (Bio-Rad, Hercules, CA, USA). Equal
amounts of protein were separated via SDS-PAGE and transferred to
nitrocellulose membranes. The membranes were blocked in
Tris-buffered saline with Tween-20 (TBST) that contained 5% skim
milk. Immunoblotting was performed by overnight incubation with the
indicated primary antibodies at 4°C and then with HRP-conjugated
secondary antibodies for 3 h. The bands were detected using the
luminescent image analyzer LAS-3000 (Fujifilm, Tokyo, Japan). The
results of the western blot bands analysis were quantified with
Multi-gauge v2.02 software (Fujifilm).
Immunohistochemistry (IHC)
Fixed tumors were embedded in 10% neutral
phosphate-buffered formalin. The paraffin- embedded tissue sections
were cut and transferred to plus slides. The sections were
deparaffinized in xylene and rehydrated through alcohol gradients.
To quench the endogenous peroxidase activity, the sections were
incubated in 3% hydrogen peroxide for 10 min. After being washed in
PBS, the sections were incubated in PBS with 3% bovine serum
albumin (BSA) for 1 h at room temperature to block non-specific
binding sites. Corresponding primary antibodies were applied and
the sections were incubated overnight at 4°C. The following day,
they were washed and incubated with secondary HRP-conjugated
antibodies (1:50; Zymed) for 3 h at room temperature. After being
washed, the sections were incubated with DAB (Vector Laboratories,
Burlingame, CA, USA) and counterstained with Mayer’s hematoxylin
(Dako, Glostrup, Denmark). The slides were then imaged under a
light microscope (Carl Zeiss, Gottingen, Germany).
OPN immunoassay
The cells were transfected with OPN WT and TM
constructs. OPN levels from a cultured medium were then determined
via Quantikine Human OPN Immunoassay from R&D Systems
(Minneapolis, MN, USA). The results were read on a microplate
reader from Bio-Rad at 450 nm OD.
Immunofluorescence assay
The cells were grown and treated on a two-well
chamber slide. The slides were washed with PBS and fixed in 4%
paraformaldehyde and methanol, and in acetone, respectively. After
being blocked with 3% BSA, the sections were incubated with
specific primary antibodies, washed and incubated with
fluorescence-conjugated secondary antibodies. Their nuclei were
stained with DAPI. The slides were visualized using a fluorescent
microscope (Carl Zeiss).
In vivo tumor xenograft studies
Six-week-old nude mice were obtained from Joongang
Laboratory Animals, Inc. (Seoul, Korea). The mice were maintained
in a pathogen-free animal facility at least one week before use.
The nude mice were inoculated subcutaneously with H226 cells that
were non-transfected or stably transfected with OPN WT or TM. Seven
mice were used in each group. The volume of the tumor in each mouse
was measured with a caliper at regular intervals and calculated as
described previously (18,19).
At the end of the experiment, the mice were sacrificed and their
individual tumors were weighed and collected for subsequent
analysis.
Luciferase assay
The treated cells were lysed in a passive lysis
buffer from Promega and centrifuged for 10 min at 4°C. A
supernatant was used for the luminescence dose. The LucR and LucF
activities were measured using a Dual Luciferase kit from Promega
according to the manufacturer’s instructions.
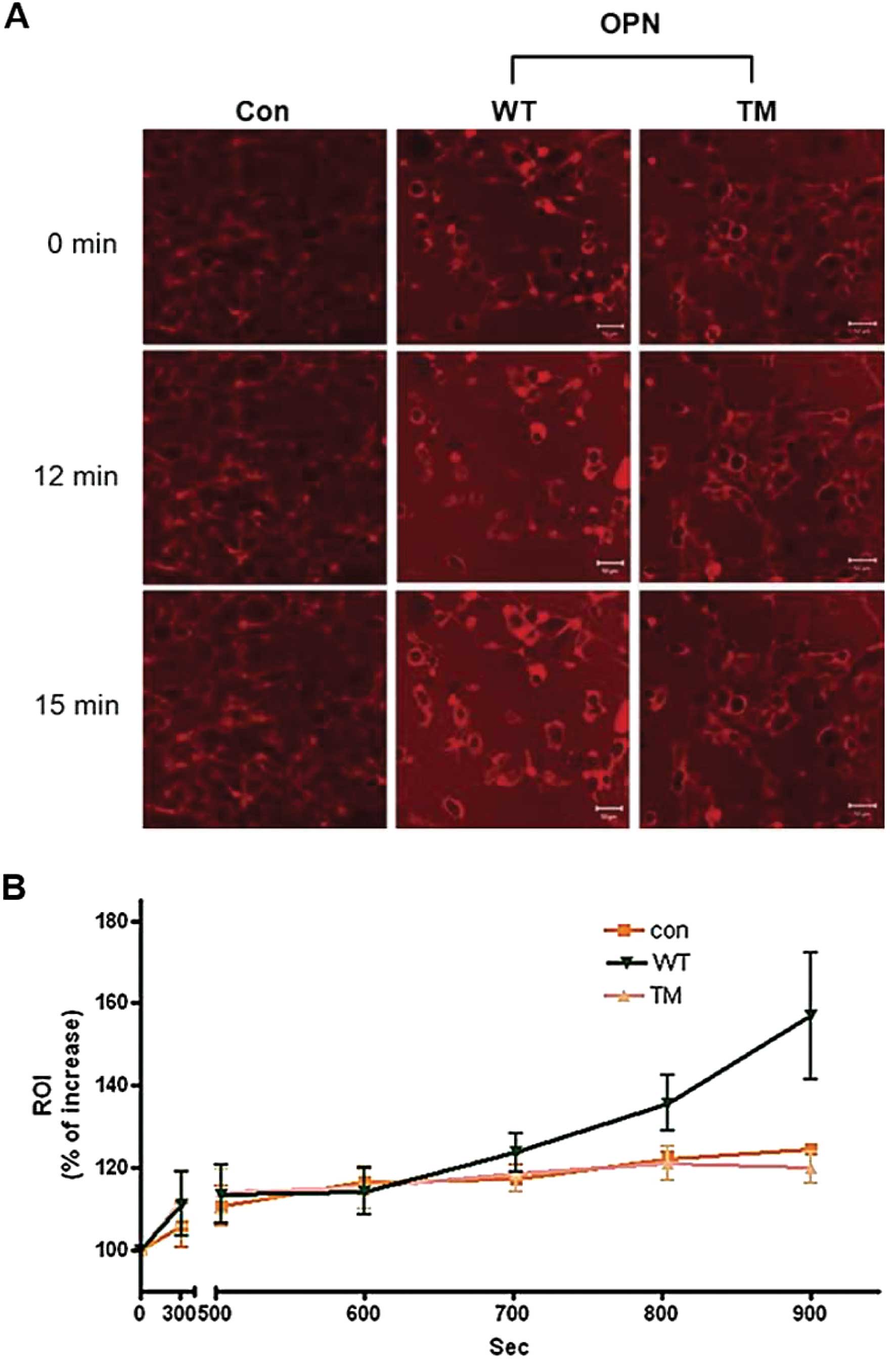
Cy3-labeled glucose uptake
H226 cells that stably expressed OPN WT and TM were
cultured on glass. Then, 6 μM of Cy3-labeled glucose (53) was added, and the viable cells were
monitored using confocal laser scanning microscopy (CLSM) within 15
min. Fluorescent images of the cells were captured every 60
sec.
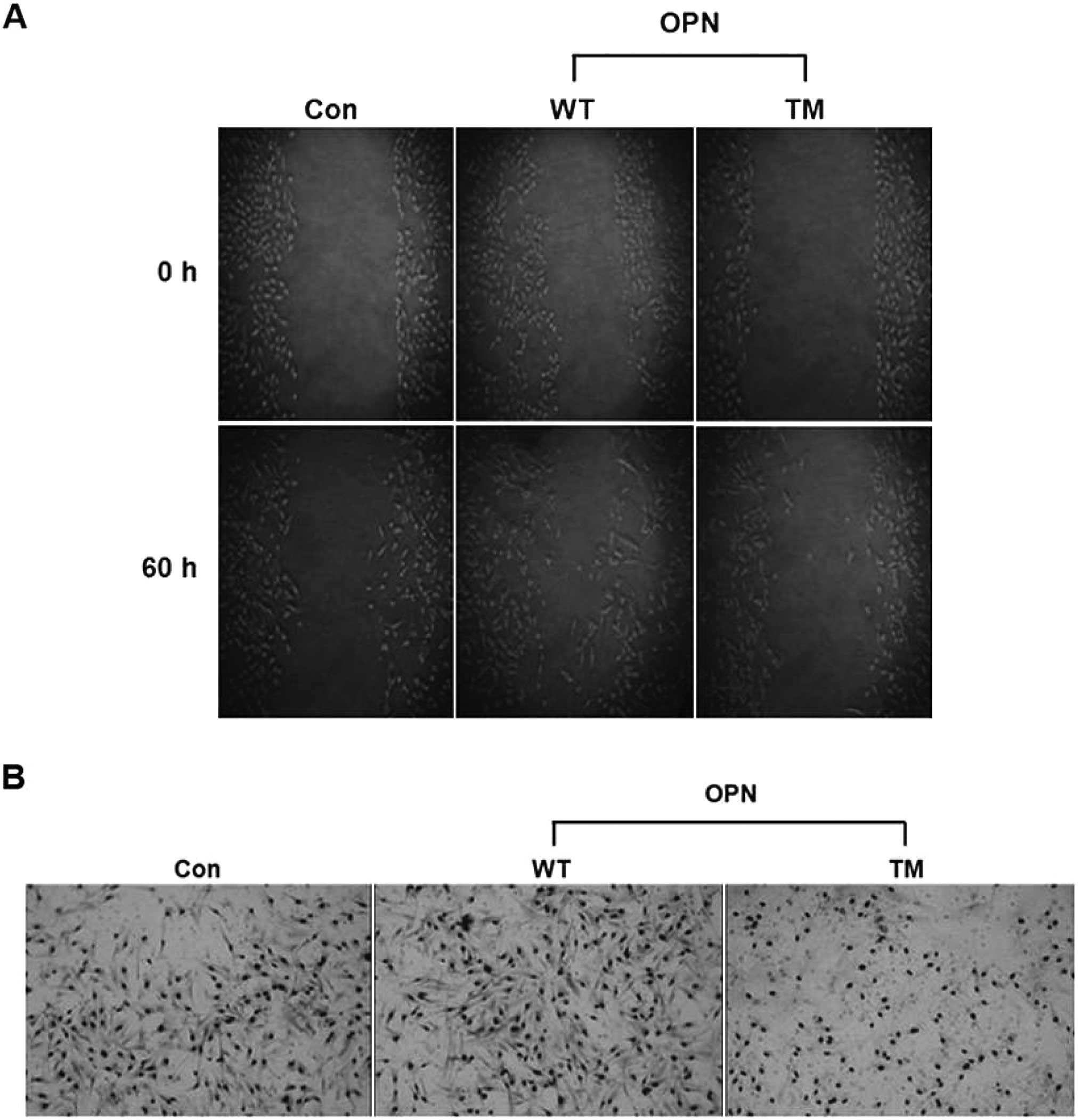
Wound healing assay
The confluent H226 cell monolayers were scratched
with a pipette tip, with 30 min pre-incubation in the presence of
mitomycin. The wound areas were observed using phase contrast
microscopy on an inverted microscope. Images of the same areas were
captured at regular intervals over the indicated time.
Invasion assay
Invasion assays were performed using BD BioCoat™
Matrigel™ Invasion Chambers with 8 μm polycarbonated filters from
BD Biosciences (San Jose, CA, USA) according to the manufacturer’s
instructions.
Nuclear extraction
The nucleus and cytosol were separated with the
Nuclear Extract kit from Active Motif (Carlsbad, CA, USA). The
cells were grown to confluence in a 100-mm tissue culture plate and
washed with 5 μl ice-cold PBS/phosphatase inhibitors. The collected
cells were gently resuspended in a 500 μl hypotonic buffer and
incubated on ice for 15 min. After their incubation, a 25 μl
detergent was added in the tube. The cytoplasmic fraction and the
nuclear pellet were separated through centrifugation. The nuclear
pellet was resuspended with a 50 μl complete lysis buffer and
incubated for 30 min on ice on a rocking platform at 150 rpm.
Following centrifugation, the supernatant containing nuclear
proteins was collected for further analysis.
Statistical analysis
Statistical analyses were performed with a Student’s
t-test for experiments on two groups using the Graphpad Software
(San Diego, CA, USA).
Results
Expression of OPN WT and TM in the H226
cells
To characterize the connection between OPN
O-glycosylation sites and their function, a series of point
mutations in O-glycosylation sites were introduced into the OPN
gene and the cells were co-transfected with OPN WT or
O-glycosylation mutant forms of OPN and NF-κB luciferase or
bicistronic luciferase construct. The results clearly showed that
the introduction of selective mutation on three O-glycosylation
sites of OPN reduced the NF-κB activity and the cap-dependent
protein translation more potently than other O-glycosylation mutant
forms of OPN (Fig. 1A and B).
Based on the results, OPN TM was chosen for subsequent
experiments.
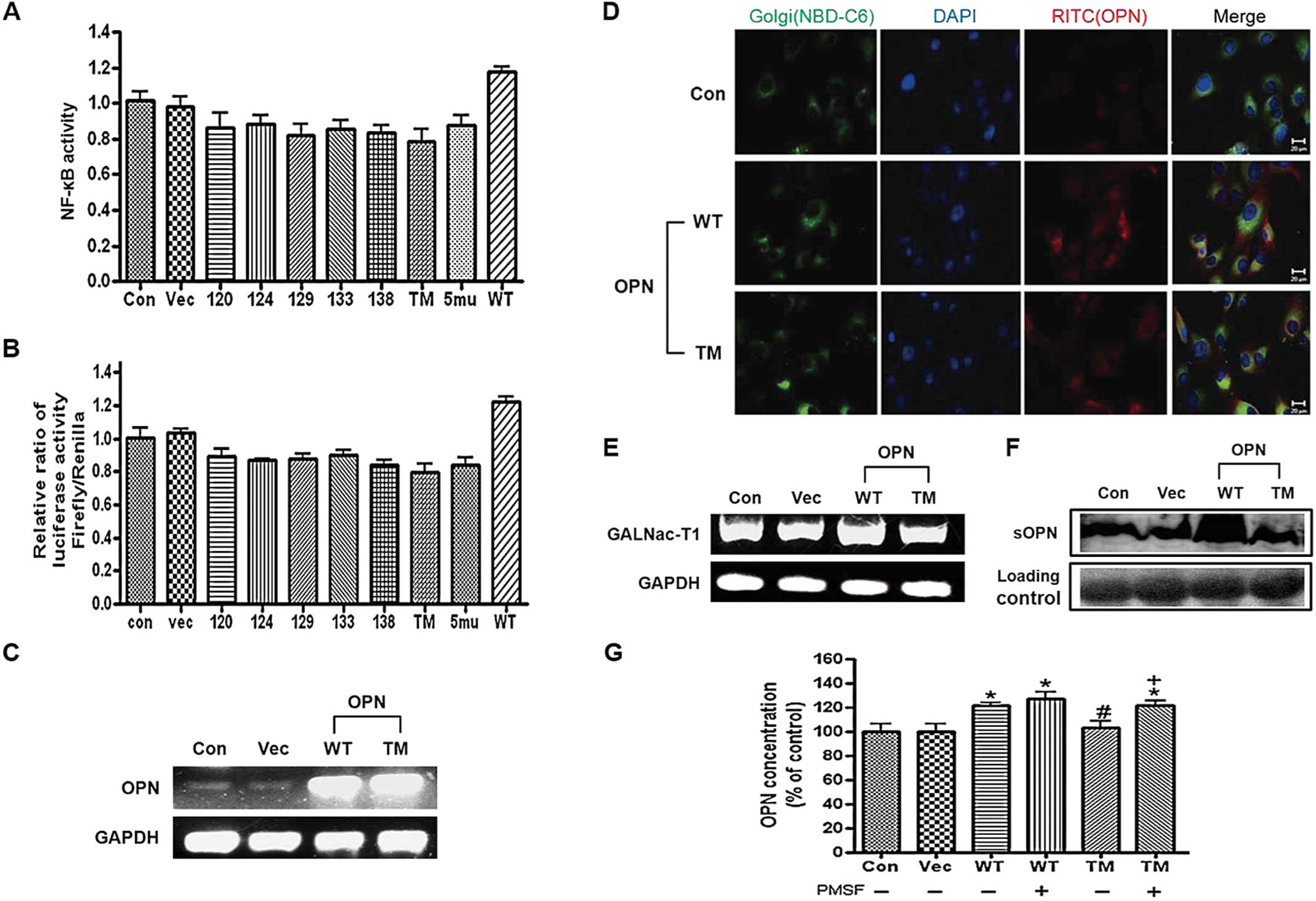 | Figure 1Expression of osteopontin (OPN)
wild-type (WT) and triple mutant (TM) in H226 cells. (A) For the
dual luciferase assay, the cells were co-transfected with different
OPN constructs, as indicated, and with the bicistronic reporter
construct pcDNAfLUC-polIRES-rLUC, and then lysed and assayed for
firefly and renilla luciferase activities. Each bar presents the
mean ± SEM (n=3). (B) Cells were co-transfected with OPN
constructs, as indicated, and the NF-κB luciferase vector with the
pRA-SV40 control vector, then lysed and assayed for firefly and
renilla luciferase activities. (C) RT-PCR analysis of OPN. H226
cells were transfected with OPN WT/TM for 24 h and then the total
RNA was isolated and subjected to RT-PCR. (D) Fluorescence imaging
of OPN after the transfection. H226 cells were incubated with OPN
WT and TM for 24 h, followed by fixing and immunostaining for the
OPN (red via TRITC), Golgi marker (green via NBD-C6 Ceramide) and
nuclei (blue via DAPI). The scale bar is 20 μm. (E) RT-PCR analysis
of GALNac-T1. H226 cells were transfected with OPN WT/TM for 24 h
and then the total RNA was isolated and subjected to RT-PCR.
Representative bands are shown. (F) Western blotting of the
secreted OPN. H226 cells were transected with OPN WT/TM and the
cultured media were collected and subjected to western blot
analysis. (G) ELISA of OPN. H226 cells were transfected with OPN
WT/TM and then treated with PMSF (0.5 mM), after which cultured
media were collected and ELISA was performed. Each bar presents the
mean ± SEM (n=3). *P<0.05 vs. Con,
#P<0.05 vs. WT [PMSF (-)], and +P<0.05
vs. TM [PMSF (-)]. Con, control (H226 cells); Vec, vector control;
WT, OPN WT-transfected cells; TM, OPN TM-transfected cells. |
The mRNA levels of OPN following the transfection of
the H226 cells with OPN WT and TM were then examined. The results
clearly showed that the mRNA level of OPN increased in the OPN
WT/TM-transfected cells (Fig.
1C). The immunofluorescence assay also showed the
overexpression of OPN in Golgi in the cells transfected with OPN WT
and TM, unlike in the control (Fig.
1D). The mRNA level of GalNAc-T1 in the H226 cells was also
determined. Unlike OPN WT, OPN TM did not increase the mRNA level
of GalNAc-T1 (Fig. 1E). The
western blot analysis and ELISA revealed that unlike OPN TM, OPN WT
increased the expression levels of OPN in conditioned media
(Fig. 1F and G). It was also
demonstrated that treatment of OPN WT/TM-transfected cells with
PMSF, a broad-spectrum protease inhibitor, increased the OPN
expression levels (Fig. 1G).
OPN WT, but not OPN TM, induced the
OPN-dependent signaling pathway
To evaluate the mechanisms of OPN regulation and the
role of OPN substrates in OPN-dependent biologic responses in cell
migration and growth, western blotting, immunofluorescence and
invasion and wound migration assays were carried out in cells
transfected with OPN WT/TM. The results clearly showed that unlike
OPN TM, OPN WT increased Akt, p-Akt (Thr308/Ser473), IKK, mTOR,
p-mTOR and Glut-1, as determined by the western blot and
densitometric analyses (Fig. 2A).
Unlike OPN WT, OPN TM did not increase the expression level of
p70S6K, p-p70S6K and uPA (Fig.
2B). The immunofluorescence assay reconfirmed this pattern
(Fig. 2C). Furthermore, the wound
healing and invasion assays demonstrated that OPN WT rather than
OPN TM increased cell migration and the number of invaded cells
(Fig. 3).
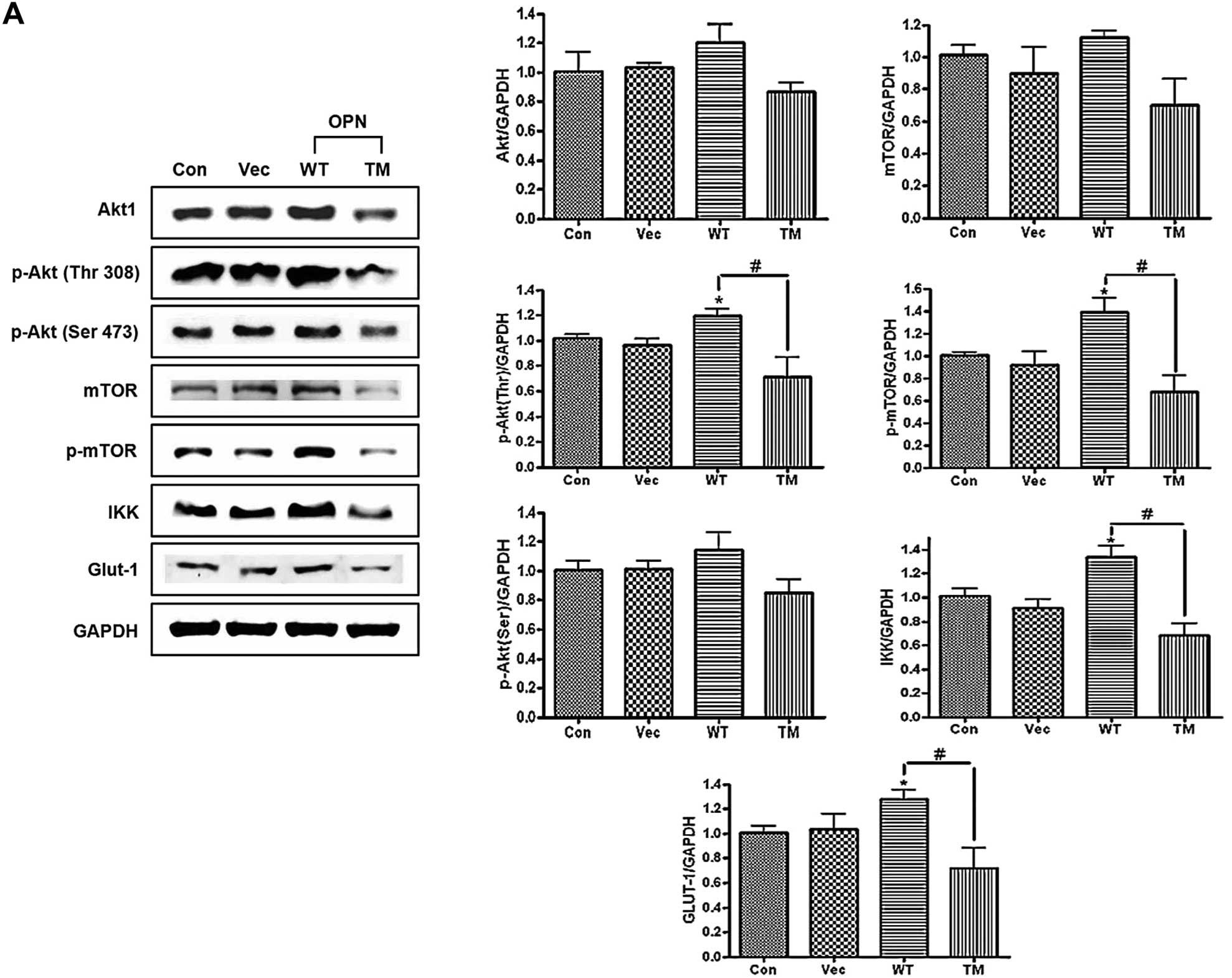 | Figure 2Western blot analysis of osteopontin
(OPN)-dependent proteins. (A) Western blot analysis of Akt, p-Akt
(Thr308), p-Akt (Ser473), mammalian target of rapamycin (mTOR),
p-mTOR, IKK and Glut-1. H226 cells were transfected with OPN WT/TM
and incubated for 48 h, followed by lysis and collection. The
lysates were used for the western blotting. The right panel shows
the results of the densitometric analysis of the bands of interest.
Each bar presents the mean ± SEM (n=3). *P<0.05 vs.
Con and #P<0.05 vs. TM. (B) Western blot analysis of
p70S6K, p-p70S6K and uPA. H226 cells were transfected with
osteopontin (OPN) wild-type (WT)/triple mutant (TM), as indicated
earlier. The lysates were used for the western blotting. (C) (Upper
panel) Fluorescence imaging of p-70S6K and p-p70S6K. (Lower panel)
Fluorescence imaging of uPA. H226 cells were incubated with OPN WT
and TM, followed by fixing and immunostaining for the indicated
proteins (red via FITC) and nuclei (blue via DAPI). The scale bar
is 20 μm. Con, control (H226 cells); Vec, vector control; WT, OPN
WT-transfected cells; TM, OPN TM-transfected cells. |
OPN TM did not increase the glucose
uptake, cap-dependent protein translation and NF-κB activity
Stable H226 cell lines that expressed OPN WT and TM
were generated to further explain the effects of OPN WT and TM on
glucose uptake and the OPN-dependent signaling pathway. The results
clearly showed that the glucose uptake increased in the cells that
stably expressed OPN WT, but did not increase in the cells that
stably expressed OPN TM (Fig. 4).
The altered expression levels of the key proteins involved in the
NF-κB signaling pathway when the stable cells were transfected with
Akt were investigated. The expression of Akt increased the
expression levels of phosphorylated forms of Akt at Thr308 and
NF-κB p65 in cytosol, whereas the NF-κB p65 expression in the
nucleus decreased. GAPDH and PARP were used as controls for the
cytosol and nucleus (Fig. 5A).
Furthermore, the Akt transfection enhanced the NF-κB activity in
the stable cells that expressed OPN WT and TM (Fig. 5B). The expression of mTOR
increased the expression levels of p-eIF4G and p-eIF4E (Fig. 5C) and the cap-dependent protein
translation in the cells that stably expressed OPN WT and TM
(Fig. 5D). The results also
showed that silencing p70S6K decreased the expression levels of
p-p70S6K and the cap-dependent protein translation in the cells
that stably expressed OPN WT and TM (Fig. 5E and F).
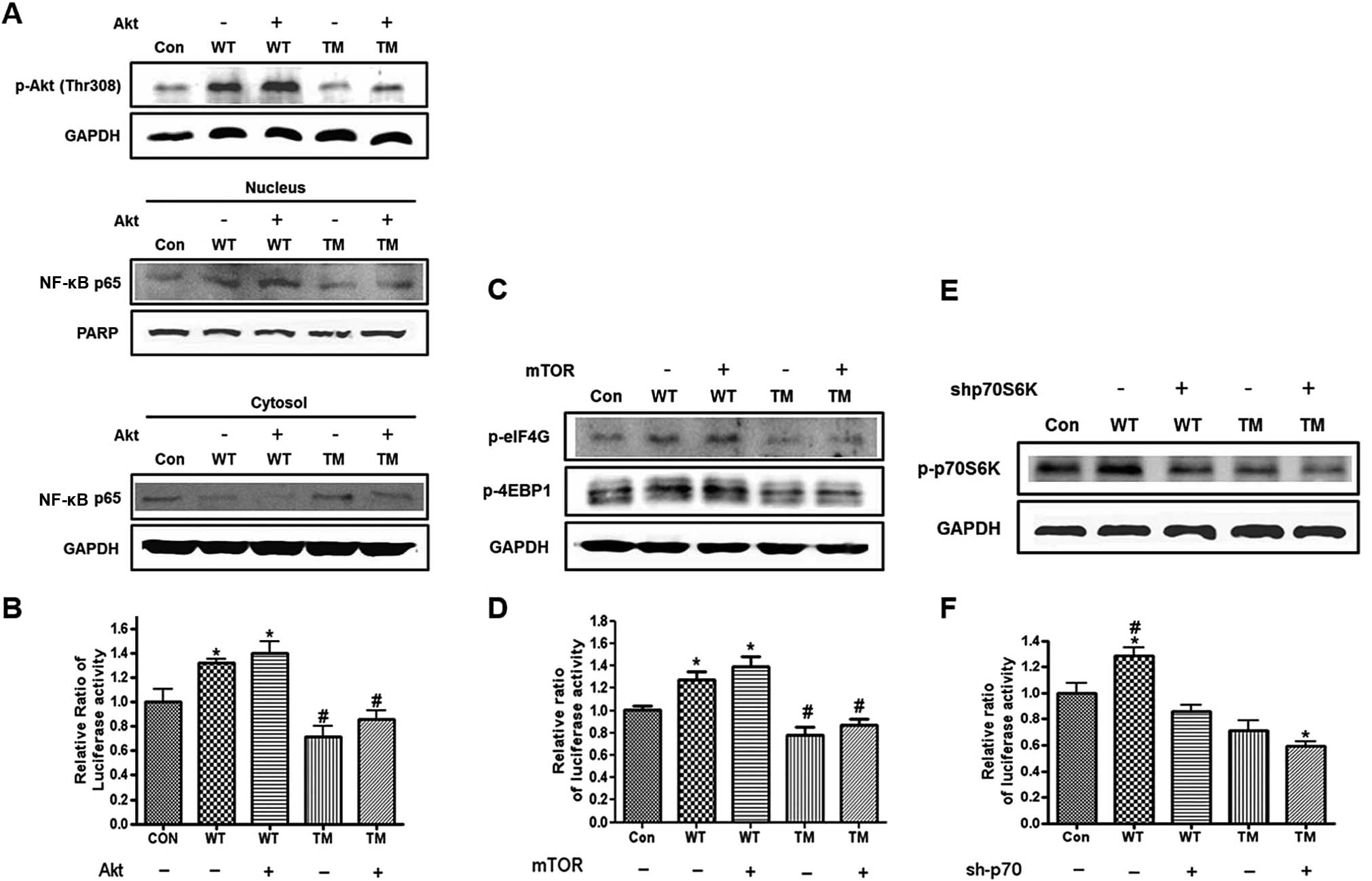 | Figure 5Western blot analysis of the proteins
that mediated the Akt/IKK and protein synthesis signaling pathway.
(A) Cells that stably expressed the osteopontin (OPN) wild-type
(WT)/triple mutant (TM) transfected with the Akt expression vector
and then subjected to western blot analysis for p-Akt (Thr308).
(Lower panel) Western blot analysis of NF-κB p65 in the cytosol and
nucleus. The cells were collected and then separated into cytosolic
and nucleic fractions using a Nuclear Extract kit. GAPDH and PARP
were used as controls for the cytosol and nucleus, respectively.
(B) For the dual luciferase assay, cells that stably expressed OPN
WT/TM were co-transfected with Akt and the NF-κB luciferase vector
with the pRA-SV40 control vector and then assayed for firefly and
renilla luciferase activities. Each bar presents the mean ± SEM
(n=3). *P<0.05 vs. Con and #P<0.05 vs.
WT+Akt and WT. Con, control (H226 cells); WT, cells that stably
expressed OPN WT; TM, cells that stably expressed OPN TM. (C) Cells
that stably expressed the OPN WT/TM that was transfected with the
mammalian target of rapamycin (mTOR) expression vector and then
subjected to western blot analysis for p-eIF4G and p-4EBP-1. (D)
For the dual luciferase assay, the cells that stably expressed the
OPN WT/TM were co-transfected with mTOR and bicistronic reporter
constructs, and then assayed for firefly and renilla luciferase
activities. Each bar presents the mean ± SEM (n=3).
*P<0.05 vs. Con and #P<0.05 vs. WT+mTOR
and WT. Con, control (H226 cells); WT, cells that stably expressed
the OPN WT; TM, cells that stably expressed the OPN TM. (E) Cells
that stably expressed the OPN WT/TM that was infected with
lentivirus which contained the shRNA that targeted p70S6K (10
ng/ml) and were then subjected to western blot analysis for
p-p70S6K. (F) For the dual luciferase assay, cells that stably
expressed the OPN WT/TM were infected and transfected with
bicistronic reporter constructs and then assayed for firefly and
renilla luciferase activities. Each bar presents the mean ± SEM
(n=3). *P<0.05 vs. Con and #P<0.05 vs.
other treated groups. Con, control (H226 cells); WT, cells that
stably expressed the OPN WT; TM, cells that stably expressed the
OPN TM. |
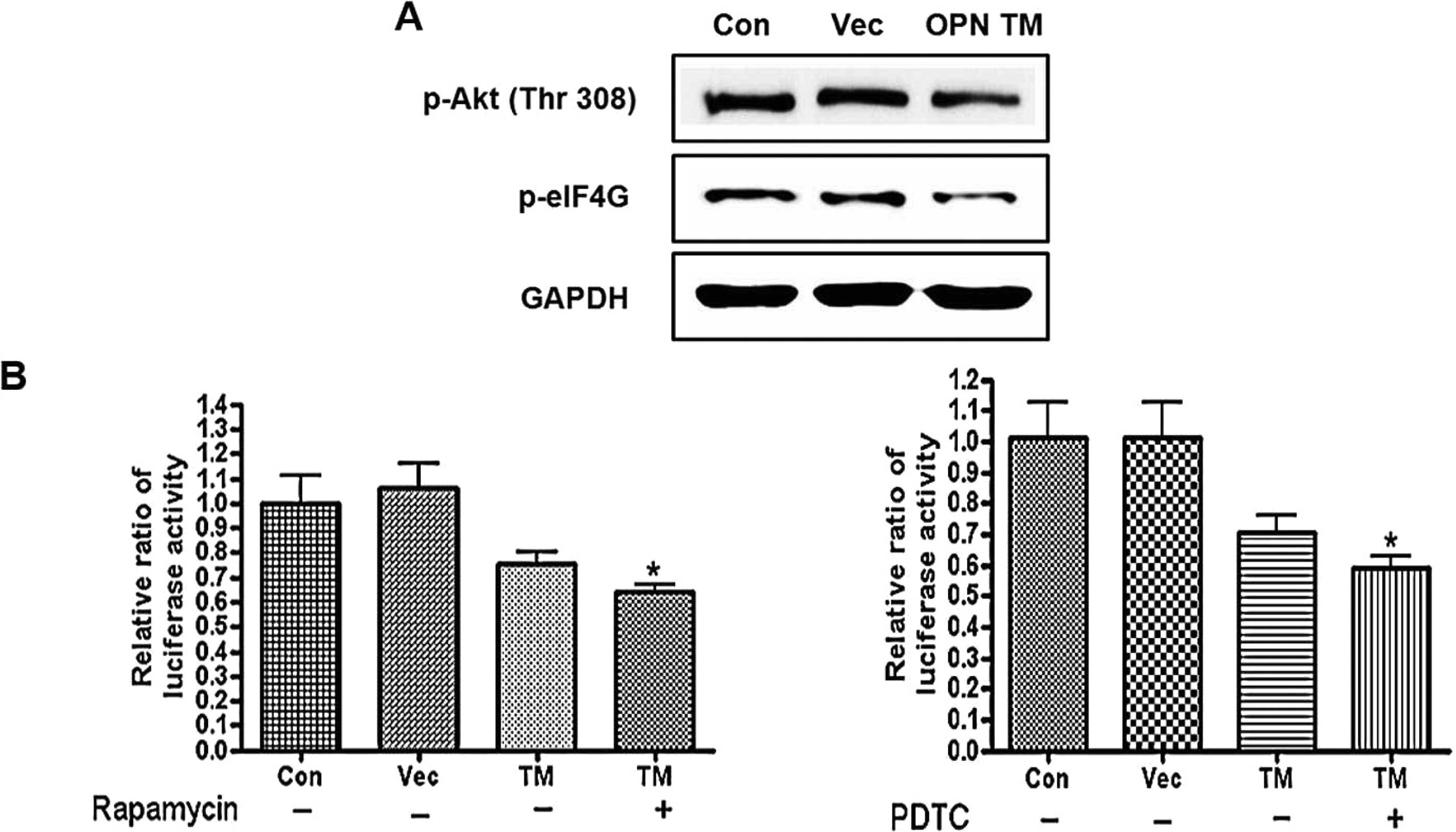
Involvement of the mTOR and IKK signaling
pathways in the OPN-mediated cap-dependent protein translation and
NF-κB activity
The effects of different inhibitors on OPN-mediated
protein translation and NF-κB activity were investigated. cis-Golgi
was blocked to the medial-Golgi transport with monensin (20) and the expression levels of p-Akt
(Thr308) and p-eIF4E were examined by western blot analysis. It was
found that the monensin treatment decreased the protein levels of
p-Akt (Thr308) and p-eIF4E in the OPN WT/TM-transfected cells
(Fig. 6A). Moreover, a
significant decrease in the cap-dependent protein translation was
observed in the OPN TM-transfected cells compared with the control.
Monensin also reduced the OPN WT-mediated cap-dependent protein
translation, as determined by the luciferase assay (Fig. 6B). To examine the role of mTOR and
the involvement of the Akt/mTOR signaling pathway in OPN-regulated
protein translation, the OPN WT/TM-transfected cells were treated
with rapamycin and the expression levels of p-eIF4G and p-4E-BP1
were examined. The results showed that rapamycin decreased the
expression levels of p-eIF4G and p-4E-BP1 in the OPN
WT/TM-transfected cells (Fig.
6C). The luciferase assay clearly demonstrated that rapamycin
attenuated the OPN-induced increase in the cap-dependent protein
translation. Moreover, a significant decrease was observed in the
OPN TM-transfected cells following rapamycin treatment (Fig. 6D). To investigate whether
OPN-mediated NF-κB activity occurs through the Akt/IKK signaling
pathway, the PI3K inhibitor LY294002 and the IKK inhibitor PDTC
were used. The results showed that LY294002 and PDTC decreased the
p-Akt (Thr308) and uPA expression levels, respectively, in the OPN
WT/TM-transfected cells (Fig. 6E and
G). The LY294002 and PDTC treatment decreased the protein level
of NF-κB p65 in the cytosol and an increase in the NF-κB p65
expression was observed in the nucleus of the cells that were
transfected with OPN WT and TM. Furthermore, PDTC or LY294002
inhibited the OPN-induced increase in NF-κB activity and a
significant decrease was observed in the OPN TM-transfected cells
following treatment with LY294002 or PDTC (Fig. 6F and H).
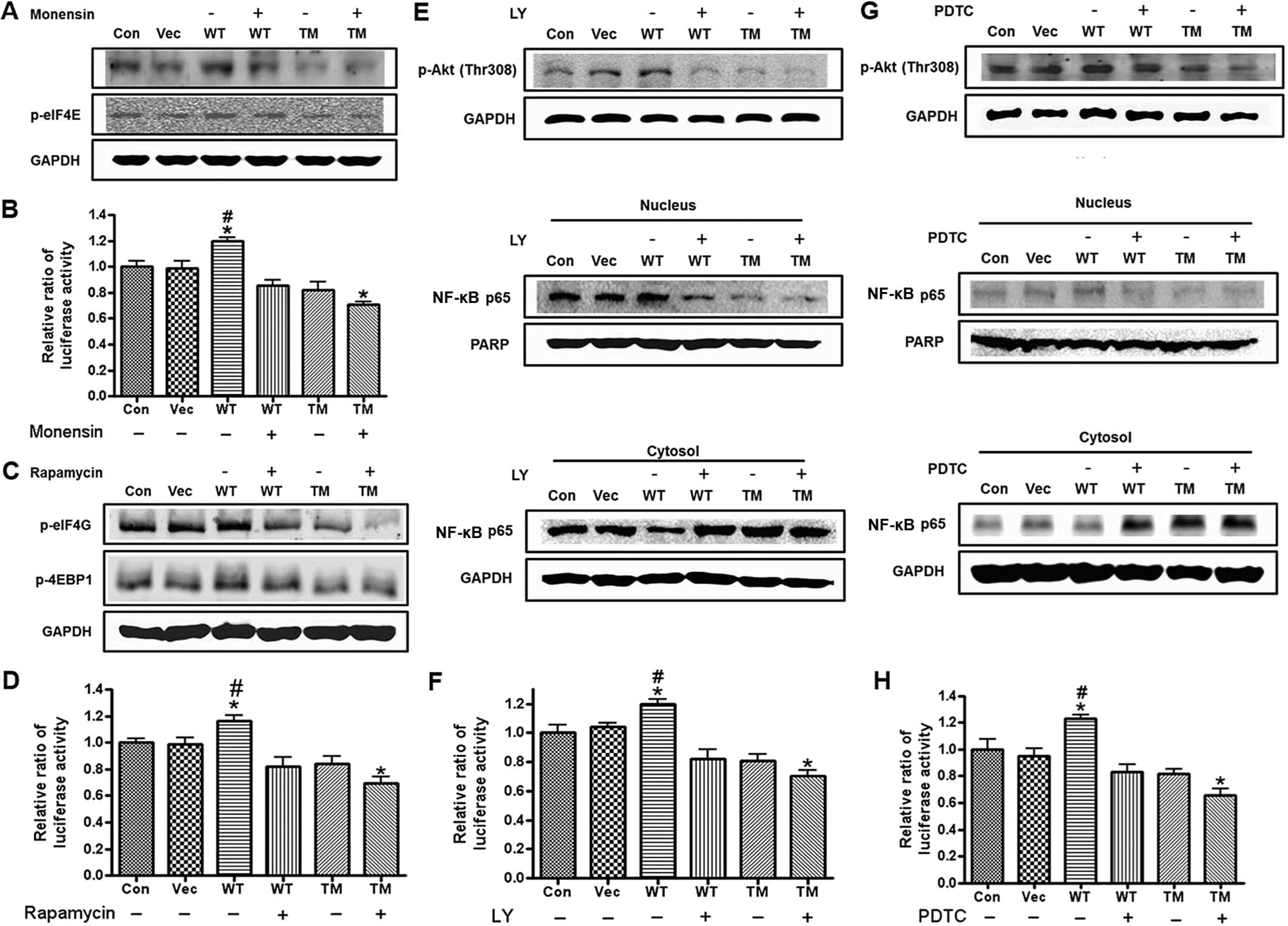 | Figure 6Effect of monensin, rapamycin,
LY294402 and PDTC in cells transfected with osteopontin (OPN)
wild-type (WT) and triple mutant (TM). (A) Western blot analysis of
p-Akt (Thr308) and p-eIF4E. H226 cells were transfected with OPN
WT/TM and then treated with monensin (0.5 mM). The lysates were
used for the western blotting. (B) For the dual luciferase assay,
cells were co-transfected with OPN constructs (vector only, OPN WT
and OPN TM) and the bicistronic reporter construct
pcDNAfLUC-polIRES-rLUC and then treated with monensin or left
untreated. The cells were lysed and assayed for firefly and renilla
luciferase activities. Each bar presents the mean ± SEM (n=3).
*P<0.05 vs. Con and #P<0.05 vs. other
treated groups. Con, control (H226 cells); Vec, vector control; WT,
OPN WT-transfected cells; TM, OPN TM-transfected cells. (C) Western
blot analysis of p-4E-BP1 and p-eIF4G. H226 cells were transfected
with OPN WT/TM and then treated with rapamycin (100 nM). The
lysates were used for the western blotting. (D) For the dual
luciferase assay, the cells were co-transfected with OPN constructs
(vector only, OPN WT and OPN TM) and the bicistronic reporter
construct pcDNAfLUC-polIRES-rLUC, and then treated with rapamycin
or left untreated. The cells were then lysed and assayed for
firefly and renilla luciferase activities. Each bar presents the
mean ± SEM (n=3). *P<0.05 vs. Con and
#P<0.05 vs. other treated groups. Con, control (H226
cells); Vec, vector control; WT, OPN WT-transfected cells; TM, OPN
TM-transfected cells. (E) Western blot analysis of p-Akt (Thr308).
H226 cells were transfected with OPN WT/TM and then treated with
LY294402 (100 nM). The lysates were used for the western blotting.
(Lower panel) Western blot analysis of NF-κB p65 in the cytosol and
nucleus. The cells were collected and separated into cytosolic and
nucleic fractions using a Nuclear Extract kit. GAPDH and PARP were
used as controls for the cytosol and nucleus, respectively. (F) For
the dual luciferase assay, the cells were co-transfected with OPN
constructs (vector only, OPN WT and OPN TM) and the NF-κB
luciferase vector with the pRA-SV40 control vector and then treated
with LY294002 or left untreated. The cells were then lysed and
assayed for firefly and renilla luciferase activities. Each bar
presents the mean ± SEM (n=3). *P<0.05 vs. Con and
#P<0.05 vs. other treated groups. Con, control (H226
cells); Vec, vector control; WT, OPN WT-transfected cells; TM, OPN
TM-transfected cells. (G) Western blot analysis of uPA. H226 cells
were transfected with OPN WT/TM and then treated with LY294402 (100
nM). The lysates were used for the western blotting. (Lower panel)
Western blot analysis of NF-κB p65 in the cytosol and nucleus. The
cells were collected and then separated into cytosolic and nucleic
fractions using a Nuclear Extract kit. GAPDH and PARP were used as
controls for the cytosol and nucleus, respectively. (H) For the
dual luciferase assay, the cells were co-transfected with OPN
constructs (vector only, OPN WT and OPN TM) and the NF-κB
luciferase vector with the pRA-SV40 control vector and then treated
with PDTC or left untreated. The cells were then lysed and assayed
for firefly and renilla luciferase activities. Each bar presents
the mean ± SEM (n=3). *P<0.05 vs. Con and
#P<0.05 vs. other treated groups. Con, control (H226
cells); Vec, vector control; WT, OPN WT-transfected cells; TM, OPN
TM-transfected cells. |
Effect of OPN TM on the OPN-mediated
signaling pathway in H322 cells
The effect of OPN TM on the NF-κB activity,
cap-dependent protein translation, and OPN-mediated signaling
pathway in H322 cells was also determined. The results showed that
OPN TM decreased the protein levels of p-Akt, p-mTOR, p-eIF4G and
p-eIF4E in the H322 cells (Fig.
7A). We also showed that OPN TM clearly decreased the NF-κB
activity and cap-dependent protein translation, but that the PDTC
and rapamycin treatment further decreased the cap-dependent protein
translation and NF-κB activity in the OPN TM-transfected cells
(Fig. 7B). These results suggest
that OPN TM is capable of altering the OPN-dependent signaling
pathway in H322 cells that reportedly have high levels of
endogenous OPN (21).
Effects of OPN WT and TM on tumor growth
in a xenograft mouse model
Evidence has shown that OPN increases tumor growth
in several in vivo cancer models (22,23). Due to the in vitro data
obtained, we investigated the effects of OPN WT and TM on tumor
growth in a xenograft mouse model. Accordingly nude mice were
injected subcutaneously with H226 cells that were alone or stably
transfected with OPN WT or TM. The data showed that OPN WT
increased whereas OPN TM decreased the tumor growth and weight in
the xenograft mouse model (Fig.
8A–C). It has been demonstrated that several OPN-dependent
proteins, such as MMP-9, VEGF, FGF-2, COX-2 and uPA, are important
in the angiogenesis, invasion and growth of tumor cells. Therefore,
the expression levels of these proteins in a tumor xenograft model
were investigated using western blotting and immunohistochemistry.
The results showed that the expression of MMP-9, VEGF and FGF-2
increased in tumors generated with OPN WT-transfected cells but not
with OPN TM-transfected cells, as determined by western blot and
densitometric analyses (Fig. 8D).
Furthermore, the IHC analysis showed a decrease in the expression
levels of uPA and COX-2 in tumors generated with OPN TM-transfected
cells and an increase in tumors generated with OPN WT-transfected
cells (Fig. 8E).
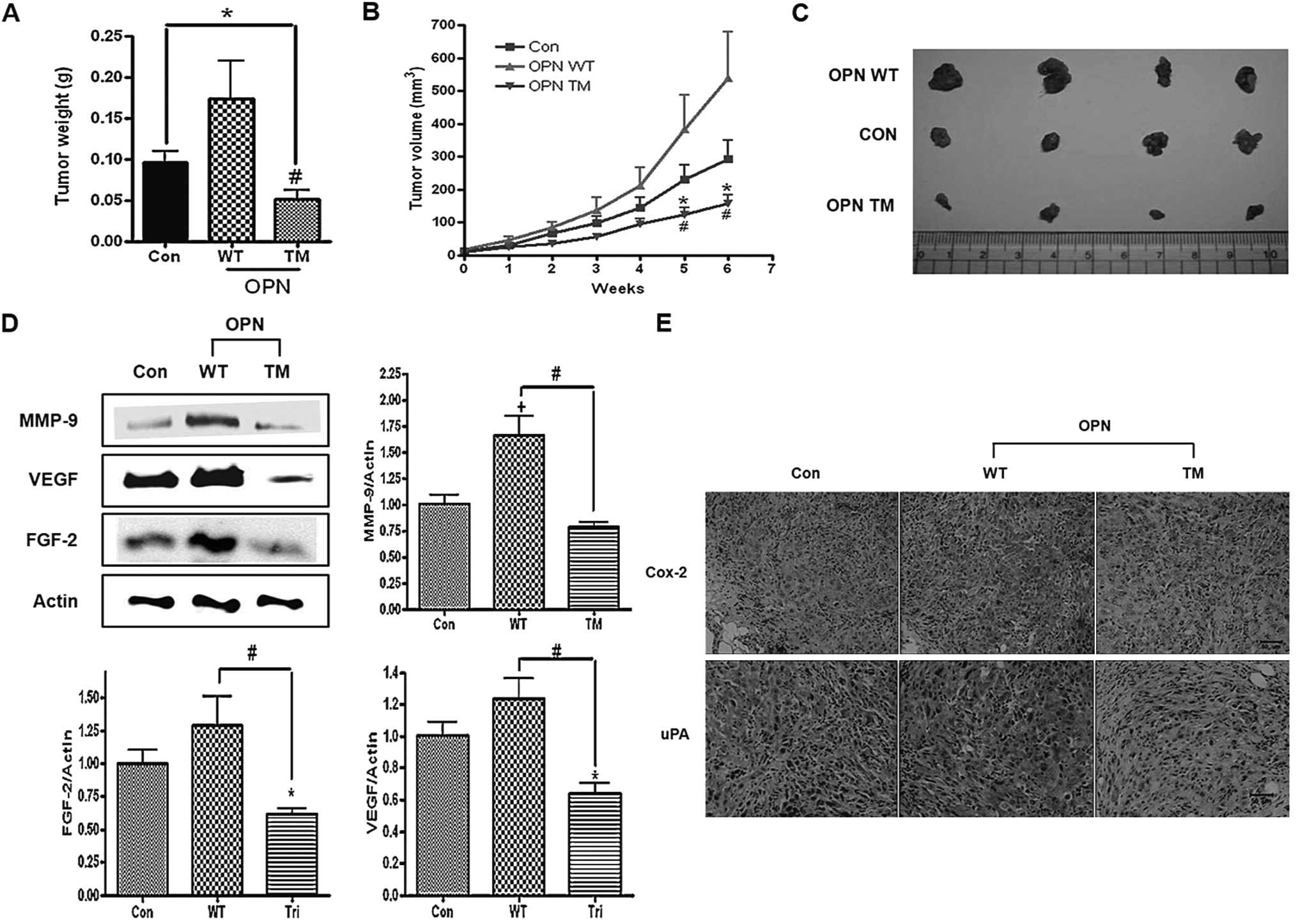 | Figure 8Effects of osteopontin (OPN)
wild-type (WT) and triple mutant (TM) on tumor growth,
angiogenesis, migration and invasion in xenograft model mice. Nude
mice inoculated subcutaneously with H226 cells that were either
non-transfected or stably transfected with OPN WT or OPN TM. (A)
Bar graph of the measured tumor weights. Each bar presents the mean
± SEM (n=7). *P<0.05 vs. Con and
#P<0.05 vs. WT. (B) The tumor growth was monitored
and the volumes were calculated, as described in Materials and
methods. *P<0.05 compared to Con and
#P<0.05 compared to WT. (C) Representative image of
the tumors in the nude mice. Con, control; WT, OPN WT xenograft
tumor; TM, OPN TM xenograft tumor. Tumor homogenates subjected to
western blot analysis. The blots were probed with antibodies, as
indicated. (D) Expression levels of MMP-9, VEGF and FGF-2. The
right panel shows the results of the densitometric analysis of the
bands of interest. Each bar presents the mean ± SEM (n=3).
*P<0.05 vs. Con and #P<0.05 vs. TM. (E)
Immunohistochemical analysis of COX-2 and uPA (magnification, ×200;
bar =50 μm; Con, control; WT, OPN WT xenograft tumor; TM, OPN TM
xenograft tumor). |
Discussion
The abundance of clinical and experimental evidence
regarding the relationship between OPN and lung cancer growth makes
OPN an attractive potential therapeutic target for combating lung
cancer development (24,25). A specific OPN inhibitor is not yet
available, however. Therefore, inhibition of the OPN-mediated
signaling pathway is a potentially attractive strategy for lung
cancer therapy.
The importance of the O-glycosylation sites in OPN
and the connection between the OPN function and its glycosylation
have yet to be adequately clarified. In the present study, the
effect of the O-glycosylation sites in OPN on cancer cell growth
and migration was functionally defined by introducing selective
mutation in three O-glycosylation sites of OPN and expressing this
mutation in lung cancer cells. Overexpression of the OPN that
contained WT glycosylation sites but not of that with the three
mutated O-glycosylation sites increased lung cancer cell growth and
migration in vitro as well as in a xenograft mouse model.
Specifically, the specified O-glysylation sites were important for
lung cancer progression and growth. Glycosylation is one of the
most important post-translational events wherein glycan chains are
connected to a specific peptide backbone. The uncertainty in
predicting the correlation of OPN O-glycosylation with its function
necessitates the examination of the relevance of O-glycosylation of
OPN in its functional environment. Therefore, the extent of
glycosylation in OPN must be determined, and the consequence of the
blocking or modification of the glycosylation of OPN must be
investigated (26–28). In the present study, this issue
was investigated by studying the connection between O-glycosylation
and the cell function of OPN by expressing OPN TM and WT in lung
cancer cells. The results clearly showed that OPN WT and TM
increased the OPN mRNA levels. The overexpression of OPN in Golgi
was also demonstrated via CLSM.
The transfer of α-N-acetylgalactosamine (GalNAc)
from UDP-GalNAc to Ser or Thr is catalyzed by
UDP-GalNAc:polypeptide N-acetylgalactosaminyltransferases
(GalNAc-Ts) in Golgi (29,30).
Miwa et al (11)
determined the O-glycosylation sites of OPN as potential acceptor
substrates of GalNAc-T1 in vitro and demonstrated that two
of its glycosylation sites, which are also conserved in humans,
were glycosylated by GalNAc-T1. The results showed that OPN WT
increased the expression levels of GalNAc-T1, whereas OPN TM did
not, suggesting that the introduction of OPN regulates the
expression of O-glycosylation-related proteins in the Golgi, which
can affect its O-glycosylation process and suggests that unlike OPN
WT, OPN TM cannot be fully glycosylated. It has been reported that
PTMs that include O-glycosylation can affect protein conformation,
stability and other properties (31–34). Glycosylation modification has been
reported to protect proteins against proteasomal degradation
(35–37). Unlike OPN WT, OPN TM did not
increase the expression of the secreted OPN; however, when the
cells were treated with PMSF, the expression levels of OPN were
recovered. The data clearly show that non-fully-glycosylated OPN,
in contrast to fully glycosylated OPN, generates different
functional forms that may alter their physiological functions.
Mutations at specific O-glycosylation sites of OPN may generally
result in lower protein stability, indicating that there is a
correlation between alteration in OPN O-glycosylation and its
stability against proteasome degradation. The results clearly show
that unlike OPN WT, OPN TM did not increase Akt and p-Akt at the
phosphorylation sites. It has been reported that OPN is able to
induce Akt and that its related signals contribute to cancer
progression (1,38).
Translational control, a crucial element in the
regulation of gene expression, has been reported to be frequently
deregulated in cancer cells (39–41). Translation is regulated via the
mTOR signaling pathway through p70S6K and 4E-BP1/eIF4E, wherein
OPN-activated Akt can be a crucial intermediate. A key step in the
regulation of protein synthesis is the assembly of the initiation
complex eIF4F that contains the initiation factors eIF4G, eIF4E
(the protein that binds to the 5′ cap structure) and eIF4A. 4E-BP1
has been reported to bind to a site on eIF4E that overlaps with the
binding site for eIF4G (42–44). Thus, 4E-BP1 competes with eIF4G in
order to bind to eIF4E, and is thus able to inhibit the eIF4F
complex formation. Phosphorylation of 4E-BP1 reduces its affinity
to eIF4E and its dissociation, making eIF4E available for binding
to eIF4G and thereby increasing the mRNA translation (45,46). The results have shown that OPN TM
decreased the expression levels of mTOR, p-mTOR, p70S6K, p-p70S6K,
p-4E-BP1, p-eIF4E and p-eIF4G, but increased the expression levels
in the OPN WT-treated cells. The results also showed that the
expression of mTOR induced the cap-dependent protein translation,
whereas rapamycin and shp70S6K inhibited the cap-dependent protein
translation in the cells treated with OPN WT and TM. These data
reveal that OPN WT induced cap-dependent protein translation
through the Akt-mTOR pathway, and that OPN TM inhibits
cap-dependent protein translation which can affect lung cancer cell
growth.
Recent studies have indicated that OPN plays a
crucial role in controlling cell motility. Moreover, NF-κB
activation has been reported to be a major effector of OPN-mediated
cell migration and motility (47,48). In an inactivated state, NF-κB is
usually kept in the cytoplasm by the inhibitor protein IκB. Upon
the NF-κB stimulation, the IκB proteins are phosphorylated by the
multi-sub-unit IKK complex, which subsequently targets IκB for
ubiquitination. Additionally, the free NF-κB translocates to the
nucleus and activates the target genes (49). The results show that OPN WT
increased NF-κB activity, which enhanced the motility of the lung
cancer cells by activating Akt, IKK, NF-κB p-65 and uPA, unlike OPN
TM. By contrast, treatment of the cells with LY294002 and PDTC
reduced the NF-κB activity induced by OPN WT and caused a further
decrease in the cells transfected with OPN TM, whereas the
expression of Akt increased NF-κB activity, suggesting that an OPN
WT-induced increase in cell motility through the NF-κB signaling
pathway can be antagonized by the introduction of OPN TM.
Glucose is the most important energy source for cell
growth. Rapidly growing cancer cells need more glucose than normal
cells. Glucose passage across the cell membrane is regulated by a
family of glucose transporters (Gluts). In non-small-cell lung
cancer (NSCLC), glucose uptake is mediated by Glut-1, a significant
indicator of poor prognosis in NSCLC (50,51). It has been reported that the NF-κB
pathway controls Glut-1 trafficking and activation (52). In the present study, a novel
fluorescence-labeled glucose analogue, Cy3-linked O-1-glycosylated
glucose, was used, which can behave as D-glucose to measure the
glucose uptake. Cy3-labeled glucose is tolerant to intense light
sources and can be applied in a bioassay system without glucose
starvation (53). The results of
the present study show that OPN TM did not increase the Glut-1
expression and the glucose uptake, which may decrease the glucose
consumption rate, unlike OPN WT.
The decreased expression of several proteins
associated with angiogenesis, invasion, and migration, such as
MMP-9, FGF-2, VEGF, uPA and COX-2, in the OPN TM xenograft tumor
suggests that OPN TM may reduce tumor progression and growth,
whereas the increased expression levels of the indicated proteins
increased tumor growth in the OPN WT xenograft mouse model.
Taken together, the results have shown that unlike
OPN WT, OPN TM did not induce Akt-mediated cap-dependent protein
translation and NF-κB activity. Overexpression of the TM of OPN may
compete with endogenous OPN for substrates that can be limiting,
which may prevent endogenous OPN from performing its proper
function. Therefore, targeting OPN and regulating the OPN signaling
and function by influencing post-translational glycosylation
through the introduction of an inactive mutant of OPN can control
lung cancer growth and migration and may have a useful application
in the treatment of lung cancer.
Acknowledgements
The present study was supported by the National
Research Foundation grants (NRF-2009-0078641 and 2012M3A9C4048819)
from the Ministry of Education, Science and Technology (MEST).
M.H.C. was also partially supported by the Research Institute for
Veterinary Science, Seoul National University.
References
|
1
|
Donati V, Boldrini L, Dell’Omodarme M, et
al: Osteopontin expression and prognostic significance in non-small
cell lung cancer. Clin Cancer Res. 11:6459–6465. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ahmed M, Behera R, Chakraborty G, et al:
Osteopontin: a potentially important therapeutic target in cancer.
Expert Opin Ther Targets. 15:1113–1126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weber GF: The metastasis gene osteopontin:
a candidate target for cancer therapy. Biochim Biophys Acta.
1552:61–85. 2001.PubMed/NCBI
|
|
4
|
Shevde LA, Das S, Clark DW and Samant RS:
Osteopontin: an effector and an effect of tumor metastasis. Curr
Mol Med. 10:71–81. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chellaiah MA, Biswas RS, Rittling SR,
Denhardt DT and Hruska KA: Rho-dependent Rho kinase activation
increases CD44 surface expression and bone resorption in
osteoclasts. J Biol Chem. 278:29086–29097. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Christensen B, Kazanecki CC, Petersen TE,
Rittling SR, Denhardt DT and Sørensen ES: Cell type-specific
post-translational modifications of mouse osteopontin are
associated with different adhesive properties. J Biol Chem.
282:19463–19472. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fukuda M: Roles of cell surface
carbohydrates in cell adhesion; in particular those for sialyl
Lewis x and polysialic acid. Seikagaku. 72:269–283. 2000.(In
Japanese).
|
|
8
|
López-Ferrer A, Barranco C and de Bolós C:
Differences in the O-glycosylation patterns between lung squamous
cell carcinoma and adenocarcinoma. Am J Clin Pathol. 118:749–755.
2002.PubMed/NCBI
|
|
9
|
Christensen B, Nielsen MS, Haselmann KF,
Petersen TE and Sørensen ES: Post-translationally modified residues
of native human osteopontin are located in clusters: identification
of 36 phosphorylation and five O-glycosylation sites and their
biological implications. Biochem J. 390:285–292. 2005. View Article : Google Scholar
|
|
10
|
Keykhosravani M, Doherty-Kirby A, Zhang C,
Brewer D, Goldberg HA, Hunter GK and Lajoie G: Comprehensive
identification of post-translational modifications of rat bone
osteopontin by mass spectrometry. Biochemistry. 44:6990–7003. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Miwa HE, Gerken TA, Jamison O and Tabak
LA: Isoform-specific O-glycosylation of osteopontin and bone
sialoprotein by polypeptide N-acetylgalactosaminyltransferase-1. J
Biol Chem. 285:1208–1219. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aggarwal BB, Vijayalekshmi RV and Sung B:
Targeting inflammatory pathways for prevention and therapy of
cancer: short-term friend, long-term foe. Clin Cancer Res.
15:425–430. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aggarwal BB and Sung B: NF-κB in cancer: a
matter of life and death. Cancer Discov. 1:469–471. 2011.
|
|
14
|
Chaturvedi MM, Sung B, Yadav VR, Kannappan
R and Aggarwal BB: NF-κB addiction and its role in cancer: ‘one
size does not fit all’. Oncogene. 30:1615–1630. 2011.
|
|
15
|
Ruggero D and Pandolfi PP: Does the
ribosome translate cancer? Nat Rev Cancer. 3:179–192. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Abraham RT and Eng CH: Mammalian target of
rapamycin as a therapeutic target in oncology. Expert Opin Ther
Targets. 12:209–222. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hagner PR, Schneider A and Gartenhaus RB:
Targeting the translational machinery as a novel treatment strategy
for hematologic malignancies. Blood. 115:2127–2135. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim YK, Kwon JT, Choi JY, et al:
Suppression of tumor growth in xenograft model mice by programmed
cell death 4 gene delivery using folate-PEG-baculovirus. Cancer
Gene Ther. 17:751–760. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Minai-Tehrani A, Jiang HL, Kim YK, et al:
Suppression of tumor growth in xenograft model mice by small
interfering RNA targeting osteopontin delivery using biocompatible
poly(amino ester). Int J Pharm. 431:197–203. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tartakoff AM: Perturbation of vesicular
traffic with the carboxylic ionophore monensin. Cell. 32:1026–1028.
1983. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shijubo N, Uede T, Kon S, et al: Vascular
endothelial growth factor and osteopontin in stage I lung
adenocarcinoma. Am J Respir Crit Care Med. 160:1269–1273. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Philip S, Bulbule A and Kundu GC:
Osteopontin stimulates tumor growth and activation of promatrix
metalloproteinase-2 through nuclear factor-kappa B-mediated
induction of membrane type 1 matrix metalloproteinase in murine
melanoma cells. J Biol Chem. 276:44926–44935. 2001. View Article : Google Scholar
|
|
23
|
Jain S, Chakraborty G and Kundu GC: The
crucial role of cyclooxygenase-2 in osteopontin-induced protein
kinase C alpha/c-Src/IkappaB kinase alpha/beta-dependent prostate
tumor progression and angiogenesis. Cancer Res. 66:6638–6648. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rodrigues LR, Teixeira JA, Schmitt FL,
Paulsson M and Lindmark-Mänsson H: The role of osteopontin in tumor
progression and metastasis in breast cancer. Cancer Epidemiol
Biomarkers Prev. 16:1087–1097. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xue YH, Zhang XF, Dong QZ, et al: Thrombin
is a therapeutic target for metastatic osteopontin-positive
hepatocellular carcinoma. Hepatology. 52:2012–2022. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chang VT, Crispin M, Aricescu AR, et al:
Glycoprotein structural genomics: solving the glycosylation
problem. Structure. 15:267–273. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Patsos G, Robbe-Masselot C, Klein A, et
al: O-glycan regulation of apoptosis and proliferation in
colorectal cancer cell lines. Biochem Soc Trans. 35:1372–1374.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Varki A: Glycan-based interactions
involving vertebrate sialic-acid-recognizing proteins. Nature.
446:1023–1029. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ten Hagen KG, Tetaert D, Hagen FK, et al:
Characterization of a UDP-GalNAc: polypeptide
N-acetylgalactosaminyltransferase that displays glycopeptide
N-acetylgalactosaminyltransferase activity. J Biol Chem.
274:27867–27874. 1999.PubMed/NCBI
|
|
30
|
Perrine CL, Ganguli A, Wu P, et al:
Glycopeptide-preferring polypeptide GalNAc transferase 10 (ppGalNAc
T10), involved in mucin-type O-glycosylation, has a unique
GalNAc-O-Ser/Thr-binding site in its catalytic domain not found in
ppGalNAc T1 or T2. J Biol Chem. 284:20387–20397. 2009. View Article : Google Scholar
|
|
31
|
Han I and Kudlow JE: Reduced O
glycosylation of Sp1 is associated with increased proteasome
susceptibility. Mol Cell Biol. 17:2550–2558. 1997.PubMed/NCBI
|
|
32
|
Lin YH and Yang-Yen HF: The
osteopontin-CD44 survival signal involves activation of the
phosphatidylinositol 3-kinase/Akt signaling pathway. J Biol Chem.
276:46024–46030. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mann M and Jensen ON: Proteomic analysis
of post-translational modifications. Nat Biotechnol. 21:255–261.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kazanecki CC, Uzwiak DJ and Denhardt DT:
Control of osteopontin signaling and function by post-translational
phosphorylation and protein folding. J Cell Biochem. 102:912–924.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jackson SP and Tjian R: O-glycosylation of
eukaryotic transcription factors: implications for mechanisms of
transcriptional regulation. Cell. 55:125–133. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gasbarri A, Del Prete F, Girnita L,
Martegani MP, Natali PG and Bartolazzi A: CD44s adhesive function
spontaneous and PMA-inducible CD44 cleavage are regulated at
post-translational level in cells of melanocytic lineage. Melanoma
Res. 13:325–337. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Solá RJ and Griebenow K: Effects of
glycosylation on the stability of protein pharmaceuticals. J Pharm
Sci. 98:1223–1245. 2009.
|
|
38
|
Dai J, Peng L, Fan K, et al: Osteopontin
induces angiogenesis through activation of PI3K/AKT and ERK1/2 in
endothelial cells. Oncogene. 28:3412–3422. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Iacoangeli A, Lin Y, Morley EJ, et al:
BC200 RNA in invasive and preinvasive breast cancer.
Carcinogenesis. 25:2125–2133. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bader AG and Vogt PK: An essential role
for protein synthesis in oncogenic cellular transformation.
Oncogene. 23:3145–3150. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kaiser C, Dobrikova EY, Bradrick SS,
Shveygert M, Herbert JT and Gromeier M: Activation of
cap-independent translation by variant eukaryotic initiation factor
4G in vivo. RNA. 14:2170–2182. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Beretta L, Gingras AC, Svitkin YV, Hall MN
and Sonenberg N: Rapamycin blocks the phosphorylation of 4E-BP1 and
inhibits cap-dependent initiation of translation. EMBO J.
15:658–664. 1996.PubMed/NCBI
|
|
43
|
Marcotrigiano J, Gingras AC, Sonenberg N
and Burley SK: Cap-dependent translation initiation in eukaryotes
is regulated by a molecular mimic of elF4G. Mol Cell. 3:707–716.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Herbert TP, Kilhams GR, Batty IH and Proud
CG: Distinct signalling pathways mediate insulin and phorbol
ester-stimulated eukaryotic initiation factor 4F assembly and
protein synthesis in HEK 293 cells. J Biol Chem. 275:11249–11256.
2000. View Article : Google Scholar
|
|
45
|
Haghighat A, Mader S, Pause A and
Sonenberg N: Repression of cap-dependent translation by 4E-binding
protein 1: competition with p220 for binding to eukaryotic
initiation factor-4E. EMBO J. 14:5701–5709. 1995.PubMed/NCBI
|
|
46
|
Brunn GJ, Hudson CC, Sekulić A, et al:
Phosphorylation of the translational repressor PHAS-I by the
mammalian target of rapamycin. Science. 277:99–101. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen YJ, Wei YY, Chen HT, et al:
Osteopontin increases migration and MMP-9 up-regulation via
alphavbeta3 integrin, FAK, ERK, and NF-kappaB-dependent pathway in
human chondrosarcoma cells. J Cell Physiol. 221:98–108. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kumar V, Behera R, Lohite K, Karnik S and
Kundu GC: p38 kinase is crucial for osteopontin-induced furin
expression that supports cervical cancer progression. Cancer Res.
70:10381–10391. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chang MS, Lee WS, Chen BC, Sheu JR and Lin
CH: YC-1-induced cyclooxygenase-2 expression is mediated by
cGMP-dependent activations of Ras, phosphoinositide-3-OH-kinase,
Akt, and nuclear factor-kappaB in human pulmonary epithelial cells.
Mol Pharmacol. 66:561–571. 2004.
|
|
50
|
Younes M, Lechago LV, Somoano JR, Mosharaf
M and Lechago J: Wide expression of the human erythrocyte glucose
transporter Glut1 in human cancers. Cancer Res. 56:1164–1167.
1996.PubMed/NCBI
|
|
51
|
Chang SH, Chung YS, Hwang SK, et al:
Lentiviral vector-mediated shRNA against AIMP2-DX2 suppresses lung
cancer cell growth through blocking glucose uptake. Mol Cells.
33:553–562. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sommermann TG, Mack HI and Cahir-McFarland
E: Autophagy prolongs survival after NFκB inhibition in B-cell
lymphomas. Autophagy. 8:265–267. 2012.PubMed/NCBI
|
|
53
|
Park J, Lee HY, Cho MH and Park SB:
Development of a cy3-labeled glucose bioprobe and its application
in bioimaging and screening for anticancer agents. Angew Chem Int
Ed Engl. 46:2018–2022. 2007. View Article : Google Scholar : PubMed/NCBI
|