Introduction
Cell cycle progression in eukaryotic organisms is
primarily controlled by cyclin-dependent kinases (CDKs), cyclins
and CDK inhibitors. Cyclin as a regulatory subunit forms a
heterodimeric complex with corresponding CDKs. The activated
cyclin/CDK complex phosphorylates a set of substrates required for
continued cell cycle progression and triggers transition to
distinct cell cycle phases. During the G1 phase, D-type cyclins
(D1, D2 and D3) accumulate in response to mitogenic stimulation and
form active kinase complexes with their catalytic partners, CDK4
and CDK6 (1). Cyclin D1 plays a
vital role in promoting the G1/S transition of the cell cycle by
inactivating the retinoblastoma protein (Rb) through
superphosphorylation. Phosphorylated Rb liberates E2F and other
Rb-bound transcription regulators, leading to the activation of S
phase-specific genes (1,2). The deregulation of cyclin D1 may
promote the development of tumors. The amplification of the cyclin
D1 gene and the overexpression of cyclin D1 proteins have been
reported in a large proportion of human cancers, including human
breast, lung and gastrointestinal malignancies. For example,
approximately 15–20% of human mammary carcinomas contain the
amplification of the cyclin D1 gene, while cyclin D1 is
overexpressed at the mRNA and protein level in >50% of breast
cancers (3–6). Furthermore, cyclin D1 is required
for tumor initiation and maintenance, as the genetic ablation of
murine cyclin D1 has been shown to result in resistance to
ErbB2-driven mammary adenocarcinomas, and the ubiquitous shutdown
of cyclin D1 in mice bearing ErbB2-driven mammary carcinomas has
been shown to trigger tumor cell senescence (7–9).
The gene encoding cyclin D1 is an oncogene and represents the
second most frequently amplified locus in a diverse set of human
cancer genomes, as revealed by recent findings (10,11). Due to its frequent deregulation
and essential requirement in cancer, cyclin D1 has been the focus
of cancer research and the potential target of drugs designed for
cancer treatment (3,5,8,11).
However, little research has been done on the specific inhibition
of cyclin D1 for cancer therapy.
Antibodies have been shown to have a number of
applications in biotechnology and clinical medicine, particularly
in the field of oncology, due to their high specificity and
affinity binding to target antigens (12,13). With the development of antibody
engineering technology, a number of novel antibodies have been
generated, including single-chain variable fragment (scFv)
antibodies (14,15). scFv antibodies have tremendous
potential in cancer prevention, diagnosis and therapy due to their
specific binding affinity to the antigen, their small size,
superior biodistribution and blood clearance, and can be easily
engineered or modified to create intrabodies (12,13,16–18). Recently, we constructed an
anti-cyclin D1 intrabody (AD5N) by linking a nuclear localization
signal sequence to the human anti-cyclin D1 scFv AD5 coding gene
obtained from the human semi-synthetic scFv phage library. The
expression of AD5N inhibited the growth and proliferation of HeLa
and MCF-7 cells (19,20). Our previous studies indicated that
the scFv antibodies may be a powerful tool which can be used to
block the function of overexpressed cyclin D1. In order to achieve
the optimal knockout of cyclin D1 by the anti-cyclin D1 scFv
antibody, further understanding the characteristics of this scFv is
required.
In this study, a human scFv antibody against cyclin
D1 (AD5) that was derived from a human semi-synthetic scFv phage
library was expressed in the soluble form in Escherichia
coli (E. coli) HB2151 cells. The purification and
identification of AD5 was conducted successfully by using affinity
chromatography, ELISA and western blot analysis. Its binding
affinity to cyclin D1 was measured by non-competitive enzyme
immunoassay. This study lays the foundation for the use of
anti-cyclin D1 scFv antibody for a variety of basic research and
potential gene therapy strategies.
Materials and methods
Materials
The helper phage, VCSM13, the E. coli
strains, HB2151 and XL1-Blue, ovalbumin (OA) and tumor necrosis
factor α (TNFα) were generously provided by Professor Yan Wang
(Navy General Hospital, Beijing, China). Phagemid AD5 encoding
anti-cyclin D1 scFv antibody and V5 tag was screened from a human
semi-synthetic scFv phage library (generously provided by Professor
Yan Wang) (21). The antibody
against cyclin D1 was purchased from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). The antibody against V5 tag was purchased
from Invitrogen (Carlsbad, CA, USA). The antibody against His tag
was purchased from HangZhou HuaAn Biotechnology Company (Hangzhou,
China). Horseradish peroxidase (HRP)-conjugated IgG was purchased
from the Protein Tech Group (Chicago, IL, USA). The HRP-conjugated
anti-M13 antibody, HisTrap HP colomns and PD-10 desalting columns
were purchased from Amersham Pharmacia Biotech, Inc. (Piscataway,
NJ, USA). Bovine serum albumin (BSA) and Coomassie blue R-250 were
purchased from Sigma (St. Louis, MO, USA).
Binding activity of phage antibodies with
cyclin D1
E. coli XL1-Blue competent cells were
transformed by the phagemid AD5 and were then allowed to grow on SB
agar plates containing 50 μg/ml ampicillin and 10 μg/ml
tetracycline overnight at 37°C. Ampicillin- and
tetracycline-resistant colonies carrying phagemid AD5
(AD5/XL1-Blue) were grown in SB medium containing 50 μg/ml
ampicillin and 10 μg/ml tetracycline at 37°C to optical density
(OD)600 0.5. VCSM13 helper phage were added to the
culture of E. coli AD5/XL1-Blue, followed by incubation
without shaking at 37°C for 1 h. The infected cells were then
incubated with shaking overnight at 30°C under 30 μg/ml kanamycin
resistance. The culture supernatants containing phage antibody AD5
were obtained by centrifugation and were then subjected to the
analysis of binding activity by ELISA assay in the microtiter
plates (Nunc) coated with purified cyclin D1 (22) as antigen or OA, TNFα and BSA as
irrelevant antigens controls. VCSM13 helper phage was added to the
microtiter plates (Nunc) coated with purified cyclin D1, OA, TNFα
and BSA instead of the culture supernatants containing phage
antibody AD5 to verify the binding specificity. Following
incubation with the above phage supernatants and HRP-conjugated
anti-M13 antibody, the reaction was developed by adding
o-phenylenediamine (OPD) and monitored by a microplate
reader (Thermo Labsystems, Waltham, MA, USA) at wavelengths of 492
nm.
Expression and identification of soluble
AD5
Amber non-suppressive E. coli HB2151 cells
were infected by the above phage clones to obtain E. coli
AD5/HB2151 that can express the soluble AD5. E. coli
AD5/HB2151 cells were stimulated by isopropyl
β-D-1-thiogalactopyranoside (IPTG) to induce the expression of
soluble AD5. The culture supernatant of AD5/HB2151 was collected by
centrifugation after IPTG induction overnight at 30°C shaking. The
expression and binding activity of soluble AD5 to cyclin D1 were
conducted by ELISA assay in the microtiter plates (Nunc) coated
with purified human recombinant cyclin D1, as previously described
(22) as antigen or OA, TNFα and
p16 as irrelevant antigens controls, followed by sequential
incubation with the above culture supernatant of AD5/HB2151, V5-tag
antibody and HRP-conjugated IgG. The reaction was developed by
adding OPD and monitored by a microplate reader (Thermo Labsystems)
at wavelengths of 492 nm. The microtiter plates (Nunc) coated with
the above proteins were incubated with PBST (0.05% Tween-20 in PBS)
containing 1% of BSA instead of anti-V5-tag antibody as the
negative controls (NC).
Purification of soluble AD5
The culture supernatant of AD5/HB2151 stimulated by
IPTG overnight at 30°C was supplemented with various concentration
of solid ammonium sulphate and stirred at 4°C for 5 h. The solution
was then centrifuged and the pellet was retained. The pellet
containing AD5 was dissolved with ice-cold PBS and dialyzed in PBS
buffer overnight at 4°C. Following centrifugation, the clear
supernatant was collected and applied to affinity chromatography
with His Trap HP column equilibrated with binding buffer (50 mM
phosphate-buffer, pH 7.4, 500 mM NaCl, 10 mM imidazole). The column
was then washed sequentially with binding buffer, washing buffer
(50 mM phosphate-buffer, pH 7.4, 500 mM NaCl, 40 mM imidazole) and
eluting buffer (50 mM phosphate-buffer, pH 7.4, 500 mM NaCl, 500 mM
imidazole). The purification of AD5 was examined by running an
aliquot of the collected samples on 12% SDS-polyacrylamide gel
electrophoresis (PAGE) and then stained with Coomassie blue R-250.
The purified AD5 was desalted by PD-10 desalting columns. The AD5
proteins were kept at −80°C after the determination of the
concentration by Bradford assay.
Immunoblotting
The samples were subjected to SDS-PAGE and the
separated proteins were electrophoretically transferred onto
nitrocellulose membranes (Bio-Rad, Hercules, CA, USA). Non-specific
binding was blocked with PBST (0.05% Tween-20 in PBS) containing 5%
non-fat milk for 1 h at room temperature. The membranes were then
incubated overnight at 4°C with antibodies against His tag or V5
tag in PBST containing 1% non-fat milk at the dilutions specified
by the manufacturers. Following 3 washes with PBST, the membranes
were then incubated with the HRP-conjugated secondary antibodies at
1:5,000 dilution in PBST containing 1% non-fat milk for 1 h at room
temperature. The membranes were then washed 3 times with PBST and
the protein bands were detected using the western blotting
detection system.
Competition ELISA
Cyclin D1 (1–10 μg/ml) was coated onto the surface
of wells in a 96-well microtitre plate (Nunc) overnight at 4°C.
Following blocking, 50 μl of AD5 (10 μg/ml in PBST containing 3%
non-fat milk) and an equal volume of rabbit anti-cyclin D1 antibody
(1:1,000 dilution in PBST containing 3% non-fat milk) were added to
each well as the test group. Wells with only 100 μl of rabbit
anti-cyclin D1 antibody (1:2,000 dilution in PBST containing 3%
non-fat milk) were used as the positive controls. A total of 50 μl
of OA or TNFα (10 μg/ml in PBST containing 3% non-fat milk) and
equal volume of rabbit anti-cyclin D1 antibody (1:1,000 dilution in
PBST containing 3% non-fat milk) were added to each well to examine
the specific binding between AD5 and cyclin D1. After 2 h of
incubation at 37°C, the plate was washed with PBST followed by
detection using HRP-conjugated goat anti-rabbit antibody as
described above. The reaction was developed by the addition of OPD
and monitored by a plate reader (Thermo Labsystems) at wavelengths
of 492 nm. The inhibition rate of AD5 to anti-cyclin D1 polyclonal
antibody was calculated using the following formula: inhibition
rate (%) = (OD492 of the positive control -
OD492 of the test group)/OD492 of the
positive control ×100%.
Determination of affinity
Non-competitive ELISA was used to determine the
affinity of AD5 to cyclin D1 as previously described (23). Briefly, a 96-well plate was coated
with serial dilutions of recombinant human cyclin D1 protein at 4°C
overnight. Four different concentrations of cyclin D1 were used.
Following incubation with serial dilutions of AD5, the plate was
sequentially incubated with anti-V5 tag antibody and HRP-conjugated
goat anti-mouse IgG. The binding reaction was detected by the
addition of OPD and monitored by a microplate reader (Thermo
Labsystems) at wavelengths of 492 nm. Three parallel wells were
used in a plate for each concentration of cyclin D1 and AD5. The
amount of AD5 (Ab′ or Ab) adherent to cyclin D1 (Ag′ or Ag) on the
plate was reflected by the enzyme product measured by
OD492. Sigmoid curves were made from OD vs. the
logarithm of total Ab added to the well. The affinity constant
(Kaff) of the antibody was calculated using the
following formula: Kaff = (n-1)/(n [Ab′] - [Ab])
{n = [Ag]/[Ag′], [Ab] and [Ab′] represent the concentration of
antibody corresponding to half the maximum OD value (OD-100)
calculated with a computerized nonlinear regression method for
plates coated with [Ag] and [Ag′], respectively}.
Statistical analysis
All conditions were performed in triplicate, and the
reported values are representative of 3 independent experiments.
All values are expressed as the means ± SD of 3 parallel
measurements. Data were analyzed using the Student's t-test and
values of P<0.05 were considered to indicate statistically
significant differences.
Results
AD5 antibody binds specifically to cyclin
D1
When the phagemids were rescued by helper phages
derived from M13 (VCSM13), the scFv-gene was fused with gene III of
the M13 phage and expressed as a scFv at the tip of the phage
particle. This is convenient for the detection of the specific
binding of an antibody to a protein. In this study, the phagemid
AD5 was transferred into E. coli XL1-Blue cells and was then
rescued by helper phage VCSM13. The ELISA results demonstrated that
the culture supernatants containing phage antibody AD5 showed
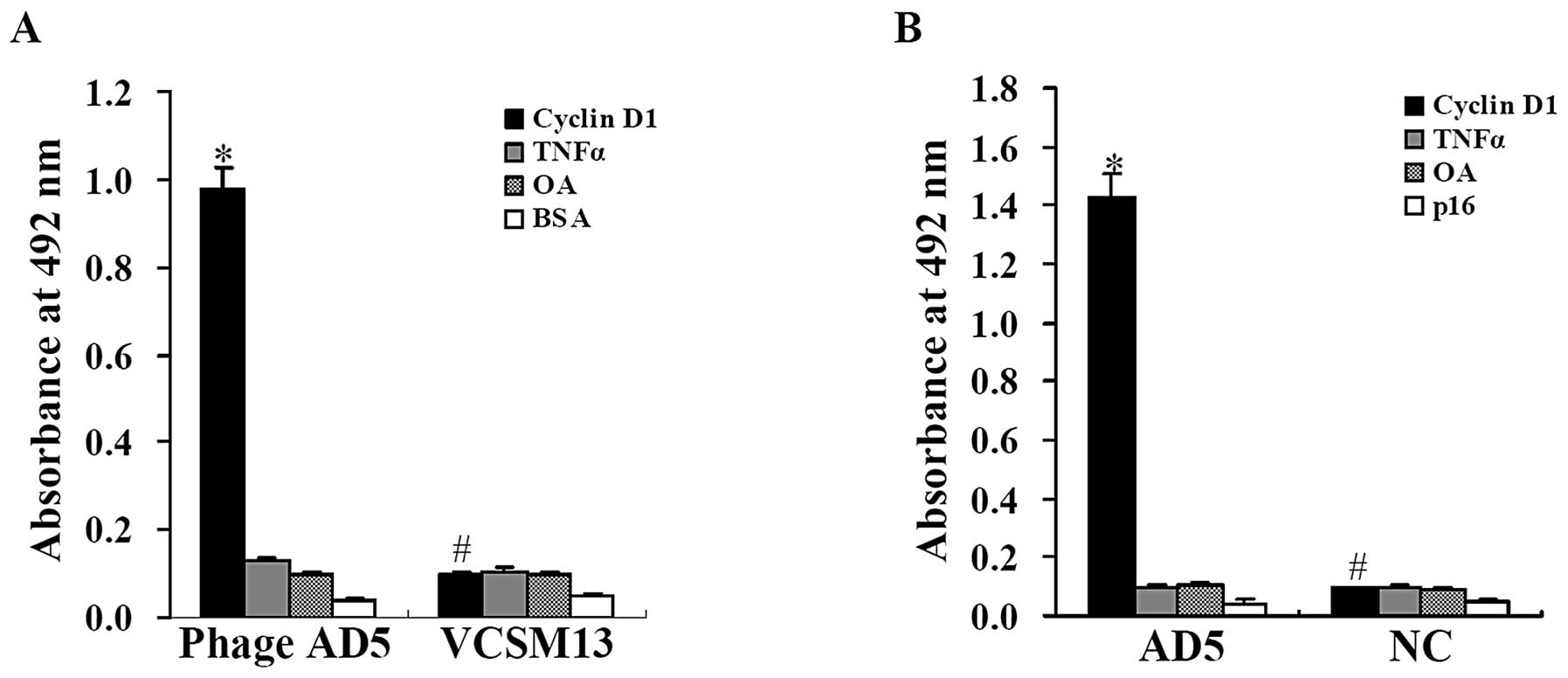
selective binding to cyclin D1. As illustrated in Fig. 1A, the absorbance at 492 nm for AD5
phage binding to cyclin D1 was 0.980±0.044, which was significantly
higher than the irrelevant antigen control groups (P<0.01).
However, compared with the AD5 phage binding activity, the
absorbance at 492 nm for the negative control VCSM13 helper phage
binding to cyclin D1 was distinctly lower (0.100±0.005; P<0.01).
Moreover, the VCSM13 helper phage showed a very similar binding
activity to cyclin D1 and the irrelevant antigen control groups
(P>0.05). This suggests that the phage antibody AD5 has specific
binding affinity to cyclin D1.
In order to identify the binding affinity of soluble
AD5 to cyclin D1, amber non-suppressive E. coli HB2151 cells
were transduced with the AD5 phage clones and stimulated by IPTG to
induce the expression of soluble AD5. The ELISA results revealed a
significantly higher binding affinity for soluble AD5 to cyclin D1.
As shown in Fig. 1B, the
absorbance at 492 nm for soluble AD5 to cyclin D1 was 1.43±0.0785
compared with 0.096±0.006, 0.105±0.007 and 0.045±0.011 to TNFα
(P<0.01), OA (P<0.01) and p16 (P<0.01), respectively. For
the negative controls, although it showed a much lower value than
that for soluble AD5 to cyclin D1 (P<0.01), there was no
siginificant difference among the lower absorbance to the above
proteins incubated with BSA instead of anti-V5-tag antibody
(P>0.05).
Purification of soluble AD5
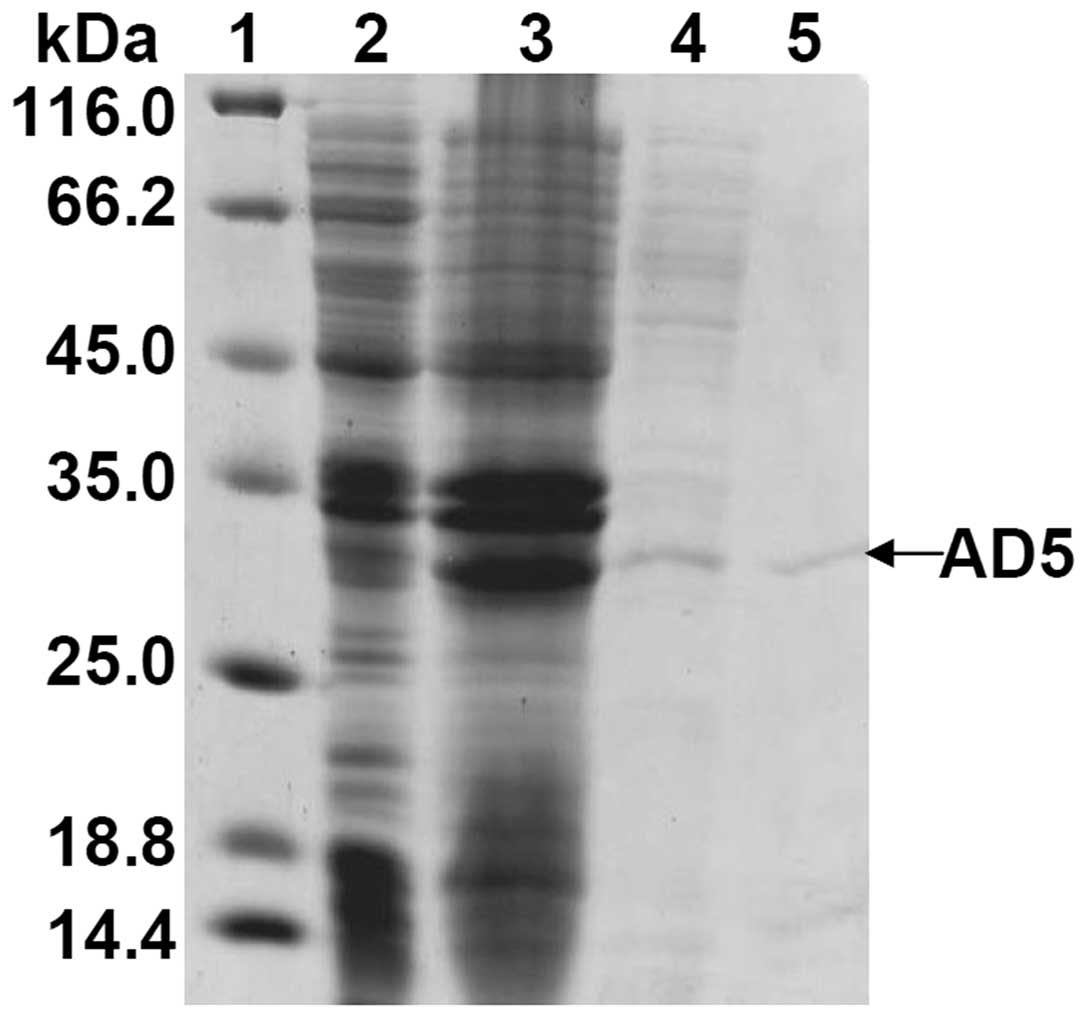
Soluble AD5 was expressed in E. coli
AD5/HB2151 and secreted into the culture medium. To purify soluble
AD5, the separation of crude proteins was first conducted by
ammonium sulfate precipitation from the culture supernatant of
E. coli AD5/HB2151 (Fig.
2). The gradient ammonium sulfate precipitation indicates that
a 40% saturation of ammonium sulfate is opitmal, which can not only
eliminate contaminations of most other proteins but can also yield
more partially purified proteins for the next purification (data
not shown). Therefore, 40% ammonium sulfate precipitation provides
the optimal conditions for further purification. The crude proteins
were then adsorbed to His Trap HP column. SDS-PAGE analysis of the
eluted fractions revealed that a single band of approximately 33
kDa was detected (Fig. 2).
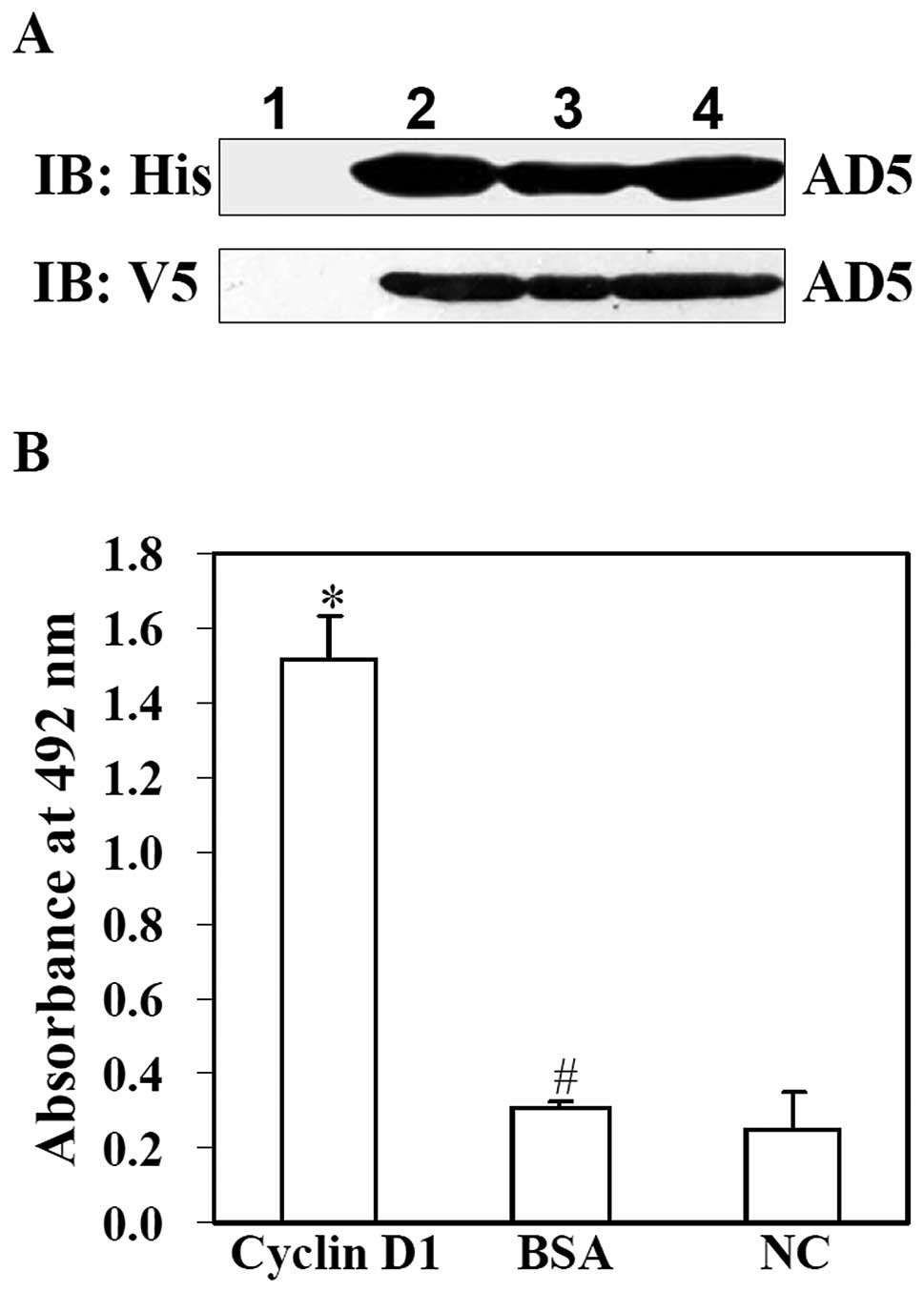
Since AD5 contains a His tag at N-terminal and V5
tag at C-terminal, AD5 as identified by western blot analysis using
anti-V5 or anti-His antibody immunoreactivity. As shown in Fig. 3A, the western blot images
displayed that following induction by IPTG, cell lysis of E.
coli AD5/HB2151, the culture supernatant of E. coli
AD5/HB215 and purified AD5 protein all showed a clear protein band
of approximately 33 kDa. The protein band was not detected in the
cell lysis of E. coli AD5/HB2151 without IPTG induction.
These results suggested that soluble AD5 was expressed in E.
coli AD5/HB2151 cells, secreted into the culture supernatant
and was purified successfully.
In order to examine the binding activity of purified
AD5 to cyclin D1, an ELISA assay was performed. As shown in
Fig. 3B, the absorbance at 492 nm
for the purified AD5 to cyclin D1 was significantly higher than
that of the irrelevant antigen BSA and the negative control (NC)
(P<0.01). However there was no siginificant difference among the
lower absorbance of BSA and the negative control (P>0.05).
AD5 competes with anti-cyclin D1
polyclonal antibody to bind to cyclin D1
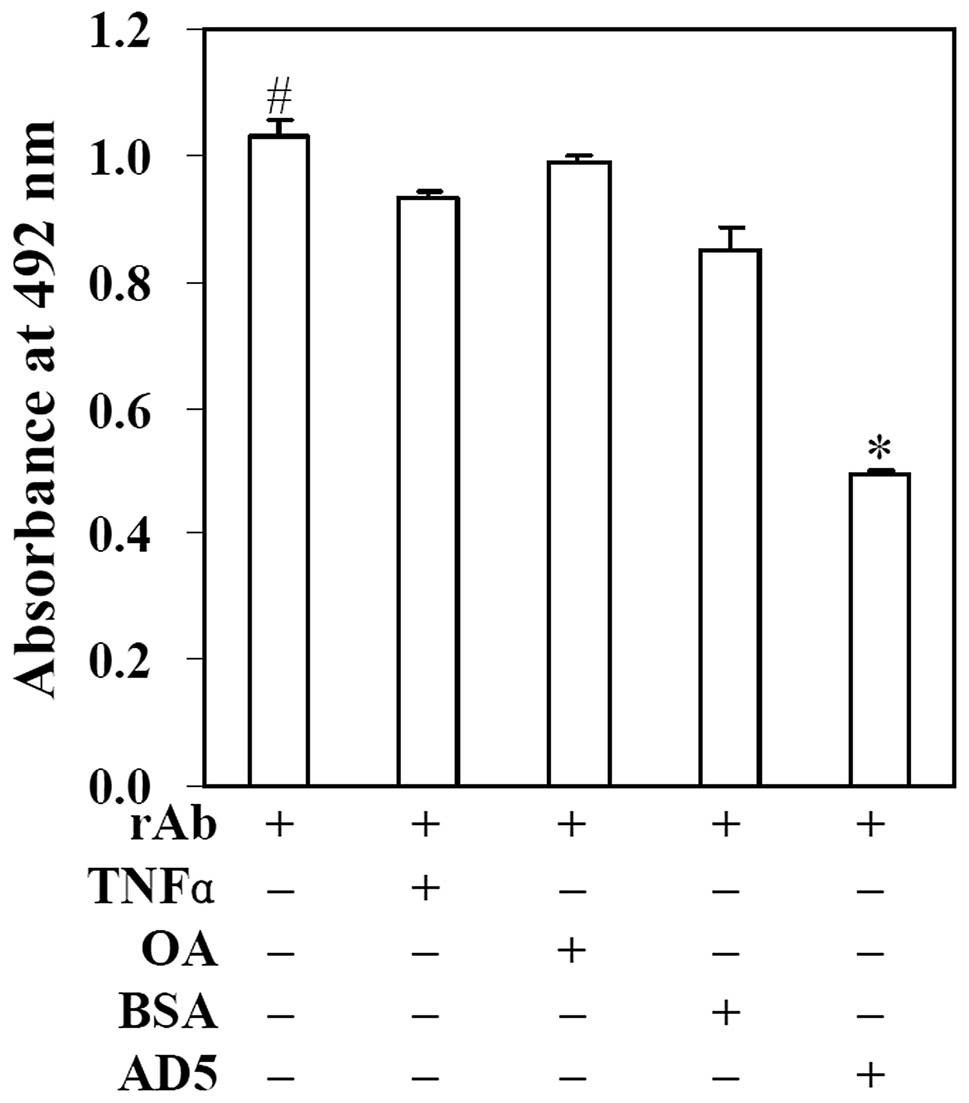
To detect the competitive inhibition ability of AD5
with the anti-cyclin D1 polyclonal antibody to bind to cyclin D1,
competition ELISA experiments were conducted. As illustrated in
Fig. 4, when cyclin D1 was
incubated only with anti-cyclin D1 polyclonal antibody, the
OD492 was 1.029±0.027. Following incubation with
anti-cyclin D1 polyclonal antibody mixture containing AD5, the
OD492 was markedly reduced to 0.495±0.006. However, when
cyclin D1 was incubated with anti-cyclin D1 polyclonal antibody
mixture containing the irrelevant antigens, TNFα, OA or BSA, the
OD492 was 0.930±0.013, 0.990±0.009 and 0.848±0.039,
respectively, which showed no significant difference compared with
the positive control group (P>0.05). These results indicated
that AD5 significantly inhibited the binding between anti-cyclin D1
polyclonal antibody and cyclin D1 (P<0.01). According to the
above formula, we found that the inhibition rate was approximately
52%. This suggests that AD5 competes with the anti-cyclin D1
polyclonal antibody to bind to cyclin D1.
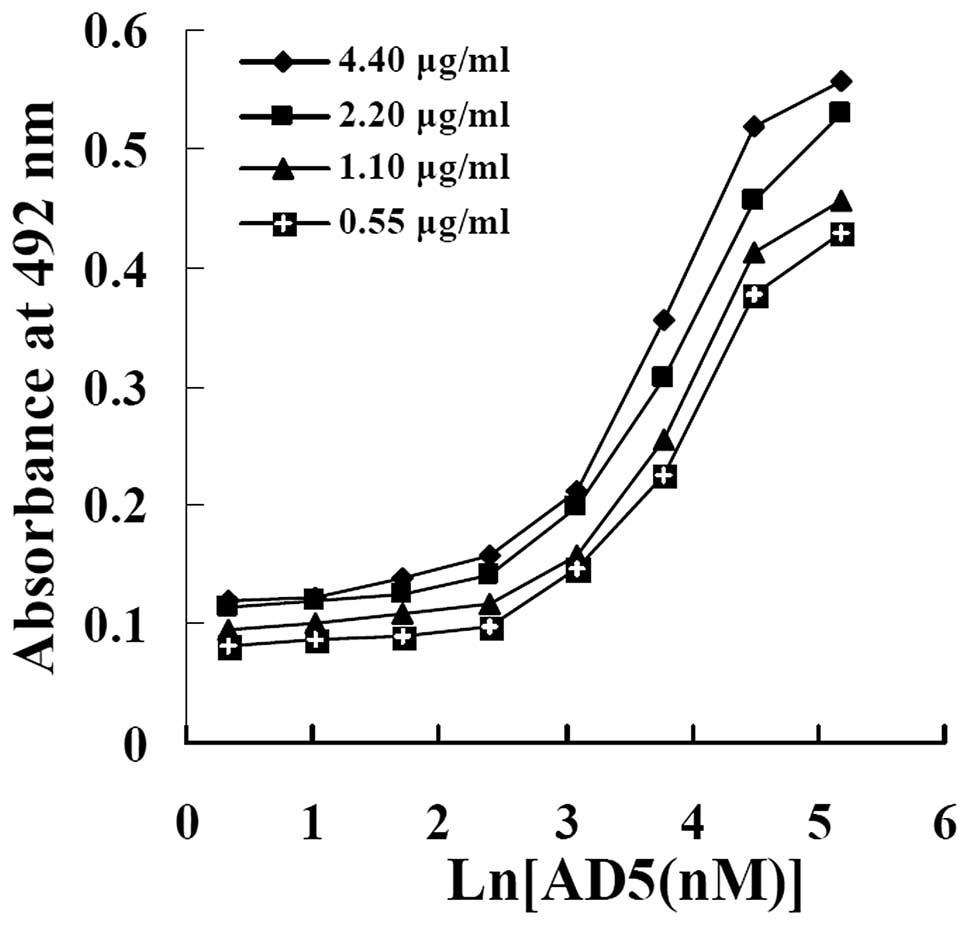
Affinity of AD5
The relative affinity constant of AD5 was measured
by non-competitive enzyme immunoassay, as previously described by
Beaty et al, which is able to measure Kaff
by solid-phase EIA using serial dilutions of antigens (coating the
plate) and antibody (23).
According to the 4 concentrations of cyclin D1 (4.40, 2.20, 1.10
and 0.55 μg/ml) coated onto the plates, 4 sigmoid curves were made
from OD vs. the logarithms of AD5 concentrations added to the wells
(Fig. 5). As shown in Table I, according to the curves and the
formula, 6 affinity constants (3 for n=2, 2 for n=4 and 1 for n=8)
were obtained for AD5. The mean affinity constant
(Kaff) of AD5 for cyclin D1 was approximately
(1.19±0.056) × 107 M−1.
 | Table IAffinity calculated from the
non-competitive ELISA curves of AD5 antibody against cyclin D1. |
Table I
Affinity calculated from the
non-competitive ELISA curves of AD5 antibody against cyclin D1.
| Affinity
(×107 M−1) |
|---|
|
|
|---|
| Antibody |
Kn=2 |
Kn=4 |
Kn=8 | Mean ± SD |
|---|
| AD5 | 1.10, 1.27,
1.16 | 1.21, 1.19 | 1.18 | 1.19 ± 0.06 |
Discussion
Cyclin D1 is a component of the core cell cycle
machinery. Cyclin D1 activates CDK4 and CDK6 and drives cell growth
and proliferation. Its deregulation may promote the development of
tumors (3,5,8,9).
Cyclin D1 may be a potential prognostic marker and a therapeutic
target in cancer (5,9,10,24). Although previous studies have
indicated that the function of cyclin D1 is inhibited by using
inhibitors to CDK4 (8,9), this is not sufficient to block the
activity of cyclin D1, as cyclin D1 has been reported to modulate
the activity of several transcription factors independent of
binding to CDK4 (25,26). Therefore, there is a pressing need
to develop inhibitory strategies targeting cyclin D1. Antibodies
have been proven to be attractive anticancer agents due to their
high specificity and affinity towards targeting antigens (12,13). In this study, a human scFv
antibody against cyclin D1 (AD5) that was derived from a human
semi-synthetic scFv phage library was expressed in the soluble form
in E. coli HB2151 cells. Soluble AD5 was isolated from
culture supernatants by the saturation of ammonium sulfate
precipitation at a concentration of 40%, and purified successfully
through the His Trap HP column. AD5 could specifically bind to
human recombinant cyclin D1 with approximately (1.19±0.056) ×
107 M−1 affinity constant and showed
approximately 52% competitive inhibition with the anti-cyclin D1
polyclonal antibody for binding to cyclin D1. These results
demonstrated that a human anti-cyclin D1 scFv antibody specifically
binding to cyclin D1 was successfully prepared.
scFv antibodies have been successfully isolated and
displayed in various expression systems, including E. coli
cells (27,28). Previous studies have demonstrated
that the majority of scFv antibodies are expressed as insoluble
inclusion bodies in the cytoplasm of E. coli cells (27–30). Although the inclusion bodies can
easily be separated from the soluble proteins of host cells and
increase the yield of the purified product, the inclusion bodies
require denaturation and refolding, which increases the difficulty
to produce active soluble scFv antibodies (27). To overcome this problem, a signal
peptide is used to direct the secretion of the scFv antibodies to
the periplasmic space and culture medium in soluble form (31). In this study, soluble scFv
antibody was expressed in non-suppressive E. coli HB2151
bacteria cells. There is a succinic acid termination codon gene
between the phage pIII gene and scFv gene. When the phage is
infected into non-suppressive E. coli HB2151, the succinic
acid termination codon will be translated to the termination codon.
Therefore, the scFv gene will not be expressed as a fusion protein
with pIII in HB2151, but it will be expressed as the soluble
protein and secreted into the periplasmic space and culture medium.
In this study, after induction with IPTG, we found there was
soluble anti-cyclin D1 scFv antibody (AD5) in the supernatant of
culture medium by ELISA assay and western blot analysis. The active
purified soluble AD5 was obtained successfully through affinity
chromatography. Therefore, the expression system for soluble AD5 we
applied in this study is certainly feasible.
Affinity of the antibody is a critical parameter
that shows the ability of the antibody to bind to its antigen. scFv
antibodies from either the semi-synthetic or naive antibody library
generaly bind to their antigens with lower affinities, ranging from
106–109 M−1 (32). In this study, the scFv antibody
AD5 showed an approximately 107 M−1 affinity
constant. Although scFv AD5 has an affinity to the antigen within
the regular range, AD5 exhibits a slightly lower binding activity.
The reason for this lower affinity of scFv antibodies may be
related to the monovalent nature of the scFv molecule (33), and the non-immunized
semi-synthetic antibody library we used (34). Although antibodies with moderate
affinity (10−7–10−9 M) may contribute to
effective penetration into tumors and moderate retention on tumor
cells (32), the moderate
antibody affinity or specificity may be insufficient for diagnostic
or certain therapeutic applications (27). In order to obtain higher affinity
for scFv, optimization of antibodies by either site-directed
mutagenesis or chain shuffling is necessary (32). Using site-directed mutagenesis of
the variable light chain and variable heavy chain CDR3 regions,
Adams et al (32)
generated a series of affinity mutants from the original C6.5 scFv.
These scFv antibodies bind to the same epitope of HER-2/neu with
affinities ranging from 10−6 to 1.5×10−11 M
(32,35). According to the intended
application, the optimization of AD5 with appropriate affinity for
antibody-based therapeutics will be carried out further using
approaches similar to those outlined above.
In conclusion, cyclin D1 is a potential target for
cancer prevention and treatment (9,10).
In this study, we successfully expressed and purified a novel scFv
antibody AD5 with specific binding affinity to cyclin D1. This
study undoubtedly provides a novel potential tool for targeting
cyclin D1 in the therapy, diagnosis and prognosis of cancer.
Acknowledgements
This study was partially supported by grants from
the National Natural Science Foundation of China (Nos. 31170882,
30972806 and 30200256), the Program for New Century Excellent
Talents in University (NCET-09-0420) and the S&T Development
Planning Program of Jilin Province (Nos. 20111806 and 20090727) (to
G.L.). We thank Professor Yan Wang of the Navy General Hospital,
Beijing, China, for his assistance with the scFv antibody
screening.
References
|
1
|
Sherr CJ: Mammalian G1 cyclins. Cell.
73:1059–1065. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: a changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Deshpande A, Sicinski P and Hinds PW:
Cyclins and CDKs in development and cancer: a perspective.
Oncogene. 24:2909–2915. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fu M, Wang C, Li Z, Sakamaki T and Pestell
RG: Minireview: cyclin D1: normal and abnormal functions.
Endocrinology. 145:5439–5447. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Musgrove EA, Caldon CE, Barraclough J,
Stone A and Sutherland RL: Cyclin D as a therapeutic target in
cancer. Nat Rev Cancer. 11:558–572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roy PG, Pratt N, Purdie CA, Baker L,
Ashfield A, Quinlan P and Thompson AM: High CCND1 amplification
identifies a group of poor prognosis women with estrogen receptor
positive breast cancer. Int J Cancer. 127:355–360. 2010.PubMed/NCBI
|
|
7
|
Yu Q, Geng Y and Sicinski P: Specific
protection against breast cancers by cyclin D1 ablation. Nature.
411:1017–1021. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Choi YJ, Li X, Hydbring P, Sanda T,
Stefano J, Christie AL, Signoretti S, Look AT, Kung AL, von Boehmer
H and Sicinski P: The requirement for cyclin D function in tumor
maintenance. Cancer Cell. 22:438–451. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Malumbres M: Cell cycle-based therapies
move forward. Cancer Cell. 22:419–420. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Beroukhim R, Mermel CH, Porter D, et al:
The landscape of somatic copy-number alteration across human
cancers. Nature. 463:899–905. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim JK and Diehl JA: Nuclear cyclin D1: an
oncogenic driver in human cancer. J Cell Physiol. 220:292–306.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Scott AM, Wolchok JD and Old LJ: Antibody
therapy of cancer. Nat Rev Cancer. 12:278–287. 2012. View Article : Google Scholar
|
|
13
|
Elvin JG, Couston RG and van der Walle CF:
Therapeutic antibodies: market considerations, disease targets and
bioprocessing. Int J Pharm. 440:83–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stewart CS, MacKenzie CR and Hall JC:
Isolation, characterization and pentamerization of alpha-cobrotoxin
specific single-domain antibodies from a naïve phage display
library: preliminary findings for antivenom development. Toxicon.
49:699–709. 2007.PubMed/NCBI
|
|
15
|
Wen S, Zhang X, Liu Y, Zhang QQ, Liu XJ
and Liang JS: Selection of a single chain variable fragment
antibody against ivermectin from a phage displayed library. J Agric
Food Chem. 58:5387–5391. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Holliger P and Hudson PJ: Engineered
antibody fragments and the rise of single domains. Nat Biotechnol.
23:1126–1136. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dai H, Gao H, Zhao X, Dai L, Zhang X, Xiao
N, Zhao R and Hemmingsen SM: Construction and characterization of a
novel recombinant single-chain variable fragment antibody against
white spot syndrome virus from shrimp. J Immunol Methods.
279:267–275. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Weisser NE and Hall JC: Applications of
single-chain variable fragment antibodies in therapeutics and
diagnostics. Biotechnol Adv. 27:502–520. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou LH, Zhu X, Cao YH, Wang L, Chen Y, Du
BR and Li GY: Construction of expression vector for anti-cyclin D1
intrabody AD5N and its inhibitory effects on cell proliferation of
breast cancer. Chin J Immunol. 24:703–706. 2008.
|
|
20
|
Zhou LH, Zhu X, Cao YH, Chen Y, Tian Y,
Wang L and Li GY: Effects of anti-cyclin D1 intrabody AD5N on HeLa
cells of uterine cervix cancer. Chin J Clin Oncol. 35:942–944.
2008.
|
|
21
|
Qiao YY, Wang Y, Chen XS, Wang YX and Hua
B: Construction of a large single-chain phage antibody library.
Chin J Microbiol Immunol. 24:194–197. 2004.
|
|
22
|
Li GY, Zou DS and Zhou LH: Expression and
purification of recombinant human cyclin D1 in E. coli BL21.
J Jilin Univ. 44:839–843. 2006.
|
|
23
|
Beatty JD, Beatty BG and Vlahos WG:
Measurement of monoclonal antibody affinity by non-competitive
enzyme immunoassay. J Immunol Methods. 100:173–179. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ding Z, Wu CJ, Chu GC, et al:
SMAD4-dependent barrier constrains prostate cancer growth and
metastatic progression. Nature. 470:269–273. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bienvenu F, Jirawatnotai S, Elias JE, et
al: Transcriptional role of cyclin D1 in development revealed by a
genetic-proteomic screen. Nature. 463:374–378. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bartek J and Lukas J: DNA repair: Cyclin
D1 multitasks. Nature. 474:171–172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ahmad ZA, Yeap SK, Ali AM, Ho WY, Alitheen
NB and Hamid M: scFv antibody: principles and clinical application.
Clin Dev Immunol. 2012:9802502012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu X, Wang D, Li S, Xiao Q, Tao K, Hu J,
Cao W and Feng W: Expression of a humanized single-chain variable
fragment antibody targeting chronic myeloid leukemia cells in
Escherichia coli and its characterization. Int J Mol Med.
29:939–945. 2012.PubMed/NCBI
|
|
29
|
Sushma K, Vijayalakshmi MA, Krishnan V and
Satheeshkumar PK: Cloning, expression, purification and
characterization of a single chain variable fragment specific to
tumor necrosis factor alpha in Escherichia coli. J
Biotechnol. 156:238–244. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang T, Yang L, Chai W, Li R, Xie J and
Niu B: A strategy for high-level expression of a single-chain
variable fragment against TNFα by subcloning antibody variable
regions from the phage display vector pCANTAB 5E into pBV220.
Protein Expr Purif. 76:109–114. 2011.PubMed/NCBI
|
|
31
|
Abdolalizadeh J, Nouri M, Zolbanin JM,
Baradaran B, Barzegari A and Omidi Y: Downstream characterization
of anti-TNF-α single chain variable fragment antibodies. Hum
Antibodies. 21:41–48. 2012.
|
|
32
|
Adams GP and Schier R: Generating improved
single-chain Fv molecules for tumor targeting. J Immunol Methods.
231:249–260. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yuan W and Parrish CR: Comparison of two
single-chain antibodies that neutralize canine parovirus: analysis
of an antibody-combining site and mechanisms of neutralization.
Virology. 269:471–480. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Neri P, Shigemori N, Hamada-Tsutsumi S,
Tsukamoto K, Arimitsu H, Shimizu T, Akahori Y, Kurosawa Y and Tsuji
T: Single chain variable fragment antibodies against Shiga toxins
isolated from a human antibody phage display library. Vaccine.
29:5340–5346. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schier R, McCall A, Adams GP, Marshall KW,
Merritt H, Yim M, Crawford RS, Weiner LM, Marks C and Marks JD:
Isolation of picomolar affinity anti-c-erbB-2 single-chain Fv by
molecular evolution of the complementarity determining regions in
the center of the antibody binding site. J Mol Biol. 263:551–567.
1996. View Article : Google Scholar
|